Embed Size (px)
Citation preview

Enzyme and Microbial Technology 33 (2003) 810–818
Effect of polyhydroxylic cosolvents on the thermostability andactivity of xylanase fromTrichoderma reeseiQM 9414
Ana Cobos, Pilar Estrada∗Departamento de Bioqu´ımica y Biolog´ıa Molecular I, Facultad de Biolog´ıa, Universidad Complutense, Ciudad Universitaria, 28040 Madrid, Spain
Received 5 November 2002; received in revised form 23 June 2003; accepted 29 June 2003
Abstract
Xylanase thermoinactivation followed first-order kinetics and the activation energy for the decay was 311 kJ mol−1. Xylanase showedat 60◦C a half-life of 2.7 min in absence of polyols but the addition of 2 M erythritol, xylitol or sorbitol increased it above 8, 32 and112 times, respectively. Moreover, xylanase showed 80% residual activity after 19 h at 60◦C with 3 M sorbitol. The thermodynamicparameters for the inactivation process in absence of additives were calculated and the increase in�G after the addition of polyhydroxyliccosolvents showed parabolic dependence on both the number of hydroxyls per molecule and the polyols concentration. Xylanase protectionby ethylene glycol (EG) and glycerol was the more effective the higher the thermoinactivation temperature. Erythritol and sorbitol didnot inhibit xylanase activity so that neither of them acted as ligand to protect xylanase. Other polyols tested showed competitive-linealinhibition in the beechwood xylan kinetics withKi = 2.13 M for glycerol andKi = 1.14 M for xylitol whereas ethylene glycol showedcompetitive-parabolic inhibition. Thus, they can protect the enzyme not only as polyols, through their interaction with the medium, butalso as ligands.© 2003 Elsevier Inc. All rights reserved.
Keywords:Xylanase; Polyol; Thermoinactivation; Thermostability; Hemicellulase
1. Introduction
Hydrolysis of xylan plays an important role in the break-down process of plant material in nature since xylan isthe major hemicellulose component in plant cell walls[1].The filamentous fungusTrichoderma reeseiproduces ex-tracellular enzymes which degrade xylan into monomers.The major components of this xylanolytic system are xy-lanase (1,4-�-d-xylan xylanohydrolase; EC 3.2.1.8) whichhydrolyses the�-1,4 bonds in the main chain generat-ing a mixture of xylo-oligosaccharides and�-xylosidase(1,4-�-d-xylan xylohydrolase; EC 3.2.1.37) which cleavesoff the terminal xylose units from the non-reducing end ofxylo-oligosaccharides and is rate-limiting in xylan hydrol-ysis [2].
The feasibility of using xylanase for xylan removal is be-ing tested in the food, feed and textile industries. The useof xylanase decreases the viscosity of bread dough increas-ing the specific bread volume[3], improves the digestibil-ity of rye-based diet of broilers[4] and is an alternativemethod to the treatment with sulfuric acid of the textile
∗ Corresponding author. Tel.:+34-91-3944620; fax:+34-91-3944672.E-mail address:[email protected] (P. Estrada).
polyester–cellulose wastes[5]. Moreover, the use of xy-lanases in the pulp and paper industry reduces the amountof chlorine or chlorine dioxide during bleaching[6]. Nowa-days, it is thought that virtually any cellulase-free xylanaseis effective in paper industry and the attention is focused onthe selection of enzymes whose characteristics, principallypH and temperature relationships are suited to pulp treat-ment regimes[7].
The fungusT. reesei(renamed asHypocrea jecorina) pro-duces and secretes an efficient xylanolytic system formed,besides a�-xylosidase and several debranching enzymes, bytwo major xylanases: the acidic XYNI with pI around 5.2,molecular mass of 19 kDa and pH optimum around 4.0 andthe alkaline xylanase II (XYNII) with pI of 9.0, molecularmass between 20 and 21 kDa and pH optimum between 5.0and 5.5[8,9]. Both enzymes belong to Glycoside HydrolaseFamily 11[10,11], their amino acid identity is approximately50%, have two catalytic Glu, one acting as general acid/baseand the other as nucleophile in a double-displacement mech-anism whereby hydrolysis of xylosidic substrates proceedswith retention of the anomeric configuration. The natureof the residue bonded to the acid/base Glu, asparagine inXYNII and aspartic acid in XYNI may explain the differ-ences in their optimum pHs[12]. The substitution of a single
0141-0229/$ – see front matter © 2003 Elsevier Inc. All rights reserved.doi:10.1016/S0141-0229(03)00220-5

A. Cobos, P. Estrada / Enzyme and Microbial Technology 33 (2003) 810–818 811
Nomenclature
Ead activation energy for the decayEG ethylene glycolD decimal reductionkd first-order reaction constantKi inhibition constantDNS dinitrosalicylict1/2 half-lifeX xylanase
amino acid in xylanase fromBacillus circulanschangedits optimum pH[13]. XYNII has, in its turn, several iso-forms [14]. Given the relevance of the xylanases stabilityin industry, the thermostability ofT. reeseiXYNII has beenstudied by electrospray ionization Fourier-transform andDLS [15] and designed mutations have been introduced inorder to increase its thermal stability[16]. However, wehave no knowledge that xylanase stability from mesophilicmicroorganisms has been achieved through the addition ofwell-known stabilizing molecules such as polyols.
We have studied the production of cellulolytic enzymesby the fungusT. reeseiQM 9414 grown on wheat strawas sole carbon source[17] and determined the kinetic[18]and chemical mechanism[19] of the �-glucosidase it se-creted. We have also purified the�-xylosidase produced bythe fungus, immobilized it on nylon powder[20], studiedits thermal stability[21] and determined its kinetic[22] andchemical mechanism[23]. We have also purified an alka-line xylanase[24] and the purpose of the present work is tostudy its thermal stability as well as the way that additivessuch as polyols affect it.
2. Materials and methods
2.1. Materials
Ethylene glycol (EG), erythritol, xylitol, sorbitol,d-xylose, bovine serum albumin, beechwood xylan, oatspelt xylan and 3,5-dinitrosalycilic acid were from Sigma(Spain). DEAE-sepharose CL-6B was from Pharmacia(Sweden) and Ultrogel AcA 44 was from LKB (Sweden).Glycerol was from Fluka (Spain) and all other reagentswere of analytical grade from Merck (Germany).
2.2. Enzyme purification
T. reeseiQM 9414 was cultured on wheat straw as pre-viously described[17]. Xylanase was purified from culturesupernatant[24] following the same procedure describedfor �-glucosidase and�-xylosidase purification[18,24] in-cluding ammonium sulfate precipitation, DEAE-sepharoseCL-6B and Ultrogel AcA 44 cromatographies. Purification
to homogeneity of the enzyme was achieved after the gelfiltration step; the fractions corresponding to the top of theelution peaks of several purification processes were pooledand employed for the experiments. The xylanase fractionhad a concentration of 0.65 mg ml−1 protein and showeda specific activity of 664�mol min−1 mg−1 with oat speltxylan and 1244�mol min−1 mg−1 with beechwood xylanwhen the activity was assayed in standard conditions.
2.3. Analytics
Protein concentration was measured according to Lowryet al. [25]. Bovine serum albumin was used as standard.
Reducing sugars were measured by the Dinitrosalicylic(DNS) reagent method[26] recording the absorbance of thereduced reagent at 640 nm in a Beckman DU-70 spectropho-tometer.d-Xylose was employed as a standard.
2.4. Enzymatic assays
Controls of non-enzymatic hydrolysis were carried outroutinely for each substrate concentration and the corre-sponding absorbance was subtracted from the absorbancemeasured in enzymatic assay tubes. All the experimentswere carried out in triplicate.
Assays were carried out in centrifuge tubes with 2 ml of0.1 M sodium citrate buffer pH 5.0 (standard buffer) con-taining 1.3�g protein and 1% (w/v) beechwood xylan at55◦C for 10 min in a water bath (Selecta). The reaction wasterminated by cooling the samples in ice for 10 min. Then,the samples were centrifuged at 3000 rpm in a desk cen-trifuge (Nahita, 2610) for 10 min. To 1 ml of the supernatant,1.5 ml of DNS was added and the samples were boiledfor 5 min, cooled in water–ice and analysed for reducingsugars.
2.5. Thermoinactivation studies
Thermoinactivation assays were carried out by preheating960�l of standard buffer at the corresponding temperature,then adding 1.3�g protein in 40�l of the same buffer andpreincubating the mixture at the same temperature. Samples(centrifuge tubes containing the mixture) were removed ev-ery 5 min for 60 min when the preincubation took place at55◦C, every minute for 10 min at 60◦C, every half minutefor 2 min at 65◦C and every 15 s for 1.5 min at 70◦C. Aftercooling the samples in ice for 10 min, 1 ml of the standardbuffer containing 2% (w/v) beechwood xylan was addedin order to assay enzymatic residual activity at 55◦C for10 min.
The polyols used to improve the thermostability of theenzyme were preheated in the standard buffer at the cor-responding temperature. Once the enzyme was added, themixture was preincubated at the same temperature. Theconcentration of polyols varied according with their solu-bilities and other preincubation conditions were as stated

812 A. Cobos, P. Estrada / Enzyme and Microbial Technology 33 (2003) 810–818
above except that 55◦C was not employed and sampleswere preincubated longer times than in absence of polyols.
2.6. Analysis of the data
Values of kinetic constants were determined throughnon-lineal regression by fitting initial velocity data to theMichaelis MentenEq. (1) with the SIGMA PLOT 2000program. Competitive inhibition data were fitted toEq. (2).The half-life of the thermoinactivated enzyme was cal-culated according toEq. (3) and the decimal reductionaccording toEq. (4). Data in parabolic plots were fitted tothe polynomial quadraticEq. (5).
v = Vs
Km + s(1)
v = Vs
Km(1 + I/Ki) + s(2)
t1/2 = A − 3.912
kd(3)
D = A − 2.3026
kd(4)
y = y0 + ax+ bx2 (5)
In Eqs. (1) and (2), v and V are initial and maximum ve-locity (�mol min−1 mg−1), Km is the Michaelis constant(mg ml−1), s is substrate concentration (mg ml−1), I is theinhibitor concentration (M) andKi is the inhibition constant(M). In Eqs. (3) and (4), t1/2 is the half-life (min),A is theintersect of the regression line,kd is the thermoinactivationconstant or first-order reaction constant (min−1), D is thedecimal reduction (min) and 3.912 and 2.3026 are the natu-ral logarithms of 50 and 10, respectively. InEq. (5), y andxare the independent and dependent corresponding variables,anda, b andy0 are constants which define the apex of theparabola, giving alsoy0 the intersect of the parabola withthe ordinate axis.
Analysis of the thermoinactivation data allowed to cal-culate the thermodynamic parameters of xylanase denat-uration: the free energy according toEq. (6), enthalpyaccording toEq. (7)and entropy according toEq. (8).
�G = −RTlnkdh
kBT(6)
�H = Ead − RT (7)
�S = �H − �G
T(8)
In Eqs. (6)–(8), �G is free energy (kJ mol−1), �H is en-thalpy (kJ mol−1), �S is entropy (J mol−1 K−1) at the corre-sponding absolute temperatureT (K), h the Planck constant(11.04× 10−36 J min), kB the Boltzmann constant (1.38×10−23 J K−1), R the gas constant (8.314 J mol−1 K−1), kd thethermoinactivation constant (min−1) andEad the energy ofactivation for the decay (kJ mol−1).
3. Results and discussion
3.1. Catalytic characterization of xylanase
TheT. reeseiQM 9414 alkaline xylanase (Mr = 20 kDa)must correspond to the alkaline XYNII described by Tenka-nen et al.[9] in T. reeseiRut C 30. We could not appreciatethrough our purification method any other alkaline xylanaseand this result agrees with a previous report of Lappalainenet al. [14] who found that the strain QM 9414 producedpractically no xylanase III (XYNIII) whereas this enzymeaccounted for 25% of the whole xylanase activity ofT. ree-seiPC-3-7.
The enzyme was characterized and results are summa-rized in the following lines (data not shown). The xylanasepool employed in this study showed no dependence ofthe ionic strength up to 1 M NaCl. Its activity varied lin-early (steady-state conditions) up to 2�g protein and upto 12 min incubation at 55◦C and pH 5.0 with 1% beech-wood xylan. The kinetic parameters determined by fittingthe experimental data (obtained by varying substrate con-centration from 1 to 14 mg ml−1 of beechwood xylan orfrom 1 to 20 mg ml−1 of oat spelt xylan, standard condi-tions) to Eq. (1) were: Km = 12.11 ± 3.1 mg ml−1 andV = 1709± 211�mol min−1 mg−1 for oat spelt xylan(substituted xylan) andKm = 2.79 ± 0.5 mg ml−1 andV = 1621± 96�mol min−1 mg−1 for beechwood xylan(non-substituted xylan). TheV reported previously forXYNII with beechwood xylan[8] agreed with the valuewe found whereas the reportedKm was about 20 timeslower than ours. This difference could be attributed to thefact that the fungus strains are different besides to possiblemethodological and substrate differences.
Xylanase was stable after preincubation (in absence ofsubstrate) at 50◦C for 30 min but lose half its activity whenpreincubation took place at 55◦C. Its optimum tempera-ture was 55◦C and the activation energy calculated fromthe Arrhenius plot in the temperature range 20–55◦C was32.1 kJ mol−1, which is quite small and suggests that thereis not a high energy barrier for catalysis. Therefore, ansmall increase in temperature will increase exponentiallythe number of activated enzyme molecules with an energysame as or over the activation energy and consequentlyable to react. The optimum pH of xylanase was 5.0 andthe enzyme was stable after 30 min preincubation at pH3.5–8.5 at room temperature whereas after 22 h preincuba-tion showed 80% residual activity in the pH range 5.0–8.5which decreased progressively at more acidic pHs at roomtemperature.
3.2. Thermoinactivation studies
Thermal inactivation of xylanase was followed at tem-peratures from 55 to 70◦C. Plots of natural logarithm ofresidual activity (lnv) versus the preincubation time showedfirst-order kinetics with correlation coefficients higher than

A. Cobos, P. Estrada / Enzyme and Microbial Technology 33 (2003) 810–818 813
103 T-1 ( K-1 )
2.90 2.95 3.00 3.05
ln k
d
-6
-4
-2
0
2
t (min)0 1 2
ln v
0
2
4
70ºC
65ºC
(A)
T ( ºC )
55 60 65 70
log
D
-1
0
1
2
z
(B)
Fig. 1. Temperature dependence of the thermoinactivation constant (A)and of the decimal reduction (B) of xylanase. Thermoinactivation assayswere carried out by preincubating at 65 or 70◦C, 1 ml of 0.1 M sodiumcitrate buffer pH 5.0 containing 1.3�g protein. Samples were removedevery 30 s for 2 min or every 15 s for 1.5 min, respectively, cooled andassayed for xylanase activity by addition of 2% (w/v) beechwood xylanin 1 ml of the same buffer followed by incubation at 55◦C for 10 min.ln v is the natural logarithm of the residual percent activity (A, inset).The thermoinactivation process was also carried out at 55 and 60◦C,being samples taken every 5 min for 60 min at 55◦C and every minute for10 min at 60◦C and the temperature dependence of the thermoinactivationconstant determined with an Arrhenius-type equation (A). The decimalreduction was calculated from experimental data withEq. (4) and itstemperature dependence fitted by linear regression (B).
0.98 when fitted by linear regression (Fig. 1A, inset).This indicates that xylanase is irreversibly inactivated by amonomolecular process[27]. Jänis et al.[15] have observedthat about 10–30% of XYNII was dimerized in buffer over50◦C as deduced from ESI spectra, but this small percentmay not be enough to cause a loss of linearity (due to pro-tein aggregation) when our biochemical assays were carriedout. The thermoinactivation constant or first-order reactionconstant (kd, in min−1) was the absolute value of the slopein the regression line. The half-life of the enzyme (t1/2,in min) was calculated by the exponential decay model at
each temperature as described by Cardoso and Emery[28]according toEq. (3).
To determine the activation energy for the decay of xy-lanase (Ead), a semilogarithmic plot of the thermoinactiva-tion constant (kd) obtained at 55, 60, 65 and 70◦C versusabsolute temperature was depicted (Fig. 1A). The fitting ofthe data by linear regression to an Arrhenius-type equationallowed to calculateEad = 311 kJ (the slope multiplied bythe gas constant,R). This is a high value, uncharacteristic ofa covalent reaction, and agrees with the existence of proteinunfolding followed by refolding into new thermodynami-cally stable structure but catalytically inactive[27]. The factthat the present experiments was carried out in the absenceof ligand also supports inactivation being due to a confor-mational process, thusEad should be the energy necessaryto change the enzyme conformation. Jänis et al.[15] havefound that XYNII dimerization was subsequent to the con-formational change of monomers and they suspected that theconformational change was responsible of the activity lossof their enzyme, just as our results seems to point to. Nev-ertheless we cannot discard that covalent changes occur inaddition to unfolding. With regard to data in literature rela-tives toEad of xylanases, 278 kJ mol−1 was theEad found forthe crude xylanase of the thermophilic fungusThermomyceslanuginosusstrain DSM 5826[29] and 102.23 kJ mol−1 atpH 6.5 for the crude xylanase from the SSBP strain of thesame fungus[30].
It is common, when one is working with enzymes em-ployed in alimentary industry, to express their inactivationby using values of parametersD and z [31]. The D ordecimal reduction value (in min) is defined as the timethe enzyme must be preincubated at a given temperatureto maintain 10% residual activity and was calculated withEq. (4). Since xylanase is added to diets for animal feedingand to dough for bread making whose preparation requireshigh temperatures, we found convenient to plot the logDversus temperature inFig. 1B. The calculation ofz (the tem-perature interval required for the thermal inactivation curvetraverses one log cycle) fromFig. 1B showedz = 6.95◦C.
The thermodynamic parameters of the thermal inacti-vation of xylanase:�G (free energy),�H (enthalpy) and�S (entropy), were calculated at 55, 60, 65 and 70◦Cfrom experimental data according toEqs. (6)–(8), re-spectively. The results are summarized inTable 1 wherewe can observe that the enthalpy shows practically no
Table 1Thermodynamic parameters of xylanase inactivation at different temper-atures
Parameter Temperature (◦C)
55 60 65 70
�G (kJ mol−1) 102.21 96.85 93.87 92.77�H (kJ mol−1) 308.40 308.36 308.32 310.26�S (J mol−1 K−1) 628.60 635.17 634.47 634.08
�G was calculated withEq. (6), �H with Eq. (7) and�S with Eq. (8).
814 A. Cobos, P. Estrada / Enzyme and Microbial Technology 33 (2003) 810–818
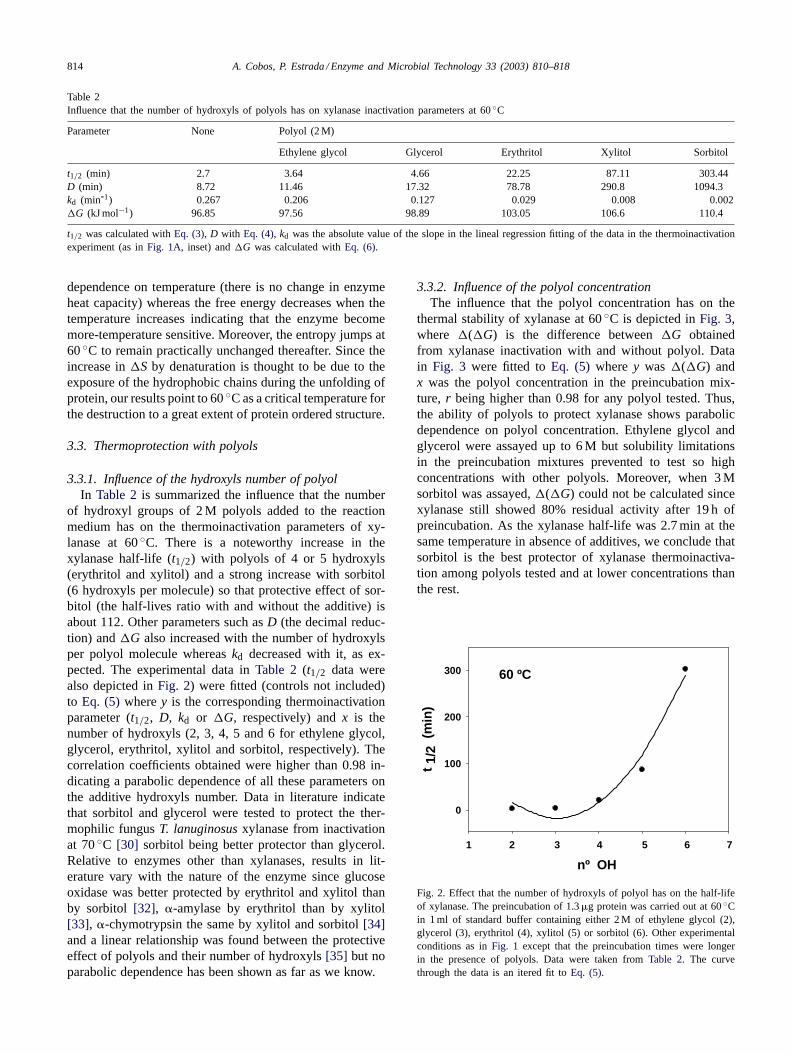
Table 2Influence that the number of hydroxyls of polyols has on xylanase inactivation parameters at 60◦C
Parameter None Polyol (2 M)
Ethylene glycol Glycerol Erythritol Xylitol Sorbitol
t1/2 (min) 2.7 3.64 4.66 22.25 87.11 303.44D (min) 8.72 11.46 17.32 78.78 290.8 1094.3kd (min-1) 0.267 0.206 0.127 0.029 0.008 0.002�G (kJ mol−1) 96.85 97.56 98.89 103.05 106.6 110.4
t1/2 was calculated withEq. (3), D with Eq. (4), kd was the absolute value of the slope in the lineal regression fitting of the data in the thermoinactivationexperiment (as inFig. 1A, inset) and�G was calculated withEq. (6).
dependence on temperature (there is no change in enzymeheat capacity) whereas the free energy decreases when thetemperature increases indicating that the enzyme becomemore-temperature sensitive. Moreover, the entropy jumps at60◦C to remain practically unchanged thereafter. Since theincrease in�S by denaturation is thought to be due to theexposure of the hydrophobic chains during the unfolding ofprotein, our results point to 60◦C as a critical temperature forthe destruction to a great extent of protein ordered structure.
3.3. Thermoprotection with polyols
3.3.1. Influence of the hydroxyls number of polyolIn Table 2is summarized the influence that the number
of hydroxyl groups of 2 M polyols added to the reactionmedium has on the thermoinactivation parameters of xy-lanase at 60◦C. There is a noteworthy increase in thexylanase half-life (t1/2) with polyols of 4 or 5 hydroxyls(erythritol and xylitol) and a strong increase with sorbitol(6 hydroxyls per molecule) so that protective effect of sor-bitol (the half-lives ratio with and without the additive) isabout 112. Other parameters such asD (the decimal reduc-tion) and�G also increased with the number of hydroxylsper polyol molecule whereaskd decreased with it, as ex-pected. The experimental data inTable 2 (t1/2 data werealso depicted inFig. 2) were fitted (controls not included)to Eq. (5) wherey is the corresponding thermoinactivationparameter (t1/2, D, kd or �G, respectively) andx is thenumber of hydroxyls (2, 3, 4, 5 and 6 for ethylene glycol,glycerol, erythritol, xylitol and sorbitol, respectively). Thecorrelation coefficients obtained were higher than 0.98 in-dicating a parabolic dependence of all these parameters onthe additive hydroxyls number. Data in literature indicatethat sorbitol and glycerol were tested to protect the ther-mophilic fungusT. lanuginosusxylanase from inactivationat 70◦C [30] sorbitol being better protector than glycerol.Relative to enzymes other than xylanases, results in lit-erature vary with the nature of the enzyme since glucoseoxidase was better protected by erythritol and xylitol thanby sorbitol [32], �-amylase by erythritol than by xylitol[33], �-chymotrypsin the same by xylitol and sorbitol[34]and a linear relationship was found between the protectiveeffect of polyols and their number of hydroxyls[35] but noparabolic dependence has been shown as far as we know.
3.3.2. Influence of the polyol concentrationThe influence that the polyol concentration has on the
thermal stability of xylanase at 60◦C is depicted inFig. 3,where �(�G) is the difference between�G obtainedfrom xylanase inactivation with and without polyol. Datain Fig. 3 were fitted toEq. (5) wherey was �(�G) andx was the polyol concentration in the preincubation mix-ture, r being higher than 0.98 for any polyol tested. Thus,the ability of polyols to protect xylanase shows parabolicdependence on polyol concentration. Ethylene glycol andglycerol were assayed up to 6 M but solubility limitationsin the preincubation mixtures prevented to test so highconcentrations with other polyols. Moreover, when 3 Msorbitol was assayed,�(�G) could not be calculated sincexylanase still showed 80% residual activity after 19 h ofpreincubation. As the xylanase half-life was 2.7 min at thesame temperature in absence of additives, we conclude thatsorbitol is the best protector of xylanase thermoinactiva-tion among polyols tested and at lower concentrations thanthe rest.
nº OH
1 2 3 4 5 6 7
t 1/
2 (
min
)
0
100
200
300 60 ºC
Fig. 2. Effect that the number of hydroxyls of polyol has on the half-lifeof xylanase. The preincubation of 1.3�g protein was carried out at 60◦Cin 1 ml of standard buffer containing either 2 M of ethylene glycol (2),glycerol (3), erythritol (4), xylitol (5) or sorbitol (6). Other experimentalconditions as inFig. 1 except that the preincubation times were longerin the presence of polyols. Data were taken fromTable 2. The curvethrough the data is an itered fit toEq. (5).

A. Cobos, P. Estrada / Enzyme and Microbial Technology 33 (2003) 810–818 815
[Polyol] (M)
0 2 4 6 8
(G
) (
kJ m
ol-
1 )
0
2
4
6
8
10
12
14
16
EthyleneGlycol
Glycerol
Sorbitol
Erythritol
Xylitol
Fig. 3. Effect of polyhydroxylic cosolvents concentration on xylanasethermostability. The preincubation of 1.3�g protein was carried out at60◦C in 1 ml of standard buffer containing the polyol concentrationindicated in the plot. Other experimental conditions as inFig. 1 exceptthat the preincubation times were longer. The enzymatic assay was carriedout as inFig. 1 by addition of 1 ml standard buffer with substrate so thatthe final polyol concentration was half the indicated in the plot.�G ofxylanase denaturation was calculated withEq. (3) and �(�G) was thedifference between�G with polyol and without it (control value, zeropoint in the plot).
3.3.3. Influence of the preincubation temperatureTo know if the protection by polyols varies with the
preincubation temperature, we have carried out the thermo-protection experiments of xylanase with ethylene glycol andglycerol at 60, 65 and 70◦C. After calculating�(�G) fromthe experimental data, results were plotted versus tempera-ture in Fig. 4. We observe a linear relationship (r > 0.985)in Fig. 4A and Bfor any polyol concentration tested, aswas expected from gazing atEq. (6). But when we considerthe slopes of the regression lines, all of them have positivevalues, which indicates that the higher the preincubationtemperature the better the enzyme is protected from ther-moinactivation by both polyols. Obviously, the effect of thepolyol concentration (2, 4 and 6 M) on�(�G) depicted pre-viously inFig. 3is also noticed inFig. 4for any temperaturetested.
Besides, as we wanted to calculate thez value for xy-lanase inactivation in the presence of ethylene glycol andglycerol, we plotted logD versus temperature (not shown),just as we have done inFig. 1Bbut now using data obtainedin the experiments described above and depicted inFig. 4.We obtained linear regression lines withr ≥ 0.98 only withethylene glycol 2 M (z = 24.81◦C) and with glycerol 6 M(z = 101.21◦C) whereas data with other polyol concentra-tions could not be fitted. The inspection of bothz values,with regard to control in absence of polyols (z = 6.95◦C,obtained fromFig. 1B) revealed thatz increased more thanthree-times with ethylene glycol 2 M and more than 14-timeswith glycerol 6 M which may be an interesting result in or-der to stabilize xylanase for poultry diets.
T (ºC)
60 65 70
(G
) (
kJ m
ol-1
)
0
5
10
15
2 M
4 M
6 M(B)
Glycerol
60 65 70
( G
) (
kJ
mol
-1)
0
5
10
15
2 M
4 M
6 M
(A)
Ethylene Glycol
Fig. 4. Effect of preincubation temperature and of ethylene glycol con-centration (A) or glycerol concentration (B) on xylanase thermostability.Experiments were carried out as inFig. 3 but xylanase was also prein-cubated at 65 and 70◦C in the presence of the indicated concentrationsof ethylene glycol (A) or glycerol (B). Other conditions and calculus asin Fig. 3.
3.3.4. Influence of the polyol/protein ratioFinally, we tried to check if the ratio polyol/protein in the
preincubation mixture affected the ability of the polyol toprotect the enzyme. The amount of protein in the preincuba-tion mixture at 65◦C was firstly varied (1.3–5.2) in absenceof polyols (controls) obtaining the same xylanase half-life(t1/2 = 0.74 min) which confirms the absence of aggrega-tion and the monomolecular nature of the process. Then,the amount of protein was varied at a fixed polyol con-centration (2 M glycerol). Preliminary results (not shown)indicate that there is an optimal ratio glycerol/protein(0.77 mol glycerol mg1 protein in 1 ml of preincubationmixture) to get the longest xylanase half-life at 65◦C.
3.4. Effect of polyols on xylanase activity
The effect that polyols have on xylanase activity is shownin Fig. 5. Ethylene glycol, glycerol and xylitol inhibit theenzyme activity whereas erythritol has no effect up to 2 Mand sorbitol increases the enzyme activity over control up to1 M decreasing it thereafter to reach again the control value.

816 A. Cobos, P. Estrada / Enzyme and Microbial Technology 33 (2003) 810–818
[Polyol] (M)0 2 4 6 8
% A
ctiv
ity
0
25
50
75
100
125
Ethylene glycol
Glycerol
XylitolA
Sorbitol
Erythritol
Fig. 5. Effect of polyhydroxylic cosolvents on xylanase activity. Enzymaticassays were carried out at 55◦C for 10 min with 1.3�g protein and 1%(w/v) beechwood xylan in 2 ml of 0.1 M sodium citrate buffer containing0–8 M ethylene glycol (�), 0–5 M glycerol (�), 0–2.4 M erythritol (�),0–3 M xylitol (�), or 0–2.2 M sorbitol (�).
With regard to the threshold value of the inhibitory polyols(C50, molar concentration at which one-half of the activity isobserved) we have calculateC50 = 8 M for ethylene glycol,C50 = 2.6 M for glycerol andC50 = 1.8 M for xylitol. Thisfirst inspection of the effect of inhibitory polyols on activityindicates that there is a relationship between their numberof hydroxyls and their inhibitory power.
To determine the type and extent of xylanase inhibitionby these polyols, we carried out kinetic studies in theirpresence. All three polyols showed competitive inhibitionin the beechwood xylan kinetics, that is, they combinewith the same enzyme form (the free enzyme) as does thesubstrate[36] and this behaviour was expected from in-hibitors possessing hydroxyls and resembling more or lessthe reaction products and/or the substrate domain involvedin binding. Competitive inhibition by ethylene glycol isdepicted inFig. 6A (in double-reciprocal plot to facilitatepattern identification). The fitting of the data by non-linearregression to competitive inhibitionEq. (2)gave a differentKi for each inhibitor concentration so that the inhibitionwas not competitive-linear. The slope replot versus inhibitorconcentrations (Fig. 6A, inset) was fitted toEq. (3) withr = 0.993 which indicates that the inhibition by ethyleneglycol was competitive-parabolic. The parabolic profile ofthe slope replot suggests that two molecules of ethyleneglycol combine with free xylanase (X) to form a dead-endcomplex X·EG2 according to Cleland[36]. The small sizeof this polyol may be responsible for the accommodationof two inhibitor molecules in a pocket designed for onesubstrate molecule. On account of this behaviour, we couldnot determine the true inhibition constant (Ki ) by ethyleneglycol so that the precise extent of its inhibition remains un-known although its threshold (C50 = 8 M, fromFig. 5) is thehighest pointing to ethylene glycol as the weakest inhibitor.
Xylitol was a competitive inhibitor (double-reciprocal plotis shown inFig. 6B) and glycerol was also a competitive
1/s
0.2 0.0 0.2 0.4 0.6 0.8 1.0 1.2
1/v
(x 1
03 )
0
2
4
6
8
10
Control
2 M
4 M
6 M
(A)
[Ethylene glycol] (M)0 2 4 6
slo
pe
(x 1
03 )
0
4
8
1/s
0.4 -0.2 0.0 0.2 0.4 0.6 0.8 1.0 1.2
1/v
(x10
3 )
0
2
4
6
8
10
Control
2 M
1 M
(B)
[Xylitol] (M)-1 0 1 2
slo
pe
(x10
3 )
0
2
4
6
8
Fig. 6. Effect of ethylene glycol (A) and xylitol (B) on the beechwoodxylan kinetics of xylanase. Double-reciprocal plots of beechwood xylankinetics (1–14 mg ml−1) were carried out at 55◦C for 10 min in 2 mlstandard buffer with 1.3�g protein and without (control,�) or with 2 M(�), 4 M (�) and 6 M (�) ethylene glycol (A). Secondary replot ofslopes vs. ethylene glycol concentration (A, inset) was fitted toEq. (5).Same experiment as in (A) but without (�) or with 1 M (�) and 2 M(�) xylitol (B). Secondary replot of slopes vs. xylitol concentration (B,inset) was fitted to linear regression.
inhibitor (not shown) and the fitting of both set of datato Eq. (2) allowed calculateKi since both polyols causedcompetitive-linear inhibitions (Ki = 2.13 M for glycerol andKi = 1.14 M for xylitol). Moreover, to confirm these val-ues, the slope replots versus inhibitor concentration weredrawn and fitted by regression linear (see the slope replotfor xylitol in Fig. 6B, inset; not shown for glycerol). Thus,we could also calculateKi as the positive value of the re-gression line intersect with the abscissa axis in the slopereplot, confirming the values obtained forKi through fittingto Eq. (2). InspectingKi values, xylitol is a more powerful

A. Cobos, P. Estrada / Enzyme and Microbial Technology 33 (2003) 810–818 817
inhibitor of xylanase activity than glycerol and this resultagrees with a published report of Graber and Combes forfungal�-amylase[33].
The ability of polyols to protect enzymes against ther-moinactivation has been related to their ability to decreasethe water activity by reducing the “free” water moleculesthrough water–polyol hydrogen bonding interactions, thusimmobilizing the water medium[34]. Besides, additivescan interact directly with the enzyme causing competitiveinhibition [35] acting as ligands, as the substrate does, andstabilizing the active centre. In relation with xylanase fromT. reeseiQM 9414, erythritol did not inhibited the enzymeactivity whereas sorbitol activated it or had no effect de-pending on its concentration (Fig. 5). So, their ability toprotect xylanase must operate through their interaction withthe medium. Ethylene glycol, glycerol and xylitol are com-petitive inhibitors so that they can partly mimic the substrateor the product that combines with the free enzyme[36]. Inthis sense, Lo Leggio et al.[37] have localized a glycerolmolecule at subsite-1 in the active centre of xylanase 10Afrom Thermoascus aurantiacuswhereas Schmidt et al.[38]observed three glycerol molecules within the active sitegroove, one of them interacting directly with the catalyticglutamate residue. Therefore, we can assume that theyprotect xylanase both by interacting with the medium andadditionally, as ligands, by interacting with xylanase.
XYNII from T. reeseiQM 9414 belongs to family 11[10,11] and must trans-xylosidate as does XYNII fromT. reeseiRut C 30 [9] and cross-linkedT. reeseiXYNIIcrystals [39]. Moreover,�-xylosidase fromT. reeseiQM9414 transfers xylose to sugars and polyols[40]. Therefore,xylanase could also catalysetrans-xylosidation to polyolsmodifying their real concentration and affecting to someunknown extent the straight biochemical explanations basedmainly in the statistical analysis of the data that we haveassumed in this work.
4. Conclusions
A conclusion can be drawn from this study: polyhydrox-ylic cosolvents thermostabilize the alkaline xylanase fromT. reeseiQM 9414 in a way which depends on three ma-jor characteristics of the polyol and/or of the reaction. First,the stabilizing effect measured as�(�G) increases parabol-ically with the number of hydroxyls of the polyol. Sec-ondly, this effect is also parabolically increased when theconcentration of the corresponding polyol is risen. Thirdly,the �(�G) in the presence of ethylene glycol and glyc-erol, increases linearly with temperature for any polyol con-centration tested, that is, the higher the thermoinactivationtemperature, the more effective protectors are both polyols.Relative to the way in which polyols protect xylanase, itappears that, as well as their classical interaction with themedium as polyhydroxylic cosolvents, ethylene glycol, glyc-erol and xylitol might protect it also as ligands since they are
competitive inhibitors of xylanase in the beechwood xylankinetics.
The best result found was 80% residual xylanase activityafter 19 h at 60◦C with 3 M sorbitol in the medium in con-trast with a half-life of 2.7 min that showed xylanase in itsabsence at the same temperature. The increases inz valuewith 2 M glycerol and 6 M ethylene glycol also allow toconsider polyols as potential protective agents of xylanaseinactivation in industry.
References
[1] Biely P. Microbial xylanolytic system. Trends Biotechnol 1985;3:286–90.
[2] Robinson PD. Cellulase and xylanase production byTrichodermareeseiRut C-30. Biotechnol Lett 1984;6:119–22.
[3] Maat J, Roza M, Verbakel J, Stam H, Santos da Silva MJ, Bosse M,et al. Xylanases and their application in bakery. In: Visser J, BeldmanG, Kusters-van Someren MA, Voragen AGJ, editors. Xylans andxylanases. Amsterdam: Elsevier; 1992. p. 371–8.
[4] Van Paridon PA, Boonman JCP, Selten GCM, Geerse C, Barug D,de Bot PHM, Hemke G. The application of fungal endoxylanases inpoultry diets. In: Visser J, Beldman G, Kusters-van Someren, MAVoragen, AGJ, editors. Xylans and xylanases. Amsterdam: Elsevier;1992. p. 371–8.
[5] Ciechanska D. Progress in biomodification of cellulose pulps bycellulases and xylanases. Fibres Textiles East Eur 2000;3:74–8.
[6] Viikari L, Kantelinen A, Sundquist J, Linko M. Xylanases inbleaching: from an idea to the industry. FEMS Microbiol Rev1994;13:335–50.
[7] Garg AP, Roberts JC, McCarthy J. Bleach boosting effect ofcellulase-free xylanase ofStreptomyces thermoviolaceusand itscomparison with two commercial enzyme preparations on birchwoodkraft pulp. Enzyme Microbiol Technol 1998;22:594–8.
[8] Törrönen A, Mach LR, Messner R, Gonzalez R, Kalkkinen N,Harkki A, et al. The two major xylanases fromTrichoderma reesei:characterization of both enzymes and genes. Biotechnology 1992;19:1461–5.
[9] Tenkanen M, Puls J, Poutanen K. Two major xylanases ofTrichoderma reesei. Enzyme Microbiol Technol. 1992;14:566–74.
[10] Henrissat B, Bairoch A. New families in the classification of glycosylhydrolases based on amino acid sequence similarities. Biochem J1993;293:781–8.
[11] http://afmb.cnrs-mrs.fr/CAZY.[12] Törrönen A, Rouvinen J. Structural comparison of two major endo-
1,4-xylanases fromTrichoderma reesei. Biochemistry 1995;34:847–56.
[13] Joshi MD, Sidhu G, Pot I, Brayer GD, Withers SG, McIntosh LP.Hydrogen bonding and catalysis: a novel explanation for how asingle amino acid substitution can change the pH optimum of aglycosidase. J Mol Biol 2000;299:255–79.
[14] Lappalainen A, Siika-Aho M, Kalkkinen N, Fagerström R,Tenkanen M. Endoxylanase II fromTrichoderma reeseihas severalisoforms with different isoelectric points. Biotechnol Appl Biochem2000;31:61–8.
[15] Jänis J, Rouvinen J, Leisola M, Turunen O, Vainiotalo P.Thermostability of endo-1,4-�-xylanase II fromTrichoderma reeseistudied by electrospray ionization Fourier-transform ion cyclotronresonance MS, hydrogen/deuterium-exchange reactions and dynamiclight scattering. Biochem J 2001;356:453–60.
[16] Turunen O, Etuaho K, Fenel F, Vehmaanpera J, Wu X,Rouvinen J, et al. A combination of weakly stabilizing mutationswith a disulfide bridge in the�-helix region of Trichodermareesei endo-1,4-�-xylanase increases the thermal stability throughsynergism. J Biotechnol 2001;88:37–46.

818 A. Cobos, P. Estrada / Enzyme and Microbial Technology 33 (2003) 810–818
[17] Acebal C, Castillon P, Estrada P, Mata I, Costa E, Aguado J, etal. Enhanced cellulase production fromTrichoderma reeseiQM9414 on physically treated wheat straw. Appl Microbiol Biotechnol1986;24:218–23.
[18] Estrada P, Mata I, Dominguez JM, Castillon MP, Acebal C. Kineticmechanism of�-glucosidase fromTrichoderma reeseiQM 9414.Biochim Biophys Acta 1990;1033:298–304.
[19] Mata I, Estrada P, Macarron R, Domı́nguez JM, Castillón MP, AcebalC. Chemical mechanism of�-glucosidase fromTrichoderma reeseiQM 9414. Biochem J 1992;283:679–82.
[20] Dueñas MJ, Estrada P. Immobilization of�-xylosidase fromTrichoderma reesei QM 9414 on nylon powder. BiocatalBiotransform 1999;17:139–61.
[21] Dueñas MJ, Estrada P, Thermoinactivation of�-xylosidaseimmobilized on nylon. In: Ballesteros A, Plou FJ, Iborra JL, HallingPJ, editors. Stability and stabilization of biocatalysts. Amsterdam:Elsevier; 1989. p. 523–8.
[22] Gómez M, Isorna P, Rojo M, Estrada P. Kinetic mechanism of�-xylosidase fromTrichoderma reeseiQM 9414. J Mol Cat B:Enzym 2001;16:7–15.
[23] Gómez M, Isorna P, Rojo M, Estrada P. Chemical mechanism of�-xylosidase fromTrichoderma reeseiQM 9414: pH-dependence ofkinetic parameters. Biochimie 2001;83:961–7.
[24] Mata I, Dominguez JM, Macarron R, Castillon MP, Estrada P.Xylanase and�-xylosidase isolation from cultures ofTrichodermareesei QM 9414. In: Grassi G, editor. Biomass for energy andindustry. Amsterdam: Elsevier; 1990. p. 2283–7.
[25] Lowry OH, Rosebrough NJ, Farr AL, Randall RT. Proteinmeasurement with the Folin phenol reagent. J Biol Chem 1981;193:256–65.
[26] Miller GL. Use of dinitrosalycilic acid reagent for determination ofreducing sugars. Anal Chem 1959;31:426–8.
[27] Klibanov A. Stabilization of enzymes against thermal inactivation.In: Laskin AI, editor. Advances in Applied Microbiology. New York:Academic Press; 1983. p. 1–28.
[28] Cardoso JP, Emery AN. A new model to describe enzyme inactiva-tion. Biotechnol Bioeng 1978;20:1471–7.
[29] Lischnig T, Purkarthofer H, Steiner W. Thermostability ofendo-�-xylanase from the thermophilic fungusThermomyceslanuginosus. Biotechnol Lett 1993;15:411–4.
[30] Singh S, Pillay B, Prior A. Thermal stability of�-xylanases producedby different Thermomyces lanuginosusstrains. Enzyme MicrobiolTechnol 2000;26:502–8.
[31] López P, Burgos J. Lipooxygenase inactivation by manothermo-sonication: effects of sonication, physical parameters, pH, KCl,glycerol and enzyme concentration. J Agric Food Chem 1995;43:620–5.
[32] Ye WN, Combes D, Monsan P. Influence of additives on thethermostability of glucose oxidase. Enzyme Microbiol Technol1988;10:498–502.
[33] Graber M, Combes D. Effect of polyols on fungal alpha-amylasethermostability. Enzyme Microbiol Technol 1989;11:673–7.
[34] Lozano P, Combes D, Iborra JL. Effect of polyols on�-chymotrypsinthermostability: a mechanistic analysis of the enzyme stabilization.J Biotechnol 1994;35:9–18.
[35] Lozano P, Can J, Iborra JL, Manjón A. Influence of polyhydroxyliccosolvents on papain thermostability. Enzyme Microbiol Technol1993;15:868–73.
[36] Cleland WW. Steady state kinetics. In: Boyer P, editor. The enzymes.New York: Academic Press; 1970. p. 1–65.
[37] Lo Leggio L, Kalogiannis S, Eckert K, Teixeira SCM, BhatMK, Andrei C, et al. Substrate specificity and subsite mobilityin Thermoascus aurantiacusxylanase 10A. FEBS Lett 2001;50:303–8.
[38] Schmidt A, Schlacher A, Steiner W, Schwab H, Kratky C. Structureof the xylanase fromPenicillium simplicissimum. Protein Sci1998;7:2081–8.
[39] Leisola M, Jokela J, Finell J, Pastinen O. Simultaneous catalysisand product separation by cross-linked enzyme crystals. BiotechnolBioeng 2001;72:501–5.
[40] Hermann MC, Vrsanská M, Jurickova M, Hirsh J, Biely P,Kubicek CP. �-d-Xylosidase of Trichoderma reeseiis a multi-functional �-d-xylan xylohydrolase. Biochem J 1997;321:375–81.





























