Embed Size (px)
Citation preview

Neuroscience Letters, 124 (1991) 49-51 © 1991 Elsevier Scientific Publishers Ireland Ltd. 0304-3940/91/$ 03.50 ADONIS 030439409100004C
NSL 07597
49
Effect of neurokinin A, substance P and calcitonin gene related peptide in peripheral hyperalgesia in the rat paw
Meire Nakamura-Cra ig and Balvinder Kaur Gill
Department of Pharmacology, Wellcome Research Laboratories, Beckenham, Kent ( U.K.)
(Received 27 August 1990; Revised version received 5 December 1990; Accepted 7 December 1990)
Key words: Peripheral hyperalgesia; Neurokinin A; Substance P; Calcitonin gene-related peptide
The effect of neurokinin A (NKA), substance P (SP) and calcitonin gene-related peptide (CGRP) in peripheral hyperalgesia was studied in rats using a modification of the Randall-Selitto paw test. NKA was l0 times more potent than SP which was 500 times more potent than CGRP in inducing hyperalgesia in the rat paw, suggesting that NKA and SP but not CGRP could have an important role in acute hyperalgesic conditions. Furthermore, sensitization induced by several injections of subthreshold doses of NKA or CGRP suggest that these neuropeptides along with SP could participate as mediators or modulators of chronic pain.
The hypothesis that SP participates in the nociceptive process has been well investigated and there appears to be enough evidence to support this theory. SP has a pre- ferential excitatory effect on the nociceptive neurones in the dorsal horn of the spinal cord [7, 9, 11, 18]. Recently we suggested that SP may participate as a neuromodula- tor in peripheral inflammatory pain probably associated with chronic conditions [14]. NKA has been shown to be distributed in parallel with SP in the dorsal horn of the spinal cord and its pharmacology has resembled SP in many ways [17]. A similar distribution is also described for CGRP [6]. Furthermore there are suggestions that NKA and CGRP are also involved in nociception [4, 8, 10, 19]. The present study investigates the possible in- volvement of NKA and CGRP in peripheral pain. The possibility that NKA and CGRP produce sensitization as does SP in the rat paw by repeated administration of a subthreshold dose [13], has also been investigated.
Male Wistar rats (150-170 g) were used throughout. Hyperalgesia was measured by a modification of the Randall-Selitto test [2]. In this test, a constant pressure of 20 mmHG is applied to the hind paw of rats (using a syringe and plunger) and is discontinued when the ani- mals present a behavioural response, characterized by a reduction of escape movements and a change of respira- tory frequency. The observation is made whilst the rat is held into the experimental position by the experi-
Correspondence: M. Nakamura-Craig, Department of Pharmacology, Wellcome Research Laboratories, Langley Court, Beckenham, Kent BR3 3BS, U.K.
menter. In control animals the time taken for such a re- sponse to occur is very reproducible (34.8 + O. 12, n = 20) but is less after injection of a hyperalgesic stimulus. Measuring the reaction time before and after injection of a hyperalgesic stimulus subcutaneously into the rat paw (subplantar, s.pl.), provides an assessment of the degree of hyperalgesi~i (expressed as the difference between the two times, d reaction time, seconds). Indomethacin (2 mg/kg, p.o.) was given 30 rain before injection of the hyperalgesic substances. The experimenter was unaware of the group treatments during the test period. All results are presented as means___ S.E.M. The level of statistical significance was determined with Student's t-test for paired samples and differences with P < 0.05 were con- sidered statistically significant. The number of animals in each experimental group was 5.
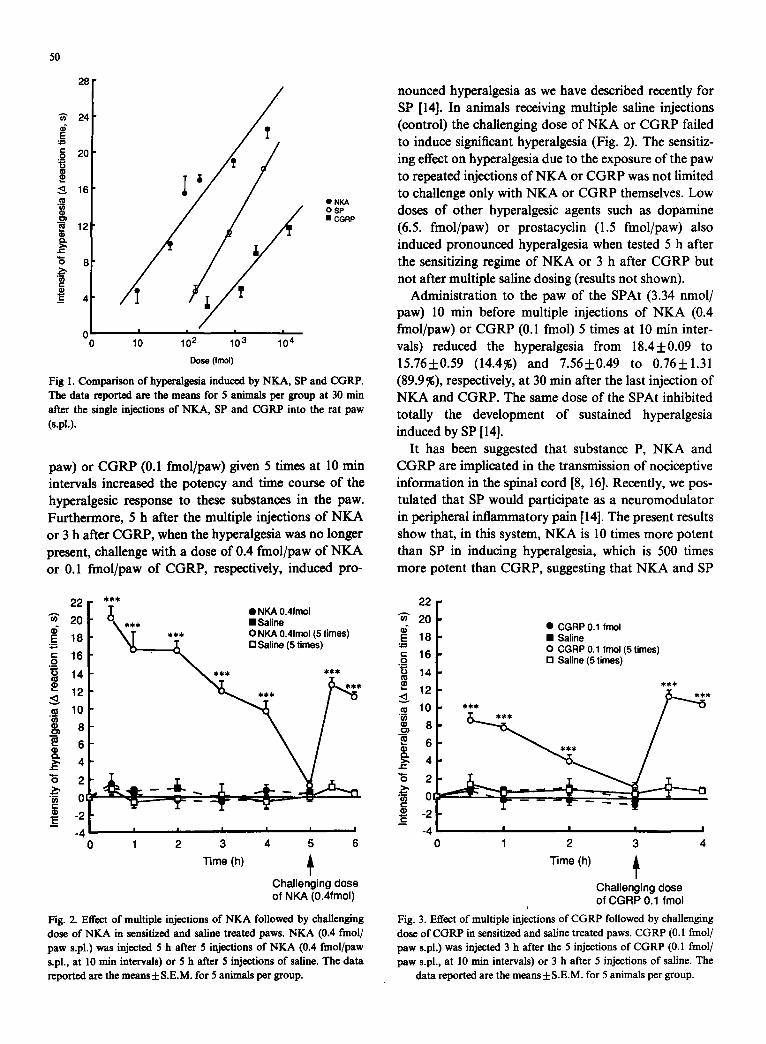
A comparison of hyperalgesia induced by SP, NKA and CGRP is shown in Fig. 1. The figure shows the hyperalgesic potency at 30 min after the subplantar injections of NKA, SP and CGRP, when the hyperalge- sic effect is maximal, the EDs0 and 95 % confidence limits being 0.35 (0-3.62), 3.49 (2.57-5.12) and 1555 nmol/ paw, respectively. The SP antagonist, [D-Arg 1, D-Pro 2, D- Trp 7,9, Leull]Sp (SPAt, 3.34 nmol/paw) 10 rain before SP reduced the hyperalgesia induced by 3.71 nmol/paw at 30 min from 18.2__.0.06 s to 1.2___0.32 s (93.4%). The hyperalgesia induced by NKA (4.41 nmol/paw, 21.1 +0.15 s) and CGRP (2.63 nmol/paw, 8.8 +_0.87 s) at 30 min was not affected by the SPAt, 3.34 nmol/paw (19.2 _ 0.78 s and 11.0 +- 0.34 s, respectively).
Multiple subthreshold injections of NKA (0.4 fmol/
50
28
~" 24 d E
~ 16 ._~
~ 12 O .
e -
"5 8
c
4
I 8
10 102
/
<,/ ! I
10 3 1 0 4
• NKA O SP • CGRP
Dose (fmol)
Fig I. Comparison of hyperalgesia induced by NKA, SP and CGRP. The data reported are the means for 5 animals per group at 30 rain after the single injections of NKA, SP and CGRP into the rat paw (s.pl.).
paw) or CGRP (0.1 fmol/paw) given 5 times at l0 • i n intervals increased the potency and time course of the hyperalgesic response to these substances in the paw. Furthermore, 5 h after the multiple injections of NKA or 3 h after CGRP, when the hyperalgesia was no longer present, challenge with a dose of 0.4 fmol/paw of NKA or 0.1 fmol/paw of CGRP, respectively, induced pro-
22 * * * ~ ( ~ , • NKA 0.4lmol
~" 20 •Saline *** O NKA 0.4fmol (5 times)
18 El Saline (5 times) =
c 16 O
14 *** ***
12 *** - .m_ 10
® 8 O)
"~ 6 O.
4 . c
.-_>' 4= ":!I , . - - T , ,
I I I I I I
0 1 2 3 4 5 6
Time (h)
Challenging dose of NKA (0.4fmol)
Fig. 2. Effect of multiple injections of NKA followed by challenging dose of NKA in sensitized and saline treated paws. NKA (0.4 fmol/ paw s.pl.) was injected 5 h after 5 injections of NKA (0.4 fmol/paw s.pl., at 10 • i n intervals) or 5 h after 5 injections of saline. The data reported are the means+ S.E.M. for 5 animals per group.
nounced hyperalgesia as we have described recently for SP [14]. In animals receiving multiple saline injections (control) the challenging dose of NKA or CGRP failed to induce significant hyperalgesia (Fig. 2). The sensitiz- ing effect on hyperalgesia due to the exposure of the paw to repeated injections of NKA or CGRP was not limited to challenge only with NKA or CGRP themselves. Low doses of other hyperalgesic agents such as dopamine (6.5. fmol/paw) or prostacyclin (1.5 fmol/paw) also induced pronounced hyperalgesia when tested 5 h after the sensitizing regime of NKA or 3 h after CGRP but not after multiple saline dosing (results not shown).
Administration to the paw of the SPAt (3.34 nmol/ paw) 10 • i n before multiple injections of NKA (0.4 fmol/paw) or CGRP (0.1 fmol) 5 times at 10 min inter- vals) reduced the hyperalgesia from 18.4+0.09 to 15.76+0.59 (14.4~;) and 7.56_+0.49 to 0.76_+1.31 (89.9%), respectively, at 30 • i n after the last injection of NKA and CGRP. The same dose of the SPAt inhibited totally the development of sustained hyperalgesia induced by SP [14].
It has been suggested that substance P, NKA and CGRP are implicated in the transmission of nociceptive information in the spinal cord [8, 16]. Recently, we pos- tulated that SP would participate as a neuromodulator in peripheral inflammatory pain [14]. The present results show that, in this system, NKA is 10 times more potent than SP in inducing hyperalgesia, which is 500 times more potent than CGRP, suggesting that NKA and SP
22 f ¢n 20
• CGRP 0.1 fmol 18 [- • Saline
'= 1R L o CGRP 0.1 fmol (5 times) ~ ; 4 ] - n Saline (S times)
12 • ***
I : -
_
-4 ' I I I I
0 1 2 3 4
Time (h)
Challenging dose of CGRP 0.1 fmol
Fig. 3. Effect of multiple injections of CGRP followed by challenging dose of CGRP in sensitized and saline treated paws. CGRP (0.1 fmol/ paw s.pl.) was injected 3 h after the 5 injections of CGRP (0.1 fmol/ paw s.pl., at 10 • i n intervals) or 3 h after 5 injections of saline. The
data reported are the means+ S.E.M. for 5 animals per group.

but not C G R P could have an impor tan t role in acute
hyperalgesic conditions. This is suppor ted by the evi-
dence that intrathecal injections o f SP and N K A but no t C G R P caused nociceptive behavioural responses in mice
[5]. When N K A or C G R P were administered to the paw
at subthreshold doses (0.4 fmol /paw or 0.1 fmol, 5 times, 10 min intervals, respectively) the development o f an in-
tense and long-lasting hyperalgesic response local to
that paw was observed following the final N K A or C G R P injection. Five hours after the last injection o f
these multiple dosing regimes o f N K A or 3 h after
C G R P , at a time when the sustained hyperalgesic effect o f that regime had subsided, the paw was still sensitized
as revealed by challenge with the previous subthreshold dose o f N K A or C G R P . Recently, we showed that
repeated administrat ion o f subthreshold doses o f SP can sensitize the paw to SP and to the act ion o f other media-
tors [14]. We have now shown that the rat paw sensitized to N K A or C G R P also responds to challenges with small doses o f other substances that p robably participate in peripheral inf lammatory hyperalgesia such as prosta-
cyclin [3] or dopamine [12]. Therefore, low endogenous levels o f N K A or C G R P could sensitize nociceptors not
only to its own effect but also to that o f different inflam- ma to ry mediators.
The effect o f a single injection o f C G R P or N K A was not inhibited by the SP antagonis t [D-Arg 1, D-Pro 2, D- Trp 7'9, Leu I I]Sp suggesting that N K A or C G R P are act-
ing at their own receptor. The fact that the substance P antagonis t [D-Arg l, D-Pro 2, D-Trp 7,9, LeuU]SP inhibited
totally and partially the development o f sustained hyper- algesia induced by C G R P and N K A respectively, sug- gests that par t o f the mechanism o f sensitization o f noci- ceptors involves part icipat ion o f SP receptors. Whether there is an involvement o f adenylate cyclase and 'de
novo ' protein synthesis in the mechanism o f sensitization o f nociceptors as has been suggested recently in similar response induced by some inf lammatory mediators [1] remains to be determined.
There is evidence that C G R P potentiates the release o f SP f rom the dorsal horn o f the spinal cord in rats [14].
We showed recently, that a SP antagonist inhibited the development o f sustained hyperalgesia induced by multi- ple injections o f prostaglandin E2 [12]. Fur thermore , the development o f sustained hyperalgesia induced by differ- ent hyperalgesic substances is inhibited in capsaicin treated rats [13]. The results presented in this paper sug-
gest that N K A and C G R P could have an impor tan t role along with SP as neurotransmit ters or neuromodula to rs mainly in chronic inf lammatory pain.
1 Ferreira, S.H., Lorenzetti, B.B. and Campos, D.I., Induction, blockade and restoration of a persistent hypersensitive state, Pain, 42 (1990) 356-371.
51
2 Ferreira, S.H., Lorenzetti, B.B. and Correa, F.M.A., Central and peripheral antialgesic action of aspirinqike drugs, Eur. J. Pharma- col., 53 (1978) 39-48.
3 Ferreira, S.H., Nakamura, M. and Castro, M.S.A., The hyperalge- sic effects of prostacyclin and prostaglandin E2, Prostaglandins, 16 (1978) 31-37.
4 Fleetwood-Walker, S.M., Mitchell, R., Hope, P.J., EI-Yassir, N. and Molony, V., Neurokinin A as a transmitter of primary after- ents involved in thermal nociception. In J.L. Henry et al. (Eds.), Substance P and Neurokinins, Springer, New York, 1987, pp. 291- 293.
5 Gamse, R. and Saria, A., Nociceptive behaviour after intrathecal injections of substance P, neurokinin A and calcitonin gene-related peptide in mice, Neurosci. Lett., 70 (1986) 143-147.
6 Gibson, S.J., Polak, J.M., Bloom, S.R., Sabate, I.M., Mulderry, P.M., Ghatei, M.A., McGregor, G.P., Morrison, J.F.B., Kelly, J.S., Evans, R.M. and Rosenfeld, M.G., Calcitonin gene-related peptide immunoreactivity in the spinal cord of man and of eight other species, J. Neurosci., 4 (1984) 3101-3111.
7 Henry, J.L., Krnjevic, K. and Morris, M.E., Substance P and spi- nal neurones, Can. J. Physiol. Pharmacol., 53 (1975) 423~32.
8 Henry. J.L. and Salter, M.W., Neurokinin A excites nociceptive and non-nociceptive dorsal horn neurons in the cat spinal cord. In J.L. Henry et al. (Eds.), Substance P and Neurokinins, Springer, New York, 1987, pp. 279-281.
9 Hokfelt, T., Kellerth, J.-O., Nilsson, G. and Pernow, B., Experi- mental immunohistochemical studies on the localization and distri- bution of substance P in cat primary sensory neurons, Brain Res., I00 (1975) 235-252.
10 Kuraishi, Y., Nanayama, T., Ohno, H., Minami, M. and Satoh, M., Antinociception induced in rats by intrathecal administration of antiserum against calcitonin gene-related peptide, Neurosci. Lett., 92 (1988) 325-329.
11 Lembeck, F., Zur Frage der Zentralen Ubertragung afferenter Im- pulse. III. Mitteilung. Das Vorkommen und die Bedeutung der Substanz P in den dorsalen Wurzeln des Ruckenmarks, Arch. Exp. Pathol. Pharmakol., 219 (1953) 197-213.
12 Nakamura, M. and Ferreira, S.H., A peripheral sympathetic com- ponent in inflammatory hyperalgesia, Eur. J. Pharmacol., 135 (1987) 145-153.
13 Nakamura-Craig, M. and Follenfant, R.L., Possible involvement of sensory neuropeptides in the mechanisms of sustained hyperal- gesia - - a new method to study chronic pain, submitted for publi- cation (1990).
14 Nakamura-Craig, M. and Smith, T.W., Substance P and peripheral inflammatory hyperalgesia, Pain, 38 (1989) 91-98.
15 Oku, R., Satoh, M., Fujii, N., Otaka, A. Yajima, H. and Takagi, H., Calcitonin gene-related peptide promotes mechanical nocicep- tion by potentiating release of substance P from the spinal dorsal horn in rats, Brain Res., 403 (1987) 350-354.
16 Solomon, R.E., Aimone, L.D., Yaksh, T.L. and Gebhart, G.F., Calcitonin gene-related peptide and substance P: co-release evoked by potassium or capsaicin from dorsal and ventral rat spinal cord in vitro and its modulation by analgesic drugs. Florence 89: An In- ternational Workshop on Sensory Neuropeptides. Florence, Italy, Abstracts, 1989.
17 Vaught, J.L., Substance P antagonists and analgesia: a review of the hypothesis, Life Sci., 43 (1988) 1419-1431.
18 Yaksh, T.L., Jessell, T.M., Gamse, R., Mudge, A.W. and Leeman, S.F., Intrathecal morphine inhibits substance P release from mam- malian spinal cord in vivo, Nature, 286 (1980) 155-156.
19 Woolf, C. and Wiesenfeld-Hallin, Z., Substance P and calcitonin gene-related peptide synergistically modulate the gain of the noci- ceptive flexor withdrawal reflex in the rat, Neurosci. Lett., 66 (1986) 226-230.





























