Embed Size (px)
Citation preview

J. PlantPhysiol. Vol. 137. pp. 323-326(1991}
Introduction
Effect of Nicotianamine on Iron Uptake and Citrate Accumulation in Two Genotypes of Tomato, Lycopersicon esculentum Mill. *
AXEL PICHI, GUNTER SCHOLZl, and KARLHEINZ SEIFERr
I Institut fUr Genetik und Kulturpflanzenforschung, Corrensstr. 3, 0-4325 Gatersleben, Germany
2 Institut fUr Biochemie der Pflanzen, Weinberg 3,0-4050 Halle (Saale), Germany
Received March 30, 1990 . Accepted August 8, 1990
Summary
The nicotianamine-Iess tomato mutant chloronerva accumulated large amounts of iron and citrate in different organs under normal iron supply of 10 ~ FeEDTA in the nutrient solution. A similar accumulation of citrate was obtained by the wild-type «Bonner Beste» only under iron deficiency and correspondingly low iron concentrations in the same organs.
Application of nicotianamine (NA) to the leaves of the mutant led to a decrease both of iron and citrate concentrations down to the levels of the wild-type.
The results are discussed in terms of a possible activation of iron by NA, resulting in the regulation of iron-dependent processes at the level of gene expression or of iron-requiring enzymes involved in citrate metabolism.
Key words: L ycopersicon esculentum Mill. cv. Bonner Beste, chloronerva, aconitase, citrate, zron, mcotianamine, organs, quantitative analysis, symplast.
Abbreviations: ChIn = mutant chloronerva; EDTA tianamine.
ethylenediaminetetra-acetate; NA OlCo-
as well as retarded growth (<<apparent iron deficiency», Scholz et al. 1988, Stephan and Prochazka 1989).
The nicotianamine-Iess tomato mutant chloronerva accumulates large amounts of iron in roots and shoots due to a defect in its iron uptake regulatory system (Scholz et al. 1985). Under normal iron supply the iron deficiency response mechanisms of strategy I, as e.g. enhancement in proton extrusion by roots and reductase activity on their surface, remain on a high level in spite of high iron concentrations in roots and shoots (Stephan and Grlin 1989). At the same time plants exhibit visual iron deficiency symptoms, such as excessive branching of roots and swelling of root tips
After supply of nicotianamine (NA) to the shoots or the roots of the mutant these morphological and physiological characters are shifted more or less completely in the direction of the wild-type, «Bonner Beste" ("phenotypical normalization», Scholz et a1. 1985, Stephan and Grlin 1989). Therefore, the existence of a NA-dependent «iron sensor» or gene repressor has been assumed, which triggers the activity of iron deficiency response mechanisms and hence, among others, prevents the excessive uptake rate of iron (Bienfait 1987, Scholz et al. 1988).
* Part 39 in the series" The <Normalizing Factor, for the Tomato Mutant chloronerva». For part 38 see Stephan and Scholz, J. Plant Physiol. 136, 631-634 (1990).
© 1991 by Gustav Fischer Verlag, Stuttgart
It is known for a long time, that organic acids, mainly citric and malic acid, are accumulated in plants grown under limited iron supply (Iljin 1951, De Kock 1958, Brown et a1. 1972, Landsberg 1981), although no data have been presented by these authors on organic acids in relation to iron concen-

324 AxEL PICH, GtlNTER SCHOLZ, and KAIu.HEINZ SEIFERT
o 5 12 22
~dark--+-- light/dark 16/8h, 25/18 OC ---1
I--water --+- half concentrated -+- Full nutrient ------t nutrient solution solution
t--O 5 I 0 ____ 30---1
f-- 1.8,Almoles/plant---i
time (days)
light/temper~ure regime
nutrition
iron supply (,uM )
NAsupply Fig. 1: Time course of the experimental program.
Table 1: Effects of iron supply and NA on the fresh weight (mg) of 22-day-old seedlings. Plants were grown without iron or with 10 pM FeEDTA in nutrient solution. Chloronerva was treated with 50 pM NAA solution via the leaves (1.8 J.'mol/plant). Results are means of six plant each ± standard deviation. (BB: wild-type Bonner Beste, chin: mutant chloronerva).
cotyledons leaves shoot tips axis roots
BB, 10,M FeEDTA 212 ±26 1180 ± 189 357±28 816 ± 81 548 ±78 BB, without iron 202 ± 16 665 ± 47 232±23 564±62 524±26 chin, 10pM FeEDTA 213 ±36 763 ± 70 320 ±42 517 ± 79 372 ± 38 chin, 10 pM FeEDTA + NA 222±40 975± 100 556±43 688 ± 81 453 ±66
trations within leaves. It was therefore imponant to investigate whether in chloronerva the level of citric acid in different organs is under the same NA-dependent control as the regulation of iron uptake and transpon into the shoot.
In the present paper we show that citrate accumulation is another indicator of «apparent iron deficiency» in the NAless tomato mutant and that exogenous NA is effective in restoring normal citrate levels in different organs.
Materials and Methods
Plant culture
Seedlings were grown in Hoagland's solution with an addition of FeEDTA and trace elements as described earlier (Scholz et al. 1987). The light/dark regime was 16/8 h with a temperature of 25/18 0c. The light intensity was 330pMm-2s- 1 and the relative humidity was about 65 %. On the 12th day 20 plants each were transferred to vessels with 2.5 L nutrient solution containing 0, 10 or 30 I'M FeEDTA (Fig. 1). This nutrient solution was replaced every 3 days.
NA supply
A 5xl0- sM NA solution with 0.05% Tween 20 (Atlas Goldschmidt GmbH, Essen, FRG) added as a wetting agent was supplied to the leaves of the mutant by a smooth brush five times per day. Over a period of 10 days each mutant received approximately 1.8pMNA.
NA was isolated from Phaseolus vulgaris 1. seeds according to Schlesier et al. (1984).
Plant growth analysis
Twenty two days after sowing, when the fourth leaf had developed, plants were harvested. Iron from the root surface was removed by immersion in a solution containing 100 pM Ca(N03h and 100 pM Na2EDTA at 4 °C for 30 min under stirring Qooste and De Bruyn 1979, modified). This procedure was followed by dissection into organs and, after fresh mass determination, by iron or citrate determination.
Iron analysis
Organs were dried and after wet digestion with H20 2/ H2S04, iron was determined with o-phenanthroline according to Humphries (1956).
Citrate determination
Ten grams of fresh material was homogenized in 30 mL perchloric acid for 1 min using an ice-cooled Bi.ihler homogenizer. The homogenate was centrifuged and 20 mL of the supernatant was adjusted to pH 9 with 5 M KOH under stirring. The sample was cooled to 4°C for quantitative precipitation of potassium perchlorate. After 15 min the precipitate was filtered and the filtrate was directly used for the citrate assay by a coupled optical test according to Boehringer (1986).
Results
At the time of harvest the wild-type plant grown without iron showed iron-deficiency symptoms, e.g. chlorosis of young leaves and stunted roots. The differences between wild-type and mutant and the effect of different treatments on fresh weight are presented in Table 1. Compared with the wild-type the mutant fresh weight was much lower with 10 ~ FeEDTA only, but considerably increased with NA treatment.
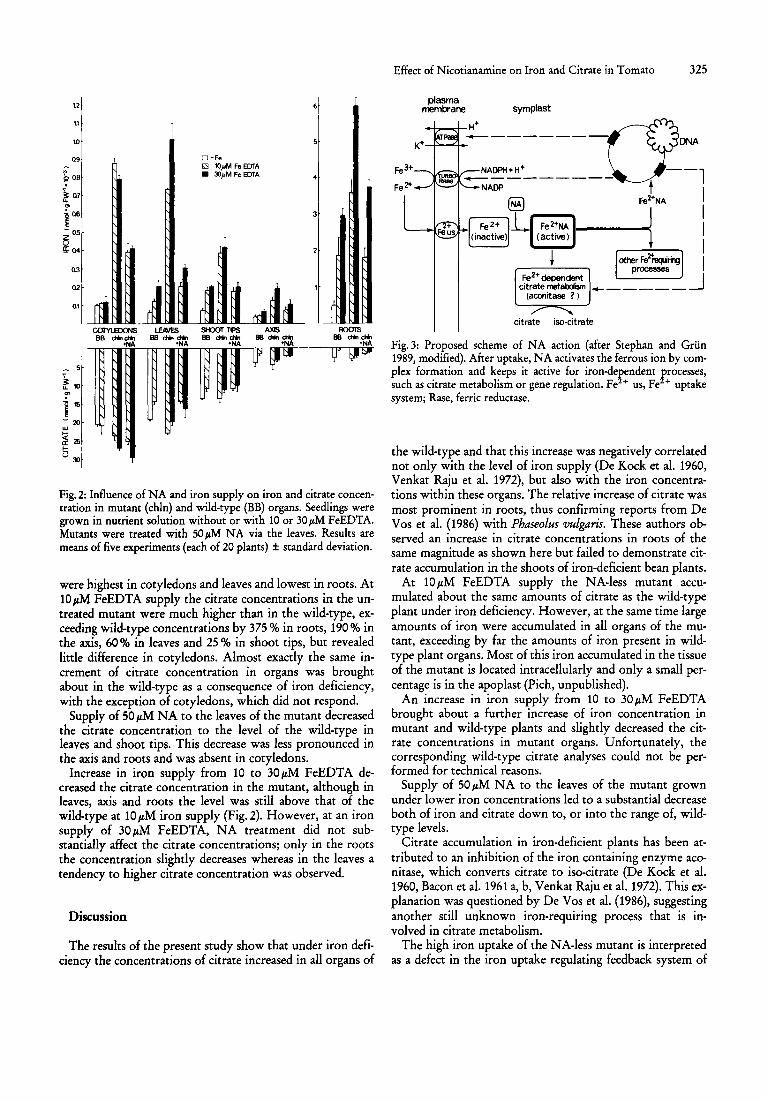
The effects of the various treatments on the concentrations of iron and citrate in different organs of the mutant and its wild-type are shown in Fig. 2. At 10 and 30 ~ FeEDTA supply in the mutant the iron concentrations per fresh weight in all organs exceeded the corresponding concentrations in wild-type organs by up to 800 % (in leaves).
Supply of NA to the mutant leaves significantly decreased the iron concentrations in all organs (except of cotyledons) to levels of the wild-type, thus confirming previous results from our laboratory (Scholz et al. 1985, Stephan and Griin 1989).
In both genotypes citrate concentrations in the fresh weight differed largely between plant organs (Fig.2). They
1.2
t1
to
09
~08 ~Q7 ...
C>
~Q6 ~ 05
g; 04
Q3
0'
COTYI..EOONS lEAVES BB cIWlclWl BB cIWl cIWl
>NIl ·NA
o -Fe ICl 10flM Fe EDTA • 30flM F. EDTA
SHOOT TI'S AXIS BBcIWlclWl lIIc1WlclWl
+NA +NA
4
ROOTS III cIWl cIWl
·NA
Fig. 2: Influence of NA and iron supply on iron and citrate concentration in mutant (chIn) and wild-type (BB) organs. Seedlings were grown in nutrient solution without or with 10 or 30 pM FeEDTA. Mutants were treated with 50 pM NA via the leaves. Results are means of five experiments (each of 20 plants) ± standard deviation.
were highest in cotyledons and leaves and lowest in roots. At 10 ~ FeEDTA supply the citrate concentrations in the untreated mutant were much higher than in the wild-type, exceeding wild-type concentrations by 375 % in roots, 190 % in the axis, 60 % in leaves and 25 % in shoot tips, but revealed little difference in cotyledons. Almost exactly the same increment of citrate concentration in organs was brought about in the wild-type as a consequence of iron deficiency, with the exception of cotyledons, which did not respond.
Supply of 50 ~ NA to the leaves of the mutant decreased the citrate concentration to the level of the wild-type in leaves and shoot tips. This decrease was less pronounced in the axis and roots and was absent in cotyledons.
Increase in iron supply from 10 to 30 ~ FeEDTA decreased the citrate concentration in the mutant, although in leaves, axis and roots the level was still above that of the wild-type at 10 I'M iron supply (Fig. 2). However, at an iron supply of 30 I'M FeEDTA, NA treatment did not substantially affect the citrate concentrations; only in the roots the concentration slightly decreases whereas in the leaves a tendency to higher citrate concentration was observed.
Discussion
The results of the present study show that under iron deficiency the concentrations of citrate increased in all organs of
Effect of Nicotianamine on Iron and Citrate in Tomato 325
plasma membrane
citrate iso-citrate
Fig.3: Proposed scheme of NA action (after Stephan and Gron 1989, modified). After uptake, NA activates the ferrous ion by complex formation and keeps it active for iron-dependent processes, such as citrate metabolism or gene regulation. Fe2 + us, Fe2 + uptake system; Rase, ferric reductase.
the wild-type and that this increase was negatively correlated not only with the level of iron supply (De Kock et al. 1960, Venkat Raju et al. 1972), but also with the iron concentrations within these organs. The relative increase of citrate was most prominent in roots, thus confirming reports from De Vos et al. (1986) with Phaseolus vulgaris. These authors observed an increase in citrate concentrations in roots of the same magnitude as shown here but failed to demonstrate citrate accumulation in the shoots of iron-deficient bean plants.
At 10 ~ FeEDTA supply the NA-Iess mutant accumulated about the same amounts of citrate as the wild-type plant under iron deficiency. However, at the same time large amounts of iron were accumulated in all organs of the mutant, exceeding by far the amounts of iron present in wildtype plant organs. Most of this iron accumulated in the tissue of the mutant is located intracellularly and only a small percentage is in the apoplast (Pich, unpublished).
An increase in iron supply from 10 to 30 ~ FeEDTA brought about a further increase of iron concentration in mutant and wild-type plants and slightly decreased the citrate concentrations in mutant organs. Unfortunately, the corresponding wild-type citrate analyses could not be performed for technical reasons.
Supply of 50 ~ NA to the leaves of the mutant grown under lower iron concentrations led to a substantial decrease both of iron and citrate down to, or into the range of, wildtype levels.
Citrate accumulation in iron-deficient plants has been attributed to an inhibition of the iron containing enzyme aconitase, which converts citrate to iso-citrate (De Kock et al. 1960, Bacon et al. 1961 a, b, Venkat Raju et al. 1972). This explanation was questioned by De Vos et al. (1986), suggesting another still unknown iron-requiring process that is involved in citrate metabolism.
The high iron uptake of the NA-Iess mutant is interpreted as a defect in the iron uptake regulating feedback system of

326 AxEL PICH, GONTER SCHOLZ, and KARLHEINZ SEIFERT
strategy I that is induced in dicots under iron deficiency. As a consequence, among others, the release of protons and reductants into the rhizosphere is enhanced as well as the activity of the .« Turbo,. reductase at the plasmalemma of root cortical cells (Kramer et al. 1980, Romheld et al. 1984, Bienfait 1985, Cakmak et al. 1987). This inducible iron uptake system is inhibited in the wild-type already at a FeEDTA supply of about 5 ~ (Stephan and Prochazka 1989, Stephan and Gron 1989). The biochemical basis of this iron uptake regulation is perhaps the reversible iron saturation of an «iron sensor» (Bienfait et al. 1987) or «iron sensor protein» (Bienfait 1988) or the binding to a (still hypothetical) regulator of genes responsible for strategy I reactions (Scholz et al. 1988).
Nicotianamine is a chelator for Fe2+ at physiological pH (Benes et al. 1983). It is therefore possible that Fe2+NA is the «active» iron that binds to the «iron sensor» or gene regulator and that Fe2+NA is necessary for other iron dependent processes as well (Fig.3). Accordingly, in the NA-Iess mutant the iron in the cells should be «inactive» or at least much less «active» in this respect. Therefore, the mutant suffers «apparent iron deficiency» and fails to repress the inductive uptake processes, unless the regulation is restored by exogenous NA.
Citrate accumulation in the mutant seems to have similar reasons. Since in the absence of NA the intracellular iron is «inactive» with respect to an iron-requiring process of citrate metabolism, citrate accumulates unless NA is supplied via the leaves. Possibly, also in these processes regulation on the genetic level is involved.
Acknowledgements
The authors wish to thank Mrs. Ch. Zappe and Mr. G. Faupel, Gatersleben, for skillful technical assistance.
References
BACON, J. S. D., P. C. DE KOCK, and M. J. PALMER: Aconitase levels in the leaves of iron-deficient mustard plants (Sinapsis alba). Biochern. J. 80, 64-70 (1961 a).
BACON, J. S. D., M. J. PALMER, and P. C. DE KOCK: The measurement of aconitase activity in the leaves of various normal and variegated plants. Biochem. J. 78, 198-204 (1961 b).
BENES, 1., K. SCHREIBER, H. RIpPERGER, and A. KIRCHEISS: Metal complex formation by nicotianamine, a possible phytosiderophore. Experientia 39,261-262 (1983).
BIENFAIT, H. F.: Regulated redox processes at the plasmalemma of plant root cells and their function in iron uptake. J. Bioenergetics and Biomembranes 17, 73-83 (1985).
- Mechanisms in Fe-efficiency reactions of higher plants. J. Plant Nutr. 11, 605-629 (1988).
BIENFAIT, H. F., L. A. DE WEGER, and D. KRAMER: Roots control the development of Fe-efficiency reactions in potato as a response to iron deficiency is located in the roots. Plant Physiol. 83, 244-247 (1987).
BOEHRINGER: UV-method for the determination of citric acid in food stuffs and other materials. Mannheim (1986).
BROWN, J. c., J. E. AMBLER, R. L. CHANEY, and C. D. Fay: Differential responses of plant genotypes of micronutrients. 1iJ.: Micronutrients in Agriculture (MORTVEDT, J. J., P. M. GIORDANO, and
W. L. LINDSAY, eds.) Soil Sci. Am., Madison, Wisconsin, 385-418 (1972).
BROWN, J. C. and R. L. CHANEY: Effect of iron on the transport of citrate into the xylem of Soybeans and Tomatoes. Plant Physiol. 47, 836-840 (1971).
CAKMAK, I., D. A. M. VAN DE WETERING, H. MARSCHNER, and H. F. BIENFAlT: Involvement of superoxide radical in extracellular ferric reduction by Iron-deficient bean roots. Plant Physiol. 85, 310-314 (1987).
DE KOCK, P. c.: The nutrient balance in plant leaves. Agric. Progress 33,88-95 (1958).
DE KOCK, P. c., A. HALL, and M. MAc DONALD: A relation between the ratios of phosphorus to iron and potassium to calcium in mustard leaves. Plant Physiol. 12, 128 -142 (1960).
DE Vos, R. C., H. J. LUBBERDING, and H. F. BIENFAIT: Rhizosphere acidification as a response to iron deficiency in bean plants. Plant Physiol. 81, 842-846 (1986).
HUMPHRIES, E. c.: Mineral components and ash analysis. In: Modern Methods of Plant Analysis (PEACH, K. and M. V. TRACEY, eds.), 1, 468-502 (1956).
lLJIN, W. S.: Metabolism of plants affected with lime-induced chlorosis (Calciose). II. Organic acids and carbohydrates. Plant and Soil 3, 339-351 (1951).
JOOSTE, J. H. and J. A. DE BRUYN: The dual mechanism of iron absorption in bean root and leaf tissues. S. Afr. J. Bot. 45, 249-256 (1979).
KRAMER, D., V. ROMHELD, E. LANDSBERG, and H. MARsCHNER: Induction of transfer-cell formation by iron deficiency in the root epidermis of Helianthus annuus L. Planta 147, 335-339 (1980).
LANDSBERG, E. c.: Organic acid synthesis and release of hydrogen ions in response to Fe-deficiency stress of mono- and dicotyledonous plant species. J. Plant Nutr. 3,579-591 (1981).
ROMHELD, V., C. MOLLER, and H Marschner: Localization and capacity of proton pumps in root of intact sunflower plants. Plant Physiol. 76,603-606 (1984).
SCHLESIER, G., A. RUDOLPH, G. SCHOLZ, and K. SEIFERT: Method for extraction of nicotianamine from seed. GDR-Patent 226880, Int. Cl. C 07 D 205 104. - C.A. 104: P 66 141 u (1984).
SCHOLZ, G., R. BECKER, U. W. STEPHAN, A. RUDOLPH, and A. PICH: The regulation of iron uptake and possible functions of nicatianamine in higher plants. Biochem. Physiol. Pflanz. 183, 257 -269 (1988).
SCHOLZ, G., G. SCHLESIER, and K. SEIFERT: Effect of nicotianamine on iron uptake by the tomato mutant «chloronerva'l>. Physiol. Plant. 63, 99-104 (1985).
SCHOLZ, G., K. H. SEIFERT, and M. GRON: The effect of nicatianamine on the uptake ofMn2+; Zn2 +, Cu2 +, Rb+ and POlby the tomato mutant chloronerva. Biochem. Physio!. Pflanz. 182, 189-194 (1987).
SCHOLZ, G., K. SEIFERT, and K. SCHREmER: Nicotianamine increases the uptake of FeEDDHA by plants at micromolar iron concentration. Biochem. Physiol. Pflanz. 180, 397 - 400 (1985).
STEPHAN, U. W. and M. GRON: Physiological disorders of the nicatianamine-auxotroph tomato mutant chloronerva at different levels of iron nutrition. II. Iron deficiency responses and heavy metal metabolism. Biochem. Physiol. Pflanz. 185, 189-200 (1989).
STEPHAN, U. W. and Z. PROCHAZKA: Physiological disorders of the nicotianamine-auxotroph tomato mutant chloronerva at different levels of iron nutrition. 1. Growth characteristics and physiological abnormalities related to iron and nicotianamine supply. Acta Bot. Neerl. 38, 147-153 (1989).
VENKAT RAju, K., H. MARSCHNER, and V. ROMHELD: Effect of iron nutritional status on iron uptake, substrate pH and production and release of organic acids and riboflavin by sunflower plants. Z. Pflanzenern. Bodenk.132, 178-191 (1972).





























