Embed Size (px)
Citation preview

Effect of Forest Fragmentation on Genetic Diversity and Mating System in a Tropical Tree, Pithecellobium elegans PAMELA HALL,* SARAH WALKER, t AND KAMALJIT BAWA:~g *Aarhus University Herbarium, University of Aarhus, Universitetsparken, Building 137, DK-8000 Aarhus C, Denmark t Smith College, Northampton, MA 01063, U.S.A.
Department of Biology, University of Massachusetts, Harbor Campus, Boston, MA 02125, U.S.A.
Abstract: Genetic diversity, population differentiation, and temporal variation in outcrosstng rates were ex- amined for Pithecellobium elegans, a Neotroptcal rain forest canopy tree. Several forest fragments and a large reserve (1500 ha) were compared for several population genetic parameters. For eight populations sampled on the Atlantic coastal plain o f Costa Rica, allozyme heterozygostty (0.13), polymorphism (35%), and effective number o f alleles (1.24) were similar to values reported fo r other tropical tree species that occur at similar densities o f less than one individual per hectare. These measures o f genetic variation were lowest in popula- tions o f the smallest size, farthest f r o m the reserve, and more isolated f rom other populations. Differentiation among samples collected in small forest fragments and the reserve population accounted fo r 10% o f the total genetic variation observed. There was a positive relationship between the level o f differentiation o f popula- tions f r o m the reserve population and their distance f r o m the reserve. Though predominantly an annually f lowering species, the number o f trees in f lower at any one time varied f r o m 80% o f observed trees to only 6%. Outcrosstng rates did not differ for two episodes in which the proportions o f f lowering trees were 33% and 80%. But periods o f low density o f f lowering adults resulted in poor seed crops or failure to set f ru i t for many individuals. Population size at many sites will he effectively decreased because o f the variation in flowering. Fragmentation o f what was once a large, continuous forested area is resulting in genetic erosion o f small, lso- latedpopulattons ofPithecellobium elegans.
Efectos de la Fragmentaci6n del Bosque sobre la Diversidad Gendtica y el Sistema de Cruza en un Arbol Tropical
Resumen: Examtnamos la dtversidad gen•tica, la diferenciact6n de la poblaci6n y la variaci6n temporal de Pithecellobium elegans, un drbol de dosel de bosque lluvtoso en el neotr6pico. Poblactones de diversos frag- mentos de bosque, ast como de una reserva grande (1,500 ha) fueron comparadas usando dtversos pardmetros de gednttca poblacionaL En ocho poblactones muestreadas de la plantcle costera del Atldntico de Costa Rica, los valores de heterocigocidad de alosomas (0.13), polimorftsmo (35%) y namero efectivo de ale- los (1.24) fueron stmtlares a los reportados para otras especies de drboles que ocurren a densidades simtlares de menos de un indivtduo por hectdrea. Estas medtciones de variact6n gendtica fueron menores en las pobla- ctones de menor tama~o, alejadas de la reserva y las mds atsladas de otras poblaciones. La dtferenctaci6n en- tre muestras colectadas en fragmentos peque~os de bosque y la poblaci6n de la reserva explica 10% de la vartaci6n gendtica total observada. Existt6 una relact6n positiva entre el nivel de dtferenctact6n de las pobla- ctones de la reserva y la distancia a la que se encontraban de la reserva. Considerada predomtnantemente como una especte de florecimiento anual, el namero de drboles en f lor vari6 entre 80% de drboles observados hasta solo 6%. Las tasas de cruza ex6gena no fueron dtferentes a dos episodtos en los cuales la proporci6n de drboles floreciendo fue de 33% Y 80%. De cualquier manera, los perfodos de baja denstdad de adultos flore-
fAddress correspondence to K. Bawa. Paper submitted December 22, 1994; revised manuscript accepted June 19, 1995.
757
conservation Biology, Pages 757-768 Volume 10, No. 3,June 1996

758 Genetic l~'versi~ and Mating System in Pithecell0bium elegans Ha//et a/.
ciendo result6 en una producci6n de semtllas pobre, o en carencta de frutos en muchos tndtvtduos. E1 tamaflo poblacional de muchos sitios decrecerd debido a la vartact6n en florecimtentos. La fragmentaci6n de lo que alguna vez fuera una vasta dtrea arbolada de Pithecellobium elegans se estd convtrtiendo en poblaciones pe- queflas gendticamente erostonadas y aisladas.
Introduction
Deforestation, including fragmentation of forested habi- tats, can result in losses of genetic variation within spe- cies by causing extinction of locally adapted populations and by reducing gene flow and effective population sizes (Ledig & Conkle 1983). The erosion of genetic di- versity as a result of deforestation may be more pro- nounced in tree species occurring in low densities in tropical lowland rain forests than for more abundant species. In many such forests more than 50% of the tree species may have adult densities of a single individual or less per hectare (Poore 1968; Lieberman et al. 1990; Hubbell & Foster 1983; Whitmore 1984; Gentry & Ter- borgh 1990). Thus, in isolated reserves of even several hundred hectares, the number of adult individuals may be less than the number required for long-term viability of population (or minimum viable population size).
To assess the genetic consequences of tropical defor- estation, information is needed on patterns of genetic di- versity and temporal variation in mating systems and gene flow (Bawa 1993). Mating systems and gene flow are important determinants of effective population size. The estimates of effective population size in turn are of critical importance in conservation planning.
Although most tropical rain forest trees are strongly outcrossed with extensive gene flow (Ashton 1969; Bawa et al. 1985; Bawa 1992; Hamrick & Murawski 1991; Hamrick et al. 1991; Hall et al. 1994), temporal variation in flowering within and among breeding sea- sons may cause fluctuations in the levels of outcrossing and gene flow (Murawski & Hamrick 1991) and may fur- ther exacerbate the effects of deforestation and forest fragmentation. The cohorts of offspring produced after an episode of flowering with low intensity may be more inbred than cohorts produced after high incidence of flowering. Thus, variation among individuals in timing of reproduction may increase inbreeding and further lower the effective population size in small fragments. Al- though tropical forest trees are known to have varied and complex flowering patterns (Bawa 1983; Newstrom et al. 1991 and references therein), the genetic conse- quences of variation in flowering within populations have remained virtually unexplored.
We examined temporal variation in reproduction and the impact of forest fragmentation on genetic variation and outcrossing rate in Pi theceUobr ium elegans, a large tree that occurs in low densities in the rain forest. Our goals were to estimate (1) genetic diversity in forest frag-
ments of various sizes, (2) the degree of genetic differen- tiation among the fragmented populations and their rela- tionship to a large population, and (3) the amount of variation in outcrossing as a function of the number of flowering individuals within a population of a large na- ture reserve.
Methods
Species Description
Pi thece l lob ium elegans (Leguminosae Mimosoideae) is a large, tropical tree, often reaching over 1 m in diame- ter and 40 m in height, with its crown emerging above the main forest canopy. This species is found at lower el- evations, usually not exceeding 400 m, from the Atlantic side of Central America and south into Amazonian Brazil. It occurs in average densities of less than one reproduc- tive individual per hectare, but it can also be found in clusters, especially on hillsides and ridge tops where in- dividuals may be separated by less than 50 m (P. Hall, personal observation). P. elegans was probably once found extensively throughout the costal plain of Costa Rica, but its distribution has been restricted and frag- mented through conversion of forests to pasture, planta- tions, and towns. As with many tropical tree species, its density and population distribution throughout its range is not well known.
In Costa Rica flowering occurs annually from Decem- ber to February, and a secondary flowering period of less intensity can also occur from May to June. But not all individuals in a population exhibit the same pheno- logical patterns. Some individuals may flower twice a year, especially if a large seed crop is not produced from the first flowering episode. Other individuals may flower at 18-month intervals, apparently alternating between the primary and secondary flowering periods. The num- ber of reproductive individuals and synchrony of repro- duction vary from year to year. Within six months of flowering, fruits containing generally between 8 and 16 seeds are produced. Fruits can be held in the tree crown for many months, and when they fall to the ground heavy predation is evident often with a loss of over 80% of the seeds in a single fruit (P. Hall, personal observa- tion). Dispersal appears to be effected by various birds feeding in the canopy and scattering seeds as they feed on them. Seedlings are rarely found in the forest except in the immediate vicinity of mother trees. P. elegans ap-
Conservation Biology Volume 10, No. 3,June 1996

Hall et d. Genetic Diversity and Mating System in Pithecell0bium degans 759
Nicaragua
Costa Rica
RIoBiJagual ~l~La ! Brmulio Carrillo
San Jo~
Cadbbean Sea
Reslfve
Pacific Ocean
8 .6 w 8,s e,4 .p
Figure 1. Location o f eight popula t ions of Pithecello- bium elegans sampled in this study. The popula t ions La Selva a n d Braulio Carrillo are located wi th in a cont iguous natural fores t reserve a n d nat ional park , respectively.
11"
pears to require a relatively high amount of light to es- tablish successfully and grow (Clark & Clark 1992).
Population Differentiation
We collected seeds and leaf material from eight popula- tion of P. elegans located throughout the Atlantic coastal plain of Costa Pica (Fig. 1). The fragmentation and isolation of these populations has been occurring for over four decades and at a particularly rapid pace in recent years (Sader & Joyce 1988). We collected mate- rial in July 1992, during a period when fruits are likely to be found in all populations. For each population we col- lected seeds from the ground or leaf material from seed- lings of various heights when there were insufficient numbers of seeds to represent a population. All the seed- lings collected were small, probably the product of re- productive episodes within the last 2-3 years, well after population fragmentation and isolation had occurred. In one population, La Selva, both adult leaves and seeds were collected, and the material was labeled appropri- ately so that its source could be determined. The num- ber of adult leaf samples collected, the number of adults present in the population, and the total number of sam- ples collected are given in Table 1.
The sampled populations of P. elegans varied in adult and juvenile density and degree of isolation. Adult densi- ties, reflecting an average distribution of individuals over an area, were extremely difficult to estimate and may be
of limited use because trees occur singly and in clumps. The adult trees are readily spotted at great distances, however, because of their distinctive crown shape. Therefore, when a clear view of an area can be obtained, the majority of large, potentially reproductive trees can be located. The area covered by such populations is dif- ficult to estimate without surveying many hectares of tropical forest. Hence, population densities were not quantified except for one sample location.
The populations in La Selva Reserve (LS) and Braulio Carrillo National Park (BC) are located within a large re- serve and national park that border each other. These sites probably represent the last large, contiguous area of forest within which P. elegans is found in Costa Pica (R. Peralta, personal communication). The LS popula- tion is located in the center of the primary forest area of the reserve. The average density of P. elegans in the area of study is approximately one individual per hectare (Clark & Clark 1992). Many other individuals occur within the reserve but are outside the sampled popula- tion. The BC population consists of trees that were not felled when parts of the forest were cleared for pastures that have been abandoned for at least six years. This area is now a mixture of secondary forest, with many individ- uals and components of the original undisturbed forest still in place. The Rio Bijagual (RB) population is located on one side of the nature reserve and park, within exist- ing pastures, and it is composed of remnant large trees scattered along hillsides and ridge tops. The average density of both the BC and RB populations, where 10 and 35 adults, respectively, were found, probably does not differ significantly from that of the LS population in terms of the number of adults in the circumscribed sam- ple area. But the presence and density of individuals in the many square kilometers of intervening landscape was not determined.
The populations of Estero Grande (EG) and Finca Abogado (A) consist of a few isolated individuals located in extensive areas of pastures at least 10 km from the large forest tract of La Selva. These populations con- sisted of five and seven adults, respectively, which we believed to be the only individuals in an area of approxi- mately 80 ha each, determined by extensive searching. The Cedral (CD) and Linda Vista (LV) populations are very near each other, but their habitats are different. The LV population grows on the sides and ridges of a se- ries of hills 200 m in elevation within a patchwork of pasture and logged forest and consists of only four adults within an area of well over 100 ha. The CD popu- lation grows at 20 m altitude and consists of remnant in- dividuals within pasture under conditions similar to those of population RB, except at about half the density and without any large forested area nearby. The last pop- ulation is located within a forest reserve (I,b) of an agri- cultural university. This forest has been heavily logged, and only six reproductive trees of P. elegans remain
Conservation Biology Volume 10, No. 3 ,June 1996
760 Genetic DiversitF and Mating System in PitheceU0bium elegans Ha//et a/.
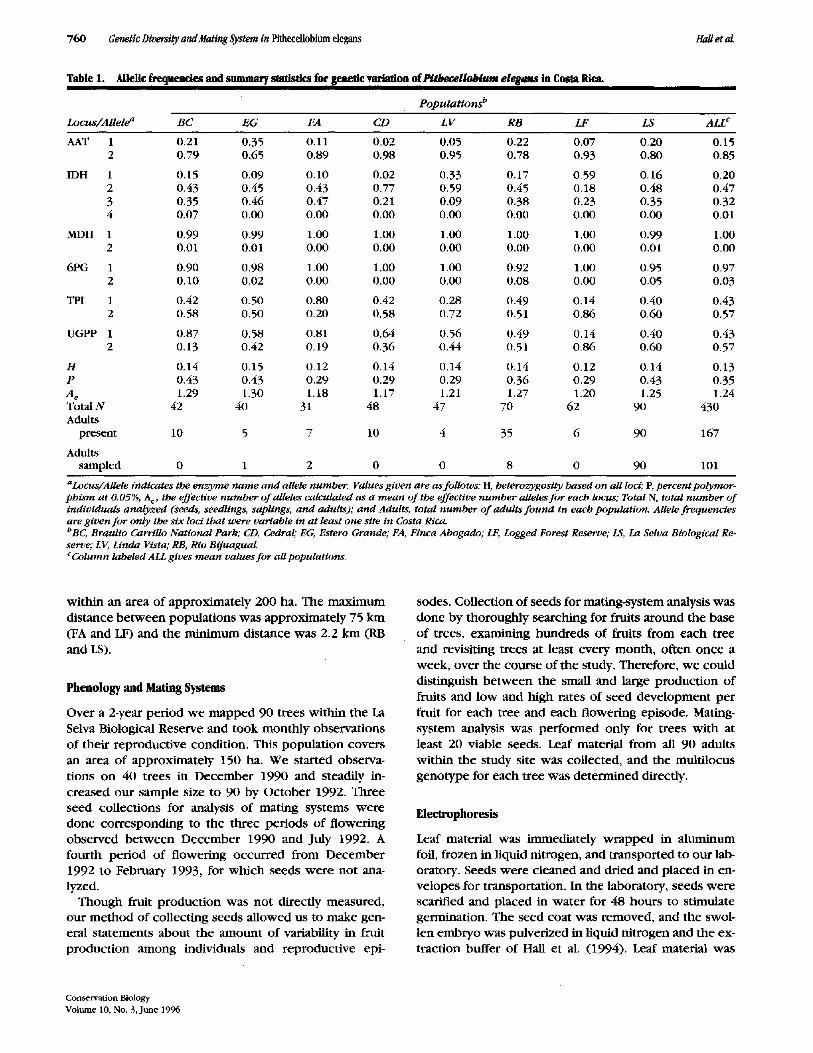
Table 1. AHelic frequencies and summary statistics for genetic variation ofPitheceilobium elegans in Costa Ric~
P o p u l a t i o n s t~
Locus /A l l e l e a B C E G FA CD L V R B LF LS ALL c
AAT 1 0.21 0.35 0.11 0.02 0.05 0.22 0.07 0.20 0.15 2 0.79 0.65 0.89 0.98 0.95 0.78 0.93 0.80 0.85
IDH 1 0.15 0.09 0.10 0.02 0.33 0.17 0.59 0.16 0.20 2 0.43 0.45 0.43 0.77 0.59 0.45 O. 18 0.48 0.47 3 0.35 0.46 0.47 0.21 0.09 0.38 0.23 0.35 0.32 4 0.07 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.01
MDH 1 0.99 0.99 1.00 1.00 1.00 1.00 1.00 0.99 1.00 2 0.01 0.01 0.00 0.00 0.00 0.00 0.00 0.01 0.00
6PG 1 0.90 0.98 1.00 1.00 1.00 0.92 1.00 0.95 0.97 2 0.10 0.02 0.00 0.00 0.00 0.08 0.00 0.05 0.03
TPI 1 0.42 0.50 0.80 0.42 0.28 0.49 0.14 0.40 0.43 2 0.58 0.50 0.20 0.58 0.72 0.51 0.86 0.60 0.57
UGPP 1 0.87 0.58 0.81 0.64 0.56 0.49 0.14 0.40 0.43 2 0.13 0.42 0.19 0.36 0.44 0.51 0.86 0.60 0.57
H 0.14 0.15 0.12 0.14 0.14 0.14 0.12 0.14 0.13 P 0.43 0.43 0.29 0.29 0.29 0.36 0.29 0.43 0.35 A e 1.29 1.30 1.18 1.17 1.21 1.27 1.20 1.25 1.24 Total N 42 40 31 48 47 70 62 90 430 Adults
present 10 5 7 10 4 35 6 90 167
Adults sampled 0 1 2 0 0 8 0 90 101
aLocus/Allele indicates the e n z y m e n a m e a n d allele number . Values g iven are as follows: H, heterozygosiCy based on all loci; P, percent polymor- p h t s m a t 0.05%, A e, the effective n u m b e r o f alleles calculated as a m e a n o f the effective n u m b e r alleles f o r each locus; Total N, total n u m b e r o f individuals ana ly zed (seeds, seedlings, saplings, a n d adults); a n d Adults, total n u m b e r o f adults f o u n d in each populat ion. Allele f requencies are g iven f o r only the six loci that were variable in a t least one site in Costa Rica. bBC, Braulio Carrillo Nat ional Park; CD, Cedral; EG, Estero Grande; FA, Finca Abogado; LF, Logged Forest Reserve; LS, La Selva Biological Re- serve; LV, Linda Vista; RB, Rio Bijuagual. C Column labeled ALL gives m e a n values f o r aU populattons.
wi th in an area o f a p p r o x i m a t e l y 200 ha. The m a x i m u m d is tance b e t w e e n popu la t i ons was a p p r o x i m a t e l y 75 k m (FA and LF) and the m i n i m u m d is tance was 2.2 k m (RB and LS).
Phenology and Mating Systems
Over a 2-year p e r i o d w e m a p p e d 90 t rees w i th in t he La Selva Biological Reserve and took m o n t h l y observa t ions o f the i r r ep roduc t i ve condi t ion . This p o p u l a t i o n covers an area o f a p p r o x i m a t e l y 150 ha. W e s tar ted observa- t ions on 40 t rees in D e c e m b e r 1990 and s teadi ly in- c r ea sed our sample size to 90 by O c t o b e r 1992. Three seed co l lec t ions for analysis of ma t ing sys tems w e r e done c o r r e s p o n d i n g to the th ree pe r i ods o f f lower ing o b s e r v e d b e t w e e n D e c e m b e r 1990 and July 1992. A four th p e r i o d o f f lower ing o c c u r r e d f rom D e c e m b e r 1992 to Februa ry 1993, for w h i c h seeds w e r e no t ana- lyzed.
T h o u g h frui t p r o d u c t i o n was no t d i rec t ly measured , ou r m e t h o d o f co l lec t ing seeds a l l owed us to make gen- eral s t a t emen t s abou t the amoun t of var iabi l i ty in fruit p r o d u c t i o n a m o n g individuals and r ep roduc t i ve epi-
sodes. Col lec t ion o f seeds for mat ing-sys tem analysis was done by t ho rough ly sea rch ing for fruits a round the base of t rees, examin ing h u n d r e d s o f fruits f rom each t ree and revisi t ing t rees at least eve ry mon th , of ten o n c e a week , over the cour se of t he study. Therefore , w e cou ld d is t inguish b e t w e e n the small and large p r o d u c t i o n o f fruits and low and h igh rates o f s eed d e v e l o p m e n t p e r fruit for each t ree and each f lower ing ep i sode . Mating- sys tem analysis was p e r f o r m e d on ly for t rees w i th at least 20 viable seeds. Leaf mater ia l f rom all 90 adul ts w i th in the s tudy site was co l lec ted , and the mul t i locus g e n o t y p e for each t ree was d e t e r m i n e d direct ly.
Electrophoresis
Leaf mater ia l was immed ia t e ly w r a p p e d in a luminum foil, f rozen in l iquid n i t rogen , and t r a n s p o r t e d to ou r lab- oratory. Seeds w e r e c l e a ne d and d r i ed and p l a c e d in en- ve lope s for t ranspor ta t ion . In the laboratory , seeds w e r e scarif ied and p l aced in w a t e r for 48 hours to s t imulate germinat ion . The seed coa t was r emoved , and the swol- len e m b r y o was pu lve r i zed in l iquid n i t rogen and the ex- t rac t ion buffer of Hall et al. (1994). Leaf mater ia l was
Conservation Biology Volume 10, No. 3, June 1996
Hall et al. Genetic OiversiCy and Mating System in PitheceUobium elegans 761
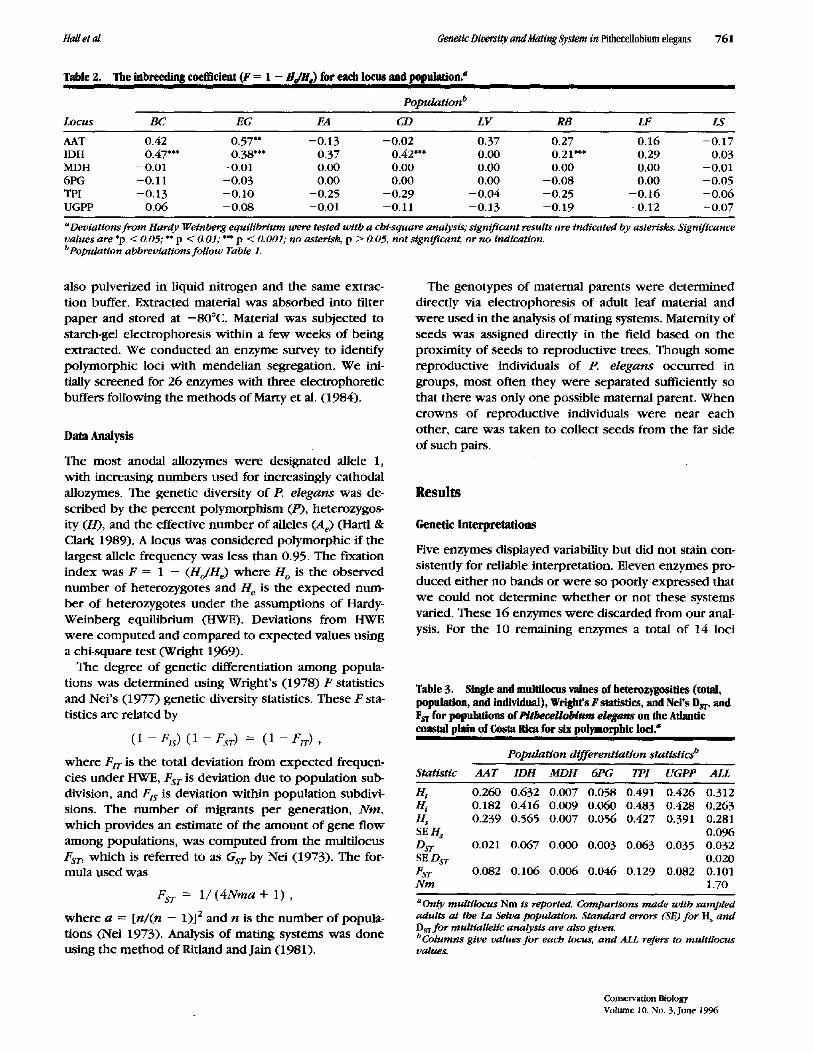
Table 2. The inbreeding coefficient (F = 1 - HdHe) for each locus and population?
Population °
Locus BC EG FA CD L V RB LF LS
AAT 0.42 0.57** - 0 . 1 3 --0.02 0.37 0.27 0.16 - 0 . 1 7 IDH 0.47*** 0.38*** 0.37 0.42*** 0.00 0.21"** 0.29 0.03 MDH -0.01 -0.01 0.00 0.00 0.00 0.00 0.00 -0.01 6PG -0.11 -0.03 0.00 0.00 0.00 -0.08 0.00 -0.05 TPI -0.13 -0.10 -0.25 -0.29 -0.04 -0.25 -0.16 -0.06 UGPP 0.06 -0.08 -0.01 -0.11 -0.13 -0.19 -0.12 -0.07
aDeviattons f ro m Hardy Wetnberg equilibrium were tested with a chi-square analysis; significant results are indicated by asterisks. Significance values are *p < 0.05; ** p ~ 0.01; *** p < 0.001; no asterisk, p > 0.05, not significant, or no indication. bPopulat~on abbreviations fol low Table 1.
also pulverized in liquid nitrogen and the same extrac- tion buffer. Extracted material was absorbed into filter paper and stored at -80°C. Material was subjected to starch-gel electrophoresis within a few weeks of being extracted. We conducted an enzyme survey to identify polymorphic loci with mendelian segregation. We ini- tially screened for 26 enzymes with three electrophoretic buffers following the methods of Marty et al. (1984).
Data Analysis
The most anodal allozymes were designated allele 1, with increasing numbers used for increasingly cathodal allozymes. The genetic diversity of P. e legans was de- scribed by the percent polymorphism (/~, heterozygos- ity (H), and the effective number of alleles (Ae) (Hartl & Clark 1989). A locus was considered polymorphic if the largest allele frequency was less than 0.95. The fixation index was F = 1 - ( H J H e ) where H o is the observed number of heterozygotes and H e is the expected num- ber of heterozygotes under the assumptions of Hardy- Weinberg equilibrium (HWE). Deviations from HWE were computed and compared to expected values using a chi-square test (Wright 1969).
The degree of genetic differentiation among popula- tions was determined using Wright's (1978) F statistics and Nei's (1977) genetic diversity statistics. These F sta- tistics are related by
(1- -F ls ) ( 1 - F s r ) = ( 1 - - F I T ) ,
where Fxr is the total deviation from expected frequen- cies under HWE, Fsr is deviation due to population sub- division, and Fxs is deviation within population subdivi- sions. The number of migrants per generation, N m , which provides an estimate of the amount of gene flow among populations, was computed from the multflocus Fsr, which is referred to as Gsr by Nei (1973). The for- mula used was
FST = 1/ ( 4 N m a + 1) ,
where a = [n/ (n - 1)] 2 and n is the number of popula- tions (Nei 1973). Analysis of mating systems was done using the method of Ritland and Jain (1981).
The genotypes of maternal parents were determined directly via electrophoresis of adult leaf material and were used in the analysis of mating systems. Maternity of seeds was assigned directly in the field based on the proximity of seeds to reproductive trees. Though some reproductive individuals of P. elegan$ occurred in groups, most often they were separated sufficiently so that there was only one possible maternal parent. When crowns of reproductive individuals were near each other, care was taken to collect seeds from the far side of such pairs.
Results
Genetic Interpretations
Five enzymes displayed variability but did not stain con- sistently for reliable interpretation. Eleven enzymes pro- duced either no bands or were so poorly expressed that we could not determine whether or not these systems varied. These 16 enzymes were discarded from our anal- ysis. For the 10 remaining enzymes a total of 14 loci
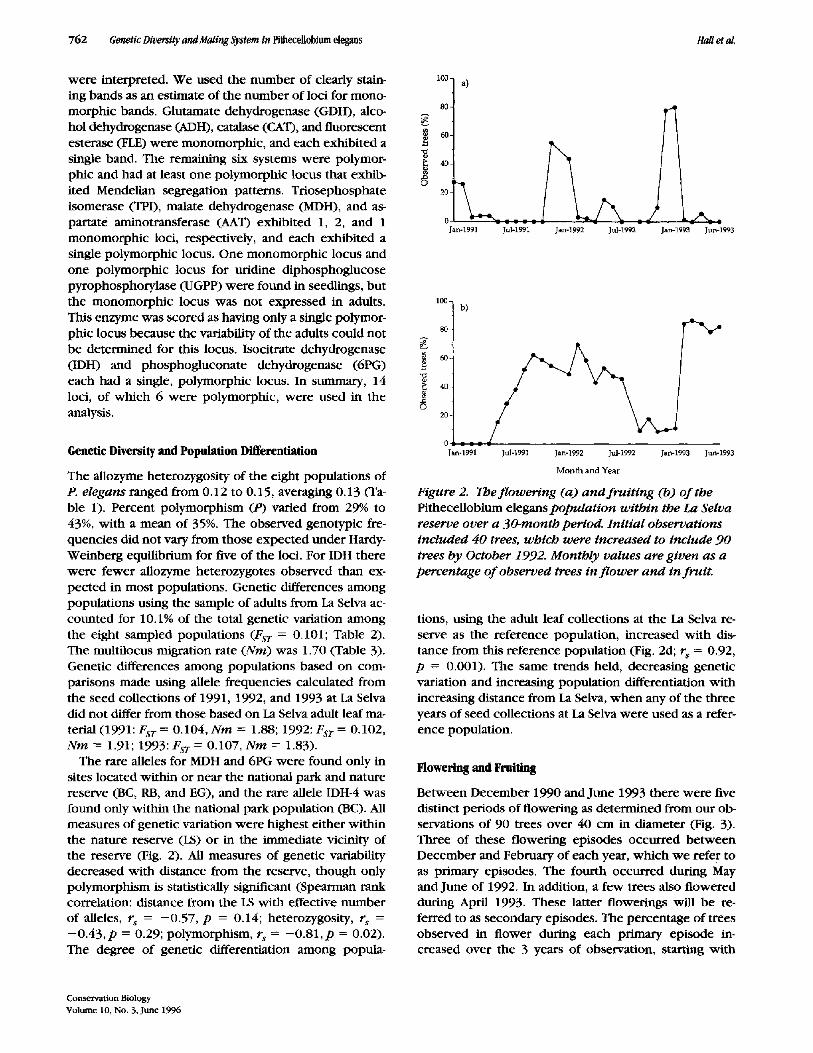
Table 3. Single and multilocus values of heterozygosities (total, population, and individual), Wright's F statistics, and Nei's DST, and FST for populations ofPitheceliobium elegans on the Atlantic coastal plain of Costa Rica for six polymorphic loci?
Popu la t ion d i f f e ren t ia t ion s ta t t s t i c~
Statistic A A T ID H M D H 6PG TPI UGPP ALL
H t 0.260 0.632 0.007 0.058 0.491 0.426 0.312 H~ 0.182 0.416 0.009 0.060 0.483 0.428 0.263 Hs 0.239 0.565 0.007 0.056 0.427 0.391 0.281 SE ns 0.096 DST 0.021 0.067 0.000 0.003 0.063 0.035 0.032 SE DST 0.020 Fsr 0.082 0.106 0.006 0.046 0.129 0.082 0.101 N m 1.70
a On/y maltaocus Nm/s reported. Comparisons made with sampled adults at the La Selva population. Standard errors (SE) for H s a n d DsT fo r multtalleltc analysis are also given.
Columns give values for each locus, and ALL refers to maltilocus values.
Conservation Biology Volume 10, No. 3,June 1996
762 Genetic Diversity and Mating System in PitheceU0bium elegans Hall et al.
were interpreted. We used the number of clearly stain- ing bands as an estimate of the number of loci for mono- morphic bands. Glutamate dehydrogenase (GDH), alco- hol dehydrogenase (ADH), catalase (CAT), and fluorescent esterase (FLE) were monomorphic, and each exhibited a single band. The remaining six systems were polymor- phic and had at least one polymorphic locus that exhib- ited Mendelian segregation patterns. Triosephosphate isomerase (TPI), malate dehydrogenase (MDH), and as- partate aminotransferase (AAT) exhibited 1, 2, and 1 monomorphic loci, respectively, and each exhibited a single polymorphic locus. One monomorphic locus and one polymorphic locus for uridine diphosphoglucose pyrophosphorylase (UGPP) were found in seedlings, but the monomorphic locus was not expressed in adults. This enzyme was scored as having only a single polymor- phic locus because the variability of the adults could not be determined for this locus. Isocitrate dehydrogenase (IDH) and phosphogluconate dehydrogenase (6PG) each had a single, polymorphic locus. In summary, 14 loci, of which 6 were polymorphic, were used in the analysis.
Genetic Diversity and Population Differentiation
The allozyme heterozygosity of the eight populations of P. e legans ranged from 0.12 to 0.15, averaging 0.13 (Ta- ble 1). Percent polymorphism (P) varied from 29% to 43%, with a mean of 35%. The observed genotypic fre- quencies did not vary from those expected under Hardy- Weinberg equilibrium for five of the loci. For IDH there were fewer allozyme heterozygotes observed than ex- pected in most populations. Genetic differences among populations using the sample of adults from La Selva ac- counted for 10.1% of the total genetic variation among the eight sampled populations (FsT = 0.101; Table 2). The multilocus migration rate ( N m ) was 1.70 (Table 3). Genetic differences among populations based on com- parisons made using allele frequencies calculated from the seed collections of 1991, 1992, and 1993 at La Selva did not differ from those based on La Selva adult leaf ma- terial (1991: FST = 0.104, N m = 1.88; 1992: Fsr = 0.102, N m -- 1.91; 1993: FST = 0.107, N m -- 1.83).
The rare alleles for MDH and 6PG were found only in sites located wRMn or near the national park and nature reserve (BC, RB, and EG), and the rare allele IDH-4 was found only within the national park population (BC). All measures of genetic variation were highest either within the nature reserve (LS) or in the immediate vicinity of the reserve (Fig. 2). All measures of genetic variability decreased with distance from the reserve, though only polymorphism is statistically significant (Spearman rank correlation: distance from the LS with effective number of alleles, r s -- -0 .57, p = 0.14; heterozygosity, r s = - 0 . 4 3 , p = 0.29; polymorphism, r s -- - 0 . 8 1 , p = 0.02). The degree of genetic differentiation among popula-
Jan-1991
100-
80-
60-
40-
20-
0
a)
Ju1-1991 Jan-1992 Ju1-1992 Jan-1993 Jun-1993
8
lO0- b)
8o-
~ 40-
8 20-
Jan-1991 Ju1-1991 J a n - 1 9 9 2 Ju1-1992 Jan-1993 Jun-1993
Month and Year
Figure 2. The f l o w e r i n g (a) a n d f r u i t i n g (b) o f the Pithecellobium elegans p o p u l a t i o n w i t h i n the La Selva reserve over a 3 0 - m o n t h per iod . In i t i a l observa t ions i nc luded 40 trees, w h i c h w e r e increased to inc lude 9 0 trees by October 1992. M o n t h l y va lues are g i v e n as a p e r c e n t a g e o f observed trees in f l o w e r a n d in f ru i t .
tions, using the adult leaf collections at the La Selva re- serve as the reference population, increased with dis- tance from this reference population (Fig. 2d; r s = 0.92, p = 0.001). The same trends held, decreasing genetic variation and increasing population differentiation with increasing distance from La Selva, when any of the three years of seed collections at La Selva were used as a refer- ence population.
Flowering and Fruiting
Between December 1990 and June 1993 there were five distinct periods of flowering as determined from our ob- servations of 90 trees over 40 cm in diameter (Fig. 3). Three of these flowering episodes occurred between December and February of each year, which we refer to as primary episodes. The fourth occurred during May and June of 1992. In addition, a few trees also flowered during April 1993. These latter flowerings will be re- ferred to as secondary episodes. The percentage of trees observed in flower during each primary episode in- creased over the 3 years of observation, starting with
Conservation Biology Volume 10, No. 3, June 1996

Haft et al. Genetic Divers#y and Mating System in Pithecell0bium elegans 763
30-
45 LS EG i
40- BC RB
P 35-
30-
25-
20
1.4-
1.3-
A e L2-
1.1-
1.0
(a)
PA CD LF • I L V •
, i , i , I , i , i , i , i , [
10 2.0 30 40 50 60 '70 80
H t
0.40-
0.35.
0.30.
0.25.
0.20
(b)
OEG
RB
OLV
FA • OLF • C D
' I ' I ' t ' ; ' u ' u ' u ' u
1 0 2 O 3 O 4 O 5 O 6 O 7 O 8 O
(c) EG
RB
LS OLV OLF
FA CD .~
' / ' u ' t ' I ' u ' I ' n ' J
10 20 30 40 50 60 70 80
o.o~5-
0.04-
0.03-
0.02-
0.012
0.O0 LS
( d ) o ~
FA • •LV
• C D •EG
' / ' I ' I ' I ' I ' I " i ' I
10 20 30 40 50 60 70 80
Distance from reserve population (krn)
Figure 3. The relationship be- tween measures of genetic vari- ab i l i t y - percent polymorphism (a); heterozygosity for polymor- phic loci (HE) (b); and effective number of alleles (c)--for popula- tions in relationship to their dis- tance from the central population of the forest reserve (La Selva, LS) and the multilocus Fsrfor each population paired with the cen- tral reserve population (LS) (d). Population symbols follow the de- scription in Table 1.
33% of the individuals and rising to 80%. A small group of 12 trees (15% of observed) and then five trees (6% of observed) flowered during the two secondary episodes, respectively.
Not all 90 trees were observed for the entire 30-month period, but our observations were sufficient to identify variation among individuals in flowering phenology. All but seven trees (8%) were observed in flower at least once. However, these seven trees did produce fruits and therefore we surmise that they flowered while not un- der observation at the beginning of this study. Most trees (73%) flowered at least once during a primary re- productive episode. But three of these trees flowered at intervals of greater than 1 year, skipping the second De- cember-to-February reproductive season. A total of 17 trees (19%) flowered at least once between April and June. Of these 17 trees, 14 (74%) had flowered within the 6 months prior to the second reproductive episodes.
Following the 1991 flowering, we collected fruits from 39 trees. Nine of these trees (23%), though produc- ing copious amounts of fruits of which many hundreds were examined, did not have fruits with mature or via- ble seeds. Though seed predation was an obvious factor, many fruits had tiny, immature seeds. In addition, we were able to collect sufficient seeds for mating-system analysis from only 19 trees, further evidence of poor seed production. Following the 1992 flowering episode, only one (2%) of the 61 sampled trees produced empty pods. We were able to collect at least 20 viable seeds from 47 individuals for mating-system analysis of the 1992 flowering with relative ease. For the 14 trees that flowered between May and June of 1992, only three (27%) produced sufficient fruit with mature seeds for
mating-system analysis. The remaining trees either failed to produce fruits, or most fruits that were found co.n- rained aborted seeds.
Mating System
An analysis of the temporal variability in the rate of out- crossing was performed using 19 trees that flowered from December 1990 to February 1991 and 47 trees that flowered from December 1991 to February 1992. Our phenological observations indicated that the percentage of flowering trees in the population was lower during the 1990-1991 episode. Allele frequencies for seeds from the May-June 1992 flowering were also obtained, but there were insufficient seeds for a complete mating- system analysis for this reproductive episode.
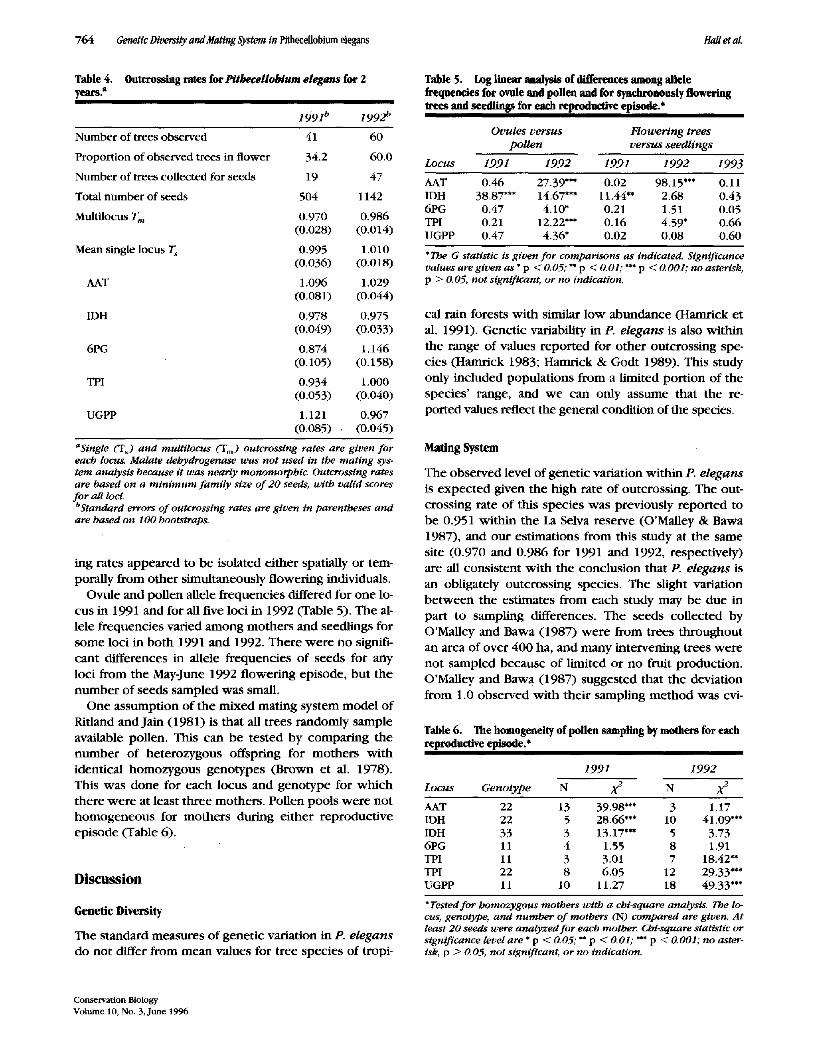
Malate dehydrogenase was nearly monomorphic within the population and was dropped from the mat- ing-system analysis, leaving five polymorphic loci for es- timation of outcrossing rates. The multilocus outcross- ing rate was 0.970 and 0.986 for 1991 and 1992, respectively (Table 4). Neither of these outcrossing rates is significantly different from 1.0 using the standard er- rors as calculated from 100 bootstraps of sampling within famih'es (Ritland & Jain 1981). The mean of sin- #e-locus estimates of outcrossing were also not signifi- cantly different from the estimates based on multilocus values. Though sample sizes were small, outcrossing rates were calculated for individual trees for each repro- ductive episode. Outcrossing rates of less than 1.0 were found for three trees in 1991 (16%) and nine trees in 1992 (19%). None of the individuals with low outcross-
C o n s e r v a t i o n B i o l o g y
V o l u m e 10, N o . 3 , J u n e 1 9 9 6
764 Genetic Dtversi~ and Mating System in PitheceU0bium elegans Hall et al.
Table 4. years, a
Outcrossing rates for Pithecellobium elegans for 2
1991 b 1992 b
Number of trees observed
Proportion of observed trees in flower
Number of trees collected for seeds
Total number of seeds
Multilocus T m
Mean single locus T s
AAT
IDH
6PG
TPI
UGPP
41 60
34.2 60.0
19 47
504 1142
0.970 0.986 (0.028) (0.014)
0.995 1.010 (0.036) (0.018)
1.096 1.029 (0.081) (0.044)
0.978 0.975 (0.049) (0.033)
0.874 1.146 (0.105) (0.158)
0.934 1.000 (0.053) (0.040)
1.121 0.967 (0.085) • (0.045)
aSingle ('Us) and multllocus (Tin) outcrosstng rates are given fo r each locus. Malate dehydrogenase was not used in the mating sys- tem analysis because it was nearly monomorphic. Outcrossing rates are based on a m i n i m u m fami ly size o f 20 seeds, with valid scores f o r all loci. O Standard errors o f outcrossing rates are given in parentheses and are based on 100 bootstraps.
ing rates appeared to be isolated ei ther spatially or tem- porally from other simultaneously flowering individuals.
Ovule and pollen allele frequencies differed for one lo- cus in 1991 and for all five loci in 1992 (Table 5). The al- lele frequencies varied among mothers and seedlings for some loci in both 1991 and 1992. There were no signifi- cant differences in allele frequencies of seeds for any loci from the May-June 1992 flowering episode, but the number of seeds sampled was small.
One assumption of the mixed mating system model of Ritland and Jain (1981) is that all trees randomly sample available pollen. This can be tested by comparing the number of heterozygous offspring for mothers with identical homozygous genotypes (Brown et al. 1978). This was done for each locus and genotype for which there were at least three mothers. Pollen pools were not homogeneous for mothers during either reproduct ive episode (Table 6).
Discussion
Genetic Diversity
The standard measures of genetic variation i n P. e l e g a n s
do not differ from mean values for tree species of tropi-
Table 5. Log linear analysis of differences among allele frequencies for ovule and pollen and for synchronously flowering trees and seedlings for each reproductive episode.*
Ovules versus F l o w e r i n g trees po l l en versus seedl ings
Locus 1991 1992 1991 1992 1993
AAT 0.46 27.39*** 0.02 98.15"** 0.11 IDH 38.87*** 14.67"** 11.44"* 2.68 0.43 6PG 0.47 4.10" 0.21 1.51 0.05 TPI 0.21 12.22"** 0.16 4.59* 0.66 UGPP 0.47 4.36* 0.02 0.08 0.60
* The G statistic is given fo r comparisons as indicated. Significance values are given as * p < 0.05; ** p < 0.01; *** p < 0.001; no asterisk, p > 0.05, not significant, or no indication.
cal rain forests with similar low abundance (Hamrick et al. 1991). Genetic variability in P. e l e g a n s is also within the range of values repor ted for other outcrossing spe- cies (Hamrick 1983; Hamrick & Godt 1989). This study only included populat ions from a limited port ion of the species ' range, and we can only assume that the re- ported values reflect the general condition of the species.
Mating System
The observed level of genetic variation within P. e l e g a n s
is expec ted given the high rate of outcrossing. The out- crossing rate of this species was previously repor ted to be 0.951 within the La Selva reserve (O'Malley & Bawa 1987), and our estimations from this study at the same site (0.970 and 0.986 for 1991 and 1992, respectively) are all consistent with the conclusion that P. e l e g a n s is an obligately outcrossing species. The slight variation be tween the estimates from each study may be due in part to sampling differences. The seeds collected by O'Malley and Bawa (1987) were from trees throughout an area of over 400 ha, and many intervening trees were not sampled because of limited or no fruit production. O'Malley and Bawa (1987) suggested that the deviation from 1.0 observed with their sampling method was evi-
Table 6. The homogeneity of pollen sampling by mothers for each reproductive episode.*
1991 1992
Locus Geno type N )(2 N X 2
AAT 22 13 39.98*** 3 1.17 IDH 22 5 28.66*** 10 41.09*** IDH 33 3 13.17"** 5 3.73 6PG 11 4 1.55 8 1.91 TPI 11 3 3.01 7 18.42"* TPI 22 8 6.05 12 29.33*** UGPP 11 10 11.27 18 49.33***
*Tested fo r homozygous mothers with a chi-square analysis. The lo- cus, genotype, and number o f mothers (N) compared are given. At least 20 seeds were analyzed fo r each mother. Cht-square statistic or significance level are * p < 0.0_5; ** p < 0.01; *** p < 0.001; no aster- isk, p > 0.05, not significant, or no indication.
Conservation Biology Volume 10, No. 3, June 1996

Hall et al. Genetic l~'vem'ty and Mating System in Pithecell0bium elegans 765
dence for populat ion subdivision. The sampling for our investigation was done in a completely enumerated con- tiguous population. Therefore, we concur with their in- ference and suggest that our higher outcrossing rate is the result of avoiding sampling across subpopulations.
The pat tern of observed flowering indicates that a population of P. elegans flowers regularly on an annual basis. But the actual percentage of the population that f lowered during the primary reproductive interval (De- cember to February) ranged from 33% to 80%. A de- crease in stand density or effective number of individu- als in f lower may lead to reduced effective outcrossing rates (Murawski et al., 1990; Murawski & Hamrick 1991). When the number of flowering individuals was halved, outcrossing rates were not reduced in P. elegans. These observations suggest that the population rate of out- crossing is always high in P. elegans.
A high rate of outcrossing for the population does not necessarily imply that outcrossing rates were identical for all individuals within the population or that mating was at random with respect to other trees. The variation in the degree of synchrony within the population will al- ter the effective distance be tween flowering trees, which in turn may limit the actual number of matings among individuals. In our study, as observed by O'Mal- ley and Bawa (1987), homozygous mothers sampled the pollen pool heterogeneously, and a few trees from each reproductive episode exhibited individual outcrossing rates far be low 1. In this population, therefore, it ap- pears that individual trees do not receive pollen at ran- dom from all synchronously flowering trees and do not breed with many other individuals.
For some loci (A_AT, IDH, and TPI), significant allellc differences existed be tween the trees that f lowered at the same time and seeds produced from that flowering episode within the La Selva population. A number of processes could produce such results: variable number of progeny per maternal tree, the migration of pollen from outside of the population, and variation in success- ful pollination and fruit development among pollen do- nors. Though the number of progeny per maternal trees used for the mating-system analysis was not identical, the variance in progeny number was not high (mean _+ standard deviation: 1991:26.5 - 5.9; 1992:24.3 -+ 5.8). There is evidence that some of the seeds could not have been fathered by trees within the sampled population (P. Hall, unpublished paternity analysis). In addition, during the 1992 primary flowering episode, flowering was also extensive at the two nearest populations, Braulio C a r ~ o and Rio Bijagual (P. Hall, personal obser- vation). These observations indicate that pollen may well be migrating into the reserve population. We can- not determine at this time, however, ff there was varia- tion in paternal reproductive success, but this is also likely to have contributed to allelic differences be tween maternal trees and seed crops.
In contrast, during the May-June 1992 flowering pe- riod, when only 12 trees (15% of the observed popula- tions) flowered, there were no differences in allele fre- quencies be tween flowering plants and the seed crop. If the number of progeny per maternal tree or differential paternal success are resulting in allelic differences, they do not appear to be of great importance during the low- density flowering period. The two nearby populations also exhibited a very low level of flowering trees during this period ( two trees at Braulio Carrillo and one tree at Rio Bijagual), providing little opportunity for pollen mi- gration because of the greater effective distance be- tween synchronously flowering trees.
Genetic Differentiation among Populations
The small amount of population differentiation (multflo- cus Fsr = 0.101) is also consistent with high rates of out- crossing in a species. The estimated number of migrants per generation, Nm, is greater than one for P. elegans, higher than the theoretical critical value be low which populations may begin to differentiate because of ge- netic drift. It is difficult to distinguish be tween current and past patterns of gene flow and their contribution to population differentiation. Samples of adults can be used to represent patterns of gene flow in the past, whereas seedlings can represent present gene flow patterns. Us- ing the La Selva population as a reference, both adults and seedlings show the same degree of population dif- ferentiation as seedlings from the other seven popula- tions. But we are missing the comparison of adults at La Selva to adults in other populations because of the pan- city of adults in these fragments. The limited results ob- tained suggest that gene flow may be important in pre- venting a large amount of genetic drift, at least for geographically close populations.
High outcrossing rates should uniformly yield low de- grees of population differentiation, yet studies of genetic differentiation among populations in tropical tree spe- cies have produced highly variable results. The majority of these studies come from populations that were sepa- rated by less than 5 km (Hamrick & Murawski 1991). In contrast, Chase et al. (1995) reported a multilocus Fsr of 0.117 for Cordia aUiodora at distances up to 1100 km. Multilocus Fsr values of 0.163 and 0.111 have been re- ported for Sternonoprous oblongifolius and Sborea tra- pezifolia, respectively, both of which are endemic to Sri Lanka (Murawski & Bawa 1994). These values cover populations separated by 10 and 44 km, respectively. In addition to P. elegans, two other species have also been studied in the same region of the Atlantic coastal plain of Costa Rica. These species, Pentacletbra macroloba and Carapa guianensis, are much more abundant than P. el- egans. The degree of population differentiation found in these two species was 22% and 5%, respectively, of the total genetic variation detected via isozyme studies (Hall
Conservation Biology Volume 10, No. 3, June 1996

766 Genetic Diversi O, and Mating System in PitheceUobium elegans Hall et al.
et al. 1994a, 1994b). In light of extensive differences in the natural history of all these species, it is difficult at this t ime to determine which aspect of their reproduc- tive biology may primarily account for the levels of pop- ulation differentiation found in each species. But the presence of high levels of outcrossing is not enough by itself to predict the degree or pattern of population dif- ferentiation.
Forest Fragmentation
The continuity of forest cover has been decreasing rap- idly in the last few decades throughout the Atlantic coastal plain of Costa Rica (Sader & Joyce 1988). The re- sult for P. elegans, and for a number of other tree spe- cies, has been reduced population sizes and greater physical isolation of populations. The fragmented popu- lations of this study were relatively small, ranging from only four trees in the logged reserve (LF) most distant to La Selva (LS) to a larger group of 35 trees remaining in pasture bordering the La Selva reserve (population RB). Though the density of the fragmented populations is dif- ficult to quantify, their degree of isolation is more easily assessed because the adults of P. elegans can be seen clearly from great distances because of their large size and distinctive crown shape. Generally, isolation of pop- ulations also increased as a function of increasing dis- tance from La Selva. Individual trees located on forested ridges be tween Braulio Carrillo, Rio Bijagual, and La Selva were clearly visible, providing possible linkages be tween these populations, whereas the logged forest population is surrounded by many square kilometers of secondary forest and agriculture. Allozyme heterozygos- ity, polymorphism, and the effective number of alleles all declined as distance of sampled populations from the reserve increased. The individuals sampled were prod- ucts of recent reproduction and hence reflect current patterns of mating within and among populations. Therefore, we conclude that fragmentation of a once large and contiguous area of tropical forest has resulted in loss of genetic diversity within populations of P. elegans.
Distinguishing be tween the multitude of effects of fragmentation can be difficult. For instance, the varia- tion in flowering intensity and synchrony appear to be a natural phenomenon, not one brought on by the circum- stances of deforestation or isolation. But the lack of com- plete flowering synchrony in P. elegans and periods of low flowering frequency in a large population mimic the effects of fragmentation in terms of low effective popu- lation size and the greater spatial isolation of individuals for at least that reproductive episode. Episodes of low flowering intensity produced fewer seeds and no evi- dence of pollen immigration into the population. There- fore, the maintenance of gene diversity within and gene flow among fragmented populations may be further
jeopardized by natural variation in flowering synchrony. Poor reproduction in spatially isolated individuals in small forest fragments is readily apparent by poor fruit product ion among trees during a t ime when fruit pro- duction was high in the La Selva population (June- August).
In spite of the fragmentation occurring in the immedi- ate vicinity of the forest reserve, there may still be im- portant genetic links be tween the reserve population and the immediately surrounding forest fragments. First, forest fragments that were very close to La Selva (LS) and Braulio Carrillo (BC) had values of genetic diversity nearly identical to those of the reserve populations for samples from recent reproductive events. Second, there was also indication that pollen was arriving from outside of the La Selva population during periods of synchro- nous intense flowering of all neighboring populations. Third, there are large, reproductive individuals in the in- tervening area be tween populations that f lowered syn- chronously with all surrounding populations. Thus, the current degree of separation be tween the two sites within the reserve and their near neighbors within pas- ture has not resulted in a disruption of gene flow, but we cannot rule out the possibility that there has been in- sufficient time to detect changes in genetic diversity of differentiation.
The critical and missing information is the effective rate of regeneration within isolated populations. For some of our studied populations, demographic and ge- netic factors probably contribute to poor or improbable regeneration. Even though the samples for this study were of seeds and seedlings, there is evidence of at least the possibility of regeneration at the logged forest and Linda Vista populations. (Our meager seed collection represented the entire reproductive effort of the year.) The product ion of a few seeds per adult tree in a single year does not bode well for successful regeneration. For population fragments near La Selva, regeneration ap- pears more probable because there were more seeds and seedlings than needed for genetic sampling. There were also a number of juveniles (saplings of greater than 1 cm diameter at breast height) in the Rio Biajagual and Estero Grande populations. We have, as yet, little evi- dence of genetic erosion at these sites. Even in the large reserve, however, where regeneration is occurring, fruit production varies over time, and there is a high inci- dence of seedlings with mutations, such as albinism and negative geotropism. This may be evidence of a high ge- netic load, limitations on the number of mates, and pos- sibly consanguineous mating.
Our study points out the fragility of small, isolated populations of tropical trees, and the greater effective isolation that can occur because of natural variability in flowering synchrony. The La Selva-Braulio Carrillo for- est is the last remaining site of a large, contiguous popu- lation of P. elegans in the Atlantic plain of Costa Rica
Conservation Biology Volume 10, No. 3,June 1996

Hall et al. Genetic Diversity and Mating System in PitheceU0bium elegans 7 6 7
(R. Peralta, personal communication; P. Hall, personal observation). As the tendrils of forest and trees standing in pasture continue to disappear, more-severe isolation will occur. Coupled with the natural temporal variation in effective reproduction, this spells the doom of small populations in forest fragments. Continuing deforesta- tion outside the reserve will eventually eliminate any possible contribution remnant populations can provide to the genetic diversity of the last, large, natural popula- tion. We conclude that small, isolated populations will not persist, that forest fragments surrounding reserves can be of important genetic value, and that it is of paramount importance to protect as much natural forest as possible to maintain sustainable populations of P. elegans.
Acknowledgments
We thank Victor Robles and Rosa Sandoval for extensive field work and Lelia C. Orrell for help in the laboratory and in preparing this manuscript. Pamela Soltis and two anonymous reviewers contributed useful suggestions and editing. Software for computat ions of mating sys- tems and pairwise comparison of populations was sup- plied by K. Ritland and M. Slatkin, respectively. This re- search was suppor ted by the National Science Foundation, the Pew Charitable Trusts' Pew Scholars Program in Conservation and the Environment, and the Conservation, Food and Health Foundation.
Literature Cited
Ashton, P. S. 1969. Speciation among tropical forest trees; some deduc- tions in the light of recent evidence. Biological Journal of the Lin- naean Society London 1:155-196.
Bawa, K. S. 1983. Patterns of flowering in tropical plants. Pages 395- 410 in C. E. Jones and R. J. Little, editors. Handbook of experimen- tal pollination biology. Van Nostrand & Reinhold, New York.
Bawa, K. S. 1992. Mating systems, genetic differentiation and specia- t ion in tropical rain forest plants. Biotropica 24:250-255.
Bawa, K. S. 1993. Effects of deforestation and forest fragmentation on genetic diversity in tropical tree populations. Pages 10-16 in R. M. Drysdale, S. E. T. John, and A. C. Yap, editors. Genetic conservation and production of tropical forest seed. Association of Southeast Asian Nations-Canada Forest Tree Seed Centre, Cniang Mai, Thailand.
Bawa, K. S., D. R. Perry, and J. H. Beach. 1985. Reproductive biology of tropical lowland rain forest trees. I. Sexual systems and self-in- compatibility mechanisms. American Journal of Botany 72:331- 345.
Brown, A. H. D., D. Zohary, and E. Nevo. 1978. Outcrossing rates and heterozygosity in natural populations of Hordeum spontoneum Koch in Israel. Heredity 41:49-62.
Chase, M. R., D. Bushier, and K. S. Bawa. 1995. Conservation implica- tions of populat ion genetic structure of Cordia .alliodora, a Neo- tropical forest tree species. American Journal of Botany 82:468- 475.
Clark, D. A., and D. B. Clark. 1992. Life history diversity of canopy and
emergent trees in a Neotropical rain forest. Ecological Monographs 62:315-344.
Gentry, A. H., andJ. Terborgh. 1990. Composit ion and dynamics of the Cocha Cashu "mature" floodplain forest. Pages 542-564 in A. H. Gentry, editor. Four Neotropical forests. Yale University Press, New Haven, Connecticut.
Hall, P., M. R. Chase, and K. S. Bawa. 1994a. Low genetic variation but nigh populat ion differentiation in a c o m m o n tropical forest tree species. Conservation Biology 8:471-482.
Hall, P., L. C. Orrell, and K. S. Bawa. 1994b. Genetic diversity and mat- ing system in a tropical tree, Carapa guianenMs. American Journal of Botany 81:1104-1111.
Hamrick, J. L. 1983. The distribution of genetic variation within and among natural plant populations. Pages 335-348 in C. M. Schone- wald-Cox, S. H. Chambers, B. MacBryde, and L. Thomas, editors. Genetics and conservation. Benjamin Cummings Publishing Com- pany, Menlo Park, California.
Hamrick, J. L., and M. J. Godt. 1989. Allozyme diversity in plant spe- cies. Pages 43-63 in A. H. D. Brown, M. T. Clegg, A. L. Kahler, and B. S. Weir, editors. Plant populat ion genetics, breeding and germ- plasm resources. Sinauer Associates, Sunderland, Massachusetts.
Hamrick, J. L., and D. A. Murawski. 1991. Levels of allozyme diversity in populat ions of u n c o m m o n Neotropical tree species. Journal of Tropical Ecology 7:395-399.
Hamrick, J. L., M. J. W. Godt, D. A. Murawski, and M. D. Loveless. 1991. Correlations be tween specieg traits and allozyme diversity: implications for conservation biology. Pages 75-86 in D. A. Falk and K. E. Holsinger, editors. Genetics and conservation of rare plants. Oxford University Press, Oxford, United Kingdom.
Hartl, D. L., and A. G. Clark. 1989. Principles of population genetics. Sinauer Associates, Sunderland, Massachusetts.
Hubbell, S. P., and R. B. Foster. 1983. Diversity of canopy trees in a Neotropical forest and implications for conservation. Pages 25-41 in S. Sutton, T. C. Whitmore, and A. Chadwick, editors. Tropical rain forest: ecology and management . Blackwell, Oxford, United Kingdom.
Ledig, F. T., and M. T. Conlde. 1983. Gene diversity and genetic struc- ture in endemic Torrey pine (Ptnus torreyana Parry ex. Carr.). Evolution 73:79-85.
Lieberman, D., G. S. Hartshorn, M. Lieberman, and R. Peralta. 1990. Forest dynamics at La Selva biological station, Costa Rica, 1969- 1985. Pages 509-521 in A. H. Gentry, editor. Four Neotropical for- ests. Yale University Press, New Have, Connecticut.
Marty, T. L., D. M. O'Malley, and R. P. Guries. 1984. A manual for starch gel electrophoresis: n e w microwave edition. Staff paper ser. no. 20. Depar tment of Forestry, University of Wisconsin, Madison.
Murawsld, D. A., and K. S. Bawa. 1994. Genetic structure and mating system of Stemonoporus oblongifolius (Dipterocarpaceae) in Sri Lanka. American Journal of Botany 81:155-160.
Murawski, D. A., andJ . L. Hamrich. 1991. The effect of the density of flowering individuals on the mating systems of nine tropical tree species. Heredity 67:167-174.
Murawsld, D. A., J. L. Hamrick, S. P. Hubbell, and R. B. Foster. 1990. Mating systems of two Bombacaceous trees of a Neotropical moist forest. Oecologia 82:501-506.
Nei, M. 1973. Analysis of gene diversity in subdivided populations. Pro- ceedings of the National Academy of Sciences USA 70:3321-3323.
Nei, M. 1977. F-statistics and analysis of gene diversity in subdivided populations. Annals of Human Genetics 47:253-259.
Newstrom, L. E., G. W. Frankie, and H. G. Baker. 1991. Survey of long- term flowering patterns in lowland tropical rain forest trees at La Selva, Costa Rica. Pages 345-366 in C. Edelin, editor. L'arbre biolo- gie developpement . No. hors serie A 7, Naturalia Monspeliensia, Montpellier, France.
Conservation Biology Volume 10, No. 3, June 1996

7 6 8 Genetic Diversi~ and Mating System in PitheceU0bium elegans Ha//et a/.
O'Malley, D. M., and K. S. Bawa. 1987. Mating system of a tropical rain forest tree. American Journal of Botany 74:1143-1149.
Poore, M. E. D. 1968. Studies in Malaysian rain forest. I. The forest on the Triassic sediments in Jengka Forest Reserve. Journal of Ecology 56:143-196.
Ritland, K., and S. K. Jain. 1981. A model for the estimation of out- crossing rate and gene frequencies using independent loci. Hered- ity 47:35-52.
Sader, S. A., and Joyce, A. T. 1988. Deforestation rates and trends in Costa Rica, 1940 to 1983. Biotropica 20:11-19.
Whitmore, T. C. 1984. Tropical rain forests of the far east. Oxford Uni- versity Press, Oxford, United Kingdom.
Wright, S. 1969. Evolution and the genetics of populations. Vol. 2. The theory of gene frequencies. University of Chicago Press, Chicago.
Wright, S. 1978. Evolution and the genetics of populations. Voi. 4. Variability within and among natural populations. University of Chicago Press, Chicago.
Conservation Biology Volume 10, No. 3,June 1996





























