Embed Size (px)
Citation preview

INFECTION AND IMMUNITY, May 1994, p. 1813-1819 Vol. 62, No. 50019-9567/94/$04.00+0Copyright ©D 1994, American Society for Microbiology
Effect of Genetic Switching of Capsular Type on Virulenceof Streptococcus pneumoniae
TANYA KELLY, JOSEPH P. DILLARD, AND JANET YOTHER*
Department of Microbiology, University ofAlabama at Birmingham, Birmingham, Alabama 35294
Received 10 November 1993/Returned for modification 19 December 1993/Accepted 2 February 1994
To assess the role of capsular serotypes in the virulence of Streptococcus pneumoniae, we have constructedisogenic derivatives differing only in the type of capsule expressed. Strains of types 2, 5, and 6B were convertedto type 3 by transformation and selection for an erythromycin resistance marker linked to the type 3 capsulelocus. Characterization studies revealed that these type 3 derivatives were indistinguishable from the type 2,type 5, and type 6B parental strains in terms of restriction enzyme fragment pattern and expression ofpneumococcal surface protein A (PspA). Expression of the type 3 capsule did not alter the mouse virulence ofthe similarly virulent type 2 strain. However, alteration of capsule type had a profound effect on the virulenceof the type 5 and type 6B derivatives. The highly virulent type 5 strain was essentially avirulent when expressingthe type 3 capsule. Conversely, the 50% lethal dose of the relatively avirulent type 6B strain was reduced> 100-fold when the type 3 capsule was expressed. Thus, the serotype of capsule expressed has a major effecton virulence, and this effect is dependent upon the genetic background of the recipient strain.
The polysaccharide capsule represents the major virulencedeterminant of Streptococcus pneumoniae. Classic studies dem-onstrated a requirement for the capsule in virulence, estab-lished its role in antiphagocytosis, and showed that mutantsproducing reduced amounts of capsule were correspondinglyreduced in virulence and antiphagocytic ability (20, 32, 39).Although more than 80 distinct capsular serotypes have nowbeen described, a small number of these account for greaterthan 90% of pneumococcal infections in humans. In children,the prevalent types are 3, 4, 6, 14, 18, 19, and 23 (14), whereasin adults, types 1, 3, 4, 6, 7, 8, 9, 12, 14, and 23 prevail (12). Theresults of opsonophagocytosis (2, 13, 19), complement activa-tion and deposition (11, 15, 23, 31, 38), and mouse virulenceassays (4, 6, 19, 21, 36, 40) have frequently been found to differwith the type of capsule expressed. In a study of 76 S.pneumoniae strains representing eight capsular serotypes orgroups, the virulence of the isolates in terms of ability and timerequired to kill infected mice was strongly associated with thecapsule type of the invading strain (4). In addition, the abilityto protect against pneumococcal challenge in mice with anti-bodies to the cell surface components phosphocholine andpneumococcal surface protein A (PspA) differs with the invad-ing strain and may be affected by the capsular serotype (5, 25,27, 33, 40).The reasons why some capsular types are better able to
evade host defenses and cause disease are not clearly under-stood. These differences may relate to specific propertiesconferred by the different capsular polysaccharides or may bedue to noncapsular determinants. Past studies examining clin-ical and laboratory isolates in mouse virulence assays havesuggested a correlation between capsule type and virulence.However, these results were based on strains of differinggenetic backgrounds. To better understand the relationshipbetween capsular type and virulence in S. pneumoniae, we haveconstructed isogenic strains differing only in capsular type. Our
* Corresponding author. Mailing address: Department of Microbi-ology-661 BBRB Box 12, University of Alabama at Birmingham,Birmingham, AL 35294-2170. Phone: (205) 934-9531. Fax: (205)975-6715. Electronic mail address: [email protected].
initial focus has been specifically on the type 3 capsule. Wehave recently determined the genetic organization of thechromosomal region required for synthesis of this capsule type(10). These studies showed that the type 3-specific genes arelinked in the chromosome and that they can be transferred asa unit during transformation. By transferring the type 3 capsuleto strains representing different capsular types and geneticbackgrounds, we expect to be able to determine whether theobserved differences in virulence properties are due to the type3 capsule itself. Here we describe the construction, character-ization, and virulence in mice of derivatives of capsule types 2,5, and 6B which express the type 3 capsule.
MATERIALS AND METHODS
Bacterial strains and growth conditions. The S. pneumoniaestrains used in this study are described in Table 1. For routineculture, bacteria were grown at 37°C in Todd-Hewitt broth(THY; Difco Laboratories, Detroit, Mich.) supplemented with0.5% yeast extract (Difco) or at 37°C in 5% CO2 on blood agarmedium (Difco) containing 3% defibrinated sheep erythro-cytes. Chemically defined medium (CDM) (34) obtained fromJRH Biosciences (Denver, Pa.) was supplemented with 0.073%cysteine-HCl, 0.025% sodium bicarbonate, and 0.0005 or 2%choline, as indicated. Erythromycin was used at a concentra-tion of 0.3 ,ug/ml.
Transformations, serotyping, and restriction enzyme frag-ment patterns. Transformations were performed as describedpreviously (42). All transformants and parental strains wereserotyped with capsule type-specific antisera (Statens Seru-minstitut, Copenhagen, Denmark) in slide agglutination as-says. Genomic DNA, prepared as described elsewhere (10),was digested with Hindlll for 4 h at 37°C and electrophoresedovernight through 0.7% agarose in Tris-borate-EDTA buffer(22).
Analysis of PspA. Bacteria were grown in CDM containing2% choline, a condition that causes release of PspA into theculture medium (43). Filtered, unconcentrated supernatantfluids (20 ,LI) were electrophoresed in sodium dodecyl sulfate(SDS)-12% polyacrylamide gels. Western blotting (immuno-blotting) was performed by using a semidry electroblotter
1813
on May 26, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

1814 KELLY ET AL.
(Bio-Rad Laboratories, Richmond, Calif.), and the blots wereprocessed as described previously (41). The PspA-specificmonoclonal antibodies XiR278, Xi126, and 2A4 were kindlyprovided by Larry McDaniel (University of Alabama at Bir-mingham). Silver staining was performed by using the SilverStain Kit from Stratagene Cloning Systems, Inc. (La Jolla,Calif.).
Characterization of morphology and capsule production.For average chain length determinations, bacteria were grownin THY to an optical density at 600 nm (OD600) of -0.3. Chainlengths were determined microscopically by using a Petroff-Hauser counting chamber (Auther C. Thomas Co., Philadel-phia, Pa.). An average of five squares was counted for eachstrain. Comparisons of average chain lengths were determinedby using the two-sample rank test (44).The number of cells per colony was determined by using
bacteria grown on blood agar medium for 18 h at 37°C in 5%CO2. A plug containing a single colony was obtained with asterile Pasteur pipette and then resuspended in 50 [L of THY.Tenfold serial dilutions were performed in THY and plated onblood agar medium. Plates were incubated overnight at 37°C in5% C02, and the number of CFU per colony was calculated.
Buoyant density determinations were performed by usingbacteria grown on blood agar medium or in THY. Bacteriagrown on solid medium were harvested by washing each platewith water, centrifuging the suspension, and then resuspendingthe pellet to an OD600 of -0.4 with water. Ten-milliliter liquidcultures, grown to an OD600 of -0.5, were harvested bycentrifugation for 10 to 15 min at 8,000 to 16,000 x g. Bacteriawere washed twice with water prior to being loaded onto 10-ml,continuous, 0 to 50% Percoll (Pharmacia, Piscataway, N.J.)gradients. As standards, 5 ,ul of density marker beads (Phar-macia) ranging in size from 1.033 to 1.076 g/ml, were alsoloaded. Gradients were centrifuged for 30 min at 8,000 x gwith the brake off. A standard curve based on the migration ofthe marker beads was generated, and the density of thebacteria was determined by extrapolation.
For determinations of total capsule content, 1.5-ml culturesgrown in CDM containing 0.0005% choline were harvested bycentrifugation at 8,000 to 16,000 x g for 10 min. Culturesupernatant fluids were filtered and saved, and the cells wereresuspended in 500 ,ul of protoplast buffer (20% sucrose, 0.005M Tris [pH 7.4], 0.0025 M MgSO4). Cell sonicates wereproduced by three cycles of a 10-s pulse, followed by a 10-sincubation on ice, with a Fisher Sonic Dismembrator model300 (Fisher Biotechnology, St. Louis, Mo.) with the intensitycontrol set at 30. Culture supernatant fluids and cell sonicateswere stored at - 20°C.
For surface localization assays, 1.5-ml cultures grown to anOD600 of -0.5 were heat killed by incubation at 65°C for 30min. Bacteria were harvested by centrifugation, and culturesupernatant fluids were filtered and saved. After the pelletswere washed twice with phosphate-buffered saline (PBS; 137mM NaCl, 2.7 mM KCl, 4.3 mM Na2HPO4 * 7H20, 1.4 mMKH2PO4), the pellets were resuspended in 1.5 ml of THY.Samples were stored at 4°C.
Relative quantities of capsular material were determined byan enzyme-linked immunosorbent assay (ELISA). Wells ofpolyvinyl microtiter plates (Falcon; Becton Dickinson, Oxnard,Calif.) were coated overnight at 4°C with 50-pl culture super-natant fluids and equivalent amounts of sonicates (for totalcapsule content) or with serial dilutions of culture supernatantfluids and heat-killed cells (for surface localization). Wellswere rinsed with PBS plus 0.05% Tween (PBST) and thenblocked overnight at 4°C with 1% bovine serum albumin inPBST. Samples were incubated at room temperature for 1 h
with the type 3-specific monoclonal antibody 16.3 (3), kindlyprovided by David E. Briles (University of Alabama at Bir-mingham). Plates were washed twice with PBST and incubatedwith alkaline phosphatase-streptavidin and biotin-conjugatedgoat anti-mouse immunoglobulin for 45 min. Wells werewashed twice with PBST, and color development with p-nitrophenyl phosphate (Sigma Chemical Co., St. Louis, Mo.)was measured at OD405.
Virulence assays. The virulence of the type 3 derivatives wascompared with that of the parental strains in BALB/ByJfemale mice (Jackson Laboratory, Bar Harbor, Maine). Bac-teria were grown to the mid-log phase in THY. Samples werediluted serially in sterile lactated Ringer's solution, and 0.2 mlwas used to infect mice intraperitoneally (i.p.) or intravenously(i.v.), as indicated. Fifty percent lethal doses (LD50s) weredetermined by the method of Reed and Muench (28) andcompared by Fisher's exact test (44). Median times to deathwere analyzed by using the two-sample rank test (44). The Pvalues were determined by using a two-tailed table.
RESULTS
Construction of derivatives of types 2, 5, and 6B expressingthe type 3 capsule. As the donor for these studies, we chose ahighly virulent type 3 strain which, unlike the recipient strains,produces large mucoid colonies when grown on blood agarplates. Strains of types 2, 5, and 6B were used as recipients.These strains were chosen because they differ significantly fromthe type 3 strain in terms of capsule structure (Fig. 1) and mousevirulence. The type 2 and type 5 strains differ in virulence fromthe type 3 strain in terms of time required to cause death(shorter with type 2) and LD50 (lower with type 5) (5, 40; thisstudy). The type 6B strain is'relatively avirulent in mice (4).
In other studies, we have shown that all of the genesnecessary for expression of the type 3 capsule are contained ina linked locus (10). In those studies, strain JD770, a derivativeof the type 3 strain WU2 that contains a nondestructiveinsertion and an erythromycin resistance gene in the capsulelocus, was constructed. We found that by transforming the type2 strain D39 with JD770 DNA and selecting for erythromycinresistance, all of the isolates obtained expressed only the type3 capsule (10). On the basis of this observation, we trans-formed strains of capsular types 5 and 6B with JD770 DNAand selected for erythromycin-resistant transformants. Slideagglutination assays using type-specific antiserum for the do-nor and for each of the recipients indicated that greater than95% of all erythromycin-resistant transformants expressed thedonor type 3 capsule but not the capsule of the recipient strain.The remainder expressed the capsule type of the recipientparent only. To lessen the possibility that unlinked noncapsulargenes from the donor type 3 strain might affect our results, eachof two isolates obtained in independent transformations wasbackcrossed three times to the parent recipient strain to produceessentially isogenic strains. All constructs were then character-ized microscopically and with respect to restriction enzymefragment patterns, PspA expression, and capsule expressionprior to testing in a mouse model of pneumococcal virulence.
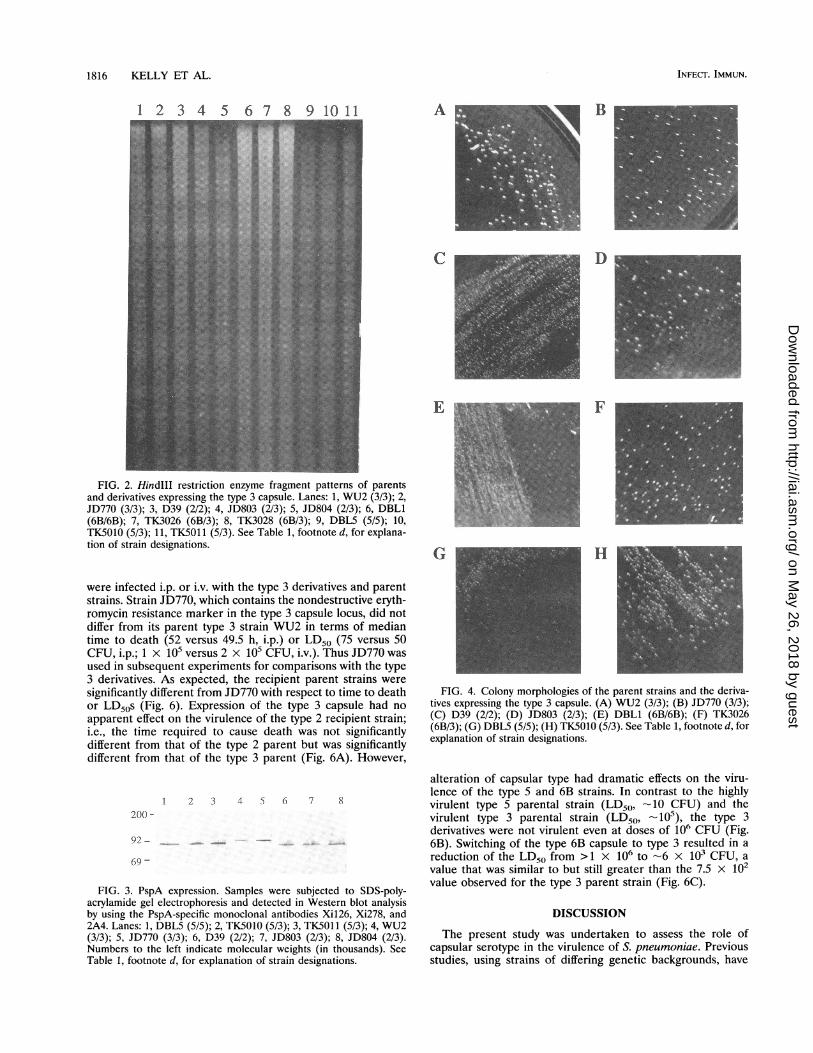
Restriction enzyme fragment patterns. HindlIl restrictionenzyme fragment patterns were used to confirm the similarityof the derivatives to the recipient parent strains. As can be seenin Fig. 2, each of the parent strains had a distinctive pattern,and the pattern for each of the type 3 derivatives of the type 2,5, and 6B strains was characteristic of its recipient parent.These results indicated that the transformations did not resultin any gross alterations in the genomic DNA and that we hadnot inadvertently reisolated the parent donor strain JD770.
INFECT. IMMUN.
on May 26, 2018 by guest
http://iai.asm.org/
Dow
nloaded from
CAPSULAR TYPE AND S. PNEUMONIAE VIRULENCE 1815
TABLE 1. Bacterial strains
Strain Derivation" Relevant phenotype Chain length' Reference
WU2 Clinical isolate Encapsulated type 3 2.3 ± 0.14 7JD770 pJD330c x WU2 Encapsulated type 3; Emr 2.4 ± 0.19 10D39 Clinical isolate Encapsulated type 2 6.2 ± 0.69 1JD803 (2/3)d JD770 x D39 Encapsulated type 3; Emr 6.6 ± 0.76 This studyJD804 (2/3) JD770 x D39 Encapsulated type 3; Emr 5.2 ± 0.74 This studyDBL5 Clinical isolate Encapsulated type 5 9.2 ± 0.80 40TK5010 (5/3) JD770 x DBL5 Encapsulated type 3; Emr 2.1 ± 0.08' This studyTK5011 (5/3) JD770 x DBL5 Encapsulated type 3; Emr 2.4 ± 0.13' This studyTK5012 (5/3) JD770 x DBL5 Encapsulated type 3; Emr NDf This studyDBL1 Clinical isolate Encapsulated type 6B 2.7 ± 0.11 4TK3026 (6B/3) JD770 x DBL1 Encapsulated type 3; Emr 2.4 ± 0.15 This studyTK3028 (6B/3) JD770 x DBLI Encapsulated type 3; Emr 3.1 ± 0.30 This study
"Crosses were done by transformation. Each derivative is the result of at least three backcrosses with the parental recipient strain.The numbers of cells per chain (means ± standard errors) were determined during this study by microscopic examination.
' pJD330 contains a Sau3AI fragment that is part of the type 3 capsule locus. It was constructed in pJY4163 (41) which contains an Em' marker. Insertion of pJD33()into the S. pneumoniae chromosome places the Emr marker in the type 3 locus but does not affect capsule expression (10)." For clarity, strains will be listed by strain number, genetic background, and capsule type expressed. For example, JD803 (2/3) represents a strain with a type 2 genetic
background expressing a type 3 capsule.' Significantly different from recipient parent strain (P < 0.001).f ND, not determined.
PspA expression. PspAs have been shown to differ in termsof molecular weight and expression of antigenic determinants(9, 26, 37). The PspAs of the donor and recipient strains usedin this study can be distinguished by molecular weight and/orreactivities with monoclonal antibodies. PspAs expressed bythe type 3 derivatives of the type 2 and type 5 strains wereshown in Western blot analysis to be identical in molecularweight to the recipient parent strain (Fig. 3). The PspA of thetype 6B strain is nonreactive with our panel of monoclonalantibodies, and its type 3 derivatives were also not detected inWestern blots. However, silver stain analysis revealed that themolecular sizes of the PspAs of these derivatives were identicalto that of the recipient parent and distinctive from that of thePspA of the type 3 donor (data not shown).Morphologic characterization and capsule production. Mi-
croscopic examination revealed that alteration of capsular typehad no effect on the chain length of the type 2 and type 6Bderivative strains. However, the chain lengths of the type 5derivatives differed significantly from that of the type 5 paren-tal strain and were almost identical to that of the type 3 parent(Table 1).
Morphologically, type 3 strains exhibit large mucoid capsuleswhen grown on blood agar plates, whereas type 2, 5, and 6B
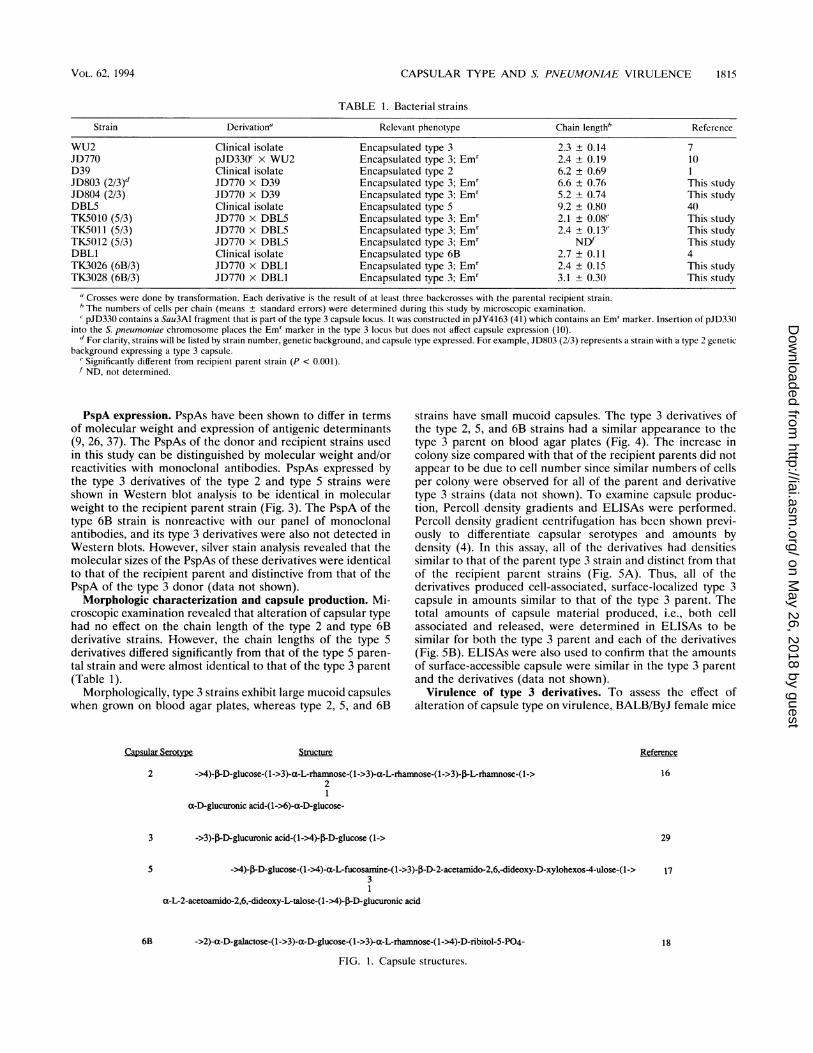
Capsular Serotype
strains have small mucoid capsules. The type 3 derivatives ofthe type 2, 5, and 6B strains had a similar appearance to thetype 3 parent on blood agar plates (Fig. 4). The increase incolony size compared with that of the recipient parents did notappear to be due to cell number since similar numbers of cellsper colony were observed for all of the parent and derivativetype 3 strains (data not shown). To examine capsule produc-tion, Percoll density gradients and ELISAs were performed.Percoll density gradient centrifugation has been shown previ-ously to differentiate capsular serotypes and amounts bydensity (4). In this assay, all of the derivatives had densitiessimilar to that of the parent type 3 strain and distinct from thatof the recipient parent strains (Fig. 5A). Thus, all of thederivatives produced cell-associated, surface-localized type 3capsule in amounts similar to that of the type 3 parent. Thetotal amounts of capsule material produced, i.e., both cellassociated and released, were determined in ELISAs to besimilar for both the type 3 parent and each of the derivatives(Fig. SB). ELISAs were also used to confirm that the amountsof surface-accessible capsule were similar in the type 3 parentand the derivatives (data not shown).
Virulence of type 3 derivatives. To assess the effect ofalteration of capsule type on virulence, BALB/ByJ female mice
Structure
2 ->4)-J3-D-glucose-(1->3)-a-L-rhamnose-(1->3)-a-L-rhamnose-(1->3)-J-L-rhamnose-(l->2
a-D-glucuronic acid-(1->6)-a-D-glucose-
3 ->3)-,B-D-glucuronic acid-(1->4)-flD-glucose (1->
5 ->4)-,lD-glucose-(1->4)-a-L-fucosamine-(1->3)-p-D-2-acetamido-2,6,-dideoxy-D-xylohexos4-ulose-(1->3
a-L-2-acetoamido-2,6,-dideoxy-L-talose-(1->4)-J3-D-glucuronic acid
6B ->2)-a-D-galactose-(1->3)-a-D-glucose-(1->3)-a-L-rhamnose-(1->4)-D-ribitol-5-P04-
FIG. 1. Capsule structures.
Reference
16
29
17
18
VOL. 62, 1994
on May 26, 2018 by guest
http://iai.asm.org/
Dow
nloaded from
1816 KELLY ET AL.
1 2 3 4 5 6 7 8 9 A
C
E
B
D
F
FIG. 2. HindIII restriction enzyme fragment patterns of parentsand derivatives expressing the type 3 capsule. Lanes: 1, WU2 (3/3); 2,JD770 (3/3); 3, D39 (2/2); 4, JD803 (2/3); 5, JD804 (2/3); 6, DBL1(6B/6B); 7, TK3026 (6B/3); 8, TK3028 (6B/3); 9, DBL5 (5/5); 10,TK5010 (5/3); 11, TK5011 (5/3). See Table 1, footnote d, for explana-tion of strain designations.
were infected i.p. or i.v. with the type 3 derivatives and parentstrains. Strain JD770, which contains the nondestructive eryth-romycin resistance marker in the type 3 capsule locus, did notdiffer from its parent type 3 strain WU2 in terms of mediantime to death (52 versus 49.5 h, i.p.) or LD50 (75 versus 50CFU, i.p.; 1 x 105 versus 2 x 105 CFU, i.v.). Thus JD770 wasused in subsequent experiments for comparisons with the type3 derivatives. As expected, the recipient parent strains weresignificantly different from JD770 with respect to time to deathor LD50s (Fig. 6). Expression of the type 3 capsule had noapparent effect on the virulence of the type 2 recipient strain;i.e., the time required to cause death was not significantlydifferent from that of the type 2 parent but was significantlydifferent from that of the type 3 parent (Fig. 6A). However,
1 2 3 4 5 6 7 8200 -
92-
69 -
FIG. 3. PspA expression. Samples were subjected to SDS-poly-acrylamide gel electrophoresis and detected in Western blot analysisby using the PspA-specific monoclonal antibodies Xi126, Xi278, and2A4. Lanes: 1, DBL5 (5/5); 2, TK5010 (5/3); 3, TK5011 (5/3); 4, WU2(3/3); 5, JD770 (3/3); 6, D39 (2/2); 7, JD803 (2/3); 8, JD804 (2/3).Numbers to the left indicate molecular weights (in thousands). SeeTable 1, footnote d, for explanation of strain designations.
G _H_
FIG. 4. Colony morphologies of the parent strains and the deriva-tives expressing the type 3 capsule. (A) WU2 (3/3); (B) JD770 (3/3);(C) D39 (2/2); (D) JD803 (2/3); (E) DBL1 (6B/6B); (F) TK3026(6B/3); (G) DBL5 (5/5); (H) TK5010 (5/3). See Table 1, footnote d, forexplanation of strain designations.
alteration of capsular type had dramatic effects on the viru-lence of the type 5 and 6B strains. In contrast to the highlyvirulent type 5 parental strain (LD50, -~10 CFU) and thevirulent type 3 parental strain (LD50, 105), the type 3derivatives were not virulent even at doses of 106 CFU (Fig.6B). Switching of the type 6B capsule to type 3 resulted in areduction of the LD50 from >1 x 106 to -6 x 103 CFU, avalue that was similar to but still greater than the 7.5 x 102value observed for the type 3 parent strain (Fig. 6C).
DISCUSSION
The present study was undertaken to assess the role ofcapsular serotype in the virulence of S. pneumoniae. Previousstudies, using strains of differing genetic backgrounds, have
INFECT. IMMUN.
on May 26, 2018 by guest
http://iai.asm.org/
Dow
nloaded from
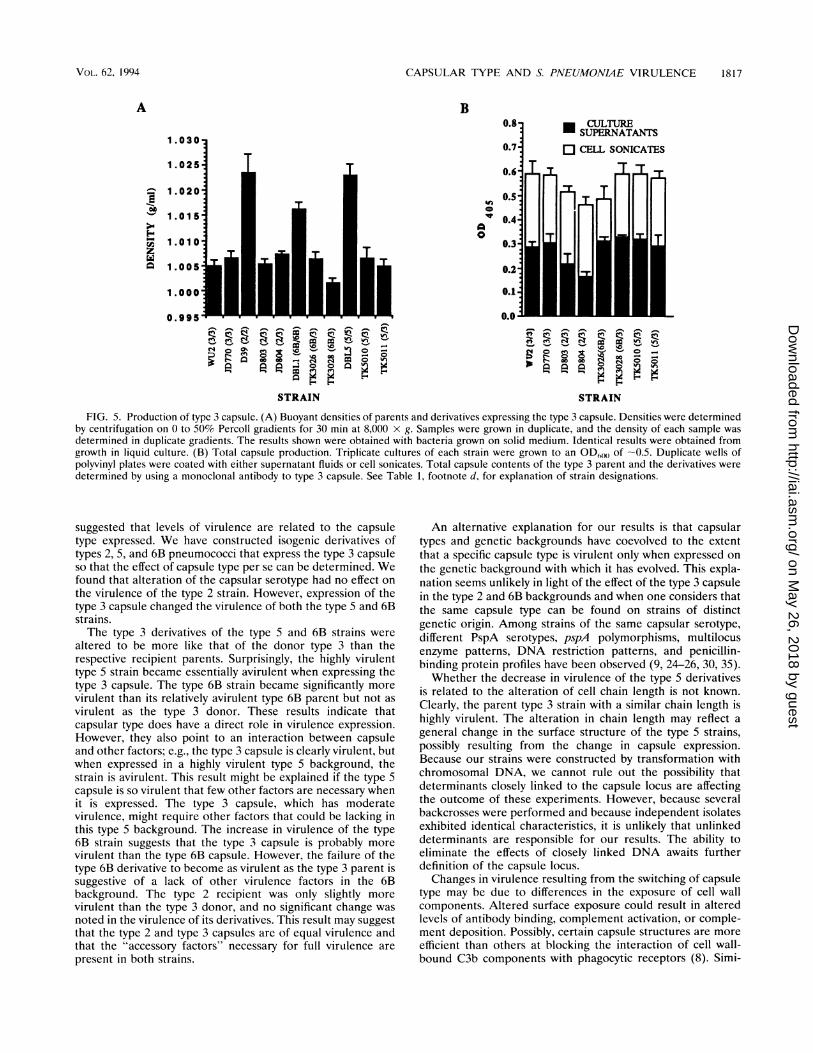
CAPSULAR TYPE AND S. PNEUMONME VIRULENCE 1817
A B
1
1I--
E-
z3
CDIV
a
0
1
1
1
1.
0.
TRaIN
STRAIN
,.,sZ -., e e
-,l-1
9 C 2 8 a . -
STRAIN
FIG. 5. Production of type 3 capsule. (A) Buoyant densities of parents and derivatives expressing the type 3 capsule. Densities were determinedby centrifugation on 0 to 50% Percoll gradients for 30 min at 8,000 x g. Samples were grown in duplicate, and the density of each sample wasdetermined in duplicate gradients. The results shown were obtained with bacteria grown on solid medium. Identical results were obtained fromgrowth in liquid culture. (B) Total capsule production. Triplicate cultures of each strain were grown to an OD(60 of -0.5. Duplicate wells ofpolyvinyl plates were coated with either supernatant fluids or cell sonicates. Total capsule contents of the type 3 parent and the derivatives weredetermined by using a monoclonal antibody to type 3 capsule. See Table 1, footnote d, for explanation of strain designations.
suggested that levels of virulence are related to the capsuletype expressed. We have constructed isogenic derivatives oftypes 2, 5, and 6B pneumococci that express the type 3 capsuleso that the effect of capsule type per se can be determined. Wefound that alteration of the capsular serotype had no effect onthe virulence of the type 2 strain. However, expression of thetype 3 capsule changed the virulence of both the type 5 and 6Bstrains.The type 3 derivatives of the type 5 and 6B strains were
altered to be more like that of the donor type 3 than therespective recipient parents. Surprisingly, the highly virulenttype 5 strain became essentially avirulent when expressing thetype 3 capsule. The type 6B strain became significantly morevirulent than its relatively avirulent type 6B parent but not asvirulent as the type 3 donor. These results indicate thatcapsular type does have a direct role in virulence expression.However, they also point to an interaction between capsuleand other factors; e.g., the type 3 capsule is clearly virulent, butwhen expressed in a highly virulent type 5 background, thestrain is avirulent. This result might be explained if the type 5capsule is so virulent that few other factors are necessary whenit is expressed. The type 3 capsule, which has moderatevirulence, might require other factors that could be lacking inthis type 5 background. The increase in virulence of the type6B strain suggests that the type 3 capsule is probably morevirulent than the type 6B capsule. However, the failure of thetype 6B derivative to become as virulent as the type 3 parent issuggestive of a lack of other virulence factors in the 6Bbackground. The type 2 recipient was only slightly morevirulent than the type 3 donor, and no significant change wasnoted in the virulence of its derivatives. This result may suggestthat the type 2 and type 3 capsules are of equal virulence andthat the "accessory factors" necessary for full virulence arepresent in both strains.
An alternative explanation for our results is that capsulartypes and genetic backgrounds have coevolved to the extentthat a specific capsule type is virulent only when expressed onthe genetic background with which it has evolved. This expla-nation seems unlikely in light of the effect of the type 3 capsulein the type 2 and 6B backgrounds and when one considers thatthe same capsule type can be found on strains of distinctgenetic origin. Among strains of the same capsular serotype,different PspA serotypes, pspA polymorphisms, multilocusenzyme patterns, DNA restriction patterns, and penicillin-binding protein profiles have been observed (9, 24-26, 30, 35).Whether the decrease in virulence of the type 5 derivatives
is related to the alteration of cell chain length is not known.Clearly, the parent type 3 strain with a similar chain length ishighly virulent. The alteration in chain length may reflect ageneral change in the surface structure of the type 5 strains,possibly resulting from the change in capsule expression.Because our strains were constructed by transformation withchromosomal DNA, we cannot rule out the possibility thatdeterminants closely linked to the capsule locus are affectingthe outcome of these experiments. However, because severalbackcrosses were performed and because independent isolatesexhibited identical characteristics, it is unlikely that unlinkeddeterminants are responsible for our results. The ability toeliminate the effects of closely linked DNA awaits furtherdefinition of the capsule locus.Changes in virulence resulting from the switching of capsule
type may be due to differences in the exposure of cell wallcomponents. Altered surface exposure could result in alteredlevels of antibody binding, complement activation, or comple-ment deposition. Possibly, certain capsule structures are moreefficient than others at blocking the interaction of cell wall-bound C3b components with phagocytic receptors (8). Simi-
VOL. 62, 1994
on May 26, 2018 by guest
http://iai.asm.org/
Dow
nloaded from
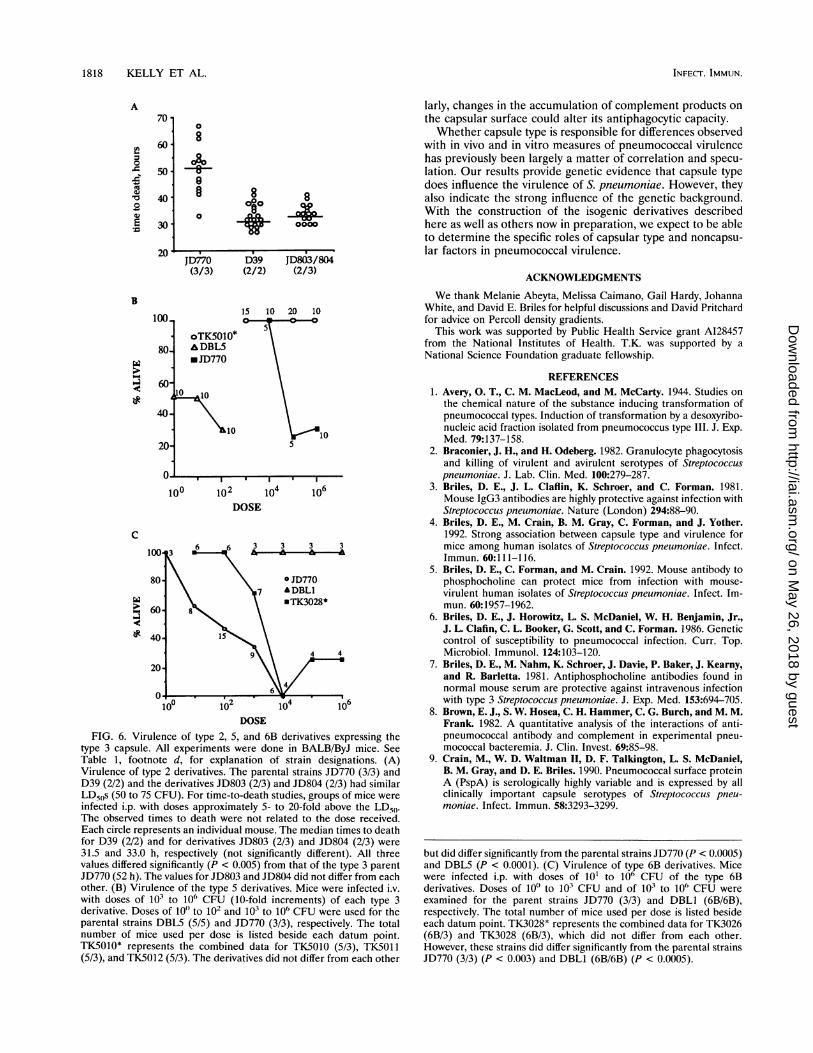
1818 KELLY ET AL.
A70 -
LI6000
X 400
.EwF- 30
20
B
100-
80-
wJ60-
40-
20-
n
0
8
-8-8
§0
8 8
0000
JD770 D39 JD80)3/804(3/3) (2/2) (2/3)
15 10 20 10
oTK5010*A&DBL5.1D770
10
10
100 102D(
I, I I
104 106OSE
C
'2 6.
10 10 10 10
DOSEFIG. 6. Virulence of type 2, 5, and 6B derivatives expressing the
type 3 capsule. All experiments were done in BALB/ByJ mice. SeeTable 1, footnote d, for explanation of strain designations. (A)Virulence of type 2 derivatives. The parental strains JD770 (3/3) andD39 (2/2) and the derivatives JD803 (2/3) and JD804 (2/3) had similarLD50s (50 to 75 CFU). For time-to-death studies, groups of mice wereinfected i.p. with doses approximately 5- to 20-fold above the LD50.The observed times to death were not related to the dose received.Each circle represents an individual mouse. The median times to deathfor D39 (2/2) and for derivatives JD803 (2/3) and JD804 (2/3) were31.5 and 33.0 h, respectively (not significantly different). All threevalues differed significantly (P < 0.005) from that of the type 3 parentJD770 (52 h). The values for JD803 and JD804 did not differ from eachother. (B) Virulence of the type 5 derivatives. Mice were infected i.v.with doses of 103 to 106 CFU (10-fold increments) of each type 3derivative. Doses of 100 to 102 and 103 to 106 CFU were used for theparental strains DBLS (5/5) and JD770 (3/3), respectively. The totalnumber of mice used per dose is listed beside each datum point.TKS010* represents the combined data for TKS010 (5/3), TKS011(5/3), and TKS012 (5/3). The derivatives did not differ from each other
larly, changes in the accumulation of complement products onthe capsular surface could alter its antiphagocytic capacity.Whether capsule type is responsible for differences observed
with in vivo and in vitro measures of pneumococcal virulencehas previously been largely a matter of correlation and specu-lation. Our results provide genetic evidence that capsule typedoes influence the virulence of S. pneumoniae. However, theyalso indicate the strong influence of the genetic background.With the construction of the isogenic derivatives describedhere as well as others now in preparation, we expect to be ableto determine the specific roles of capsular type and noncapsu-lar factors in pneumococcal virulence.
ACKNOWLEDGMENTS
We thank Melanie Abeyta, Melissa Caimano, Gail Hardy, JohannaWhite, and David E. Briles for helpful discussions and David Pritchardfor advice on Percoll density gradients.
This work was supported by Public Health Service grant A128457from the National Institutes of Health. T.K. was supported by aNational Science Foundation graduate fellowship.
REFERENCES1. Avery, 0. T., C. M. MacLeod, and M. McCarty. 1944. Studies on
the chemical nature of the substance inducing transformation ofpneumococcal types. Induction of transformation by a desoxyribo-nucleic acid fraction isolated from pneumococcus type III. J. Exp.Med. 79:137-158.
2. Braconier, J. H., and H. Odeberg. 1982. Granulocyte phagocytosisand killing of virulent and avirulent serotypes of Streptococcuspneumoniae. J. Lab. Clin. Med. 100:279-287.
3. Briles, D. E., J. L. Claflin, K. Schroer, and C. Forman. 1981.Mouse IgG3 antibodies are highly protective against infection withStreptococcus pneumoniae. Nature (London) 294:88-90.
4. Briles, D. E., M. Crain, B. M. Gray, C. Forman, and J. Yother.1992. Strong association between capsule type and virulence formice among human isolates of Streptococcus pneumoniae. Infect.Immun. 60:111-116.
5. Briles, D. E., C. Forman, and M. Crain. 1992. Mouse antibody tophosphocholine can protect mice from infection with mouse-virulent human isolates of Streptococcus pneumoniae. Infect. Im-mun. 60:1957-1962.
6. Briles, D. E., J. Horowitz, L. S. McDaniel, W. H. Benjamin, Jr.,J. L. Clafin, C. L. Booker, G. Scott, and C. Forman. 1986. Geneticcontrol of susceptibility to pneumococcal infection. Curr. Top.Microbiol. Immunol. 124:103-120.
7. Briles, D. E., M. Nahm, K. Schroer, J. Davie, P. Baker, J. Kearny,and R. Barletta. 1981. Antiphosphocholine antibodies found innormal mouse serum are protective against intravenous infectionwith type 3 Streptococcus pneumoniae. J. Exp. Med. 153:694-705.
8. Brown, E. J., S. W. Hosea, C. H. Hammer, C. G. Burch, and M. M.Frank. 1982. A quantitative analysis of the interactions of anti-pneumococcal antibody and complement in experimental pneu-mococcal bacteremia. J. Clin. Invest. 69:85-98.
9. Crain, M., W. D. Waltman II, D. F. Talkington, L. S. McDaniel,B. M. Gray, and D. E. Briles. 1990. Pneumococcal surface proteinA (PspA) is serologically highly variable and is expressed by allclinically important capsule serotypes of Streptococcus pneu-moniae. Infect. Immun. 58:3293-3299.
but did differ significantly from the parental strains JD770 (P < 0.0005)and DBLS (P < 0.0001). (C) Virulence of type 6B derivatives. Micewere infected i.p. with doses of 101 to 106 CFU of the type 6Bderivatives. Doses of 100 to 103 CFU and of 103 to 106 CFU wereexamined for the parent strains JD770 (3/3) and DBLI (6B/6B),respectively. The total number of mice used per dose is listed besideeach datum point. TK3028* represents the combined data for TK3026(6B/3) and TK3028 (6B/3), which did not differ from each other.However, these strains did differ significantly from the parental strainsJD770 (3/3) (P < 0.003) and DBL1 (6B/6B) (P < 0.0005).
INFECT. IMMUN.
on May 26, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

CAPSULAR TYPE AND S. PNEUMONIAE VIRULENCE 1819
10. Dillard, J., and J. Yother. Mol. Microbiol., in press.11. Fine, D. P. 1975. Pneumococcal type associated variability in
alternative complement pathway activation. Infect. Immun. 12:772-778.
12. Finland, M., and M. Barnes. 1977. Changes in occurrence ofcapsular serotypes of Streptococcus pneumoniae at Boston CityHospital during selected years between 1935 and 1974. J. Clin.Microbiol. 5:154-166.
13. Giebink, G. S., J. Verhoef, P. K. Peterson, and P. G. Quie. 1977.Opsonic requiremcnts for phagocytosis of Streptococcus pneu-moniae types VI, XVIII, XXIII, and XXV. Infect. Immun. 18:291-297.
14. Gray, B. M., and H. C. Dillon, Jr. 1986. Clinical and epidemiologicstudies of pneumococcal infection in children. Pediatr. Infect. Dis.J. 5:201-207.
15. Hostetter, M. K. 1986. Serotypic variation among virulent pneu-mococci in deposition and degradation of covalently bound C3b:implication for phagocytosis and antibody production. J. Infect.Dis. 153:682-693.
16. Jansson, P. E., B. Lindberg, M. Anderson, U. Lindquist, and J.Henrichsen. 1988. Structural studies of the capsular polysaccha-ride from Streptococcus pneurnoniae type 2, a reinvestigation.Carbohydr. Res. 182:111-117.
17. Jansson, P. E., B. Lindberg, and U. Lindquist. 1985. Structuralstudies of the capsular polysaccharide from Streptococcus pneu-moniae type 5. Carbohydr. Res. 140:101-110.
18. Kenne, L., B. Lindberg, and J. K. Madden. 1979. Structural studiesof the capsular antigen from Streptococcus pneumoniae type 26.Carbohydr. Res. 184:288-291.
19. Knecht, J. D., G. Schiffman, and R. Austrian. 1970. Some biolog-ical properties of pneumococcus serotype 37 and the chemistry ofits capsular polysaccharide. J. Exp. Med. 132:475-487.
20. Kruse, W., and S. Pansini. 1891. Untersuchungen uber denDiplococcus pneumoniae und verwandte Streptokoken. Z. Hyg.Infektionskr. 11:279-280.
21. MacLeod, C. M. 1965. Pneumococci, p. 391-411. In R. J. Duboseand J. G. Hirsh (ed.), Bacterial and mycotic infections of man. J. B.Lippincott Co., Philadelphia.
22. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecularcloning: a laboratory manual. Cold Spring Harbor Laboratory,Cold Spring Harbor, N.Y.
23. Matthay, K. K., W. C. Mentzer, D. W. Wara, H. K. Preisler, N. B.Lameris, and A. J. Ammann. 1981. Evaluation of opsonic require-ments for phagocytosis of Streptococcus pneumoniae serotypes VII,XIV, and XIX by chemiluminescence assay. Infect. Immun. 31:228-235.
24. McDaniel, L. S., and D. E. Briles. 1993. Molecular localization ofvariable and conserved regions of pspA, and identification ofadditional pspA homologous sequences in Streptococcus pneu-moniae. Microb. Pathog. 13:261-269.
25. McDaniel, L. S., G. Scott, J. F. Kearney, and D. E. Briles. 1984.Monoclonal antibodies against protease sensitive pneumococcalantigens can protect mice from fatal infection with Streptococcuspneumoniae. J. Exp. Med. 160:386-397.
26. McDaniel, L. S., G. Scott, K. Widenhofer, J. Carroll, and D. E.Briles. 1986. Analysis of surface protein of Streptococcus pneu-moniae recognized by protective monoclonal antibodies. Microb.Pathog. 1:519-531.
27. McDaniel, L. S., J. S. Sheffield, P. Delucchi, and D. E. Briles. 1991.PspA, a surface protein of Streptococcus pneurnoniae, is capable ofeliciting protection against pneumococci of more than one capsu-lar type. Infect. Immun. 59:222-228.
28. Reed, L. J., and H. Muench. 1938. A simple method of estimatingfifty percent endpoints. Am. J. Hyg. 27:493-497.
29. Reeves, R. E., and W. F. Goebel. 1941. Chemoimmunologicalstudies in the soluble specific substance of pneumococcus. V. Thestructure of the type III polysaccharide. J. Biol. Chem. 139:511-519.
30. Sibold, C., J. Wang, J. Henrichsen, and R. Hakenbeck. 1992.Genetic relationships of penicillin-susceptible and -resistant Strep-tococcus pneumoniae strains isolated on different continents. In-fect. Immun. 60:119-126.
31. Stephens, C. G., R. C. Williams, and W. P. Reed. 1977. Classicaland alternative complement pathway activation by pneumococci.Infect. Immun. 17:296-302.
32. Stryker, L. M. 1916. Variations in the pneumococcus induced bygrowth in immune serum. J. Exp. Med. 24:49-68.
33. Szu, S. C., S. Clarke, and J. B. Robbins. 1983. Protection againstpneumococcal infection in mice conferred by phosphocholine-binding antibodies: specificity of the phosphocholine binding andrelation to several types. Infect. Immun. 39:993-999.
34. van de Rijn, I., and R. E. Kessler. 1980. Growth characteristics ofgroup A streptococci in a new chemically defined medium. Infect.Immun. 27:444-448.
35. Viering, T. P., and D. P. Fine. 1989. Genetic analysis of Strepto-coccus pneumoniae serotypes with the use of DNA fingerprinting.J. Infect. Dis. 160:76-82.
36. Walter, A. W., V. H. Guerin, M. W. Beattie, H. Y. Cotler, and H. G.Bucca. 1941. Extension of the separation of types among pneumo-cocci: description of 17 types in addition to types I to 32 (Cooper).J. Immunol. 41:279.
37. Waltman, W. D., II, L. S. McDaniel, and D. E. Briles. 1990.Variation in the molecular weight of PspA (pneumococcal surfaceprotein A) among Streptococclus pneumoniae. Microb. Pathog.8:61-69.
38. Winkelstein, J. A., A. S. Abramaritz, and A. Tomasz. 1980.Activation of C3 via the alternative complement pathway results infixation of C3b to the pneumococcal cell wall. J. Immunol.124:2502-2506.
39. Wood, W. B., and M. R. Smith. 1949. Inhibition of surfacephagocytosis by the capsular "slime layer" of pneumococcus typeIII. J. Exp. Med. 90:85-99.
40. Yother, J., C. Forman, B. M. Gray, and D. E. Briles. 1982.Protection of mice with Streptococcus pneumoniae by anti-phos-phocholine antibody. Infect. Immun. 36:184-188.
41. Yother, J., G. L. Handsome, and D. E. Briles. 1992. Truncatedforms of PspA that are secreted from Streptococcus pneumoniaeand their use in functional studies and cloning of the pspA gene. J.Bacteriol. 174:610-618.
42. Yother, J., L. S. McDaniel, and D. E. Briles. 1986. Transformationof encapsulated Streptococcus pneumoniae. J. Bacteriol. 168:1463-1465.
43. Yother, J., and J. M. White. Submitted for publication.44. Zar, J. H. 1984. Biostatistical analysis, p. 718. Prentice Hall, Inc.,
Englewood Cliffs, N.J.
VOL. 62? 1994
on May 26, 2018 by guest
http://iai.asm.org/
Dow
nloaded from