Embed Size (px)
Citation preview

8/14/2019 Effect of Hibernation and Reproductive Status on Body Mass and Condition of Coastal Brown Bears
http://slidepdf.com/reader/full/effect-of-hibernation-and-reproductive-status-on-body-mass-and-condition-of 1/7
Effect of Hibernation and Reproductive Status on Body Mass and Condition of Coastal BrownBearsAuthor(s): Grant V. Hilderbrand, Charles C. Schwartz, Charles T. Robbins, Thomas A. HanleySource: The Journal of Wildlife Management, Vol. 64, No. 1 (Jan., 2000), pp. 178-183Published by: Allen PressStable URL: http://www.jstor.org/stable/3802988Accessed: 17/09/2009 14:50
Your use of the JSTOR archive indicates your acceptance of JSTOR's Terms and Conditions of Use, available athttp://www.jstor.org/page/info/about/policies/terms.jsp . JSTOR's Terms and Conditions of Use provides, in part, that unlessyou have obtained prior permission, you may not download an entire issue of a journal or multiple copies of articles, and youmay use content in the JSTOR archive only for your personal, non-commercial use.
Please contact the publisher regarding any further use of this work. Publisher contact information may be obtained athttp://www.jstor.org/action/showPublisher?publisherCode=acg .
Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printedpage of such transmission.
JSTOR is a not-for-profit organization founded in 1995 to build trusted digital archives for scholarship. We work with the
scholarly community to preserve their work and the materials they rely upon, and to build a common research platform thatpromotes the discovery and use of these resources. For more information about JSTOR, please contact [email protected].
Allen Press is collaborating with JSTOR to digitize, preserve and extend access to The Journal of Wildlife Management.
http://www.jstor.org

8/14/2019 Effect of Hibernation and Reproductive Status on Body Mass and Condition of Coastal Brown Bears
http://slidepdf.com/reader/full/effect-of-hibernation-and-reproductive-status-on-body-mass-and-condition-of 2/7
178 WINTER RANGE SHIFT BY CARIBOU Ferguson and Messier78 WINTER RANGE SHIFT BY CARIBOU Ferguson and Messier
parametric statistics for the behavioral sciences.Second edition. McGraw-Hill, New York, NewYork, USA.
SKOOG, R. 0. 1968. Ecology of the caribou (Rangifertarandus granti) in Alaska. Dissertation, Univer-sity of California, Berkeley, California, USA.
THOMAS, D. C. 1982. The relationship between fer-tility and fat reserves of Peary caribou. CanadianJournal of Zoology 60:597-602.
TYLER, N. J. C. 1987. Natural limitation of the abun-dance of the High Arctic Svalbard reindeer. Dis-sertation, University of Cambridge, Cambridge,United Kingdom.
parametric statistics for the behavioral sciences.Second edition. McGraw-Hill, New York, NewYork, USA.
SKOOG, R. 0. 1968. Ecology of the caribou (Rangifertarandus granti) in Alaska. Dissertation, Univer-sity of California, Berkeley, California, USA.
THOMAS, D. C. 1982. The relationship between fer-tility and fat reserves of Peary caribou. CanadianJournal of Zoology 60:597-602.
TYLER, N. J. C. 1987. Natural limitation of the abun-dance of the High Arctic Svalbard reindeer. Dis-sertation, University of Cambridge, Cambridge,United Kingdom.
WELLS, J. V, AND M. E. RICHMOND. 995. Popula-tions, metapopulations, and species populations:What are they and who should care? Wildlife So-ciety Bulletin 23:458-462.
ZAR, J. H. 1984. Biostatistical analysis. Second edition.Prentice-Hall, Englewood Cliffs, New Jersey,USA.
Received 11 December 1998.Accepted 11 May 1999.Associate Editor: Merrill.
WELLS, J. V, AND M. E. RICHMOND. 995. Popula-tions, metapopulations, and species populations:What are they and who should care? Wildlife So-ciety Bulletin 23:458-462.
ZAR, J. H. 1984. Biostatistical analysis. Second edition.Prentice-Hall, Englewood Cliffs, New Jersey,USA.
Received 11 December 1998.Accepted 11 May 1999.Associate Editor: Merrill.
EFFECT OF HIBERNATION ND REPRODUCTIVE TATUS ON BODYMASS AND CONDITION OF COASTAL BROWN BEARS
GRANT V. HILDERBRAND, laska Department f Fish and Game, 333 Raspberry Road, Anchorage, AK 99518, USACHARLES C. SCHWARTZ,1 laska Department f Fish and Game, 34828 Kalifornsky each Road, Suite B, Soldotna, AK
99669, USACHARLES . ROBBINS, Departments f Natural Resource Sciences and Zoology, Washington State University, ullman, WA
99164, USATHOMAS A. HANLEY, nited States Forest Service, Pacific Northwest Research Station, 2700 Sherwood Lane, Suite 2A,
Juneau, AK 99801, USA
Abstract: We investigated the effect of hibernation and reproductive status on changes in body mass andcomposition of adult female brown bears (Ursus arctos) on the Kenai Peninsula, Alaska. This information is
fundamental to understanding nutritional ecology of wild brown bear populations. Six adult females handledin the fall and following spring (paired samples) lost 73 + 22 kg (x + SD; 32 ? 10%) of fall body mass over208 ? 19 days. Of this mass loss, 56 ? 22% (55 ? 22 kg) was lipid and 44 ? 22% (43 ? 21 kg) was leanbody mass. Catabolism of lipid stores accounted for 88.4 ? 8.1% of the body energy used to meet maintenancedemands. Overwinter differences in body composition of adult females assessed only once in either the fall (n= 21) or spring (n = 32) were similar to those of paired samples. Relative fatness of bears entering the denwas positively related to the contribution of fat (%) to body mass (P < 0.01) and body energy (P < 0.01) lossesduring hibernation. Thus, relative fatness at the onset of fasting influences the relative proportion of lipid storesand lean body mass catabolized to meet protein and energy demands during hibernation. In the spring, lonefemales had greater body and lean masses than females with cubs of the year or yearlings. Lipid content wasgreatest in lone females in the fall. Studies using body mass and composition as indices of population healthshould consider season or reproductive class.
JOURNAL OF WILDLIFE MANAGEMENT 64(1):178-183
Key words: body composition, body condition, brown bear, hibernation, Kenai Peninsula, reproductive class,Ursus arctos.
EFFECT OF HIBERNATION ND REPRODUCTIVE TATUS ON BODYMASS AND CONDITION OF COASTAL BROWN BEARS
GRANT V. HILDERBRAND, laska Department f Fish and Game, 333 Raspberry Road, Anchorage, AK 99518, USACHARLES C. SCHWARTZ,1 laska Department f Fish and Game, 34828 Kalifornsky each Road, Suite B, Soldotna, AK
99669, USACHARLES . ROBBINS, Departments f Natural Resource Sciences and Zoology, Washington State University, ullman, WA
99164, USATHOMAS A. HANLEY, nited States Forest Service, Pacific Northwest Research Station, 2700 Sherwood Lane, Suite 2A,
Juneau, AK 99801, USA
Abstract: We investigated the effect of hibernation and reproductive status on changes in body mass andcomposition of adult female brown bears (Ursus arctos) on the Kenai Peninsula, Alaska. This information is
fundamental to understanding nutritional ecology of wild brown bear populations. Six adult females handledin the fall and following spring (paired samples) lost 73 + 22 kg (x + SD; 32 ? 10%) of fall body mass over208 ? 19 days. Of this mass loss, 56 ? 22% (55 ? 22 kg) was lipid and 44 ? 22% (43 ? 21 kg) was leanbody mass. Catabolism of lipid stores accounted for 88.4 ? 8.1% of the body energy used to meet maintenancedemands. Overwinter differences in body composition of adult females assessed only once in either the fall (n= 21) or spring (n = 32) were similar to those of paired samples. Relative fatness of bears entering the denwas positively related to the contribution of fat (%) to body mass (P < 0.01) and body energy (P < 0.01) lossesduring hibernation. Thus, relative fatness at the onset of fasting influences the relative proportion of lipid storesand lean body mass catabolized to meet protein and energy demands during hibernation. In the spring, lonefemales had greater body and lean masses than females with cubs of the year or yearlings. Lipid content wasgreatest in lone females in the fall. Studies using body mass and composition as indices of population healthshould consider season or reproductive class.
JOURNAL OF WILDLIFE MANAGEMENT 64(1):178-183
Key words: body composition, body condition, brown bear, hibernation, Kenai Peninsula, reproductive class,Ursus arctos.
Knowledge of seasonal cianges in body massand composition provides important insightsinto the nutritional ecology of bears. Femalebrown bears fast during hibernation, give birthduring this winter dormancy, and lactate for 2to 3 months before den
emergence (Farley andRobbins 1995). Additionally, the cubs may stay
1 Present Address: Interagency Grizzly Bear StudyTeam, Forestry Sciences Laboratory, Montana StateUniversity, Bozeman, MT 59717, USA.
Knowledge of seasonal cianges in body massand composition provides important insightsinto the nutritional ecology of bears. Femalebrown bears fast during hibernation, give birthduring this winter dormancy, and lactate for 2to 3 months before den
emergence (Farley andRobbins 1995). Additionally, the cubs may stay
1 Present Address: Interagency Grizzly Bear StudyTeam, Forestry Sciences Laboratory, Montana StateUniversity, Bozeman, MT 59717, USA.
with the mother, who continues to lactate, forup to 3 years. These life-history traits create sig-nificant energy demands for the female. Femalebody mass and body fat content are positivelyrelated to reproductive success of individualsand, thus, to
parametersof
population produc-tivity such as litter size, interval between litters,and age of first reproduction (Rogers 1976,1987; Bunnell and Tait 1981; Blanchard 1987;Stringham 1990; Noyce and Garshelis 1994; At-kinson and Ramsay 1995; Samson and Huot
with the mother, who continues to lactate, forup to 3 years. These life-history traits create sig-nificant energy demands for the female. Femalebody mass and body fat content are positivelyrelated to reproductive success of individualsand, thus, to
parametersof
population produc-tivity such as litter size, interval between litters,and age of first reproduction (Rogers 1976,1987; Bunnell and Tait 1981; Blanchard 1987;Stringham 1990; Noyce and Garshelis 1994; At-kinson and Ramsay 1995; Samson and Huot
I
J. Wildl. Manage. 64(1):2000. Wildl. Manage. 64(1):2000

8/14/2019 Effect of Hibernation and Reproductive Status on Body Mass and Condition of Coastal Brown Bears
http://slidepdf.com/reader/full/effect-of-hibernation-and-reproductive-status-on-body-mass-and-condition-of 3/7
J. Wildl. Manage. 64(1):2000 BODY MASS AND CONDITION OF BROWN BEARS * Hilderbrand et al.
1995; Hilderbrand et al. 1999). Quantificationof the impact of hibernation and reproductiveclass on body mass and composition, specifically
lipidstores and lean
bodymass, is important n
understanding population productivity.The impacts of hibernation and cub produc-
tion on body mass and composition have beenquantified for brown bears in captivity (Wattsand Jonkel 1988, Farley and Robbins 1995, Bar-boza et al. 1997) but not in the wild. Wild polarbears lose lipids and lean body mass while fast-ing, and the proportional oss of these 2 bodycomponents s influenced by the relative atnessof the bear at the onset of the fasting period(Atkinson et al. 1996). Body composition alsodiffers across reproductive classes in wild polarbears (Atkinson and Ramsay 1995). The objec-tives of this study were to quantify changes inbody mass and composition during the periodof winter dormancy and assess the effect of re-productive status on seasonal body mass andcomposition in wild, adult female brown bears.
METHODS
During 1996-98, adult female brown bearson the Kenai Peninsula, Alaska -60?N, 1500W)
were located with fixed-winged aircraft andanesthetized (Telazol, Fort Dodge AnimalHealth, Fort Dodge, Iowa, USA; 5-10 mg/kgbody mass; Taylor et al. 1989) by darting froma helicopter in the early spring (24 Apr-1 Jun)just after den emergence, and in the fall (6-27Oct) just before den entry. The time of capturewas typically within a few days of den entry orden exit; however, in 2 cases, bears remainedoutside the den for 7 to 14 days after capturein the fall. Bears were categorized as lone fe-
males, females with cubs of the year, femaleswith yearlings, or females with >2-year-old off-spring based on the age of the offspring accom-panying her at the time of capture or duringsubsequent aerial surveys.
At each capture, bears were weighed with anelectronic load cell (?0.2 kg) and body com-position was determined by bioelectrical mped-ance analysis (BIA, Model 101A, RJL Systems,Detroit, Michigan, USA) and-or isotopic dilu-tion (Farley and Robbins 1994, Hilderbrand etal.
1998).Both methods estimate
bodywater
content, which is inversely proportional o fatcontent (Farley and Robbins 1994). Thus, mea-sures of water content by BIA and dilution al-low accurate estimation of body fat content(Standard error <3.3%; Farley and Robbins
1994, Hilderbrand t al. 1998). The mean of the2 methods is the most accurate predictor ofbody composition (Hilderbrand et al. 1998).However, because isotopic dilution requires 1.5to 2.5 hrs to complete, dilutions were only per-formed on 15 of the bears in the current study.Bioelectrical impedance analysis was also per-formed on these 15 bears and the mean of the2 methods was used in analyses. Bioelectricalimpedance analysis measures alone were usedfor an additional 38 bears. The decision ofwhether or not to perform a dilution was basedon field logistics and not skewed by reproduc-tive class or season. Gut fill may produce erro-neous measures using BIA or isotopic dilution(Farley and Robbins 1994). However, the effectof gut fill on body composition estimates has notbeen quantified. Additionally, he bears in thecurrent study were likely already fasting due totemporal proximity to den entry or exit. Thecontribution of fat to losses of body energy dur-ing the hibernation period was estimated ac-cording to Atkinson and Ramsay (1995). Rela-tive fatness was calculated as the proportion offat to lean body mass in the fall (Atkinson et al.1996).
Estimates of body fat and lean body mass byBIA and isotopic dilution were compared usingpaired t-tests. Changes in body compositionduring hibernation, ength of time between cap-tures, proportional energy loss due to fat, rela-tive fatness, and seasonal body mass and com-position by reproductive status were calculatedand reported as means (?SD). The relation-ships between relative fatness n the fall and theproportional contribution of fat to loss of bodymass and body energy were assessed by power
regression analysis. Data were pooled acrossyears and seasonal body composition was com-pared across reproductive lass using analysis ofvariance (ANOVA) and Fisher's east significantdifference (LSD) analyses (Zar 1984).
RESULTS
Body fat and lean body mass estimates byBIA and isotopic dilution did not differ acrossmethods (t = 1.23, df = 14, P = 0.24). Six adultfemales that were handled during the fall andfollowing spring lost an average of 73 ? 22 kg(32 ? 10%) of fall body mass. Mass loss con-sisted of body fat (56 ? 22%) and lean bodymass (44 ? 22%). Catabolism of lipid stores ac-counted for 88.4 ? 8.1% (range of 77 to 98%)of the body energy used to meet maintenance
179
8/14/2019 Effect of Hibernation and Reproductive Status on Body Mass and Condition of Coastal Brown Bears
http://slidepdf.com/reader/full/effect-of-hibernation-and-reproductive-status-on-body-mass-and-condition-of 4/7
180 BODY MASS AND CONDITION OF BROWN BEARS Hilderbrand et al. J. Wildl. Manage. 64(1):2000
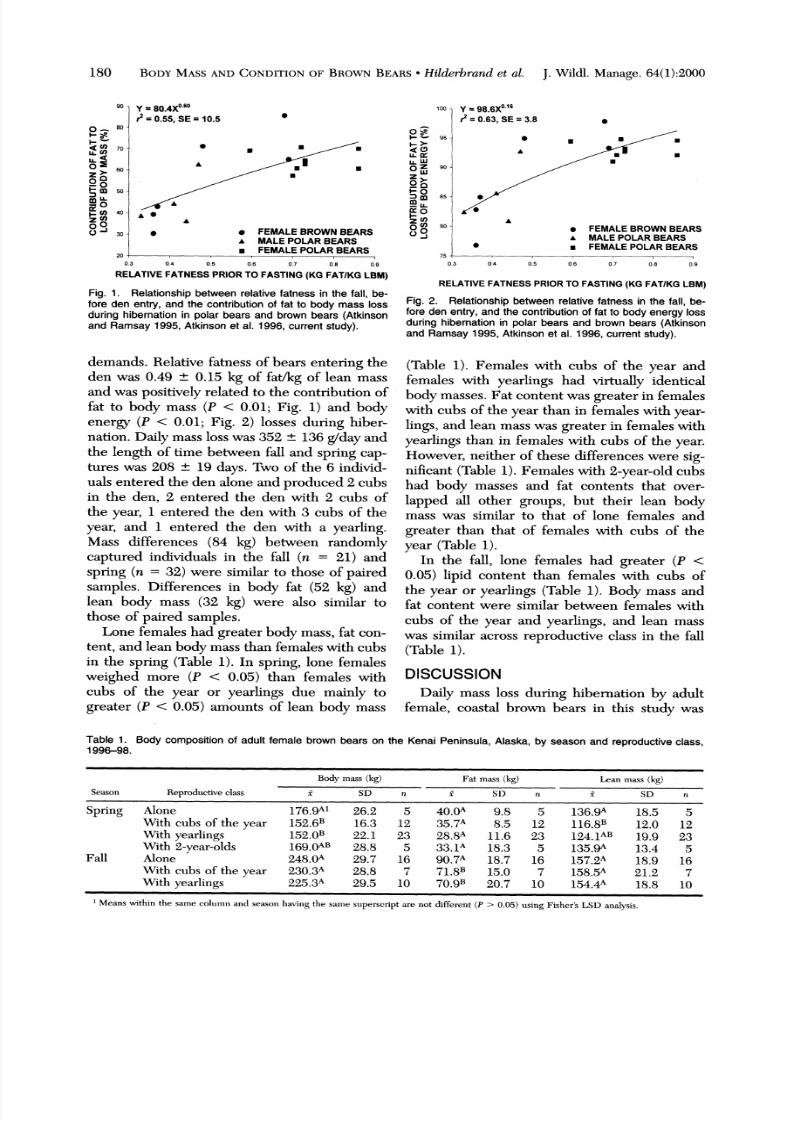
Y = 80.4X6?0r = 0.55, SE= 10.5 0
A . *
* *
' 30 * * FEMALE ROWN BEARSA MALE OLAR BEARS* FEMALE OLAR BEARS0 Ii , , i
0.3 0.4 0.5 0.6 0.7 0.8 0.9
RELATIVE ATNESS PRIOR TO FASTING KG FATIKG BM)
Fig. 1. Relationship etween relative atness in the fall, be-fore den entry, and the contribution f fat to body mass lossduring hibernation n polar bears and brown bears (Atkinsonand Ramsay 1995, Atkinson t al. 1996, current tudy).
demands. Relative fatness of bears entering theden was 0.49 ? 0.15 kg of fat/kg of lean massand was positively related to the contribution offat to body mass (P < 0.01; Fig. 1) and bodyenergy (P < 0.01; Fig. 2) losses during hiber-nation. Daily mass loss was 352 ? 136 g/day andthe length of time between fall and spring cap-tures was 208 ? 19 days. Two of the 6 individ-uals entered the den alone and produced 2 cubsin the
den,2 entered the den with 2
cubs ofthe year, I entered the den with 3 cubs of theyear, and 1 entered the den with a yearling.Mass differences (84 kg) between randomlycaptured individuals in the fall (n = 21) andspring (n = 32) were similar to those of pairedsamples. Differences in body fat (52 kg) andlean body mass (32 kg) were also similar tothose of paired samples.
Lone females had greater body mass, fat con-tent, and lean body mass than females with cubsin the spring (Table 1). In spring, lone femalesweighed more (P < 0.05) than females withcubs of the year or yearlings due mainly togreater (P < 0.05) amounts of lean body mass
100
I- I
LL I
uZ 90
z-o
75
05-
O
750.3
Y = 98.6X?01'? = 0.63, SE = 3.8 0
s *
A/
* FEMALE BROWN BEARSA MALE POLAR BEARS
* * FEMALE POLAR BEARS
0.4 0.5 0.6 0.7 0.8 0.9
RELATIVE FATNESS PRIOR TO FASTING (KG FAT/KG LBM)
Fig. 2. Relationship etween relative atness in the fall, be-fore den entry, and the contribution f fat to body energy lossduring hibernation n polar bears and brown bears (Atkinsonand Ramsay 1995, Atkinson t al. 1996, current tudy).
(Table 1). Females with cubs of the year andfemales with yearlings had virtually identicalbody masses. Fat content was greater n femaleswith cubs of the year than in females with year-lings, and lean mass was greater n females withyearlings than in females with cubs of the year.However, neither of these differences were sig-nificant (Table 1). Females with 2-year-old ubshad body masses and fat contents that over-
lapped all other groups, but their lean bodymass was similar to that of lone females andgreater than that of females with cubs of theyear (Table 1).
In the fall, lone females had greater (P <0.05) lipid content than females with cubs ofthe year or yearlings (Table 1). Body mass andfat content were similar between females withcubs of the year and yearlings, and lean masswas similar across reproductive class in the fall(Table 1).
DISCUSSIONDaily mass loss during hibernation by adult
female, coastal brown bears in this study was
Table 1. Body composition of adult female brown bears on the Kenai Peninsula, Alaska, by season and reproductive class,1996-98.
Body mass (kg) Fat mass (kg) Lean mass (kg)Season Reproductive class x SD n x SD n x SD n
Spring Alone 176.9A1 26.2 5 40.OA 9.8 5 136.9A 18.5 5With cubs of the year 152.6B 16.3 12 35.7A 8.5 12 116.8B 12.0 12With
yearlings152.0B
22.1 23 28.8A 11.6 23 124.1AB 19.9 23With 2-year-olds 169.0AB 28.8 5 33.1A 18.3 5 135.9A 13.4 5Fall Alone 248.0A 29.7 16 90.7A 18.7 16 157.2A 18.9 16
With cubs of the year 230.3A 28.8 7 71.8B 15.0 7 158.5A 21.2 7With yearlings 225.3A 29.5 10 70.9B 20.7 10 154.4A 18.8 10
1 Means within the same column and season having the same superscript are not different (P > 0.05) using Fisher's LSD analysis.
90
0 80
< ?) 70
ILI
I--"'
<(0 4060
O
0?

8/14/2019 Effect of Hibernation and Reproductive Status on Body Mass and Condition of Coastal Brown Bears
http://slidepdf.com/reader/full/effect-of-hibernation-and-reproductive-status-on-body-mass-and-condition-of 5/7
J. Wildl. Manage. 64(1):2000 BODY MASS AND CONDITION OF BROWN BEARS * Hilderbrand et al.
similar to that reported in previous studies ofcaptive nonlactating (335 g/day) and lactating(490 g/day) brown bears of similar mass (Farleyand Robbins 1995). While virtually all of themass lost during hibernation by nonlactating,captive females was lipid (Farley and Robbins1995), the demands of lactation can increaseboth energy and protein catabolism during hi-bernation (Farley and Robbins 1995, Barbozaet al. 1997, Tinker et al. 1998). Two bears usedto assess the costs of hibernation n the currentstudy gave birth in the den, and therefore, lac-tated during part of the dormancy. As expected,these bears utilized lean body mass during hi-bernation (11.6 and 35.0 kg, 14.3 and 34.9% ofmass loss, respectively). However, each of the 4remaining bears in the current study that didnot give birth during hibernation also lost leanbody mass (19.0 to 54.6 kg, 28.8 to 69.7% ofmass loss) and lipid stores. Thus, lean bodymass was catabolized by hibernating adult fe-male brown bears in this study in the absenceof lactation demands. Potential causes of leanbody mass losses by bears in the current studyinclude catabolism during hibernation, catabo-lism during the brief period bears spent outside
the den after fall captures and before springcaptures, and water losses due to dehydration(Farley and Robbins 1994).
Atkinson et al. (1996) reported that 74 to99% of the energy used by polar bears to meetmaintenance costs during fasting was derivedfrom lipid stores. We report a similar range (77to 98%) in this study. Relative fatness also influ-enced the proportion of body lipids and leanbody mass used to meet maintenance costs (At-kinson et al. 1996). Our findings support this
argument n brown bears (Fig. 2). Atkinson andRamsay (1995) suggested that increases in leanbody mass in polar bears before the reproduc-tive fast serve 2 major roles: (1) to support theprotein and mineral demands of cub productionand lactation, and (2) to provide the muscula-ture necessary to support the large fat stores. Arole in supporting cub production has been doc-umented in captive studies quantifying he en-ergy and protein demands of cub productionand lactation (Farley and Robbins 1995). In thecurrent study, lone (pregnant) females enteredthe den with a lean body mass virtually denticalto other reproductive classes. However, femalesexiting the den with cubs of the year had thelowest lean body mass of any reproductive class(Table 1). The second major role above is evi-
denced by the increase in lean body mass thatoccurs between spring and fall, regardless of re-productive class (Table 1). Furthermore, astingmale polar bears lose a greater proportion oflean body mass than fasting females faced withthe additional protein demands of cub produc-tion and lactation (Atkinson and Ramsay 1995,Atkinson et al. 1996). While increased proteinstores accumulated before fasting can be mo-bilized to meet these 2 roles, "normal" roteinstores are likely conserved (Atkinson and Ram-say 1995). A general maintenance of smoothand skeletal muscle function and integrity dur-ing winter dormancy occurs in bears despite ca-tabolism of protein during hibernation Barbozaet al. 1997, Tinker et al. 1998).
Relative fatness at the onset of fasting s likelya critical factor impacting the relative propor-tion of lipid stores and lean body mass used tomeet protein and energy demands in both maleand female bears. Thus, studies of fat and pro-tein metabolism in captive, hibernating bears(Lundberg et al. 1976, Farley and Robbins1995, Barboza et al. 1997) that have concludedthat weight loss is due almost entirely to fat ca-tabolism are likely skewed by the obesity of cap-
tive bears relative to wild bears.The reduced body mass and lean body mass
of females with cubs, particularly cubs of theyear and yearlings, reflects the combined costsof cub production, actation, and increased foodacquisition. Atkinson and Ramsay (1995) re-ported differences similar to those in the cur-rent study across reproductive classes in adultfemale polar bears. Females with cubs of theyear accumulate ean body mass during the ac-tive period and, by fall, have lean body mass
contents virtually dentical to those of lone fe-males (Table 1). However, the presence of cubsmay force females to feed in less productivehabitats or at less productive imes to avoid cubpredation by more dominant bears, particularlymales (Schoen and Beier 1990, McLellan 1994,Mattson and Reinhardt 1995), in turn leadingto reduced fat depots relative to lone females.The lean body mass of females with yearlingsthe following spring is intermediate to that oflone females and females with cubs of the year(Table 1). This result suggests that the contin-ued nursing of cubs 9-15 months of age duringhibernation ncreases protein and-or energy de-mands relative to lone bears. The similar bodycomposition of females with 2-year-old cubsand lone females suggests a decreased energy
181

8/14/2019 Effect of Hibernation and Reproductive Status on Body Mass and Condition of Coastal Brown Bears
http://slidepdf.com/reader/full/effect-of-hibernation-and-reproductive-status-on-body-mass-and-condition-of 6/7
182 BODY MASS AND CONDITION OF BROWN BEARS * Hilderbrand et al. J. Wildl. Manage. 64(1):2000
and protein demand as the cubs are weaned.Unfortunately, studies of body composition ofmothers and offspring and mass transfer be-tween the 2 have not been conducted with cubs>9 months of age.
Female brown bears throughout their geo-graphic range face similar energetic demands ofhibernation and reproduction, despite variednutritional resources (McLellan 1994, Hilder-brand et al. 1999). In some populations, thecosts of hibernation and reproduction may limitreproductive effort and population productivityas reproductive success is positively related tobody mass and fat content. Studies that usebody mass and-or composition as indices of
population health should ensure that the sam-ple is representative of the population and notskewed by season or reproductive class.
ACKNOWLEDGMENTS
We wish to thank biologists T. Bailey, R.Ernst, G. Del Frate, L. Lewis, B. Scotten, E.Shochat, and T. Spraker and pilots T. Cambier,J. DeCreeft, J. Larivee, C. Loftsted, V. Loftsted,L. Nichols, L. Rogers, and K. Solloy for theirassistance with sample collection. E. C. Hell-gren, S. D. Farley, and 2 anonymous reviewersprovided insightful comments that improvedthis manuscript. Funding for this project wasprovided by the Intermountain Forest andRange Experiment Station and Pacific North-west Research Station of the U.S. Forest Ser-vice, U.S. Fish and Wildlife Service, NationalPark Service, Alaska Department of Fish andGame, National Fish and Wildlife Foundation,Kenai Peninsula Chapter of Safari Club, Inter-national, and Raili Korkka Brown Bear Endow-
ment.
LITERATURE CITED
ATKINSON, . N., R. A. NELSON, AND M. A. RAMSAY.1996. Changes in the body composition of fastingpolar bears (Ursus maritimus): the effect of rel-ative fatness on protein conservation. Physiologi-cal Zoology 69:304-316.
, AND M. A. RAMSAY. 995. The effect of pro-longed fasting on the body composition and re-productive success of female polar bears (Ursusmaritimus). Functional Ecology 9:559-567.
BARBOZA, . S., S. D. FARLEY, AND C. T. ROBBINS.1997. Whole-body urea cycling and protein turn-over during hyperphagia and dormancy in grow-ing bears (Ursus americanus and U. arctos). Ca-nadian Journal of Zoology 75:2129-2136.
BLANCHARD, . M. 1987. Size and growth patterns ofthe Yellowstone grizzly bear. International Con-
ference on Bear Research and Management 7:99-107.
BUNNELL, F. L., AND D. E. N. TAIT. 1981. Populationdynamics of bears-implications. Pages 75-98 inC. W. Fowler and T. D. Smith, editors. Dynamicsof large mammal populations. John Wiley & Sons,New York, New York, USA.
FARLEY, S. D., AND C. T. ROBBINS. 1994. Develop-ment of two methods to estimate body composi-tion of bears. Canadian Journal of Zoology 72:220-226.
, AND . 1995. Lactation, hibernation,and mass dynamics of American black bears andgrizzly bears. Canadian Journal of Zoology 73:2216-2222.
HILDERBRAND, . V., S. D. FARLEY, AND C. T. ROB-BINS. 1998. Predicting body condition of bears viatwo field methods. Journal of Wildlife Manage-
ment 62:406-409., C. C. SCHWARTZ, . T. ROBBINS, M. E. JA-COBY, T. A. HANLEY, . M. ARTHUR, AND C. SER-VHEEN. 1999. Importance of dietary meat, partic-ularly salmon, to body size, population productiv-ity, and conservation of North American brownbears. Canadian Journal of Zoology 77:132-138.
LUNDBERG, D. A., R. A. NELSON, H. W. WAHNER,AND J. D. JONES. 1976. Protein metabolism in theblack bear during hibernation. Mayo Clinic Pro-ceedings 51:716-722.
MATTSON, D. J., AND D. P. REINHART. 995. Influ-ence of cutthroat trout (Oncorhynchus clarki) onbehavior and reproduction of Yellowstone grizzlybears (Ursus arctos), 1975-1989. Canadian Jour-nal of Zoology 73:2072-2079.
MCLELLAN, B. N. 1994. Density-dependent popula-tion regulation of brown bears. InternationalConference on Bear Research and ManagementMonographs Series 3:15-37.
NOYCE, K. V., AND D. L. GARSHELIS. 994. Body sizeand blood characteristics as indicators of condi-tion and reproductive performance in blackbears. International Conference on Bear Re-search Management 9:481-496.
ROGERS, L. 1976. Effects of mast and berry crop fail-ures on survival, growth, and reproductive suc-cess of black bears. Transactions of the NorthAmerican Wildlife and Natural Resource Confer-ence 41:431-438.
. 1987. Effects of food supply and kinship onsocial behavior, movements, and populationgrowth of black bears in northeastern Minnesota.Wildlife Monographs 97.
SAMSON, C., AND J. HUOT. 1995. Reproductive biol-ogy of female black bears in relation to body sizein early winter. Journal of Mammalogy 76:68-77.
SCHOEN, ., AND L. BEIER. 1990. Brown bear habitatpreferences and brown bear logging and miningrelationships in southeast Alaska. Alaska Depart-ment of Fish and Game, Federal Aid in WildlifeRestoration Study 4.17.
STRINGHAM, . F. 1990. Grizzly bear reproductiverate relative to body size. International Confer-ence on Bear Research and Management 8:433-443.
TAYLOR, W. P., H. V. REYNOLDS, II, AND W. B. BAL-LARD. 1989. Immobilization of grizzly bears with

8/14/2019 Effect of Hibernation and Reproductive Status on Body Mass and Condition of Coastal Brown Bears
http://slidepdf.com/reader/full/effect-of-hibernation-and-reproductive-status-on-body-mass-and-condition-of 7/7
J. Wildl. Manage. 64(1):2000 BODY MASS AND CONDITION OF BROWN BEARS * Hilderbrand et al.J. Wildl. Manage. 64(1):2000 BODY MASS AND CONDITION OF BROWN BEARS * Hilderbrand et al.
tiletamine hydrochloride and zolazepam hydro-chloride. Journal of Wildlife Management 53:978-981.
TINKER, D. B., H. J. HARLOW, NDT. D. BECK. 1998.Protein use and muscle fiber
changesin free-
ranging, hibernating black bears. PhysiologicalZoology 71:414-424.
WATTS, P. D., AND C. JONKEL. 1988. Energetic costs
tiletamine hydrochloride and zolazepam hydro-chloride. Journal of Wildlife Management 53:978-981.
TINKER, D. B., H. J. HARLOW, NDT. D. BECK. 1998.Protein use and muscle fiber
changesin free-
ranging, hibernating black bears. PhysiologicalZoology 71:414-424.
WATTS, P. D., AND C. JONKEL. 1988. Energetic costs
of winter dormancy in grizzly bear. Journal ofWildlife Management 52:654-656.
ZAR, J. H. 1984. Biostatistical analysis. Prentice Hall,Englewood Cliffs, New Jersey, USA.
Received 18 February 1999.Accepted 3 August 1999.Associate Editor: Hellgren.
of winter dormancy in grizzly bear. Journal ofWildlife Management 52:654-656.
ZAR, J. H. 1984. Biostatistical analysis. Prentice Hall,Englewood Cliffs, New Jersey, USA.
Received 18 February 1999.Accepted 3 August 1999.Associate Editor: Hellgren.
ESTIMATING OPULATION IZE OF GRIZZLY EARS USING HAIRCAPTURE, DNA PROFILING, ND MARK-RECAPTURE NALYSIS
GARTH MOWAT, ish and Wildlife Division, Timberland onsultants, Box 171, Nelson, BC V1L 5P9 CanadaCURTIS TROBECK, eparment f Biological Sciences, University f Alberta, Edmonton, AB T6G 2E9 Canada
Abstract: We used DNA analysis to estimate grizzly bear (Ursus arctos) population size in a 9,866-km2 area
in southeast British Columbia and a 5,030-km2 area in southwest Alberta. We sampled bears by removing hair
at bait sites surrounded by a single strand of barbed wire. DNA profiling with microsatellites of the root portionof the hair was used to identify individuals. We collected hair from 109 different bears and had 25 recapturesin 5 10-day trapping sessions in British Columbia. In Alberta we collected hair from 37 bears and had 9
recaptures in 4 14-day sessions. A model in program CAPTURE (Mh) that accommodates heterogeneity in
individual capture probabilities estimated the population size in British Columbia as 262 (95% CI = 224-313)
and in Alberta as 74 (60-100). We believe that hair capture combined with DNA profiling is a promising
technique for estimating distribution and abundance of bears and potentially many other species. This approachis of special interest to management biologists because it can be applied at the scale conservation and man-
agement decisions are made.
JOURNAL OF WILDLIFE ANAGEMENT 4(1):183-193
Key words: Alberta, British Columbia, Canada, density, DNA profiling, grizzly bears, hair removal, mark-
recapture, microsatellite analysis, population size, Ursus arctos.
ESTIMATING OPULATION IZE OF GRIZZLY EARS USING HAIRCAPTURE, DNA PROFILING, ND MARK-RECAPTURE NALYSIS
GARTH MOWAT, ish and Wildlife Division, Timberland onsultants, Box 171, Nelson, BC V1L 5P9 CanadaCURTIS TROBECK, eparment f Biological Sciences, University f Alberta, Edmonton, AB T6G 2E9 Canada
Abstract: We used DNA analysis to estimate grizzly bear (Ursus arctos) population size in a 9,866-km2 area
in southeast British Columbia and a 5,030-km2 area in southwest Alberta. We sampled bears by removing hair
at bait sites surrounded by a single strand of barbed wire. DNA profiling with microsatellites of the root portionof the hair was used to identify individuals. We collected hair from 109 different bears and had 25 recapturesin 5 10-day trapping sessions in British Columbia. In Alberta we collected hair from 37 bears and had 9
recaptures in 4 14-day sessions. A model in program CAPTURE (Mh) that accommodates heterogeneity in
individual capture probabilities estimated the population size in British Columbia as 262 (95% CI = 224-313)
and in Alberta as 74 (60-100). We believe that hair capture combined with DNA profiling is a promising
technique for estimating distribution and abundance of bears and potentially many other species. This approachis of special interest to management biologists because it can be applied at the scale conservation and man-
agement decisions are made.
JOURNAL OF WILDLIFE ANAGEMENT 4(1):183-193
Key words: Alberta, British Columbia, Canada, density, DNA profiling, grizzly bears, hair removal, mark-
recapture, microsatellite analysis, population size, Ursus arctos.
Estimating carnivore abundance s central totheir conservation, however, options for esti-mating carnivore population size are few andoften require specific circumstances or assump-
tionsthat are difficult to meet.
Estimatingbear
population size is difficult and only has beenachieved in conjunction with intensive effort(McLellan 1989, Garshelis 1992, Stirling et al.1997). Recent efforts to develop bear inventorymethods have involved the use of mark-recap-ture modeling. Researchers have used live cap-ture to mark bears and then recaptured bearsusing camera stations (Mace et al. 1994), aerialsurvey (Larsen and Markel 1989, Stirling et al.1997), and hair removal and DNA fingerprint-ing analysis (Proctor 1995). Most recently, hairremoval and DNA fingerprinting have beenused to mark and recapture grizzly bears(Woods et al. 1996, 1999). This latter methodhas several benefits as live capture of bears isunnecessary, ndividuals can be identified with
Estimating carnivore abundance s central totheir conservation, however, options for esti-mating carnivore population size are few andoften require specific circumstances or assump-
tionsthat are difficult to meet.
Estimatingbear
population size is difficult and only has beenachieved in conjunction with intensive effort(McLellan 1989, Garshelis 1992, Stirling et al.1997). Recent efforts to develop bear inventorymethods have involved the use of mark-recap-ture modeling. Researchers have used live cap-ture to mark bears and then recaptured bearsusing camera stations (Mace et al. 1994), aerialsurvey (Larsen and Markel 1989, Stirling et al.1997), and hair removal and DNA fingerprint-ing analysis (Proctor 1995). Most recently, hairremoval and DNA fingerprinting have beenused to mark and recapture grizzly bears(Woods et al. 1996, 1999). This latter methodhas several benefits as live capture of bears isunnecessary, ndividuals can be identified with
a small risk of error, and hair removal sites arefaster to set and are checked less often thanlive-capture sites. Simpler logistics allow astudy design that comes closer to meeting theassumptions and sample size requirements ofcurrent mark-recapture models.
There are several important assumptions n-volved with mark-recapture nalysis o estimatepopulation size. The most important may be theassumption of population closure, which Whiteet al. (1982) separate into demographic andgeographic closure. Demographic closure as-sumes there are no births or deaths or perma-nent immigration or emigration during thestudy. Errors due to demographic changes inpopulation size are likely to be small for bears,especially if the duration of study is restrictedto 6-10 weeks. Geographic closure is violated ifindividuals move on and off the study area be-tween trapping sessions. A positive bias resultswhen animals have home ranges that span the
a small risk of error, and hair removal sites arefaster to set and are checked less often thanlive-capture sites. Simpler logistics allow astudy design that comes closer to meeting theassumptions and sample size requirements ofcurrent mark-recapture models.
There are several important assumptions n-volved with mark-recapture nalysis o estimatepopulation size. The most important may be theassumption of population closure, which Whiteet al. (1982) separate into demographic andgeographic closure. Demographic closure as-sumes there are no births or deaths or perma-nent immigration or emigration during thestudy. Errors due to demographic changes inpopulation size are likely to be small for bears,especially if the duration of study is restrictedto 6-10 weeks. Geographic closure is violated ifindividuals move on and off the study area be-tween trapping sessions. A positive bias resultswhen animals have home ranges that span the
18383





























