Embed Size (px)
Citation preview

Endocrine Journal 2013, 60 (11), 1241-1249
INSULIN resistance associated with obesity, particu-larly visceral obesity, has been revealed to be involved in the pathogenesis of lifestyle-related diseases, such as type 2 diabetes, hypertension, non-alcoholic steato-hepatitis, chronic kidney disease and arteriosclerosis. In addition, inflammation caused by chronic infections, such as periodontal disease, is known to cause deterio-ration in glucose tolerance and insulin sensitivity [1]. Chronic infections, such as periodontal disease, have been shown to induce increased circulating levels of interleukin (IL) -6, IL-1β, IL-21 and C reactive protein. Overproduction of these cytokines leads to reduced insulin sensitivity and deterioration of glucose toler-ance. Moreover, a local inflammation can later give
Effect of nematode Trichinella infection on glucose tolerance and status of macrophage in obese mice
Hideyuki Okada1), Takahide Ikeda1), Kazuo Kajita1), Ichiro Mori1), Takayuki Hanamoto1), Kei Fujioka1), Masahiro Yamauchi1), Taro Usui1), Noriko Takahashi1), Yoshihiko Kitada1), Koichiro Taguchi1), Yoshihiro Uno1), Hiroyuki Morita1), Zhiliang Wu2), Isao Nagano2), Yuzo Takahashi2), Takuya Kudo1), Kazuki Furuya1), Takahiro Yamada1) and Tatsuo Ishizuka1)
1) Department of General Internal Medicine, Gifu University Graduate School of Medicine, Gifu 501-1194, Japan2) Department of Parasitology, Gifu University Graduate School of Medicine, Gifu 501-1194, Japan
Abstract. We investigated the effect of Trichinella infection on glucose tolerance and (pro- or anti-inflammatory) macrophage status in adipose tissue. Ob/ob mice and high fat-fed mice (obesity model) and C57/BL mice (control mice) were orally infected with (infected group) or without (uninfected group) 400 Trichinella per mouse. Four weeks later, the mice were subjected to investigation, which showed that fasting plasma glucose levels decreased in the infected group of C57/BL and ob/ob mice. Glucose tolerance, evaluated with intraperitoneal GTT, improved in the infected group of ob/ob mice and high fat-fed mice compared with the uninfected groups. Additional assay included anti-inflammatory macrophage (M2) markers and pro-inflammatory macrophage (M1) markers, with the aim to explore the effect of Trichinella infection on adipose tissue inflammation, since our previous study identified anti-inflammatory substances in secreted proteins by Trichinella. The result showed that mRNA levels of M2 markers, such as CD206, arginase and IL-10, increased, whereas M1 markers, such as CD11c, iNOS and IL-6, decreased in the stromal vascular fraction (SVF) isolated from epididymal fat in ob/ob mice. Residential macrophages obtained from the peritoneal lavage exhibited lower M1 markers and higher M2 markers levels in the infected group than in the uninfected group. Trichinella infection increases the ratio of M2/M1 systemically, which results in an improvement in pro-inflammatory state in adipose tissue and amelioration of glucose tolerance in obese mice.
Key words: Parasite infection, Insulin resistance, Adipose tissue inflammation, Macrophage
rise to a systemic inflammatory state. However, the effect of a chronic parasitic infection on glucose tol-erance remains unclear. Parasitic helminth is known to secrete an immunosuppressive agent and anti-in-flammatory materials to suppress an attack from a host [2]. Therefore, it is possible that a parasitic infection may provide the opposite effect to a bacterial infection regarding insulin sensitivity.
On the other hand, a chronic low grade inflamma-tion in adipose tissue is currently considered as a main cause of insulin resistance associated with obesity. Monocyte chemoattractant protein-1 (MCP-1) over-expressing mice exhibit insulin resistance and mac-rophage infiltration in adipose tissue, whereas high fat diet-induced insulin resistance, inflammatory pro-file of adipose tissue and hepatic steatosis are pre-vented in MCP-1 deficient mice, as well as MCP-1 receptor deficient mice [3, 4]. These data indicate that MCP-1, which is secreted in hypertrophic adipocytes,
Submitted Feb. 12, 2013 as EJ13-0056; Accepted Jul. 30, 2013 as EJ13-0312. Released online in J-STAGE as advance publication Aug. 29, 2013Correspondence to: Tatsuo Ishizuka, Department of General Internal Medicine, Gifu University Graduate School of Medicine, Yanagido 1-1, Gifu, 501-1194, Japan. E-mail: [email protected]
Original
©The Japan Endocrine Society

1242 Okada et al.
cloning the cytokine macrophage migration inhibitory factor (MIF) homologue from Trichinella [7]. Another line of our study demonstrated that Trichinella infection improves glucose tolerance in control and obese mice without affecting adiposity.
Materials and Methods
1. Animals and infectionOb/ob mice and C57/BL mice were fed CE2 pow-
der (CLEA Japan Inc, Tokyo) ad libitum. In addition, C57/BL mice were fed a high fat diet (High Fat Diet 32, CLEA Japan Inc., Fat: 31.9%) for four weeks as high fat diet-induced obese mice (HF mice). They were divided into two groups (10 mice each): infected orally with 400 Trichinella larvae/mouse (infected group) and a control group (uninfected group) (Fig. 1). Four weeks later, body weight, fat weight (subcutaneous fat, epididymal fat and perirenal fat), fasting plasma glucose and insulin levels were measured as follows. To evaluate glucose tolerance, intraperitoneal glucose tolerance test (ipGTT, glucose: 2g/kg) was performed. Plasma glucose lev-els were measured with Medisafe Mini Blood Glucose Monitoring System (TERUMO, Tokyo, Japan), and plasma insulin levels were assayed with Insulin RIA kit
-induced macrophage accumulation in adipose tissue causes insulin resistance. Moreover, classically acti-vated macrophage (M1 macrophage) expressing tumor necrosis factor (TNF) -α and inducible nitric oxide synthase (iNOS) observed in obese adipose tissue have been shown to be more harmful than alternatively acti-vated macrophage (M2 macrophage) expressing argi-nase 1 and IL-10 in lean adipose tissue [5].
In this study, we investigated the parasite, Trichinella spiralis, for its ability to effect glucose tolerance and affect the status of macrophage in adipose tissue. T. spi-ralis is a parasitic nematode which infects a wide vari-ety of vertebrate hosts. After consumption of infected muscles, the larvae develop into adult worms in the host intestine where the females give rise to the second gen-eration. The newborn larvae migrate through the blood vessels and invade striated muscle cells. Trichinella infection causes local and systemic changes in the host. The former includes muscle cell transformation to a nurse cell in the cyst [6], and the latter includes immu-nological responses which act in two ways: one is resist-ing the second challenge of Trichinella infection and the other is suppressing the immune attack against the already-existing Trichinella in the host muscle. In rela-tion to this research interest, we have previously studied
Fig. 1 Procedure for preparation of mice infected with (infected group) or without (uninfected group) Trichinella. C57/BL and ob/ob mice were fed with normal diet or high fat diet for 4weeks. They were divided into two groups (10 mice each): infected orally with 400 Trichinella larvae/mouse (infected group) and uninfected group (control group). 4weeks or 12weeks later, experiments described in Materials and Methods were performed.

1243Trichinella infection improves glucose tolerance
CTCTGAGGGCTGACACAAGG; Arginase 1: CTC CAAGCCAAAGTCCTTAGAG/AGGAGCTGTCAT TAGGGACATC; G3PDH: GGCATTGTGGAAGGGC TCAT/GACACATTGGGGGTAGGAACAC
3. Immunohistochemical stainingImmunostaining of paraffin sections of the fat tis-
sues was performed using anti-CD11c antibody and anti-CD206 antibody (Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA), and Vectastain ABC kit (Vector Laboratories, Burlingame, CA, USA).
4. StatisticsStatistical comparisons were performed with Student’s
t-test (Fig. 2, Fig. 3C, Fig. 3F, Fig. 4C, Fig. 5 and Fig. 6). Other data were analyzed with one-factor ANOVA (Dunnett test). Data are given as mean ± SEM. Values of P < 0.05 were considered statistically significant.
Results
1. Effects of Trichinella infection on fasting plasma glucose level and glucose tolerance
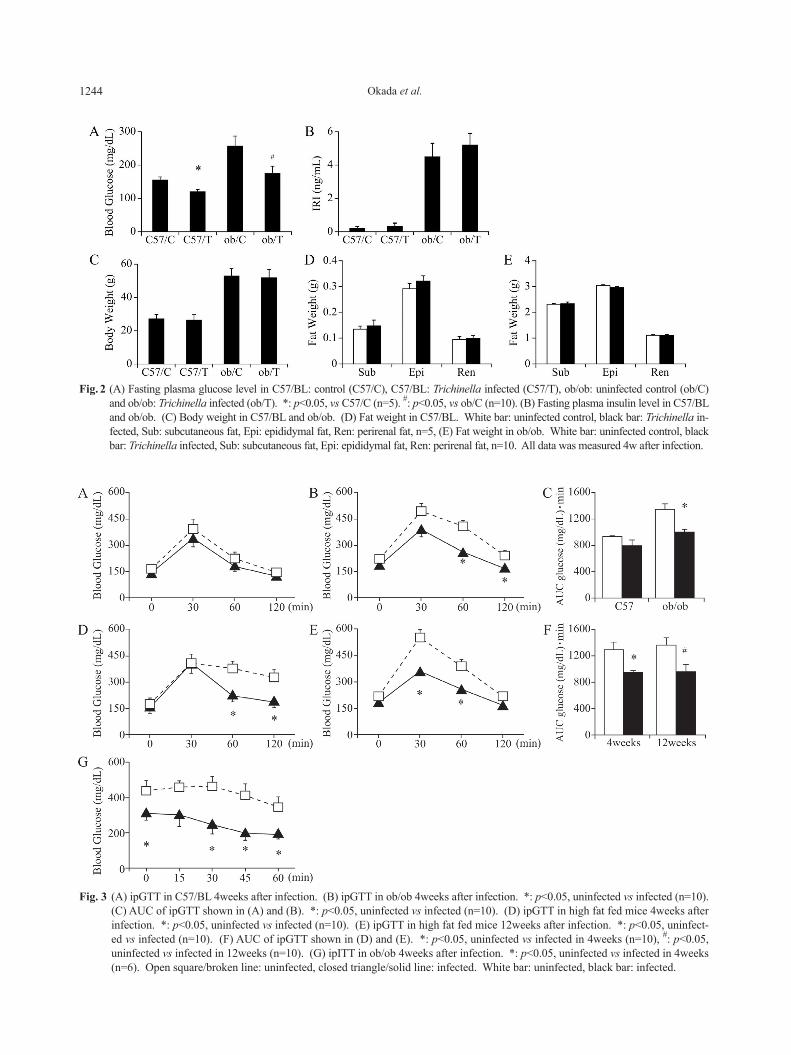
Trichinella infection for four weeks reduced fasting plasma glucose level without influence on plasma insu-lin levels in C57/BL and ob/ob mice (Fig. 2A, B). These results implied that Trichinella infection improved insu-lin sensitivity. Trichinella infection did not affect body weight or fat weight (Fig. 2C-E). To evaluate the effect of Trichinella infection on glucose tolerance, ipGTT was performed. As shown in Fig. 3A-C, Trichinella infec-tion for four weeks suppressed plasma glucose levels during GTT in ob/ob mice, but not in lean control mice as shown in Fig. 3A. It also improved glucose toler-ance in high fat fed mice (Fig. 3D). Moreover, this glu-cose-reducing effect continued for 3 months (12 weeks) after the infection in high fat fed mice (Fig. 3E). On the other hand, ipITT performed in ob/ob mice revealed that insulin sensitivity was improved in the infected group compared with the uninfected group (Fig. 3G). Taken together, Trichinella infection improved glucose tolerance and insulin sensitivity without weight reduc-tion especially in obese mice.
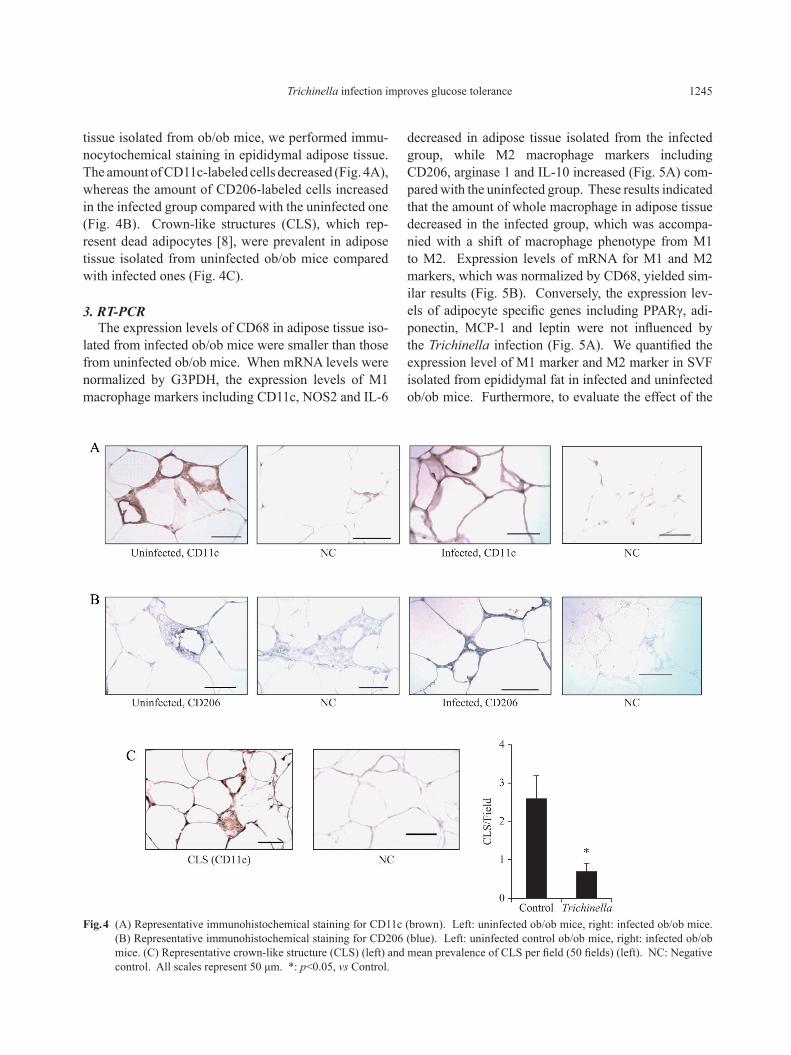
2. Immunocytochemical stainingAs Trichinella infection decreased plasma glucose
level more effectively in obese animals, we specu-lated that it may act on adipose tissue inflammation. To observe infiltration of inflammatory cells in adipose
(Linco Research, St. Charles, MO, USA). In addition, intraperitoneal insulin tolerance test (ipITT, insulin: 0.3 IU/kg) was performed in ob/ob mice. An area under the curve (AUC) was measured from glucose levels at 0, 30, 60 and 120 min during each ipGTT.
2. Real time PCRTo determine the expression levels of adipocyte spe-
cific genes (peroxisome proliferator-activated recep-tor gamma (PPARγ), leptin and adiponectin), cytokine genes (IL-6, IL-10 and MCP-1), M1 macrophage mark-ers (CD11c and NOS) and M2 macrophage marker (CD206, arginase 1), real time PCR (RT-PCR) was per-formed. Total RNA was isolated from epididymal adi-pose tissue. In addition, adipocytes and stromal vas-cular fraction (SVF) were obtained with collagenase digestion. Residential macrophages were harvested by lavaging the peritoneal cavity with ice cold PBS. Total RNA was isolated from samples using TRIZOL (GIBCO BRL, Life Technologies, Inc., Carlsbad, CA, USA) according to the manufacturer’s instructions. To reduce genomic DNA contamination, the isolated RNA was treated with RQ1 RNase-free DNase (Promega, Madison, WI, USA), which was monitored with PCR using primers for the subsequent RT-PCR. Reverse transcription was performed using a SuperScript Reverse Transcriptase (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. PCR amplification was performed as follows: prede-nature for 1 cycle at 95°C for 30s, and 40 cycles at 95°C for 5s, 60-62°C for 30s using a Thermal Cycler Dice (TAKARA, Ohtsu, Japan). Expression levels, calculated as copy numbers in each sample, were nor-malized to the expression level of G3PDH or CD68. Oligonucleotide primers were designed as follows:PPARγ: CCCTTTGGTGACTTTATGGA/CTGCCTGA GGTCTGTCATCT; Adiponectin: AAGGACAAGGCC GTTCTCT/TATGGGTAGTTGCAGTCAGTTGG; IL-6: GACTTCCATCCAGTTGCCTTCT/TTTTCTCT TTCCACGATTTCC; IL-10: TCCTCTTCACCTGCT CCAC/GCTGGACAACATACTGCTAACC; MCP-1: TCACCTGCTGCTACTCATTCAC/CCATTCCTTC TTGGGGTCAG; Leptin: GACACCAAAACCCTCAT CAA/TCTCCAGGTCATTGGCTATC; CD68: CTTCC CACAGGCAGCACAG/AATGATGAGAGGCAGCA AGAGG; CD11c: CTGGATAGCCTTTCTTCTGCTG/GCACACTGTGTCCGAACTC; CD206: CTGTGTAG TTGTGATTGGTGGC/CTTTTGGAGTAGTGGT TGGAGAA; NOS2: CCAAGCCCTCACCTACTTCC/
1244 Okada et al.
Fig. 3 (A) ipGTT in C57/BL 4weeks after infection. (B) ipGTT in ob/ob 4weeks after infection. *: p<0.05, uninfected vs infected (n=10). (C) AUC of ipGTT shown in (A) and (B). *: p<0.05, uninfected vs infected (n=10). (D) ipGTT in high fat fed mice 4weeks after infection. *: p<0.05, uninfected vs infected (n=10). (E) ipGTT in high fat fed mice 12weeks after infection. *: p<0.05, uninfect-ed vs infected (n=10). (F) AUC of ipGTT shown in (D) and (E). *: p<0.05, uninfected vs infected in 4weeks (n=10), #: p<0.05, uninfected vs infected in 12weeks (n=10). (G) ipITT in ob/ob 4weeks after infection. *: p<0.05, uninfected vs infected in 4weeks (n=6). Open square/broken line: uninfected, closed triangle/solid line: infected. White bar: uninfected, black bar: infected.
Fig. 2 (A) Fasting plasma glucose level in C57/BL: control (C57/C), C57/BL: Trichinella infected (C57/T), ob/ob: uninfected control (ob/C) and ob/ob: Trichinella infected (ob/T). *: p<0.05, vs C57/C (n=5). #: p<0.05, vs ob/C (n=10). (B) Fasting plasma insulin level in C57/BL and ob/ob. (C) Body weight in C57/BL and ob/ob. (D) Fat weight in C57/BL. White bar: uninfected control, black bar: Trichinella in-fected, Sub: subcutaneous fat, Epi: epididymal fat, Ren: perirenal fat, n=5, (E) Fat weight in ob/ob. White bar: uninfected control, black bar: Trichinella infected, Sub: subcutaneous fat, Epi: epididymal fat, Ren: perirenal fat, n=10. All data was measured 4w after infection.
1245Trichinella infection improves glucose tolerance
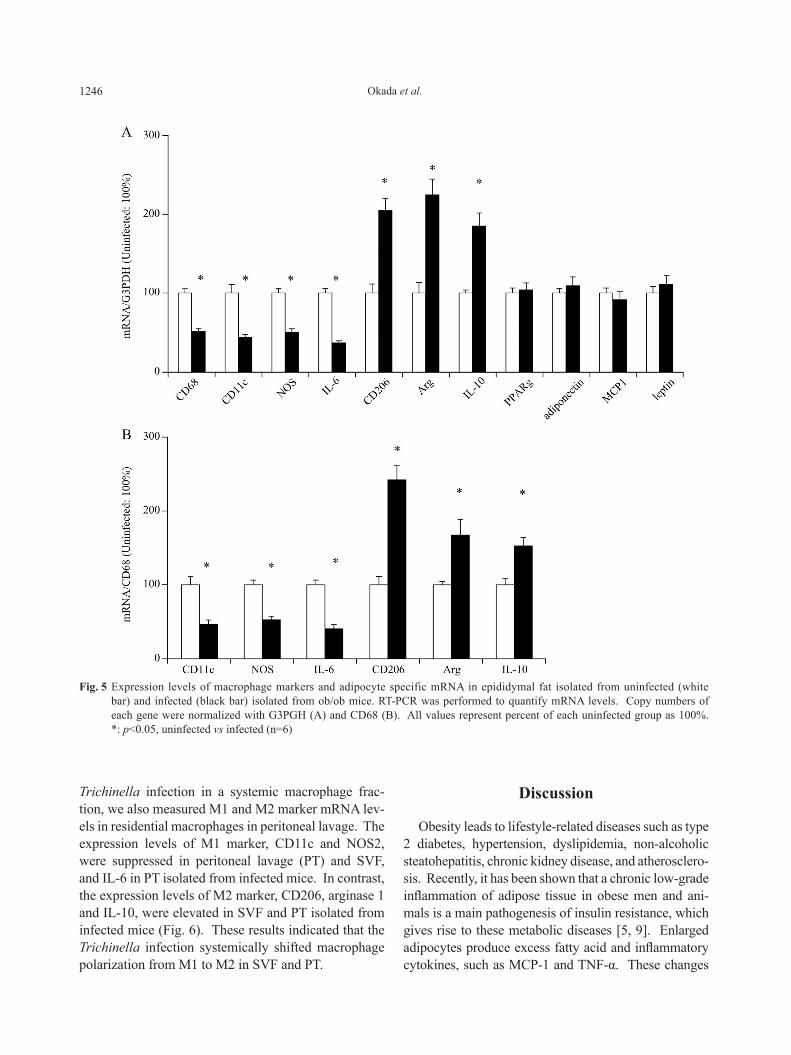
decreased in adipose tissue isolated from the infected group, while M2 macrophage markers including CD206, arginase 1 and IL-10 increased (Fig. 5A) com-pared with the uninfected group. These results indicated that the amount of whole macrophage in adipose tissue decreased in the infected group, which was accompa-nied with a shift of macrophage phenotype from M1 to M2. Expression levels of mRNA for M1 and M2 markers, which was normalized by CD68, yielded sim-ilar results (Fig. 5B). Conversely, the expression lev-els of adipocyte specific genes including PPARγ, adi-ponectin, MCP-1 and leptin were not influenced by the Trichinella infection (Fig. 5A). We quantified the expression level of M1 marker and M2 marker in SVF isolated from epididymal fat in infected and uninfected ob/ob mice. Furthermore, to evaluate the effect of the
tissue isolated from ob/ob mice, we performed immu-nocytochemical staining in epididymal adipose tissue. The amount of CD11c-labeled cells decreased (Fig. 4A), whereas the amount of CD206-labeled cells increased in the infected group compared with the uninfected one (Fig. 4B). Crown-like structures (CLS), which rep-resent dead adipocytes [8], were prevalent in adipose tissue isolated from uninfected ob/ob mice compared with infected ones (Fig. 4C).
3. RT-PCRThe expression levels of CD68 in adipose tissue iso-
lated from infected ob/ob mice were smaller than those from uninfected ob/ob mice. When mRNA levels were normalized by G3PDH, the expression levels of M1 macrophage markers including CD11c, NOS2 and IL-6
Fig. 4 (A) Representative immunohistochemical staining for CD11c (brown). Left: uninfected ob/ob mice, right: infected ob/ob mice. (B) Representative immunohistochemical staining for CD206 (blue). Left: uninfected control ob/ob mice, right: infected ob/ob mice. (C) Representative crown-like structure (CLS) (left) and mean prevalence of CLS per field (50 fields) (left). NC: Negative control. All scales represent 50 μm. *: p<0.05, vs Control.
1246 Okada et al.
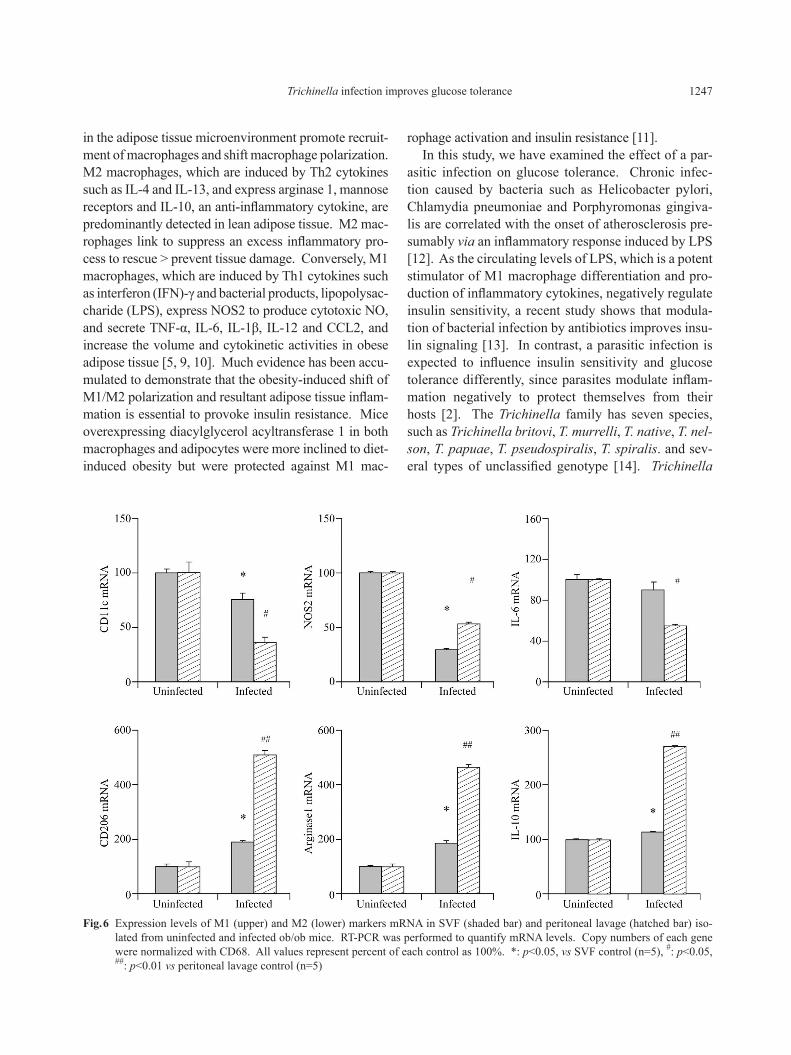
Trichinella infection in a systemic macrophage frac-tion, we also measured M1 and M2 marker mRNA lev-els in residential macrophages in peritoneal lavage. The expression levels of M1 marker, CD11c and NOS2, were suppressed in peritoneal lavage (PT) and SVF, and IL-6 in PT isolated from infected mice. In contrast, the expression levels of M2 marker, CD206, arginase 1 and IL-10, were elevated in SVF and PT isolated from infected mice (Fig. 6). These results indicated that the Trichinella infection systemically shifted macrophage polarization from M1 to M2 in SVF and PT.
Discussion
Obesity leads to lifestyle-related diseases such as type 2 diabetes, hypertension, dyslipidemia, non-alcoholic steatohepatitis, chronic kidney disease, and atherosclero-sis. Recently, it has been shown that a chronic low-grade inflammation of adipose tissue in obese men and ani-mals is a main pathogenesis of insulin resistance, which gives rise to these metabolic diseases [5, 9]. Enlarged adipocytes produce excess fatty acid and inflammatory cytokines, such as MCP-1 and TNF-α. These changes
Fig. 5 Expression levels of macrophage markers and adipocyte specific mRNA in epididymal fat isolated from uninfected (white bar) and infected (black bar) isolated from ob/ob mice. RT-PCR was performed to quantify mRNA levels. Copy numbers of each gene were normalized with G3PGH (A) and CD68 (B). All values represent percent of each uninfected group as 100%. *: p<0.05, uninfected vs infected (n=6)
1247Trichinella infection improves glucose tolerance
rophage activation and insulin resistance [11].In this study, we have examined the effect of a par-
asitic infection on glucose tolerance. Chronic infec-tion caused by bacteria such as Helicobacter pylori, Chlamydia pneumoniae and Porphyromonas gingiva-lis are correlated with the onset of atherosclerosis pre-sumably via an inflammatory response induced by LPS [12]. As the circulating levels of LPS, which is a potent stimulator of M1 macrophage differentiation and pro-duction of inflammatory cytokines, negatively regulate insulin sensitivity, a recent study shows that modula-tion of bacterial infection by antibiotics improves insu-lin signaling [13]. In contrast, a parasitic infection is expected to influence insulin sensitivity and glucose tolerance differently, since parasites modulate inflam-mation negatively to protect themselves from their hosts [2]. The Trichinella family has seven species, such as Trichinella britovi, T. murrelli, T. native, T. nel-son, T. papuae, T. pseudospiralis, T. spiralis. and sev-eral types of unclassified genotype [14]. Trichinella
in the adipose tissue microenvironment promote recruit-ment of macrophages and shift macrophage polarization. M2 macrophages, which are induced by Th2 cytokines such as IL-4 and IL-13, and express arginase 1, mannose receptors and IL-10, an anti-inflammatory cytokine, are predominantly detected in lean adipose tissue. M2 mac-rophages link to suppress an excess inflammatory pro-cess to rescue > prevent tissue damage. Conversely, M1 macrophages, which are induced by Th1 cytokines such as interferon (IFN)-γ and bacterial products, lipopolysac-charide (LPS), express NOS2 to produce cytotoxic NO, and secrete TNF-α, IL-6, IL-1β, IL-12 and CCL2, and increase the volume and cytokinetic activities in obese adipose tissue [5, 9, 10]. Much evidence has been accu-mulated to demonstrate that the obesity-induced shift of M1/M2 polarization and resultant adipose tissue inflam-mation is essential to provoke insulin resistance. Mice overexpressing diacylglycerol acyltransferase 1 in both macrophages and adipocytes were more inclined to diet-induced obesity but were protected against M1 mac-
Fig. 6 Expression levels of M1 (upper) and M2 (lower) markers mRNA in SVF (shaded bar) and peritoneal lavage (hatched bar) iso-lated from uninfected and infected ob/ob mice. RT-PCR was performed to quantify mRNA levels. Copy numbers of each gene were normalized with CD68. All values represent percent of each control as 100%. *: p<0.05, vs SVF control (n=5), #: p<0.05, ##: p<0.01 vs peritoneal lavage control (n=5)

1248 Okada et al.
a parasitic infection may influence systemic immune function. As mesenteric fat links to systemic glucose and lipid metabolism more closely than epididymal fat [15], macrophage polarization in mesenteric fat was a hoped for result and could be estimated. However, the contamination of small lymph nodes located in the mes-entery interfered with the measurement of macrophage markers in SVF, making our method inappropriate to assess macrophage polarization in mesenteric fat.
No difference was observed in gene expression lev-els of adipocyte specific genes including PPARγ, adi-ponectin, leptin and MCP-1, in total RNA isolated from epididymal fat, between infected and uninfected groups as shown in Fig. 5. These results suggest that improved insulin sensitivity in the infected group may be due to the changes of adipocyte specific genes. Our results have been consistent with recent data suggesting that a shift of macrophage polarization toward M2 is sufficient to protect mice against diet-induced insulin resistance [11]. Although the expression of adiponectin, an insu-lin-sensitizing adipokine, was not influenced, another mechanism may be involved in Trichinella infection-in-duced improvement of insulin resistance. Since circu-lating IL-10 enhances insulin sensitivity in vivo and in vitro [16, 17], increasing the expression level of IL-10 in adipose tissue and peritoneal lavage (Fig. 5 and 6) may contribute to prevent insulin resistance. In this study, we examined the effects of Trichinella infection on inflammation of adipose tissue, however, the possi-bility cannot be excluded that other organs, such as the liver and skeletal muscle may be involved.
We have cloned the immunosuppressive agent in the excretory-secretory (ES) products from Trichinella [7]. Other researchers have demonstrated that a par-asite such as Trichinella spiralis, Schistosoma man-soni, Theromyzon tessulatum and Filaria secretes sub-stances that modulate immune response [18, 19]. Our preliminary experiments showed that treatment with ES decreased IL-6 mRNA level and increased IL-10 mRNA level in cultured macrophages (RAW246.7) (data not shown). Taken together, we hypothesize that secreted substances by Trichinella may induce the changes of systemic macrophage polarization followed by increased M2/M1 ratio in SVF, which may result in improved insulin sensitivity and glucose tolerance in obese mice. Identification of a specific substance may lead to a new strategy to treat type 2 diabetes and other inflammatory diseases.
spiralis utilizes almost all carnivores and omnivores as hosts. Trichinella spiralis has a distinctive life cycle that is an intermediate host as well as a final host. If a host ingests Trichinella orally, they perform sexual reproduction in the small intestine, with adult females laying their eggs in the small intestine mucosa, and then the newborn larvae are transported throughout the whole body mainly through the bloodstream, form a cyst in striated muscle fibers, and wait for the opportu-nity to be predated to the next host.
Trichinella infection decreased fasting plasma glu-cose level without any changes in body weight, or fat weight as shown in Fig. 2. As blood glucose levels were more rapidly and markedly reduced by insulin injec-tion, insulin sensitivity improved in the infected group. Improvement of glucose tolerance evaluated by ipGTT was observed in obese mice including ob/ob mice and high fat fed mice. Ingestion of Trichinella causes body weight loss due to enteritis in the first week. It is gradu-ally restored following the movement of the worms to skeletal muscle. The enteritis-induced weight loss could not be recognized four weeks after infection. Since no difference was observed in body weight and fat weight between control and infected groups, the improvement of glucose tolerance in the infected group is not due to malnutrition caused by the parasitic infection.
We performed immunostaining for CD11c and CD206 in adipose tissue of ob/ob mice as shown in Fig. 4. A more abundant expression of CD206 was detected in the infected group than in the uninfected group. A crown-like structure was less frequently observed in adipose tissue isolated from the infected group. These results suggest that obesity-induced chronic inflamma-tion may be suppressed by Trichinella infection. The quantitative analysis of gene expression by RT-PCR showed that mRNA levels of CD68 was reduced in the infected group. Moreover, expression levels of M1 macrophage markers including CD11c, NOS2 and IL-6 were decreased, while M2 macrophage markers including CD206, arginase 1, IL-10 increased in whole adipose tissue, SVF and peritoneal lavage cells of res-idential macrophage isolated from the infected group as shown in Fig. 5, 6. These results indicated that improvement of glucose tolerance in the infected group was associated with reduced infiltration of macrophage and the shift of macrophage polarization toward anti-in-flammatory phenotype in adipose tissue. Because resi-dential macrophage in peritoneal lavage exhibited sim-ilar changes to macrophage in SVF, it is suggested that

1249Trichinella infection improves glucose tolerance
References
1. Marchetti E, Monaco A, Procaccini L, Mummolo S, Gatto R, et al. (2012) Periodontal disease: the influence of metabolic syndrome. Nutr Metab 9: 88.
2. Schmid-Hempel S (2009) Immune defence, parasite evasion strategies and their relevance for ‘macroscopic phenomena’ such as virulence. Philos Trans R Soc Lond B Biol Sci 364: 85-98.
3. Weisberg SP, Hunter D, Huber R, Lemieux J, Slaymaker S, et al. (2006) CCR2 modulates inflammatory and met-abolic effects of high-fat feeding. J Clin Invest 116: 115-124.
4. Kanda H, Tateya S, Tamori Y, Kotani K, Hiasa K, et al. (2006) MCP-1 contributes to macrophage infiltration into adipose tissue, insulin resistance, and hepatic ste-atosis in obesity. J Clin Invest 116: 1494-1505.
5. Lumeng CN, Bodzin JL, Saltiel AR (2007) Obesity induced a phenotypic switch in adipose tissue mac-rophage polarization. J Clin Invest 117: 175-184.
6. Despommier DD, Gold AM, Buck SW, Capo V, Silberstein D (1990) Trichinella spiralis: secreted anti-gen of the infective L1 larva localizes to the cytoplasm and nucleoplasm of infected host cells. Exp Parasitol 71: 27-38.
7. Wu Z, Boonmars T, Nagano I, Nakada T, Takahashi Y (2003) Molecular expression and characterization of a homologue of host cytokine macrophage migration inhibitory factor from Trichinella spp. J Parasitol 89: 507-515.
8. Murano I, Barbatelli G, Parisani V, Latini C, Muzzonigro G (2008) Dead adipocytes, detected as crown-like struc-ture, are prevalent in visceral fat depots of genetically obese mice. J Lipid Res 49: 1562-1568.
9. Hotamisligil GS, et al. (2006) Inflammation and meta-bolic disorders. Nature 444: 860-867.
10. Nguyen MT, Favelyukis S, Nguyen AK (2007) A sub-population of macrophages infiltrates hypertrophic adi-pose tissue and is activated by free fatty acids via Toll-like receptors2 and 4 and JNK-dependent pathways. J Biol Chem 282: 35279-35292.
11. Koliwad SK, Streeper RS, Monetti M, Cornelissen I, Chan L, et al. (2010) DGAT-dependent triacylglycerol storage by macrophages protects mice from diet-in-duced insulin resistance and inflammation. J Clin Invest 120: 756-767.
12. Fujimoto Y, Shimoyama A, Suda Y, Fukase K (2012) Synthesis and immunomodulatory activites of Helicobacter pylori lipophilic terminus of lipopolysac-charide including lipid A. Carbohydr Res 356: 37-43.
13. Carvalho BM, Guadagnini D, Tsukumo DML, Schenka AA, Latuf-Filho P, et al. (2012) Modulation of gut microbiota by antibiotics improves insulin signaling in high-fat fed mice. Diabetologia 55: 2823-2834.
14. Murell KD, et al. (2000) The systematics of the genus Trichinella with a key to species. Vet Parasitol 93: 293-307.
15. Björntrop P (1990) “Portal” adipose tissue as a genera-tor of risk factors for cardiovascular disease and diabe-tes. Arteriosclerosis 10: 493-496.
16. Blüher M, Fasshauer M, Tönjes A, Kratzsch J, Schön M, et al. (2005) Association of interleukin-6, C-reactive protein, interleukin-10, and adiponectin plasma con-centrations with measures of obesity, insulin sensitivity and glucose metabolism. Exp Clin Endocrinol Diabetes 113: 534-537.
17. Cintra DE, Pauli JR, Araújo EP, Moraes JC, de Souza CT, et al. (2008) Interleukin-10 is a protective fac-tor against diet-induced insulin resistance in liver. J Hepatol 48: 628-637.
18. Pastrana DV, Raghavan N, Fitzgerald P, Eisinger SW, Metz C, et al. (1998) Filarial nematode parasites secret a homologue of the human cytokine macrophage migra-tion inhibitory factor. Infect Immun 66: 5955-5963.
19. Pryor SC, Elizee R (2000) Evidence of opiates and opi-oid neuropeptides and their immune effects in para-sitic invertebrates representing three different phyla: Schistosoma mansoni, Theromyzon tessulatum, Trichinella spiralis. Acta Biol Hung 51: 331-341.





























