Embed Size (px)
Citation preview

R E S E A R C H A R T I C L E
E¡ectofacetic acidand pHon the cofermentationofglucoseandxylose to ethanol byageneticallyengineered strain ofSaccharomyces cerevisiaeElizabeth Casey1,2, Miroslav Sedlak1,2, Nancy W.Y. Ho1,3 & Nathan S. Mosier1,2
1Laboratory of Renewable Resources Engineering, Purdue University, West Lafayette, IN, USA; 2Department of Agricultural and Biological Engineering,
Purdue University, West Lafayette, IN, USA; and 3Department of Chemical Engineering, Purdue University, West Lafayette, IN, USA
Correspondence: Nathan S. Mosier,
Laboratory of Renewable Resources
Engineering, Purdue University, West
Lafayette, IN, USA. Tel.: 11 765 494 7022;
fax: 11 765 494 7023;
e-mail: [email protected]
Received 8 September 2009; revised 13 January
2010; accepted 22 February 2010.
Final version published online 14 April 2010.
DOI:10.1111/j.1567-1364.2010.00623.x
Editor: Jens Nielsen
Keywords
Saccharomyces cerevisiae; xylose; acetic acid;
ethanol; cellulose; inhibition.
Abstract
A current challenge of the cellulosic ethanol industry is the effect of inhibitors
present in biomass hydrolysates. Acetic acid is an example of one such inhibitor
that is released during the pretreatment of hemicellulose. This study examined the
effect of acetic acid on the cofermentation of glucose and xylose under controlled
pH conditions by Saccharomyces cerevisiae 424A(LNH-ST), a genetically engi-
neered industrial yeast strain. Acetic acid concentrations of 7.5 and 15 g L�1,
representing the range of concentrations expected in actual biomass hydrolysates,
were tested under controlled pH conditions of 5, 5.5, and 6. The presence of acetic
acid in the fermentation media led to a significant decrease in the observed
maximum cell biomass concentration. Glucose- and xylose-specific consumption
rates decreased as the acetic acid concentration increased, with the inhibitory effect
being more severe for xylose consumption. The ethanol production rates also
decreased when acetic acid was present, but ethanol metabolic yields increased
under the same conditions. The results also revealed that the inhibitory effect of
acetic acid could be reduced by increasing media pH, thus confirming that the
undissociated form of acetic acid is the inhibitory form of the molecule.
Introduction
Historically, petroleum has been the major source for liquid
transportation fuels. However, declining oil reserves and
environmental concerns have led to interest in alternative,
renewable energy sources. One promising alternative is the
conversion of plant biomass into ethanol. The primary
biomass feedstocks for the current ethanol industry have
been corn grain and sugar cane. However, interest has
recently shifted to replacing these traditional feedstocks with
more abundant, non-food-based cellulosic biomass feed-
stocks such as agricultural wastes (e.g. corn stover) or energy
crops (e.g. switchgrass). The use of cellulosic biomass as
feedstock for the production of ethanol via biochemical
routes presents many technical hurdles not faced with the
use of corn or sugar cane as feedstock. One significant
challenge is the development of efficient and economical
pretreatment and enzymatic hydrolysis steps for the release
of fermentable sugars from the biomass (Mosier et al., 2005;
Stephanopoulos, 2007). Another obstacle is engineering
robust, process-relevant industrial microorganisms that are
capable of mixed sugar fermentation and are tolerant to
inhibitors (Hahn-Hagerdal et al., 2007).
The complexity of cellulosic biomass hydrolysates as
compared with corn starch or sugar cane hydrolysates leads
to the challenge of engineering a suitable microorganism.
Cellulosic biomass hydrolysates contain a variety of sugars
(primarily glucose and xylose) and inhibitory compounds
such as acetic acid and various phenolic compounds. The
primary microorganism used in industrial fermentations,
Saccharomyces cerevisiae, is not able to ferment xylose
(Barnett, 1976), which represents about 40% of the sugars
found in biomass hydrolysates. This problem of limited
sugar utilization has been successfully addressed through
metabolic engineering of S. cerevisiae with genes for the
xylose metabolic pathways of xylose-fermenting bacteria, i.e.
xylose isomerase (Kuyper et al., 2005), or yeasts, i.e. xylose
reductase and xylitol dehydrogenase (Ho & Chen, 1997;
FEMS Yeast Res 10 (2010) 385–393 c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
YEA
ST R
ESEA
RC
H

Ho et al., 1998; Eliasson et al., 2000; Sonderegger et al.,
2004). Nontraditional microorganisms for industrial etha-
nol production (such as Escherichia coli, Zymomonas mobi-
lis, and Thermoanaerobacterium saccharolyticum) have also
been engineered to convert mixed sugar streams to ethanol
(Lindsay et al., 1995; Zhang et al., 1995; Shaw et al., 2008).
With strains now capable of mixed sugar fermentation, the
primary challenge has become the development of industrial
microorganisms with tolerance to the inhibitors that are
formed and released as biomass is broken down in the
pretreatment and hydrolysis steps (Hahn-Hagerdal et al.,
2007). The inhibitors found in biomass hydrolysates can be
primarily classified as weak acids, furan derivatives, or
phenolic compounds. These inhibitors have been shown to
negatively impact the fermentative performance (cell
growth, ethanol yield and productivity, and/or sugar con-
sumption rates) of non-pentose-fermenting strains of
S. cerevisiae (Palmqvist & Hahn-Hagerdal, 2000; Klinke
et al., 2004; Almeida et al., 2007) and Z. mobilis (Lawford &
Rousseau, 1993), as well as non-ethanol-producing strains
of E. coli (Luli & Strohl, 1990). The previously studied
strains are not suitable for industrial ethanol production
because of their substrate and product limitations. It would
be more valuable to understand how inhibitors impact
microorganisms that have been engineered to ferment
multiple substrates to ethanol. For example, Bellissimi et al.
(2009) recently published a study discussing the effect of
acetic acid on a strain of S. cerevisiae that was engineered for
xylose fermentation with bacterial xylose isomerase. How-
ever, few inhibition studies of engineered organisms have
been published.
Acetic acid is a weak acid generated from the deacetyla-
tion of hemicellulose during pretreatment (Palmqvist &
Hahn-Hagerdal, 2000; Klinke et al., 2004; Almeida et al.,
2007). It is known to inhibit microbial growth and has been
used as an antimicrobial agent in the food and beverage
industries (Luck & Jager, 1997). Acetic acid is present in
varying concentrations in all types of biomass, for example
corn stover and poplar contain 5.6% and 3.6% acetyl by
mass, respectively (Lu et al., 2009). When studying the
inhibitory effect that acetic acid can have on microorgan-
isms, process-relevant conditions for the production of
cellulosic ethanol at an industrial scale must be considered.
Assuming that the minimum concentration of ethanol for
economic distillation is 5% ethanol, the initial unhydrolyzed
biomass concentration at the start of the process must be
approximately 20% by weight. This would result in theore-
tical acetic acid concentrations of 11.2 and 7.2 g L�1 in the
hydrolysates of corn stover and poplar, respectively, assum-
ing no accumulation due to recycling of process streams. An
actual acetic acid concentration of 13 g L�1 has been ob-
served in dilute acid-pretreated corn stover hydrolysate (Lu
et al., 2009). The removal of acetic acid and other inhibitors
would add cost to the overall process. Therefore, a detailed
study of the effect of these inhibitors on ethanol yields and
production rates, especially for xylose fermentation, is
important for ongoing microorganism development efforts
and cellulosic ethanol commercialization.
The goal of the study reported in this paper was to
determine the effect of acetic acid at relevant industrial
process concentrations on the cofermentation of glucose
and xylose under controlled pH conditions by S. cerevisiae
424A(LNH-ST), a polyploid industrial yeast strain capable
of fermenting glucose and xylose. Saccharomyces cerevisiae
424A(LNH-ST) was genetically engineered for xylose meta-
bolism by overexpressing xylose reductase, xylitol dehydro-
genase from Pichia stipitis, and xylulose kinase from
S. cerevisiae (Ho et al., 1998; Ho et al., 2000). We report the
effects of acetic acid and pH on biomass growth, substrate
consumption rates, and ethanol production rates.
Materials and methods
Yeast strain
All fermentations utilized S. cerevisiae 424A(LNH-ST), a
recombinant industrial yeast strain capable of the cofermen-
tation of glucose and xylose (Ho et al., 1998; Ho et al., 2000).
Fermentation experiments
Batch fermentations were completed in 1-L New Brunswick
BioFlo 110 benchtop fermentors (Edison, NJ) equipped
with pH control. The culture volume was 80% of the
fermentor volume. No sparging of any gas was performed.
Our measurements of dissolved oxygen during fermentation
showed that the oxygen concentration declined to below 1%
of saturation within 40 min of the beginning of the fermen-
tation. The inoculum for the fermentor was prepared by
pregrowing yeast aerobically in a shaker set at 28 1C and
200 r.p.m. in 2-L flasks containing 500 mL YEPD media (1%
yeast extract, 2% peptone, and 2% glucose) (Mallinckrodt
Chemicals, Phillipsburg, NJ).
YEP media (1% yeast extract, 2% peptone) were used as
the fermentation media. YEP was chosen as the fermenta-
tion medium because the extensive prior data for the
performance of this yeast were obtained using this medium.
This allowed for comparisons between this work and prior
work. In addition, YEP is a rich medium. Thus, the most
significant stress on the fermenting yeast was the presence of
acetic acid unencumbered by the metabolic stress due to the
synthesis of minor metabolites. Glucose and xylose concen-
trations in the starting fermentation media were 60 g L�1
each. The acetic acid concentrations examined were 0 (for
control), 7.5, and 15 g L�1. These concentrations were
selected to represent the range of concentrations that can
be expected in hydrolysates from a variety of biomass
FEMS Yeast Res 10 (2010) 385–393c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
386 E. Casey et al.

sources (Takahashi et al., 1999). The pH of the media was
adjusted to the desired value (5, 5.5, or 6) with ammonium
hydroxide (28–30% NH3) (Mallinckrodt Chemicals) or
14.8 M phosphoric acid (Fisher Scientific, Pittsburgh, PA).
Once the cell density of the growth culture reached
approximately 6 g dry cells L�1, the culture was centrifuged
for 5 min at 3100 g. The cell pellet was resuspended in YEP
and used to inoculate the fermentor to an initial cell density
of 4.75 g dry cells L�1. The temperature and r.p.m. for the
fermentation were set to 28 1C and 200 r.p.m., respectively.
The media pH was continuously controlled within � 0.1
from the desired value using the pH control system provided
with the BioFlo 110 fermentor using 1 M phosphoric acid
and 1 M ammonium hydroxide. BIOCOMMAND PLUS software
(New Brunswick Scientific Co.) was used to record the real-
time media pH. The fermentation then proceeded for a
maximum of 240 h and all fermentations were carried out in
duplicate.
Analysis of fermentation substrates andproducts
The fermentation metabolites were analyzed by HPLC using
the method outlined by Lu et al. (2009) using a Waters
Alliance 2695 HPLC system with an Aminexs HPX-87H
300� 7.8 mm column (Bio-Rad Laboratories, Hercules,
CA). The HPLC column operating conditions were 60 1C at
a flow rate of 0.6 mL min�1 for the mobile phase, 5 mM
sulfuric acid in water.
Results
We were interested in determining the combined effects of
acetic acid and pH on various fermentation performance
characteristics of S. cerevisiae 424A(LNH-ST). Specifically,
we examined the impact of these factors on biomass growth,
glucose and xylose consumption rates, and ethanol produc-
tivity rates and yields. To accomplish this, a 32 factorial
experimental fermentation design was selected. Two factors,
acetic acid concentration and media pH, at three different
levels were chosen to provide a total of nine different
fermentation conditions, with each fermentation condition
repeated twice. Figure 1 shows the representative fermenta-
tion profiles for each of the nine conditions. A visual
comparison of the profiles revealed the effect of acetic acid
and pH on the cofermentation of glucose and xylose. The
maximum biomass concentrations decreased and the xylose
consumption rates slowed as the acetic acid concentration
increased and pH decreased. The increase in the acetic acid
concentration and the decrease in pH also corresponded to a
decrease in ethanol production rates. Acetic acid had a
minimal inhibitory effect on glucose consumption, with
the exception of the most severe condition tested (pH 5 and
15 g L�1 acetic acid). More detailed results for each of these
fermentation performance characteristics are provided in
the following sections.
Impact of acetic acid and pH on biomass growth
Because of the high cell concentration at inoculation,
minimal cell growth was observed. However, the extent of
this growth for each of the nine fermentation conditions was
compared to determine the effect of acetic acid and pH on
the growth of S. cerevisiae 424A(LNH-ST). The maximum
biomass concentrations under each condition are summar-
ized in Table 1. At a given pH, the maximum biomass
concentration was shown to decrease significantly in fer-
mentations with acetic acid as compared with the control
(with the exception of the least severe condition tested, pH 6
and 7.5 g L�1 acetic acid).
Slight reductions in the biomass concentration were also
observed as the pH decreased for fermentations at a given
acetic acid concentration. This effect was not observed in the
control fermentations, suggesting that the reductions in
biomass when acetic acid was present cannot be explained
solely by the decrease in pH. This hypothesis was confirmed
by a previous study that reported that pH had no effect on
biomass growth in the range examined here (Phowchinda
et al., 1995).
The biomass yields observed (0.016–0.025 g g�1 sugar
consumed) were less than the expected yield of 0.055 g g�1
sugar consumed for anaerobic metabolism (Davis et al.,
2006). A carbon balance of all fermentations following the
convention of Wang et al. (1979) closed at or above 90%
(data not shown). Therefore, the lower yield of biomass was
offset by the higher yield of fermentation products such as
ethanol, CO2, xylitol, and glycerol.
Impact of acetic acid and pH on specific glucoseand xylose consumption rates
To explore the effect of acetic acid and pH on substrate
consumption, the initial specific sugar consumption rates
(both glucose and xylose) were calculated for each fermen-
tation condition. This rate was calculated by determining
the slope of the steepest portion of the substrate concentra-
tion curve and dividing that by the average cell concentra-
tion during that period to yield a rate with the units of
g substrate g�1 dry cell h�1. The initial specific glucose con-
sumption rates are summarized in Table 1. The glucose
consumption rates for a single pH condition decreased
significantly with increasing acetic acid concentration, with
the exception of pH 6. Under the most severe condition (pH
5 and 15 g L�1 acetic acid), the glucose consumption rate was
only 12% that of the control. However, the inhibitory effect
of acetic acid on the glucose consumption rates decreased as
pH increased. For example, the glucose consumption rate of
the fermentation with 7.5 g L�1 acetic acid at pH 5 was about
FEMS Yeast Res 10 (2010) 385–393 c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
387Effect of acetic acid/pH on xylose fermentation

Fig. 1. Time course profiles for the cofermentation of glucose and xylose by Saccharomyces cerevisiae 424A(LNH-ST) in the presence of varying
concentrations of acetic acid and pH values: (a) 0 g L�1 acetic acid, pH 6; (b) 0 g L�1 acetic acid, pH 5.5; (c) 0 g L�1 acetic acid, pH 5; (d) 7.5 g L�1 acetic
acid, pH 6; (e) 7.5 g L�1 acetic acid, pH 5.5; (f) 7.5 g L�1 acetic acid, pH 5; (g) 15 g L�1 acetic acid, pH 6; (h) 15 g L�1 acetic acid, pH 5.5; and (i) 15 g L�1
acetic acid, pH 5. �, glucose; �, xylose; ., acetic acid; n, ethanol; ~, biomass.
FEMS Yeast Res 10 (2010) 385–393c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
388 E. Casey et al.
half that of the control, compared with almost 90% of the
control when the pH was increased to 6.
The initial specific xylose consumption rates are also
summarized in Table 1. It is evident that xylose consump-
tion was strongly inhibited by acetic acid; a significant
decrease in the rate was observed with both increasing acetic
acid concentrations and decreasing pH values. Under the
harshest condition (pH 5 and 15 g L�1 acetic acid), no
significant xylose consumption was observed. Results from
a similar study (Bellissimi et al., 2009) did not show such a
severe decrease in the xylose consumption rate for the
fermentation conditions of pH 5 with 3 g L�1 acetic acid.
However, the concentrations examined in this study were
2.5–5 times greater than those examined by Bellissimi and
colleagues.
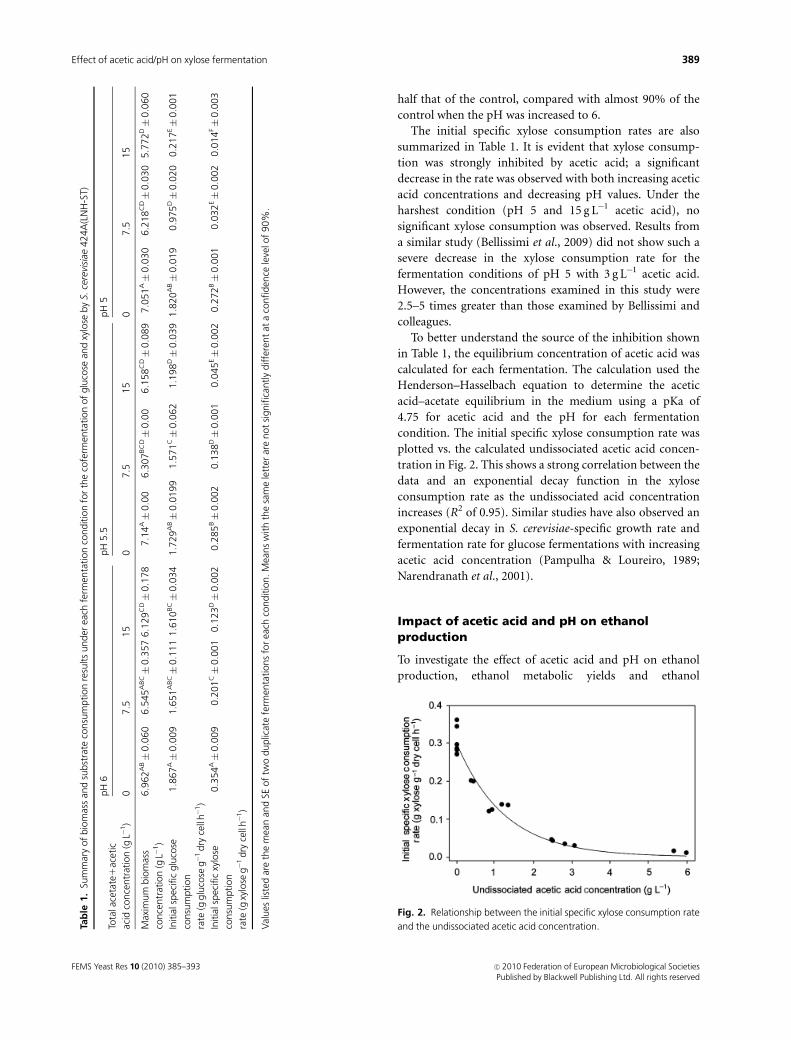
To better understand the source of the inhibition shown
in Table 1, the equilibrium concentration of acetic acid was
calculated for each fermentation. The calculation used the
Henderson–Hasselbach equation to determine the acetic
acid–acetate equilibrium in the medium using a pKa of
4.75 for acetic acid and the pH for each fermentation
condition. The initial specific xylose consumption rate was
plotted vs. the calculated undissociated acetic acid concen-
tration in Fig. 2. This shows a strong correlation between the
data and an exponential decay function in the xylose
consumption rate as the undissociated acid concentration
increases (R2 of 0.95). Similar studies have also observed an
exponential decay in S. cerevisiae-specific growth rate and
fermentation rate for glucose fermentations with increasing
acetic acid concentration (Pampulha & Loureiro, 1989;
Narendranath et al., 2001).
Impact of acetic acid and pH on ethanolproduction
To investigate the effect of acetic acid and pH on ethanol
production, ethanol metabolic yields and ethanol
Fig. 2. Relationship between the initial specific xylose consumption rate
and the undissociated acetic acid concentration.Tab
le1.
Sum
mar
yof
bio
mas
san
dsu
bst
rate
consu
mption
resu
lts
under
each
ferm
enta
tion
conditio
nfo
rth
eco
ferm
enta
tion
of
glu
cose
and
xylo
seby
S.ce
revi
siae
424A
(LN
H-S
T)
Tota
lace
tate
1ac
etic
acid
conce
ntr
atio
n(g
L�1)
pH
6pH
5.5
pH
5
07.5
15
07.5
15
07.5
15
Max
imum
bio
mas
s
conce
ntr
atio
n(g
L�1)
6.9
62
AB�
0.0
60
6.5
45
ABC�
0.3
57
6.1
29
CD�
0.1
78
7.1
4A�
0.0
06.3
07
BC
D�
0.0
06.1
58
CD�
0.0
89
7.0
51
A�
0.0
30
6.2
18
CD�
0.0
30
5.7
72
D�
0.0
60
Initia
lspec
ific
glu
cose
consu
mption
rate
(gglu
cose
g�1
dry
cell
h�1)
1.8
67
A�
0.0
09
1.6
51
ABC�
0.1
11
1.6
10
BC�
0.0
34
1.7
29
AB�
0.0
199
1.5
71
C�
0.0
62
1.1
98
D�
0.0
39
1.8
20
AB�
0.0
19
0.9
75
D�
0.0
20
0.2
17
E�
0.0
01
Initia
lspec
ific
xylo
se
consu
mption
rate
(gxy
lose
g�1
dry
cell
h�1)
0.3
54
A�
0.0
09
0.2
01
C�
0.0
01
0.1
23
D�
0.0
02
0.2
85
B�
0.0
02
0.1
38
D�
0.0
01
0.0
45
E�
0.0
02
0.2
72
B�
0.0
01
0.0
32
E�
0.0
02
0.0
14
F�
0.0
03
Val
ues
liste
dar
eth
em
ean
and
SEof
two
duplic
ate
ferm
enta
tions
for
each
conditio
n.M
eans
with
the
sam
ele
tter
are
not
signifi
cantly
diffe
rent
ata
confiden
cele
velo
f90%
.
FEMS Yeast Res 10 (2010) 385–393 c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
389Effect of acetic acid/pH on xylose fermentation

volumetric production rates were calculated. The ethanol
metabolic yields, calculated by dividing the observed etha-
nol concentrations by the theoretical ethanol concentrations
(0.51 times the total concentration of consumed glucose and
xylose), are provided in Table 2. If all of the consumed
glucose and xylose was converted to ethanol, the ethanol
metabolic yields would be equal to 1. However, a portion of
the consumed sugars resulted in the production of glycerol,
xylitol, and/or additional cell mass. Thus, the metabolic
yields are all o 1. The presence of acetic acid enhanced
rather than inhibited the yield of consumed sugars to
ethanol, as seen by an increase in the ethanol metabolic
yields as the concentration of acetic acid increased. At pH 5,
ethanol metabolic yield improvements of 20% and 30%
were observed in the presence of 7.5 and 15 g L�1 acetic acid,
respectively, as compared with the control.
In addition, ethanol volumetric production rates
(Table 2) were calculated by dividing the maximum ethanol
concentration by the fermentation time required to reach
that concentration. Significant decreases in productivity
were seen in the presence of acetic acid. Comparing the least
and the most severe conditions (0 g L�1 acetic acid at pH 6
and 15 g L�1 at pH 5, respectively), a 67% decrease in
volumetric productivity was observed. The results also show
a linear increase in the production rates with increasing pH
(R2 of 0.89 and 0.99 for 7.5 and 15 g L�1 acetic acid
concentrations, respectively, plots not shown).
Discussion
The effect of acetic acid on non-pentose-fermenting
S. cerevisiae strains has been studied widely (Maiorella
et al., 1983; Pampulha & Loureirodias, 1989; Phowchinda
et al., 1995; Taherzadeh et al., 1997; Thomas et al., 2002).
Prior studies examined a range of acetic acid concentrations
(up to 12 g L�1) under a variety of uncontrolled or con-
trolled pH values (pH 2.8–5.5). A common observation was
that acetic acid resulted in a decrease in biomass yield
coupled to an increase in ethanol yield (Maiorella et al.,
1983; Taherzadeh et al., 1997; Thomas et al., 2002). How-
ever, the increased ethanol yields came at the cost of a
decreased ethanol production rate (Phowchinda et al.,
1995). The decrease in the fermentation rate was explained
by a decrease in intracellular pH, leading to the conclusion
that it is the concentration of the undissociated, uncharged
form of acetic acid that governs the inhibitory effect
(Pampulha & Loureirodias, 1989). Undissociated acetic acid
freely diffuses across the cell membrane and rapidly dis-
sociates because of the higher intracellular pH, resulting in
the release of protons into the cytoplasm. Plasma ATPase
pumps these protons out of the cell at the cost of ATP to
avoid intracellular acidification until the influx of protons
exceeds the cell’s proton-pumping capability and Tab
le2.
Sum
mar
yof
ethan
oly
ield
and
pro
duct
ivity
resu
lts
under
each
ferm
enta
tion
conditio
nfo
rth
eco
ferm
enta
tion
of
glu
cose
and
xylo
seby
Sacc
har
om
yces
cere
visi
ae424A
(LN
H-S
T)
Tota
lace
tate
1ac
etic
acid
conce
ntr
atio
n(g
L�1)
pH
6pH
5.5
pH
5
07.5
15
07.5
15
07.5
gL�
115
gL�
1
Ethan
olm
etab
olic
yiel
d0.7
85
BC�
0.0
12
0.8
57
ABC�
0.0
02
0.8
75
AB�
0.0
08
0.7
93
BC�
0.0
06
0.8
26
BC�
0.0
12
0.8
80
AB�
0.0
08
0.7
55
C�
0.0
02
0.8
92
AB�
0.0
62
0.9
56
A�
0.0
18
Ethan
olv
olu
met
ric
pro
duct
ion
rate
(get
han
olL�
1h�
1)
1.2
34
A�
0.0
27
0.7
84
C�
0.0
37
0.5
88
D�
0.0
09
1.1
77
A�
0.0
28
0.5
74
D�
0.0
44
0.4
74
DE�
0.0
13
0.9
41
B�
0.0
06
0.4
53
E�
0.0
08
0.4
03
E�
0.0
06
Val
ues
liste
dar
eth
em
ean
and
SEof
two
duplic
ate
ferm
enta
tions
for
each
conditio
n.M
eans
with
the
sam
ele
tter
are
not
signifi
cantly
diffe
rent
ata
confiden
cele
velo
f90%
.
FEMS Yeast Res 10 (2010) 385–393c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
390 E. Casey et al.

acidification of the cytoplasm cannot be avoided (Russell,
1992). ATP is also needed for the removal of excess acetate
from the cytoplasm through the energy-dependent weak
acid efflux pump, Pdr12 (Piper et al., 1998). The need for
ATP to maintain intracellular pH homeostasis and reduce
the internal acetate concentration explains the decreased
biomass and increased ethanol yields; glucose was diverted
from biomass generation to ethanol production to generate
the ATP needed for cell maintenance. Similar to the studies
discussed above, a decreased biomass concentration and
increased metabolic ethanol yields were seen in the glucose/
xylose cofermentations with acetic acid for the given study.
These results can also be explained by the diversion of
carbon from biomass growth to ethanol production for
ATP generation. The ethanol production rates also de-
creased in the presence of acetic acid in this study. Upon
comparison of the results, the rate Phowchinda et al. (1995)
achieved with 6 g L�1 acetic acid and uncontrolled pH
conditions was approximately the rate we achieved under
the harshest condition tested in the present work (15 g L�1 at
pH 5). This suggests that S. cerevisiae 424A(LNH-ST) was
more tolerant to acetic acid than the strain they used and/or
controlling the media pH can significantly mitigate inhibi-
tion by acetic acid.
Although there were many similarities in the experimen-
tal procedures and results from this study and the previous
studies, the difference in microorganisms (pentose vs. non-
pentose-fermenting strains) makes a direct comparison
difficult. A limited number of studies have been published
that investigate the impact of acetic acid on genetically
engineered S. cerevisiae strains capable of both glucose and
xylose fermentation (Helle et al., 2003; Bellissimi et al.,
2009). Bellissimi et al. (2009) looked at the effect of acetic
acid on an S. cerevisiae strain engineered with the xylose
isomerase pathway as opposed to the xylose reductase and
xylitol dehydrogenase pathway in the strain used in this
study. The conditions they examined were an acetic acid
concentration of 3 g L�1 under controlled pH values of 3.5 or
5. Helle et al. (2003) studied the effect of acetic acid on an
S. cerevisiae strain that was engineered with the xylose
reductase and xylitol dehydrogenase genes for xylose meta-
bolism, similar to the strain used in this study. Their
experimental conditions were 3 g L�1 acetic acid and initial
pH values of 4, 4.7, and 5.5. Similar to the results with the
non-pentose-fermenting S. cerevisiae strains, biomass yields
decreased in the presence of acetic acid for both studies.
Bellissimi et al. (2009) also showed that xylose consumption
rates were more affected by acetic acid than glucose con-
sumption rates, while Helle et al. (2003) noted a decrease in
ethanol production rates from the presence of acetic acid in
the fermentation media. Similar observations were made in
the present study, confirming the impact of acetic acid on
the cofermentation of glucose and xylose. The primary
difference from these results, when compared with the
results using non-pentose-fermenting S. cerevisiae strains,
was the increased inhibitory effect on xylose consumption
vs. glucose consumption. The rate of xylose consumption in
S. cerevisiae 424A(LNH-ST) is approximately 20% that of
glucose and the ATP yield is less (1.67 mol ATP mol�1 xylose
compared with 2.0 mol ATP mol�1 glucose). Thus, the esti-
mated ATP generation rate when xylose is the sole carbon
source is approximately 17% of the ATP generation rate
when glucose is fermented. This is likely a major reason why
the inhibitory effect of acetic acid on xylose consumption is
more severe than on glucose consumption. Bellissimi and
colleagues showed that a continuous feed of glucose at low
concentrations releases some of the inhibition of xylose
consumption, further supporting this conclusion. Bellissimi
et al. (2009) also observed increasing glucose consumption
rates with 3 g L�1 acetic acid present, a trend not seen in this
study. Low amounts of acetic acid have been shown to
stimulate fermentation (Taherzadeh et al., 1997; Thomas
et al., 2002). However, the acetic acid concentrations tested
in this study exceeded those amounts by a factor of 2 or
more.
Conclusions
The presence of inhibitory compounds in biomass hydro-
lysates is a major obstacle facing the cellulosic ethanol
industry because they impact the fermentative performance
of microorganisms negatively. The effect of acetic acid, one
of these inhibitors, on the cofermentation of glucose and
xylose by S. cerevisiae 424A(LNH-ST) under controlled pH
conditions was examined. Acetic acid inhibited biomass
growth, substrate consumption, and ethanol volumetric
productivity. However, acetic acid enhanced ethanol meta-
bolic yield. Significant decreases in the maximum biomass
concentration were observed in the presence of acetic acid,
with this effect becoming less severe as the pH was increased.
Similar trends were noted with glucose and xylose initial
specific consumption rates, with the effect being more
significant on xylose consumption. An exponential relation-
ship was found between the initial specific xylose consump-
tion rates and the concentration of undissociated acetic acid,
confirming that the inhibitory effect of acetic acid is linked
to the undissociated form of acetic acid. Ethanol production
rates decreased considerably in the presence of acetic acid.
However, no inhibitory effect was seen with the ethanol
metabolic yields; rather, acetic acid was shown to improve
these yields. The impact of media pH was also investigated
in this study. The results suggest that increasing media pH
can alleviate some of the inhibitory effect of the acetic acid
as this causes a decrease in the concentration of undisso-
ciated acetic acid, the inhibitory form of acetic acid for
S. cerevisiae fermentations.
FEMS Yeast Res 10 (2010) 385–393 c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
391Effect of acetic acid/pH on xylose fermentation

Acknowledgements
This work was supported by the US Department of Energy
Biomass Program, Contract GO17059-16649, and Purdue
Agricultural Research Programs. This material is based on
work supported under a National Science Foundation
Graduate Research Fellowship. Any opinions, findings, con-
clusions, or recommendations expressed in this publication
are those of the authors and do not necessarily reflect the
views of the National Science Foundation. The authors
thank Haroon Mohammad for the preliminary work he
completed on this project. They also thank Eduardo Xi-
menes and Chialing Wu for their internal review of this
manuscript.
References
Almeida JRM, Modig T, Petersson A, Hahn-Hagerdal B, Liden G
& Gorwa-Grauslund MF (2007) Increased tolerance and
conversion of inhibitors in lignocellulosic hydrolysates by
Saccharomyces cerevisiae. J Chem Technol Biot 82: 340–349.
Barnett JA (1976) The utilization of sugars by yeasts. Adv
Carbohyd Chem Bi 32: 125–234.
Bellissimi E, van Dijken JP, Pronk JT & van Maris AJA (2009)
Effects of acetic acid on the kinetics of xylose fermentation by
an engineered, xylose-isomerase-based Saccharomyces
cerevisiae strain. FEMS Yeast Res 9: 358–364.
Davis L, Rogers P, Pearce J & Peiris P (2006) Evaluation of
Zymomonas-based ethanol production from a hydrolysed
waste starch stream. Biomass Bioenerg 30: 809–814.
Eliasson A, Christensson C, Wahlbom CF & Hahn-Hagerdal B
(2000) Anaerobic xylose fermentation by recombinant
Saccharomyces cerevisiae carrying XYL1, XYL2, and XKS1 in
mineral medium chemostat cultures. Appl Environ Microb 66:
3381–3386.
Hahn-Hagerdal B, Karhumaa K, Fonseca C, Spencer-Martins I &
Gorwa-Grauslund MF (2007) Towards industrial pentose-
fermenting yeast strains. Appl Microbiol Biot 74: 937–953.
Helle S, Cameron D, Lam J, White B & Duff S (2003) Effect of
inhibitory compounds found in biomass hydrolysates on
growth and xylose fermentation by a genetically engineered
strain of S. cerevisiae. Enzyme Microb Tech 33: 786–792.
Ho NWY & Chen ZD (1997) Stable recombinant yeasts capable of
effective fermentation of both glucose and xylose. PCT Patent
No. WO97/42307.
Ho NWY, Chen ZD & Brainard AP (1998) Genetically engineered
Sacccharomyces yeast capable of effective cofermentation of
glucose and xylose. Appl Environ Microb 64: 1852–1859.
Ho NWY, Chen Z, Brainard AP & Sedlak M (2000) Genetically
engineered Saccharomyces yeasts for conversion of cellulosic
biomass to environmentally friendly transportation fuel
ethanol. ACS Symposium Series 767 (Anastas PT, Heine LG &
Williamson TC, eds), pp. 143–159. American Chemical
Society, Washington, DC.
Klinke HB, Thomsen AB & Ahring BK (2004) Inhibition of
ethanol-producing yeast and bacteria by degradation products
produced during pre-treatment of biomass. Appl Microbiol
Biot 66: 10–26.
Kuyper M, Hartog MMP, Toirkens MJ, Almering MJH, Winkler
AA, Dijken JP & Pronk JT (2005) Metabolic engineering of a
xylose-isomerase-expressing Saccharomyces cerevisiae strain
for rapid anaerobic xylose fermentation. FEMS Yeast Res 5:
399–409.
Lawford HG & Rousseau JD (1993) The effect of acetic acid on
fuel ethanol production by Zymomonas. Appl Biochem Biotech
39/40: 687–699.
Lindsay SE, Bothast RJ & Ingram LO (1995) Improved strains of
recombinant Escherichia coli for ethanol production from
sugar mixtures. Appl Microbiol Biot 43: 70–75.
Lu Y, Warner R, Sedlak M, Ho N & Mosier NS (2009)
Comparison of glucose/xylose cofermentation of poplar
hydrolysates processed by different pretreatment technologies.
Biotechnol Progr 25: 349–356.
Luck E & Jager M (1997) Antimicrobial Food Additives:
Characteristics, Uses, Effects. Springer, New York.
Luli GW & Strohl WR (1990) Comparison of growth, acetate
production, and acetate inhibition of Escherichia coli strains in
batch and fed-batch fermentations. Appl Environ Microb 56:
1004–1011.
Maiorella B, Blanch HW & Wilke CR (1983) By-product
inhibition effects on ethanolic fermentation by Saccharomyces
cerevisiae. Biotechnol Bioeng 25: 103–121.
Mosier N, Wyman C, Dale B, Elander R, Lee YY, Holtzapple M &
Ladisch M (2005) Features of promising technologies for
pretreatment of lignocellulosic biomass. Bioresource Technol
96: 673–686.
Narendranath NV, Thomas KC & Ingledew WM (2001) Effects of
acetic acid and lactic acid on the growth of Saccharomyces
cerevisiae in a minimal medium. J Ind Microbiol Biot 26:
171–177.
Palmqvist E & Hahn-Hagerdal B (2000) Fermentation of
lignocellulosic hydrolysates. II: inhibitors and mechanisms of
inhibition. Bioresource Technol 74: 25–33.
Pampulha ME & Loureiro V (1989) Interaction of the effects of
acetic acid and ethanol on inhibition of fermentation in
Saccharomyces cerevisiae. Biotechnol Lett 11: 269–274.
Pampulha ME & Loureirodias MC (1989) Combined effect of
acetic acid, pH, and ethanol on intracellular pH of fermenting
yeast. Appl Microbiol Biot 31: 547–550.
Phowchinda O, Deliadupuy ML & Strehaiano P (1995) Effects of
acetic acid on growth and fermentative activity of
Saccharomyces cerevisiae. Biotechnol Lett 17: 237–242.
Piper P, Mahe Y, Thompson S et al. (1998) The Pdr 12 ABC
transporter is required for the development of weak organic
acid resistance in yeast. EMBO J 17: 4257–4265.
Russell JB (1992) Another explanation for the toxicity of
fermentation acids at low pH – anion accumulation versus
uncoupling. J Appl Bacteriol 73: 363–370.
FEMS Yeast Res 10 (2010) 385–393c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
392 E. Casey et al.

Shaw AJ, Podkaminer KK, Desai SG et al. (2008) Metabolic
engineering of a thermophilic bacterium to produce ethanol at
high yield. P Natl Acad Sci USA 105: 13769.
Sonderegger M, Jeppsson M, Larsson C et al. (2004) Fermentation
performance of engineered and evolved xylose-fermenting
Saccharomyces cerevisiae strains. Biotechnol Bioeng 87.
Stephanopoulos G (2007) Challenges in engineering microbes for
biofuels production. Science 315: 801–804.
Taherzadeh MJ, Niklasson C & Liden G (1997) Acetic acid – friend
or foe in anaerobic batch conversion of glucose to ethanol by
Saccharomyces cerevisiae? Chem Eng Sci 52: 2653–2659.
Takahashi CM, Takahashi DF, Carvalhal MLC & Alterthum F
(1999) Effects of acetate on the growth and fermentation
performance of Escherichia coli KO11. Appl Biochem Biotech
81: 193–203.
Thomas KC, Hynes SH & Ingledew WM (2002) Influence of
medium buffering capacity on inhibition of Saccharomyces
cerevisiae growth by acetic and lactic acids. Appl Environ
Microb 68: 1616–1623.
Wang DIC, Clooney CL, Demain AL, Dunnill P, Humphrey AE &
Lilly MD (1979) Fermentation and Enzyme Technology. John
Wiley & Sons, New York.
Zhang M, Eddy C, Deanda K, Finkelstein M & Picataggio S
(1995) Metabolic engineering of a pentose metabolism
pathway in ethanologenic Zymomonas mobilis. Science 267:
240.
FEMS Yeast Res 10 (2010) 385–393 c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
393Effect of acetic acid/pH on xylose fermentation





























