Embed Size (px)
Citation preview

American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
1
Effect of Winery Yeast Lees on Touriga Nacional 1 Red Wine Color and Tannin Evolution 2
Ana Rodrigues,1,2 Jorge Manuel Ricardo-Da-Silva,3* Carlos Lucas,4 and Olga Laureano5 3 1PhD Student, 3Associate Professor of Enology, 5Full Investigator and Professor of Enology, Technical University of 4 Lisbon, Instituto Superior de Agronomia, Laboratório Ferreira Lapa (Sector de Enologia), 1349-017 Lisboa, 5 Portugal; and 2PhD Student in a company-based PhD program, 4Winemaker, Dão Sul – Sociedade Vitivinícola, S.A., 6 Quinta das Sarzedas, 3430-909 Carregal do Sal, Portugal. 7 *Corresponding author (email: [email protected]; tel: +351 213 653 542) 8 Acknowledgments: The authors thank to Fundação para a Ciência e Tecnologia for the A. Rodrigues PhD grant 9 (SFRH/BDE/15598/2006). 10 Manuscript submitted Oct 2011, revised May 2012, accepted Jul 2012 11 Copyright © 2012 by the American Society for Enology and Viticulture. All rights reserved. 12 13
Abstract: Red wine aging on lees is a winemaking practice used to achieve more rounded and less 14
astringent resulting wines. In the present work the effect of adding two different winery external yeast lees 15
to a red wine on its color and tannin evolution during earlier stage of aging was studied. The results 16
indicated that the yeast lees addition did not show an effect on color stabilization, during the studied period 17
of time. Both color compounds and condensed tannins were quickly adsorbed to the yeast lees at the 18
beginning of the experiment. There was a retarding effect on the proanthocyanidin polymerization reaction 19
by the addition of yeast lees, leading to the maintenance of the low and medium size tannins in solution. 20
Thus, it was observed two different interactions: first, the proanthocyanidin adsorption by the yeast lees, 21
namely the ones with the highest polymerization degree and second, the retarding of proanthocyanidin 22
polymerization probably by the yeast lees released mannoproteins. The yeast lees age was a conditioning 23
factor in the mannoprotein release and on its effect in wine. 24
Keywords: Red wine, yeast lees, color, proanthocyanidins, mannoproteins 25
26
27
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
AJEV Papers in Press. Published online October 5, 2012.

American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
2
Introduction 28
Red wine polyphenols are one of the most important and studied themes in oenology, namely anthocyanins 29
and proanthocyanidins. Anthocyanins are responsible for wine red color and they exist at wine pH in 30
colored forms and especially condensed with flavanol units and acetaldehyde in polymeric pigments 31
(Somers 1971, Dallas et al. 1996). Proanthocyanidins or condensed tannins are related with wine 32
astringency, bitterness and color and they are polymers resulting from the condensation between flavan-3-33
ol units. They are composed by procyanidins ((+)-catechin and (-)-epicatechin) and prodelphinidins ((-)-34
epigallocatechin), sometimes esterified by gallic acid (Haslam 1980, Ricardo da Silva et al. 1990, Fulcrand 35
et al. 1999). 36
Red wine aging on lees turn out to be a winery practice very used all over the wine producing world 37
(Fornairon-Bonnefond et al. 2002). The objective of this procedure is to improve wine mouthfeel 38
characteristics, by the achievement of a more rounded and less astringent wine. Yeast lees also enhance 39
bacterial activity and growth during malolactic fermentation (Ribéreau-Gayon et al. 1998). Besides the 40
usage of the red wine fine lees, it can be added external yeast lees to the wine. The external yeast lees can 41
be used in the form of a commercial oenological product or by taking advantage of the winery alcoholic 42
fermentation by-products like the yeast lees produced in other wines fermentation and aging. 43
Wine fine lees are a microorganism’s reservoir, constituted mainly by the alcoholic fermentation yeasts 44
and some bacteria, but also by several other different components, mainly tartaric salts and organic 45
residues (Fornairon-Bonnefond et al. 2002). Their composition is variable and it depends on the yeasts 46
autolysis process, namely on the characteristics of the medium, the strain of yeasts used and the 47
fermentation temperature (Feuillat and Charpentier 1982). Autolysis is the breakdown of the yeast cell 48
components by the action of endogenous enzymes and it is characterized by the loss of cell organization, 49
degradation of cell macromolecules and by the releasing of the breaking products into the extracellular 50
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S

American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
3
environment (Hernawan and Fleet 1995). The autolysis main products are amino acids, proteins, 51
polysaccharides, lipids and nucleic acids. 52
One of the principal products derived from the yeast autolysis are mannoproteins. These polysaccharides 53
are released into the medium after the action of the β-(1→3) glucanases on the cell wall glucans (Feuillat 54
et al. 1989). Escot et al. (2001) verified that the tannins structure is affected by the addition of 55
mannoproteins, reducing the wine astringency and suggesting that these polysaccharides retain 56
anthocyanins and tannins, preventing their precipitation. Riou et al. (2002) studied the interaction between 57
seed tannins and wine polysaccharides in a model solution, and concluded that mannoproteins have a 58
protective effect on tannin aggregation, though they did not prevent their initial aggregation. Poncet-59
Legrand et al. (2007) have shown in a model wine that mannoproteins of low and medium molecular 60
weight (51 and 62 kDa) are more effective probably due to a steric hindrance mechanism although all 61
mannoprotein fractions prevented tannins aggregation and precipitation. On the other hand, Guadalupe et 62
al. (2007) verified that commercial added mannoproteins did not protected polyphenols from precipitation 63
neither did protected wine color, though they contributed to a better mouthfeel and structure, resulting in a 64
more rounded and sweet wine. On another work, Guadalupe and Ayestarán (2008) have shown that during 65
wine aging there is a precipitation of mannoprotein-tannin aggregates as well as of mannoprotein-pigments 66
aggregates, suggesting that mannoproteins do not act as protective colloids. 67
Yeast lees can also react directly with wine polyphenols mainly by adsorption mechanisms (Salmon et al. 68
2002) and this kind of interference results in a decrease of the wine free polyphenols. Medina et al. (2005) 69
have demonstrated that the yeast strain used in the alcoholic fermentation is an important choice in what 70
matters to preserve the wine color during the vinification process. Polar condensed tannins, with more 71
epigallocatechin units, are preferentially adsorbed by yeast lees and there is no preferential adsorption 72
between low and high tannin polymers (Mazauric and Salmon 2005). The anthocyanins that remain 73
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S

American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
4
adsorbed to the lees have a dominant blue color, (Morata et al. 2003, Mazauric and Salmon 2006) resulting 74
in an enhancement of the yellow anthocyanins in the medium. Vasserot et al. (1997) verified that the 75
adsorption of anthocyanins to yeast lees results from the establishment of hydrogen bonds. They suggested 76
that the adsorption capacity depended on the anthocyanin polarity, being the polar anthocyanins the more 77
reactive. On the other hand, Mazauric and Salmon (2006) verified that anthocyanin adsorption on yeast 78
lees does not follow a simple mechanism of hydrogen bonding and that there is no relationship between 79
anthocyanin polarity and adsorption capacity. 80
Empirically, some winemakers add the yeast lees resulting from the white wines winemaking process to 81
red wines in aging period in order to achieve more rounded and better mouthfeel wines instead of adding 82
commercial enological products. Considering that wine yeast lees are a great source of mannoproteins and 83
that this winery by-product is a possible resource to obtain naturally and economically produced 84
mannoproteins, the aim of this work was to understand the effect of adding two different winery derived 85
yeast lees to a Touriga Nacional (Vitis vinifera , L. cv.) grapevine variety red wine on its color and tannin 86
evolution during aging, according to the normal practices used at Dão Sul’s winery. The two types of yeast 87
lees used resulted from the alcoholic fermentation of an Encruzado (Vitis vinifera , L. cv.) grapevine 88
variety white wine: the first ones coming from the end of the white wine aging process in oak barrels and 89
the others coming from the very end of the white wine alcoholic fermentation. 90
Materials and Methods 91
Chemicals. All chemicals used were of analytical reagent grade. All solvents were of HPLC grade. (+)-92
catechin, (-)-epicatechin, (-)-epicatechin-3-O-gallate and (-)-epigallocatechin were purchased from 93
Extrasynthese (Genay, France). Toluene-α-thiol (phenylmethanethiol) was purchased from Fluka (Buchs, 94
Switzerland). 95
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S

American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
5
Wines. Grapes from Touriga Nacional (Vitis vinifera , L. cv.) were harvested manually and vinified at Dão 96
Sul winery (Dão Region, Portugal) in 2009. The grapes (9000 kg) were de-stemmed and crushed into a 97
stainless steel vessel and a preparation of commercial maceration enzymes (Vinozym Vintage FCE, 98
Novozymes, Denmark) was added. After 24 hours of cold pre-fermentative maceration at 15ºC the must 99
was inoculated with a commercial activated Saccharomyces cerevisiae cerevisiae preparation (Lalvin 100
QD145, Lallemand, Canada). The alcoholic fermentation occurred for ten days at 20 to 25ºC. After the 101
beginning of the alcoholic fermentation, the must was punched down for 20 minutes every three hours, and 102
it was submitted to a rack and return program for half an hour every day, until the end of alcoholic 103
fermentation. After the end of alcoholic fermentation, the free-run wine was transferred to another 104
stainless steel vessel in order to perform malolactic fermentation. The wine was analyzed for ethanol 105
content, pH, volatile acidity, titratable acidity and free and total SO2 according to the Organisation 106
International de la Vigne et du Vin official methods (OIV 2006). The wine chemical parameters were the 107
following: alcohol content 13.6% v/v, titratable acidity 7.8 g/L expressed in tartaric acid, volatile acidity 108
0.26 g/L expressed in acetic acid, pH 3.75, residual sugars 2.3 g/L, 15 mg/L of free SO2, 32 mg/L of total 109
SO2. 110
White wine yeast lees preparation and trials in red wines. Two different white wine derived yeast lees 111
were prepared in order to be added to the red wine. The first yeast lees were prepared from winery 112
resulting lees from the technological process of an Encruzado (Vitis vinifera, L. cv.) grape variety 113
harvested in 2009 in the Dão Sul winery. The Encruzado grapes free running juice was cooled at 5ºC, 114
added with pectinolytic enzymes (Novoclair Speed, Novozymes, Denmark) and it settled down for 24 115
hours. After decantation, the must was inoculated with a commercial active dry yeasts preparation 116
(Saccharomyces cerevisiae var. bayanus, Enoferm QA23, Lallemand, Canada), fermenting at 15 to 18ºC, 117
for three weeks. At the end of the fermentation, the wine was submitted to bâtonnage by manual stirring 118
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S

American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
6
with a stainless steel stick for 30 seconds per barrel, in an interval between two and three days during one 119
month after the end of alcoholic fermentation. Then the lees were allowed to settle down until being stirred 120
once again 100 days after the end of alcoholic fermentation. Yeast lees were taken from the white wine at 121
the time it was separated from lees for racking, six months after the end of alcoholic fermentation. The lees 122
were then centrifuged at 3500 r.p.m. for ten minutes and the resulting pellet was added to the red wine in 123
the normal winery used proportion of 5% (w/v) of the total volume – BE wine. In order to understand if 124
the age of the lees would have any influence in the color and tannins stabilization, a second trial was 125
prepared using newly fermented yeast lees. These yeast lees were prepared from a newly fermented 126
Encruzado clarified must as described above. Four liters of Encruzado must was fermented with the same 127
commercial active dry yeast preparation than the white wine obtained for BE trial. The fermentation 128
occurred at controlled temperature (35ºC) for four days. This high temperature was chosen in order to 129
accelerate the fermentation process. One day after the end of alcoholic fermentation the yeast lees were 130
centrifuged at 3500 r.p.m. for ten minutes. The resulting pellet was added to the wine in the proportion of 131
0.8% (w/v), the same proportion that yeast lees represented in the white fermented wine – LN wine. T 132
wine refers to the wine with no addition of yeast lees. The three wines were kept in amber flasks (2.5 L), at 133
35ºC, for sixty days. Only one trial was performed for each wine modality as the usage of the same yeast 134
lees, the same wine and the procedure of stirring, the experimental variability concerning the 135
mannoproteins release was expected to be the same. Samples of each wine were analyzed for their color 136
characterization and proanthocyanidins composition according to their degree of polymerization at 0, 6, 137
13, 19, 26, 47 and 60 days. The analyses were performed in duplicate except if the duplicated values were 138
quite different and it would be repeated once again. 139
Color characterization. In order to characterize the wine color compounds it was used the 140
spectrophotometrical method described by Somers and Evans (1977). The wines were centrifuged for ten 141
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S

American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
7
minutes at 3.500 r.p.m. and the absorbances were measured using a Unicam UV–Vis UV4 142
spectrophotometer (Unicam, Cambridge, UK). 143
Isolation of proanthocyanidins. The isolation of wine proanthocyanidins was made according to the 144
methods described by Sun et al. (1999) and Labarbe et al. (1999). Four milliliters of wine were injected 145
onto a Toyopearl TSK HW-40F (Tosoh Corp., Tokyo, Japan) packed column (100x10 mm) and first 146
washed with a solution of ethanol:water:TFA (55:45:0.05 v/v/v) in order to remove small molecules and 147
monomeric flavanols. The wine oligomeric and polymeric proanthocyanidins were eluted with a solution 148
of acetone:water (40:60 v/v). After evaporation of the tannin fraction at 30ºC under vacuum, the 149
proanthocyanidins were resuspended in 1 ml of methanol to be used in further analysis. 150
Fractionation of proanthocyanidins. The proanthocyanidins were separated according to their degree of 151
polymerization as described by Labarbe et al. (1999) during the experiment time. One milliliter of 152
proanthocyanidins methanol solution was precipitated by chloroform on the top of a glass powder column 153
(50x10 mm). The elution gradient (chloroform/methanol) was applied as following: FI –75:25 (v/v); FII – 154
70:30 (v/v); FIII – 65:35 (v/v); FIV – 60:40 (v/v); FV – 55:45 (v/v); FVI – 50:50 (v/v); FVII – 45:55 (v/v); 155
FVIII – 0:100 (v/v). After evaporation of each tannin fraction at 30ºC under vacuum, the 156
proanthocyanidins were resuspended in 0.5 ml of methanol. The eight fractions were analyzed by HPLC 157
after thiolysis in order to determine their structural characteristics. 158
Characterization of proanthocyanidins. The proanthocyanidins were submitted to depolymerization in 159
the presence of toluene-α-thiol in an acidic medium as described by Monagas et al. (2003). One hundred 160
microliters of toluene-α-thiol 5% in methanol 0.2 M HCl were added to 100 µL of proanthocyanidins 161
solution in a hermetically sealed glass tube. The mixture was warmed up to 55ºC on a water bath for 7 162
min. Ten microliters of the prepared solution were immediately injected in a HPLC system equipped with 163
a Merck Hitachi Lachrom L-7100 Pump (Merck Hitachi, Tokyo, Japan), followed by a Phenomenex 164
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S

American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
8
Gemini C18 110 A 150 x 3.0 mm column (Phenomenex, Torrance, California, USA) and a UV/Vis 165
Waters 2487 Dual Wavelength detector (Waters, Milford, Massachusetts, USA), in the following elution 166
conditions: 0.8 ml/min flow with a linear gradient of 15 to 75% of a solution of acetonitrile/water/formic 167
acid (80:18:2, v/v/v) in a solution of water/formic acid (98:2, v/v), in an 18 minutes run. The detection was 168
monitored at 280 nm and 320 nm. The amounts of monomers and of toluene-α-thiol adducts released were 169
calculated from the areas of the chromatographic peaks at 280 nm by comparison with calibration curves 170
(Rigaud et al. 1991). 171
Polysaccharide characterization. In order to characterize the white wine derived lees for their 172
polysaccharide composition, both lees were digested with a commercial preparation of β-glucanases from 173
Trichoderma harzanium as described by Moine-Ledoux et al. (1997). The schematic procedure for the 174
characterization of the yeast lees is represented in Figure 1. The yeast lees were diluted in distillated water 175
in a concentration of 15% (v/v). Glucanex (Novozymes, Bagsvaerd, Denmark) was added in a 176
concentration of 1% of the lees weight and the mixture was kept at 42ºC for five hours. After this period 177
the mixture was centrifuged, dialyzed against water and freeze-dried. The mixture was dissolved in water 178
and injected onto a Concanavalin-A Sepharose 4B (GE Healthcare Bio-Sciences, Uppsala, Sweden) 179
packed column (100x10 mm), and eluted with a sodium acetate-HCL 50 mM pH 5.6, NaCl 150 mM, 180
CaCl2 1 mM, MgCl2 1 mM and MnCl2 1 mM buffer solution at 0.8 ml/min, monitored by a refraction 181
index and a 254 nm wavelength detectors as described by Gonçalves et al. (2002), constituting the 182
unretained fraction. The retained fraction was eluted with the same buffer solution added with methyl α-183
D-mannopyranoside 500 mM. The two fractions (unretained and retained fractions) were dialyzed against 184
water for seven days, at 4ºC. The process was monitored with a Denver Instrument Model 220 185
condutivimeter (Denver Instrument, New York, USA). After freeze drying, each fraction (1 g/L) was 186
injected onto a FPLC system equipped with a Pharmacia LKB Pump P-500, a 12HR 10/30 FPLC size-187
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S

American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
9
exclusion column (GE Healthcare Bio-Sciences, Uppsala, Sweden) and eluted with an ammonium acetate 188
0.3 M buffer solution at 0.3 ml/min and monitored by a Perkin-Elmer LC-30 RI refractive index detector 189
(Perkin-Elmer, Waltham, Massachusetts, USA) modified with a LED (light-emitting diode) light source 190
(Cromolab, Queijas, Portugal) and a Knauer WellChrom Spectro-Photometer K-2501 wavelength detector 191
(Knauer, Berlin, Germany) at 254 nm. Calibration of the system was performed with Shodex P-82 Pullulan 192
standards (Showa Denko K.K., Kanagawa, Japan). 193
The carbohydrate composition was determined by gas chromatography after derivatization of the samples 194
into their alditol acetates according to Albersheim et al. (1967). One hundred microliters of a μ-inositol 195
solution (1 mg/ml) and 1 ml of trifluoroacetic acid 2 M were added to 1 ml of polysaccharides solution (1 196
mg/ml). After hydrolysis at 120ºC, for 75 minutes, the mixture was washed with 5 ml of water and dried. 197
Five hundred microliters of a saturated sodium borohydride solution in ammonia were added and the 198
mixture reacted for two hours at room temperature. The reaction was stopped by adding some drops of 199
glacial acetic acid and the mixture was washed with 5 ml of a solution of 1% HCl in methanol and dried. 200
One hundred and fifty microliters of pyridine and 150 μL of acetic anhydride were added to the mixture 201
and it reacted for 12 hours at room temperature. The reaction was stopped by adding a drop of water in an 202
ice bath. The mixture was washed with 5 ml water, followed by 1 ml of ethanol, and dried. The alditol 203
acetates were extracted to 200 μL of chloroform and were quantified on a CE Instruments GC 8000 Top 204
gas chromatographer (Thermo Fisher Scientific, Milan, Italy) with a capillary column Zebron ZB-Wax 10 205
60x0.25 mm, 0.25 µm film (Phenomenex, Torrance, California, USA) and a FID detector. The column 206
temperature was initially set at 220ºC for four minutes and raised to 235ºC at 10ºC/min, maintaining this 207
temperature for five minutes. Hydrogen was used as carrier gas at 1mL/min. µ-inositol was used as the 208
internal standard and sugars quantification was made after determination of each sugar response factor 209
using pure sugars for this purpose. 210
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S

American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
10
The total protein content was determined as described by Lowry et al. (1951) using bovine serum albumin 211
fraction V (Sigma-Aldrich, St. Louis, Missouri, USA) for the calibration curve. The total polysaccharides 212
content was determined by the phenol-sulfuric method as described by Dubois et al. (1956) using glucose 213
(Panreac, Barcelona, Spain) for the calibration curve as glucose was the most common sugar for 214
calibration curve referred in the literature. 215
Sensory Analysis. Wines were submitted to a tasting panel of eleven expert tasters from the Technical 216
University of Lisbon and Dão Sul wine producing company. The tasters had previous training in sensory 217
analysis. The three wines were tasted (T, BE and LN) at the end of the experiment and at 18ºC, in an 218
acclimatized tasting room at 20ºC. The wines were tasted one time by each taster in one single session. 219
The glasses used were an official tasting glass in tulip shape with 60 ml of wine. It was used a descriptive 220
tasting sheet considering the following parameters: Color – intensity; Aroma – fruity, floral, intensity, 221
persistency, equilibrium; Mouthfeel – body, bitterness, astringency, acidity, persistency, equilibrium; 222
overall balance. These parameters were generated according to the objective of the experiment. All 223
parameters were classified in a scale from 1 (absent) to 5 (very intense), except for aroma equilibrium, 224
mouthfeel equilibrium and overall balance parameters where the classification was made in a scale from 1 225
(bad) to 5 (excellent). All attributes were classified according to their perceived intensity by the tasters. 226
Significant differences between results were analyzed using the Statistica 6.0 software (Statsoft, Tulsa, 227
Oklahoma, USA). The values for each tasting descriptor were submitted to a single-way analysis of 228
variance (ANOVA) as the tasting panel was a trained one. Differences between samples always refer to 229
significant differences with at least p < 0.05. 230
Results 231
Characterization of the white wine yeast lees. The results obtained in the characterization of white wine 232
yeast lees are represented in Table 1. The characterization of mannoproteins regarding the sugars profile is 233
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S

American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
11
given from column 6 to 15, obtained in percentages from the GC analysis of each fraction (peak) achieved 234
from the FPLC size exclusion chromatography. The molecular weight for each of these fractions is 235
presented in column 3. The main goal of this characterization was to understand if the BE and LN lees 236
were chemically different although the grape must and the yeast strain used in the alcoholic fermentation 237
were the same for both lees. The main goal of this characterization was to understand if the BE and LN 238
lees were chemically different although the grape must and the yeast strain used in the alcoholic 239
fermentation were the same for both lees. The total colloids extracted from the yeast lees by the action of 240
β-glucanase represented 1.2% and 1.3% (w/w) of the centrifuged yeast lees total weight for BE and LN, 241
respectively. In both cases, the unretained Concanavalin-A fractions represented the lesser amount of the 242
total colloids obtained from the digestion of the yeast lees (21% for BE yeast lees and 13% for LN yeast 243
lees). The retained fraction, where the mannoproteins should be, represented 56% and 65% of the BE and 244
LN total colloids, respectively. 245
The molecular weight separation of the concanavalin-A unretained (BE FNR) and retained (BE FR) 246
fractions is shown in Figures 2 a) and b)). Concerning BE FNR sample there were three different peaks 247
with an average molecular weight of 177, 23 and 3 kDa, respectively. These peaks presented high values 248
in arabinose and galactose. The 177 kDa and 23 kDa peaks presented around 9% of arabinose and 90% of 249
galactose. The 3 kDa peak presented 18% of arabinose and nearly 55% of galactose. Considering the 250
residual sugars and protein composition of each molecular weight separated fraction, the three different 251
peaks were possibly polysaccharides rich in arabinose and galactose (PRAGs). It was possible that due to 252
the fact that the yeast lees were obtained from the fermentation of a white wine must, they also had grape 253
derived polysaccharides on their composition, like PRAGs. 254
The BE FR sample showed two well defined peaks with average molecular weights of 151 and 23 kDa and 255
an intermediate zone that seemed to be a third compound with an average molecular weight of 54 kDa. As 256
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S

American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
12
it is possible to see on Table 1, the 151 kDa peak had a similar content of mannose (52%) and glucose 257
(47%), and a proteic part. As the added yeast lees are not the pure fermented yeasts, the polysaccharide 258
found can come from a non-completed hydrolysis of this polysaccharide (glucan-mannoprotein polymer) 259
or its origin is other than the yeasts. The two other peaks, with 54 and 23 kDa average molecular weights 260
had a higher content in mannose (95% and 96% respectively) and also a proteic part in their composition. 261
As the two other peaks had a high content in mannose as well as a proteic part in their composition, they 262
represented mannoproteins. 263
Concerning LN yeast lees, the concanavalin-A retained (LN FR) and unretained (LN FNR) fractions 264
molecular weight distributions are represented on Figures 2 c) and d). The molecular weight distribution 265
showed the existence of three different macromolecules in both samples. The LN FNR average molecular 266
weights were 159, 23 and 2 kDa. The first peak had a high content in arabinose (15%) and galactose 267
(84%), the second peak had very high protein content and the third peak had a very high content in 268
arabinose (40%) and galactose (60%). The first peak of the LN FNR, corresponding to an average 269
molecular weight of 159 kDa compound, could possibly be a PRAG due to the high content in arabinose 270
and galactose. The second peak, with an average molecular weight of 23 kDa, had a very high protein 271
content, possibly being a glycoprotein. The third peak, with an average molecular weight of 2 kDa, also 272
had a very high content in arabinose and galactose, seeming to be a PRAG of small molecular weight. 273
The LN FR fraction molecular weight distribution had the following average molecular weights: 154, 50 274
and 25 kDa. The first peak had a high content in proteins with glucose being the main sugar (60%). The 275
second peak had a high content in mannose (89%) as well as a proteic part. The third peak had mannose 276
(67%) as the main sugar and also a proteic part. The LN FR first peak, corresponding to a 154 kDa average 277
molecular weight, had glucose as the main sugar and was possibly a glycoprotein. The second peak, with 278
an average molecular weight of 50 kDa, had a high content in mannose as well as a proteic part, being a 279
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S

American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
13
mannoprotein. The third peak, with an average molecular weight of 25 kDa, had mannose as the main 280
sugar and also a proteic part, being a mannoprotein. 281
Mannoprotein content in the newly fermented yeast lees (LN) was higher than in the old fermented yeast 282
lees (BE). This difference was probably due to the fact that BE spent more time in contact with the wine, 283
releasing a great quantity of mannoproteins into the medium which resulted in a decrease in the 284
mannoprotein content in the lees at the time they were added to the red wine. 285
Effect of white wine yeast lees addition on total polyphenol index and color compounds evolution. 286
The color parameters evolution is represented on Figure 3. Regarding total polyphenol index (I280), T 287
sample had some variations during the time, and it evolved for a final higher value than BE and LN 288
samples. BE slightly decreased in the first day of the experiment, increasing until the 19th day, and 289
stabilizing after this day until the end. LN sample maintained the same total polyphenol index until the end 290
of the experiment. 291
Regarding color intensity (CI), T maintained a constant evolution with no variations until the end of the 292
experiment, where it slightly increased. Both BE and LN samples had an initial quick decrease for CI due 293
to the yeast lees addition. Polymeric pigments (PP) represents longer chain colored compounds resulting 294
from the condensation reactions between anthocyanins and flavanol units (Somers 1971). PP content 295
evolution was similar to color intensity as well as the evolution of total anthocyanin content (TA) and total 296
pigments (TP). 297
Regarding hue evolution, all modalities showed a similar behavior, increasing through time and with 298
similar values. BE had a slightly higher final value, followed by T, and finally LN sample. 299
In spite of the results obtained during the planned trial, two years later it was observed that the LN wine 300
had a color intensity slightly higher (4.5) than the control (4.3) and the BE (3.9) wines. It was also 301
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S

American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
14
observed at long term a lower degree of polymerization of color pigments in LN wine (53%) compared 302
with control (59%) and BE (57%) wines. 303
Effect of white wine yeast lees on proanthocyanidins evolution. The evolution of total 304
proanthocyanidin content is shown in Figure 4 a). Wine with no yeast lees added (T wine) showed a 305
continuous decrease on total proanthocyanidins from the beginning until the end of the experiment. In 306
comparison, both wines added with yeast lees (BE and LN) had an initial greater decrease in the 307
proanthocyanidin content, observable from day 0 to day 5. The proanthocyanidin content in the wine 308
added with old fermented yeast lees (BE) decreased around 690 mg/L between day 0 and day 5, and in the 309
wine added with newly fermented yeast lees (LN) it decreased around 521 mg/L in the same period. 310
Concerning the evolution of BE wine between day 5 and the end of the experiment, there was a constant 311
proanthocyanidin content with almost no variation between the values through this period of time. LN 312
wine proanthocyanidin content decreased around 136 mg/L from day 5 until day 60. In comparison, T 313
wine had a decrease of 270 mg/L from day 5 until the end of the experiment, although it finished with a 314
higher quantity of proanthocyanidins. 315
The evolution of the overall degree of polymerization (mDP) of wines is shown in Figure 4b). The mDP 316
evolution of the wine with no added yeast lees (T wine) showed a slow decrease through time, starting 317
with an mDP of 15.2 and finishing with 11.1. Wine added with old fermented lees (BE) had a faster 318
decrease in the beginning of the experiment from day 0 to day 5, after which it stabilized until the end of 319
the experiment (9.1). Wine added with newly fermented yeast lees (LN) had a similar evolution to T wine, 320
finishing with the highest mDP value (12.5). 321
Figure 5 represents the quantity of proanthocyanidins per mDP interval and by experiment. In each day of 322
sampling, the wines were fractioned regarding their proanthocyanidins and the respective mean degree of 323
polymerization. The mDP values obtained for each of the eight purified fractions varied between 3.6 and 324
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S

American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
15
33.0 (data not shown), resulting in a very large range of values for this parameter. The results for the eight 325
tannin fractions obtained on the glass powder separation column were grouped in intervals of mDP values, 326
from 3 to 8, 8 to 14, 14 to 20 and 20 to 33 in order to facilitate the results evaluation. 327
Concerning the control wine (T wine), on day 5, the proanthocyanidin distribution was similar to day 0 for 328
all mDP intervals, but on the 26th day, was observed an increase of the percentage of proanthocyanidins in 329
the interval [8-14] and a decrease in the interval [20-33]. On the last day of the experiment, the presence 330
of proanthocyanidins with an mDP ranging [20-33] was no longer observed. Comparing with the control 331
wine (T), in the BE samples the fraction [20-33] disappeared earlier since this fraction was no longer 332
present on the 26th day. It was possible to observe mainly the presence of proanthocyanidins belonging to 333
the fraction [3-8] and [8-14]. Together they represented 92% of the total tannins. On the other hand, wines 334
added with LN yeast lees kept at the end of the experiment mainly proanthocyanidins belonging to the [14-335
20] interval which represented 54% of the total tannins. 336
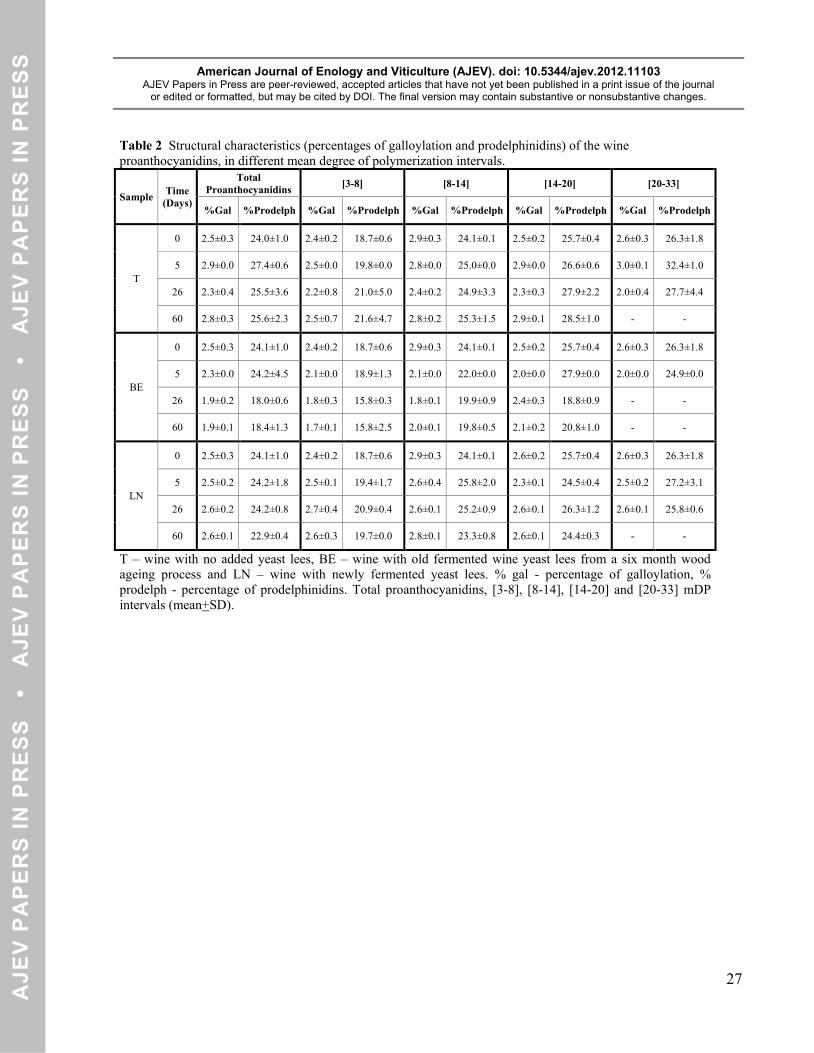
Table 2 shows the evolution of wines proanthocyanidins galloylation and prodelphinidins percentage. 337
Concerning wine with no yeast lees added (T wine) no differences were achieved between the several 338
sampling dates both for galloylation and prodelphinidins percentage. Wine with newly fermented yeast 339
lees added (LN) had the same behavior than T wine. Wine with old fermented yeast lees added (BE) had a 340
decrease in both galloylation and prodelphinidins percentage between day 0 and day 60. It was possible to 341
observe that proanthocyanidins belonging to [3-8] mDP interval had a decrease of 0.7% in the galloylation 342
percentage between day 0 and day 60 and proanthocyanidins belonging to [8-14] mDP interval had a 343
decrease of 0.9% in the same period. Regarding to the evolution of the prodelphinidins percentage from 344
the beginning until the end of the experiment, proanthocyanidins belonging to [8-14] mDP interval had a 345
decrease of 4.3% and proanthocyanidins belonging to [14-20] mDP interval had a decrease of 4.9%. 346
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S

American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
16
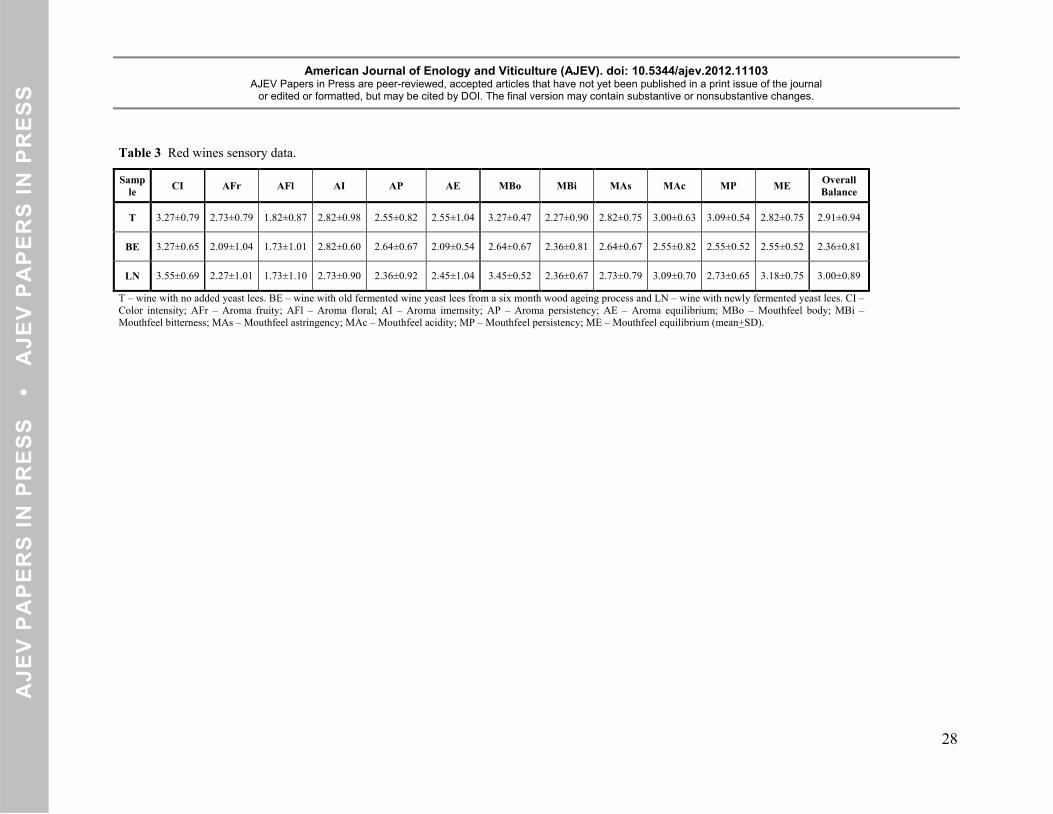
Sensory analysis. Table 3 shows the sensory analysis data obtained for the three wines. In what concerns 347
to the color intensity and aroma attributes, no significant (p<0.05) differences were found among the three 348
wines. In mouthfeel evaluation, some significant differences were obtained between the three wines. The 349
wine body was significantly lower (p<0.05) in BE (2.64) wine than in T (3.27) and LN (3.45) wines, with 350
no significant difference (p<0.05) between the last two. The BE wine showed also a significant lower 351
(p<0.05) persistence than the control wine and a lower (p<0.05) mouthfeel equilibrium than the LN wine. 352
Although there were no significant differences (p<0.05) among the three wines for the overall balance, the 353
LN yeast lees added wine was considered by the tasting panel as the favorite wine with a classification of 354
3.00. The other two wines were classified with 2.91 for the no yeast lees added wine and with 2.36 for the 355
BE wine. 356
Discussion 357
Effect of white wine yeast lees addition on total polyphenols index and color compounds evolution. 358
The T wine presented a higher total polyphenol index (I280) than the other two wines. Probably this is due 359
to some initial adsorption of polyphenols to lees. This adsorption was more evident in wine added with BE 360
yeast lees. This difference was probably due to the fact that the yeast lees quantity added in LN wine was 361
much smaller than in BE, as the concentration of LN yeast lees added was 0.8% (w/v) and BE was 5% 362
(w/v). Guadalupe et al. (2007) found that wines added with external mannoproteins had a higher decrease 363
in the total polyphenol index due to a possible precipitation of polyphenol-mannoproteins aggregates. 364
Salmon et al. (2002) verified that yeast lees are highly reactive with wine polyphenols if the lees were 365
cultivated in the absence of polyphenols. As the yeast lees used on this experiment were produced in a 366
white wine fermentation, the polyphenols interacted immediately with the added yeast lees, resulting in a 367
decrease of the wine polyphenol content. 368
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S

American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
17
Also the color intensity (CI) had an initial quick decrease at the beginning of the experiment for BE and 369
LN wines. CI is mainly due to colored anthocyanins and polymeric pigments content. Vasserot et al. 370
(1997) verified that the quantity of adsorbed anthocyanins linearly increases with their initial 371
concentration, with no saturation on the adsorption process. In the same way, if the quantity of added yeast 372
lees is higher, there will be a higher anthocyanin content decrease. Mazauric and Salmon (2005) have 373
shown that polyphenol adsorption by yeast lees occurs in two-step kinetics, starting with a fast polyphenol 374
fixing followed by a saturation fixing slow and constant which is in accordance with the results obtained in 375
this work. This decrease also occurs in the polymeric pigments (PP) initially. 376
Concerning the hue evolution, the fact that BE had a higher value at the end of the experiment that the 377
other trials could also be due to the highest yeast lees quantity added, resulting in the preferential 378
adsorption of the blue anthocyanins and maintaining more yellow compounds in the wine medium (Morata 379
et al. 2003; Mazauric and Salmon 2006). 380
The results obtained for the color parameters evolution showed that the addition of external yeast lees to 381
this red wine did not influence color stabilization during the studied period of time, as already reported in 382
other works (Guadalupe et al. 2007; Guadalupe and Ayestarán 2008). The only major observation was the 383
interaction between yeast lees and color compounds, resulting in a decrease of colored anthocyanins as 384
already reported by other authors (Vasserot et al. 1997; Salmon et al. 2002; Morata et al. 2003; Mazauric 385
and Salmon 2005, 2006). The color stability is mainly due to the presence of polymeric pigments, 386
molecules resulting from the reaction between anthocyanins and tannins. The phenomenons that enhance 387
the formation of polymeric pigments or that avoid its precipitation result in a stabilization of color. 388
Regarding the wine color during the studied period of time, it was not yet observed the precipitation of 389
polymeric pigments in any modality. On the other hand, it was not expected that the presence of 390
polysaccharides could enhance the formation of polymeric pigments. 391
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S

American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
18
Effect of white wine yeast lees on proanthocyanidins evolution. Comparing the evolution of the total 392
proanthocyanidins content, it was possible to observe an initial decrease between day 0 and day 5 in all 393
wines. This decrease is more noticeable in wine added with yeast lees. This fact can be due to the 394
adsorption of condensed tannins by the lees, as already referred by other authors (Mazauric and Salmon 395
2005; Fernández et al. 2011). After the 5th day the proanthocyanidin content decrease was faster in the T 396
wine, although it ends up with higher proanthocyanidin content. If the decrease rate observed in the first 397
three months of this experiment was maintained along the aging time, then the addition of yeast lees 398
stabilize the decrease of proanthocyanidins. Riou et al. (2002) have already observed that some 399
mannoproteins can influence the aggregation and growing of tannin particles. 400
The proanthocyanidin content distribution profile evolution for the T wine showed that the more 401
polymerized proanthocyanidins precipitated along the time. The inexistence of proanthocyanidins with an 402
mDP between 20 and 33 at the 60th day was probably due to: a) the polymerization of flavan-3-ol units 403
results in the formation of high molecular weight polymers that precipitate; b) the association between 404
tannins and anthocyanins as T-A+ complexes promotes the stability of the molecule without any further 405
growth (Vidal et al. 2002); c) as tannin tends to aggregate through time and further to precipitate, leads to 406
a reduction of the number of flavan-3-ol units existing on solution, limiting the reaction between 407
monomers and monomers with oligomers in order to form higher chain polymers. 408
Comparing the evolution of BE wine proanthocyanidins with the one observed in T wine, it was possible 409
to say that the addition of old yeast lees promoted an initial adsorption of the highly polymerized 410
proanthocyanidins, followed by the maintenance of the low and medium size proanthocyanidins in the 411
wine medium. This fact confirmed that the yeast lees have a stronger interaction with highly polymerized 412
condensed tannins as already reported by Mazauric and Salmon (2006) although the highest mDP found 413
by these authors was 16. Guadalupe and Ayestarán (2008) also verified that in a wine treated with 414
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S

American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
19
commercial mannoproteins there was a decrease of high molecular weight mannoproteins at the same time 415
that proanthocyanidins decreased, suggesting that high molecular weight mannoproteins aggregate to 416
proanthocyanidins and that this co-aggregates precipitate or flocculate. On the other hand, the low and 417
medium size proanthocyanidins maintenance in the wine medium can be attributed to the low-medium 418
mannoproteins that were extracted from the yeast lees during the experiment period. Riou et al. (2002) 419
verified that in a wine model solution, polysaccharides, namely mannoproteins, interfere on the particle 420
size of proanthocyanidins, although they do not protect the tannins initial aggregation. 421
On LN wine proanthocyanidins evolution, the fact that the proanthocyanidins with an mDP between 14 422
and 20 were the ones existing in a higher percentage in the last day of the trial could be due to a retarding 423
of higher molecular weight proanthocyanidins polymerization promoted by mannoproteins released by the 424
added yeast lees. 425
Concerning the evolution of wines proanthocyanidins galloylation and prodelphinidins percentage, the 426
decrease of prodelphinidins percentage is in accordance with the results shown by Mazauric and Salmon 427
(2005) who reported the condensed tannins that remain in the wine have less epigallocatechin units than 428
the initial tannins, indicating that the more polar condensed tannins are preferentially adsorbed by yeast 429
lees. The decrease of galloylation percentage was already reported by Mazauric and Salmon (2006) as they 430
showed that the condensed tannins adsorbed by yeast lees have a high galloylation percentage when 431
compared with the wine initial condensed tannins. It seemed like proanthocyanidins with a higher number 432
of hydroxyl groups tend to disappear faster from the BE wine medium. Cheynier et al. (1992) showed that 433
the more galloylated proanthocyanidins were faster involved in condensation reactions than the non-434
galloylated compounds. 435
Comparing the evolution of the total proanthocyanidin content and the evolution of the different 436
proanthocyanidins mDP intervals content and galloylation and prodelphinidins percentages, it was possible 437
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S

American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
20
to conclude that there were two different mechanisms acting as a result of the external yeast lees addition: 438
a) the first one was observable in the transition from day 0 to day 5, where the external lees addition 439
promoted a quick adsorption of the proanthocyanidins, namely the ones that were more polymerized as it 440
was possible to see mainly in the results obtained for the BE wine; b) after this fast adsorption, it seemed 441
that there was a retarding of the proanthocyanidins polymerization to a higher degree, namely maintaining 442
a relatively high quantity of proanthocyanidins with mDP [8-14] for the BE sample and in the [14-20] 443
interval for the LN sample, probably due to the released mannoproteins by yeast lees. It is important to 444
notice that the first effect, related to the yeast lees adsorption, was more evident on the BE wine, and it 445
was the result of adding a higher quantity of yeast lees to this modality than to the LN wine. The second 446
effect, related to the retarding of further polymerization due to the presence of mannoproteins was more 447
evident on the LN wine. As the LN wine yeast lees were fresh, removed at the very end of alcoholic 448
fermentation, they possibly had more effective mannoproteins available for the stabilization phenomenon 449
than the ones coming from BE wine yeast lees. It should be also noticed that the protein quantity in LN FR 450
fraction is higher than in the BE FR fraction. LN FR peak 1 contained 74.1% of proteins and peak 2 451
contained 18.2%. In BE FR the same peaks contained respectively 14.8% and 7.3%. Possibly the protein 452
composition of these macromolecules may have influence in their interaction with proanthocyanidins. 453
Poncet-Legrand et al. (2007) verified that low-medium mannoproteins (51 and 62 kDa) prevented 454
polyphenol precipitation by preventing tannin aggregation. Both used yeast lees presented a mannoprotein 455
with an average molecular weight of 54 and 50 kDa, respectively, however this mannoprotein was more 456
perceptible in the newly fermented yeast lees polysaccharide profile. The tannin polymerization retarding 457
effect could have been promoted by the interaction of these particular mannoprotein with the 458
proanthocyanidins in the wine medium. 459
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S

American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
21
Sensory analysis. Wine added with newly fermented yeast lees (LN) was considered by the tasting panel 460
as the favorite wine. Wine added with old fermented yeast lees (BE) had a less wine body, persistence and 461
mouthfeel equilibrium. These results were in accordance with results obtained for the proanthocyanidins 462
content evolution. The added yeast lees took a great quantity of condensed tannins from the medium 463
resulting in a wine with lower mouthfeel equilibrium. This way, even if the lees addition contributed to an 464
enrichment of wine mannoproteins, they could not compensate, regarding the sensory characteristics, the 465
high decrease in wine tannins. Probably in BE wine the quantity of added yeast lees was enough to remove 466
some tannins that could contribute to a more aggressive wine, softening the mouthfeel. Possibly the 467
mannoproteins that have been extracted to this wine contributed also to a better mouthfeel. 468
469
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S

American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
22
Conclusions 470
The external white wine yeast lees addition did not have an effect on wine color stabilization. The only 471
remarkable effect was the decrease of anthocyanins and polymeric pigments content, specially the colored 472
anthocyanins, immediately after the yeast lees addition. This decrease was due to the adsorptive 473
interactions that were established between anthocyanins and yeast lees. The same kind of effect was 474
observed between proanthocyanidins and yeast lees at the beginning of the experiment. During the time, 475
after the initial decay, it seemed like there was a retarding effect on the proanthocyanidins polymerization 476
reactions by the addition of external yeast lees. This effect could occur due to two different factors: a) the 477
fast initial removal of the more polar proanthocyanidins by adsorption to yeast lees and b) the 478
proanthocyanidins polymerization retarding effect promoted by the low-medium molecular weight 479
mannoproteins extracted from the added yeast lees. Regarding the proanthocyanidins structure, the 480
addition of old fermented yeast lees obtained from a six month aging process (BE wine) resulted in the 481
prevalence of less polar tannins in wine and an adsorption of the highest molecular weight 482
proanthocyanidins. The fact that BE yeast lees added wine was considered by the tasting panel as the less 483
equilibrated in the mouthfeel parameters could be due to the addition in excess of the external white wine 484
lees. On the other hand, the wine added with newly fermented yeast lees (LN) had, at the end of the 485
experiment, a higher percentage of proanthocyanidins with [14-20] mDP, and, possibly due to this fact, a 486
better mouthfeel, as reported by the tasting panel. These results showed that the addition of external yeast 487
lees in the winery has to be carefully planned and both the working concentration and the yeast lees age 488
should be tested in a small volume before adding the yeast lees to the wine in the industrial process. 489
490
491
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S

American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
23
Literature Cited 492
Albersheim, P., D.J. Nevis, P.D. English, and A. Karr. 1967. A method for the analysis of sugars in plant 493 cell-wall polysaccharides by gas liquid chromatography. Carbohydr. Res. 5:340-345. 494
Cheynier, V., J. Rigaud, and J.M. Ricardo da Silva. 1992. Structure of procyanidin oligomers isolated from 495 grape seeds in relation to some of their chemical properties. In Plant Polyphenols. R.W. Hemingway and 496 P.E. Laks (eds.), pp. 281-294. Plenum Press, New York, USA. 497
Dallas, C., J.M. Ricardo da Silva, and O. Laureano. 1996. Products formed in model wine solutions 498 involving anthocyanins, procyanidin B2, and acetaldehyde. J. Agric. Food Chem. 44:2402-2407. 499
Dubois, M., K. A. Gilles, J. K. Hamilton, P. A. Rebers, and S. Smith. 1956. Colorimetric method for 500 determination of sugars and related substances. Anal. Chem. 28:350-356. 501
Escot, S., M. Feuillat, L. Dulau, and C. Charpentier. 2001. Release of polysaccharides by yeasts and the 502 influence of released polysaccharides on color stability and wine astringency. Austr. J. Grape Wine Res. 503 7:153-159. 504
Fernández, O., O. Martínez, Z. Hernández, Z. Guadalupe, and B. Ayestarán. 2011. Effect of the presence 505 of lysated lees on polysaccharides, color and main phenolic compounds of red wine during barrel ageing. 506 Food Res. Int. 44:84-91. 507
Feuillat, M., and C. Charpentier. 1982. Autolysis of yeasts in champagne. Am. J. Enol. Vitic. 33:6-13. 508
Feuillat, M., M. Freyssinet, and C. Charpentier. 1989. L’élevage sur lies des vins blancs de Bourgogne. II. 509 Evolution des macromolécules: polysaccharides et protéines. Vitis 28:161-176. 510
Fornairon-Bonnefond, C., C. Camarasa, M. Moutounet, and J.M. Salmon. 2002. New trends on yeast 511 autolysis and wine ageing on lees: a bibliographic review. J. Int. Sci. Vigne Vin 36:49-69. 512
Fulcrand, H., S. Remy, J.-M. Souquet, V. Cheynier, and M. Moutounet. 1999. Study of wine tannin 513 oligomers by on-line liquid chromatography electrospray ionization mass spectrometry. J. Agric. Food 514 Chem. 47:1023–1028. 515
Gonçalves, F., A. Heyraud, M.N. Pinho, and M. Rinaudo. 2002. Characterization of white wine 516 mannoproteins. J. Agric. Food Chem. 50:6097-6101. 517
Guadalupe, Z., A. Palacios, and B. Ayestarán. 2007. Maceration enzymes and mannoproteins: a possible 518 strategy to increase colloidal stability and color extraction in red wines. J. Agric. Food Chem. 55:4854-519 4862. 520
Guadalupe, Z. and B. Ayestarán. 2008. Effect of commercial mannoprotein addition on polysaccharide, 521 polyphenolic, and color composition in red wines. J. Agric. Food Chem. 56:9022–9029. 522
Haslam, E. (1980). In vino veritas: Oligomeric procyanidins and the ageing of red wines. Phytochem. 523 19:2577-2582. 524
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S

American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
24
Hernawan, T. and G. Fleet. 1995. Chemical and cytological changes during the autolysis of yeasts. J. Ind. 525 Microbiol. Biotechnol. 14:440-450. 526
Labarbe, B., V. Cheynier, F. Brossaud, J.M. Souquet, M. Moutounet. 1999. Quantitative fractionation of 527 grape proanthocyanidins according to their degree of polymerization. J. Agric. Food Chem. 47:2719-2723. 528
Lowry, O. H., N. J. Roserbrough, A. L. Farr, and R. J. Randall. 1951. Protein measurement with the Folin 529 phenol reagent. J. Biol. Chem. 193:265-275. 530
Mazauric, J.P. and J.M. Salmon. 2005. Interactions between yeast lees and wine polyphenols during 531 simulation of wine aging: I. Analysis of remnant polyphenolic compounds in the resulting wines. J. Agric. 532 Food Chem. 53:5647–5653. 533
Mazauric, J.P. and J.M. Salmon. 2006. Interactions between yeast lees and wine polyphenols during 534 simulation of wine aging. II. Analysis of desorbed polyphenol compounds from yeast lees. J. Agric. Food 535 Chem. 54:3876–3881. 536
Medina, K., E. Boido, E. Dellacassa, and F. Carrau. 2005. Yeast interactions with anthocyanins during red 537 wine fermentation. Am. J. Enol. Vitic. 56 :104-109. 538
Moine-Ledoux, V., A. Perrin, I. Paladin, and D. Dubourdieu. 1997. Premiers résultats de stabilisation 539 tartrique des vins par addition de mannoprotéines purifiées (Mannostab). J. Int. Sci. Vigne Vin 31:23-31. 540
Monagas, M., C. Gómez-Cordovés, B. Bartolomé, O. Laureano, and J. M. Ricardo-da-Silva, 2003. 541 Monomeric, oligomeric, and polymeric flavan-3-ol composition of wines and grapes from Vitis vinifera L. 542 Cv. Graciano, Tempranillo, and Cabernet Sauvignon. J. Agric. Food Chem. 51:6475–6481. 543
Morata, A., M.C. Gómez-Cordovés, J. Suberviola, B. Bartolomé, B. Colomo, and J.A. Suárez. 2003. 544 Adsorption of Anthocyanins by Yeast Cell Walls during the Fermentation of Red Wines. J. Agric. Food 545 Chem. 51:4084-4088. 546
OIV. 2006. Recueil des Méthodes Internationales d’Analyse des Vins et Moûts. Organisation International 547 de la Vigne et du Vin, Paris. 548
Poncet-Legrand, C., T. Doco, P. Williams, and A. Vernhet. 2007. Inhibition of grape seed tannin 549 aggregation by wine mannoproteins: effect of polysaccharide molecular weight. Am. J. Enol. Vitic. 58:87-550 91. 551
Ribéreau-Gayon, P., D. Dubourdieu, B. Donéche, and A. Lonvaud. 1998. La développement des bactéries 552 lactiques dans le vin. In Traité d’Œnologie – 1. Microbiologie du Vin – Vinifications, pp. 197-223. Dunod, 553 Paris, France. 554
Ricardo da Silva, J.M., J.P. Rosec, M. Bourzeix, and N. Heredia. 1990. Separation and quantitative 555 determination of grape and wine procyanidins by high performance reversed phase liquid chromatography. 556 J. Sci. Food Agric. 53:85-92. 557
Rigaud, J., J. Perez-Ilzarbe, J. M. Ricardo da Silva, and V. Cheynier. 1991. Micro method for the 558 identification of proanthocyanidin using thiolysis monitored by high-performance liquid chromatography. 559 J. Chromatogr. 540:401–405. 560
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S

American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
25
Riou, V., A. Vernhet, T. Doco, and M. Moutounet. 2002. Aggregation of grape seed tannins in model wine 561 – effect of wine polysaccharides. Food Hydrocoll. 16:17-23. 562
Salmon, J.M., C. Fornairon-Bonnefond, and J.P. Mazauric. 2002. Interactions between wine lees and 563 polyphenols: influence on oxygen consumption capacity during simulation of wine aging. J. Food Sci. 564 67:1604-1609. 565
Somers, T.C. 1971. The polymeric nature of wine pigments. Phytochem. 10:2175-2186. 566
Somers, T. C., and M. E. Evans. 1977. Spectral evaluation of young red wines: anthocyanin equilibria, 567 total phenolics, free and molecular SO2, "chemical age". J. Sci. Food Agric. 28:279-287. 568
Sun, B., G.P. Belchior, J.M. Ricardo da Silva, and M.I. Spranger. 1999. Isolation and purification of 569 dimeric and trimeric procyanidins from grape seeds. J. Chromatogr. A. 841:115-121. 570
Vasserot Y., S. Caillet and A. Maujean. 1997. Study of anthocyanin adsorption by yeast lees. Effect of 571 some physicochemical parameters. Am. J. Enol. Vitic. 48:433 –437. 572
Vidal, S., D. Cartalade, J.-M. Souquet, H. Fulcrand, and V. Cheynier. 2002. Changes in proanthocyanidin 573 chain length in winelike model solutions. J. Agric. Food Chem. 50:2261-2266. 574
575
576
577
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S

American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
26
Table 1 White wine yeast lees – old fermented wine yeast lees from a six month wood ageing process (BE) and newly fermented yeast lees (LN) – characterization of the concanavalin-A non retained (FNR) and retained fractions (FR) for its molecular weight (MW), polysaccharides (Polysacc. %), proteins (%) and residual sugars (%) (fucose (Fuc), rhamnose (Rha), arabinose (Ara), xylose (Xyl), mannose (Man), glucose (Glc) and galactose (Gal)), n.d. represents non detected.
Sample Total
colloids a
Distribution in total
colloidsb
MW (kDa)c
Polysacc. (%)c
Proteins (%)c
Fuc (%)c
Rha (%)c
Ara (%)c
Xyl (%)c
Man (%)c
Glc (%)c
Gal (%)c
BE
F NR
Peak 1
1.2%
21%
177±17 71.1±0.2 28.9±0.2 n.d. n.d. 9.0±0.0 n.d. n.d. n.d. 91.0±0.0
Peak 2 23±1 85.5±0.3 14.5±0.3 0.7±0.9 1.8±2.5 9.2±2.0 1.4±2.0 n.d. n.d. 86.9±1.5
Peak 3 3±0 67.0±0.0 32.2±0.0 2.7±0.0 8.9±0.0 18.3±0.0 15.1±0.0 n.d. n.d. 54.9±0.0
F R
Peak 1
56%
151±11 85.2±0.3 14.8±0.3 n.d. n.d. 0.1±0.1 0.4±0.2 52.2±6.4 47.1±6.4 0.2±0.3
Peak 2 54±4 92.7±0.3 7.3±0.3 n.d. n.d. 0.4±0.2 0.1±0.1 95.3±0.9 3.8±1.3 0.5±0.1
Peak 3 23±1 94.0±0.5 6.0±0.5 n.d. n.d. 0.4±0.0 n.d. 95.9±0.3 3.7±0.3 n.d.
LN
F NR
Peak 1
1.3%
13%
159±2 80.1±2.3 19.9±2.3 0.9±1.2 0.0±0.0 14.5±4.2 0.4±0.5 n.d. n.d. 84.3±2.4
Peak 2 23±1 21.3±1.8 78.7±1.8 3.3±0.0 8.1±3.0 23.1±0.0 n.d. n.d. n.d. 63.3±0.0
Peak 3 2±0 46.3±3.5 53.7±3.5 n.d. n.d. 40.5±0.0 n.d. n.d. n.d. 59.5±0.0
F R
Peak 1
65%
154±11 25.9±2.6 74.1±2.6 n.d. 0.2±0.0 0.2±0.0 n.d. 32.9±0.3 60.4±1.3 6.5±2.0
Peak 2 50±3 81.8±0.3 18.2±0.3 n.d. 0.1±0.1 0.1±0.0 n.d. 88.6±5.8 8.9±4.3 2.2±1.5
Peak 3 25±1 92.6±0.3 7.4±0.3 n.d. n.d. 0.4±0.1 0.1±0.0 67.1±7.0 29.5±6.7 2.8±0.4
a – extracted from yeast lees after digestion with β-glucanase in aqueous medium; b – after Concanavalin-A affinity chromatography; c – after FPLC size exclusion chromatography.
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
27
Table 2 Structural characteristics (percentages of galloylation and prodelphinidins) of the wine proanthocyanidins, in different mean degree of polymerization intervals.
Sample Time (Days)
Total Proanthocyanidins [3-8] [8-14] [14-20] [20-33]
%Gal %Prodelph %Gal %Prodelph %Gal %Prodelph %Gal %Prodelph %Gal %Prodelph
T
0 2.5±0.3 24.0±1.0 2.4±0.2 18.7±0.6 2.9±0.3 24.1±0.1 2.5±0.2 25.7±0.4 2.6±0.3 26.3±1.8
5 2.9±0.0 27.4±0.6 2.5±0.0 19.8±0.0 2.8±0.0 25.0±0.0 2.9±0.0 26.6±0.6 3.0±0.1 32.4±1.0
26 2.3±0.4 25.5±3.6 2.2±0.8 21.0±5.0 2.4±0.2 24.9±3.3 2.3±0.3 27.9±2.2 2.0±0.4 27.7±4.4
60 2.8±0.3 25.6±2.3 2.5±0.7 21.6±4.7 2.8±0.2 25.3±1.5 2.9±0.1 28.5±1.0 - -
BE
0 2.5±0.3 24.1±1.0 2.4±0.2 18.7±0.6 2.9±0.3 24.1±0.1 2.5±0.2 25.7±0.4 2.6±0.3 26.3±1.8
5 2.3±0.0 24.2±4.5 2.1±0.0 18.9±1.3 2.1±0.0 22.0±0.0 2.0±0.0 27.9±0.0 2.0±0.0 24.9±0.0
26 1.9±0.2 18.0±0.6 1.8±0.3 15.8±0.3 1.8±0.1 19.9±0.9 2.4±0.3 18.8±0.9 - -
60 1.9±0.1 18.4±1.3 1.7±0.1 15.8±2.5 2.0±0.1 19.8±0.5 2.1±0.2 20.8±1.0 - -
LN
0 2.5±0.3 24.1±1.0 2.4±0.2 18.7±0.6 2.9±0.3 24.1±0.1 2.6±0.2 25.7±0.4 2.6±0.3 26.3±1.8
5 2.5±0.2 24.2±1.8 2.5±0.1 19.4±1.7 2.6±0.4 25.8±2.0 2.3±0.1 24.5±0.4 2.5±0.2 27.2±3.1
26 2.6±0.2 24.2±0.8 2.7±0.4 20.9±0.4 2.6±0.1 25.2±0.9 2.6±0.1 26.3±1.2 2.6±0.1 25.8±0.6
60 2.6±0.1 22.9±0.4 2.6±0.3 19.7±0.0 2.8±0.1 23.3±0.8 2.6±0.1 24.4±0.3 - -
T – wine with no added yeast lees, BE – wine with old fermented wine yeast lees from a six month wood ageing process and LN – wine with newly fermented yeast lees. % gal - percentage of galloylation, % prodelph - percentage of prodelphinidins. Total proanthocyanidins, [3-8], [8-14], [14-20] and [20-33] mDP intervals (mean+SD).
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
28
Table 3 Red wines sensory data.
Sample CI AFr AFl AI AP AE MBo MBi MAs MAc MP ME Overall
Balance
T 3.27±0.79 2.73±0.79 1.82±0.87 2.82±0.98 2.55±0.82 2.55±1.04 3.27±0.47 2.27±0.90 2.82±0.75 3.00±0.63 3.09±0.54 2.82±0.75 2.91±0.94
BE 3.27±0.65 2.09±1.04 1.73±1.01 2.82±0.60 2.64±0.67 2.09±0.54 2.64±0.67 2.36±0.81 2.64±0.67 2.55±0.82 2.55±0.52 2.55±0.52 2.36±0.81
LN 3.55±0.69 2.27±1.01 1.73±1.10 2.73±0.90 2.36±0.92 2.45±1.04 3.45±0.52 2.36±0.67 2.73±0.79 3.09±0.70 2.73±0.65 3.18±0.75 3.00±0.89
T – wine with no added yeast lees. BE – wine with old fermented wine yeast lees from a six month wood ageing process and LN – wine with newly fermented yeast lees. CI – Color intensity; AFr – Aroma fruity; AFl – Aroma floral; AI – Aroma imemsity; AP – Aroma persistency; AE – Aroma equilibrium; MBo – Mouthfeel body; MBi – Mouthfeel bitterness; MAs – Mouthfeel astringency; MAc – Mouthfeel acidity; MP – Mouthfeel persistency; ME – Mouthfeel equilibrium (mean+SD).
A
JE
V P
AP
ER
S I
N P
RE
SS
•
A
JE
V P
AP
ER
S I
N P
RE
SS

American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
29
Figure 1 Schematic representation of the yeast lees characterization procedure – winery resulting lees (BE) and newly fermented lees (LN).
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
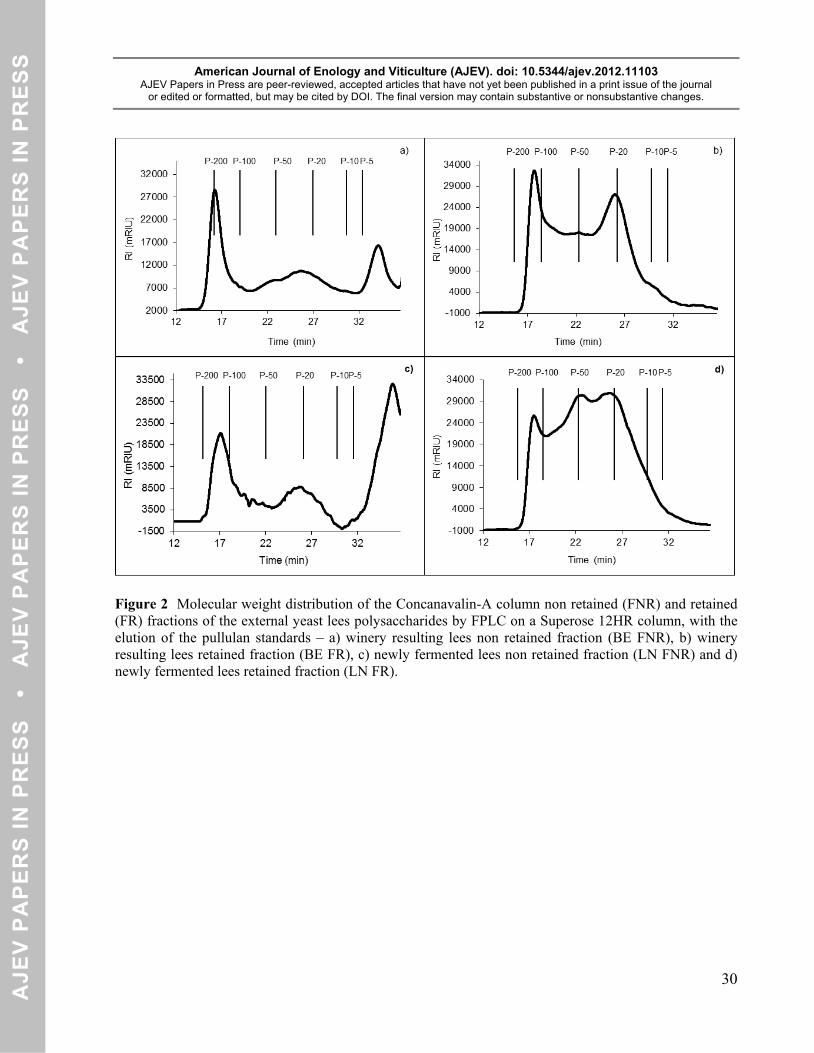
American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
30
Figure 2 Molecular weight distribution of the Concanavalin-A column non retained (FNR) and retained (FR) fractions of the external yeast lees polysaccharides by FPLC on a Superose 12HR column, with the elution of the pullulan standards – a) winery resulting lees non retained fraction (BE FNR), b) winery resulting lees retained fraction (BE FR), c) newly fermented lees non retained fraction (LN FNR) and d) newly fermented lees retained fraction (LN FR).
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S

American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
31
Figure 3 a) Polyphenol index (I280), b) color intensity (CI), c) hue, d) polymeric pigments (PP), e) total anthocyanins (TA), f) colored anthocyanins (CA), g) total pigments (TP) evolutions on wines with no added lees (T), winery resulting lees (BE) and newly fermented lees (LN).
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S

American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
32
Figure 4 a) global proanthocyanidins content (PA) and b) global mDP evolutions on wines with no added lees (T), from a six months wood ageing process (BE) and newly fermented lees (LN).
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S

American Journal of Enology and Viticulture (AJEV). doi: 10.5344/ajev.2012.11103 AJEV Papers in Press are peer-reviewed, accepted articles that have not yet been published in a print issue of the journal
or edited or formatted, but may be cited by DOI. The final version may contain substantive or nonsubstantive changes.
33
Figure 5 Evolution of the quantity of proanthocyanidins per mDP interval and for each modality (T – with no added lees wine, BE – winery resulting lees from a six months wood ageing process added wine, 8 LN – newly fermented lees added wine), (a) day 0, (b) day 5, (c) day 26 and (d) day 60. The relative percentages of the proanthocyanidins quantities per mDP interval for each sample are also shown. 0
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S
•
AJ
EV
PA
PE
RS
IN
PR
ES
S





























