Embed Size (px)
Citation preview

EFFECT OF TEMPERATURE ON THE GROWTH RATE OF GRIFFITHSIA TENUIS C. AGARDH(RHODOPHYTA: CERAMIALES)
William W. REYNOLDS & Martha E. CASTERLIN
The Pennsylvania State University, Wilkes-Barre, Pennsylvania i87o8 U .S.A.
Received April 13, 1977
Keywords : algae, temperature, growth, Grillithsia tenuis
Abstract
Clonal cultures of Griffithsia tenuis were grown for 18 days(Erdschreiber solution, LD 12 : 12, 2200 lux) at 13, 18, 22 and25°C . The optimum temperature for growth (increase in numberof cells) under these conditions was 22'C .
Introduction
Grii ithsia tenuis is one of approximately 30 species ofthis genus found worldwide (Levring, Hoppe & Schmid,1969). It is a filamentous, benthic summer annual,forming loose tufts in very quiet, warm bays (Taylor,1957). Cosmopolitan in warm seas (Dawson, 1966a), G .tenuis is common during the spring and summer inbays of the upper Gulf of California (Mexico) .
The mean sea surface temperature for the upper Gulfranges from WC in January to 30CC in August andSeptember (Thomson, 1969), a range of 17° C. This sug-gests that G. tenuis in the upper Gulf must be relativelyeurythermal, since stenothermal algae have been definedas having a thermal tolerance range of less than io°C(Levring, Hoppe & Schmid, 1969) . G. globulifera cultureshave been reported as having lower and upper tempera-ture tolerances of 8-11 and 32-35°C, respectively (Biebl,1962) .
Water temperature is of great importance to the devel-opment, growth, and geographical distribution of ma-rine algae (Levring, Hoppe & Schmid, 1969), acting as a
Dr. W. Junk b .v. Publishers - The Hague, The Netherlands
Hydrobiologia vol. 56, 3, pag. 225-227, 1977
controlling and/or lethal factor (Fry, 1947) . Most speciesof algae are confined to a 5°C isothere (Dawson, 1966b),undergoing a condition of quiescence or `rigor' in regionsof their occurrence in which the temperature rises aboveor descends below the normal range .
Temperature (as a controlling factor) directly influ-ences the rates of photosynthesis and respiration, therebyaffecting growth (McCombie, 1960 ; Goldman & Carpen-ter, 1974; Pope & Berger, 1974) . Indirectly, temperaturedecreases the solubility of oxygen in water, whereby itmay limit growth by restricting dark-hour respiration(Dawson, 1966b) . Since temperature is so obviously im-portant to the growth of algae, this investigation wasconducted to determine the controlling effect of tem-perature on the growth rate of G . tenuis, and to ascertainthe optimum temperature for growth under controlledconditions .
Methods
Forty sections, 3-8 mm long and containing 1-8 intactcells each, were taken from filaments of G . tenuis grow-ing in a clonal culture at 20°C . The culture was derivedfrom algae collected in the upper Gulf of California(Cholla Bay, Puerto Peiiasco, Sonora, Mexico) in Feb-ruary 1970, when the water temperature was 15 .5°C .
Each section was placed in a standard culture tube halffilled with Erdschreiber solution, and kept in a growthchamber providing 12 hours of 2200 lux illuminationalternating with 12 hours of darkness, for 18 days . Tencultures were grown at each of the following tempera-tures: 13, 18, 22 and 25°C . After 18 days, the numbers of
22 5
226
.10
.08
2I-
.06
z0U
c) .04o!
w
aJX .02
2510
15
20TEMPERATURE (°C)
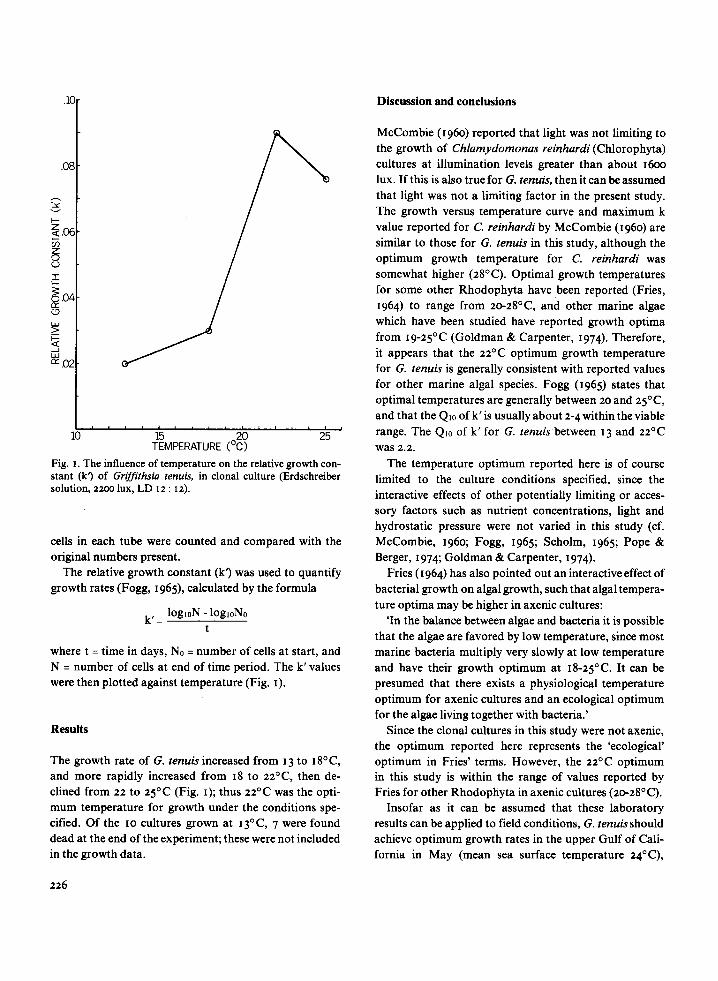
Fig . i . The influence of temperature on the relative growth con-stant (k') of Griffithsia tenuis, in clonal culture (Erdschreibersolution, 2200 lux, LD 12 : 12) .
cells in each tube were counted and compared with theoriginal numbers present.
The relative growth constant (k) was used to quantifygrowth rates (Fogg, 1965), calculated by the formula
k'=logioN - logioNo
t
where t = time in days, N o = number of cells at start, andN = number of cells at end of time period . The k' valueswere then plotted against temperature (Fig . 1) .
Results
The growth rate of G. tenuis increased from 13 to 18°C,and more rapidly increased from 18 to 22°C, then de-clined from 22 to 25°C (Fig. 1) ; thus 22°C was the opti-mum temperature for growth under the conditions spe-cified. Of the 1o cultures grown at 13°C, 7 were founddead at the end of the experiment ; these were not includedin the growth data.
Discussion and conclusions
McCombie (1960) reported that light was not limiting tothe growth of Chlamydomonas reinhardi (Chlorophyta)cultures at illumination levels greater than about 16oolux. If this is also true for G. tenuis, then it can be assumedthat light was not a limiting factor in the present study .The growth versus temperature curve and maximum kvalue reported for C. reinhardi by McCombie (1960) aresimilar to those for G. tenuis in this study, although theoptimum growth temperature for C. reinhardi wassomewhat higher (28°C). Optimal growth temperaturesfor some other Rhodophyta have been reported (Fries,1964) to range from 20-28°C, and other marine algaewhich have been studied have reported growth optimafrom 19-25°C (Goldman & Carpenter, 1974) . Therefore,it appears that the 22°C optimum growth temperaturefor G . tenuis is generally consistent with reported valuesfor other marine algal species . Fogg (1965) states thatoptimal temperatures are generally between 2o and 25°C,and that the Qio of k' is usually about 2-4 within the viablerange. The Q, o of k' for G. tenuis between 13 and 22°Cwas 2.2 .
The temperature optimum reported here is of courselimited to the culture conditions specified, since theinteractive effects of other potentially limiting or acces-sory factors such as nutrient concentrations, light andhydrostatic pressure were not varied in this study (cf .McCombie, 1960 ; Fogg, 1965; Scholm, 1965; Pope &Berger, 1974; Goldman & Carpenter, 1974).
Fries (1964) has also pointed out an interactive effect ofbacterial growth on algal growth, such that algal tempera-ture optima may be higher in axenic cultures :
`In the balance between algae and bacteria it is possiblethat the algae are favored by low temperature, since mostmarine bacteria multiply very slowly at low temperatureand have their growth optimum at 18-25°C. It can bepresumed that there exists a physiological temperatureoptimum for axenic cultures and an ecological optimumfor the algae living together with bacteria .'
Since the clonal cultures in this study were not axenic,the optimum reported here represents the `ecological'optimum in Fries' terms. However, the 22°C optimumin this study is within the range of values reported byFries for other Rhodophyta in axenic cultures (2o-28°C) .Insofar as it can be assumed that these laboratory
results can be applied to field conditions, G. tenuis shouldachieve optimum growth rates in the upper Gulf of Cali-fornia in May (mean sea surface temperature 24°C),

with decreased growth rates from June-October (26-31 ° C) .The death of 7 out of to cultures at 13°C indicates thatG. tenuis may sometimes approach a condition of coldrigor in the upper Gulf of California in January, when themean sea surface temperature is 14'C .
References
Biebl, R . 1962 . Temperaturresistenz tropischer Meeresalgen.Bot. Mar., Vol. 4, pp . 241-254 .
Dawson, E. Y. 1966a. Marine Algae in the Vicinity of PuertoPenasco, Sonora, Mexico . University of Arizona Press,Tucson, 57 pp .
Dawson, E . Y . 1966b . Marine Botany . Holt, Rinehart & Winston,New York, 371 PP .
Fogg, G . E . 1965 . Algal Cultures and Phytoplankton Ecology .University of Wisconsin Press, Madison, 126 pp .
Fries, L . 1964 . Temperature optima of some red algae in axeniccultures . Bot . Mar ., Vol. 4, pp . 12-14 .
Fry, F . E . J . 1947 . Effects of the Environment on Animal Activi-ty . University of Toronto Studies, Biol . Ser . No. 55, 62 pp.(Publ. Ontario Fish. Res. Lab . No 68) .
Goldman, J . C . & Carpenter, E . J . 1974. A kinetic approach tothe effect of temperature on algal growth . Limnol. Oceanogr .,Vol . 19, PP. 756-766 .
Levring, T ., Hoppe, H . A . & Schmid, O . J . 1969 . Marine Algae :A Survey of Research and Utilization. Cram, de Gruyter &Co., Hamburg, 421 pp .
McCombie, A. M. 1960. Actions and interactions of tempera-ture, light intensity and nutrient concentration on the growthof the green alga, Chlamydomonas reinhardi Dangeard . J .Fish . Res . Bd . Canada, Vol. 17, pp . 871-894 .
Pope, D. H . & Berger, L . R . 1974 . Effect of temperature andhydrostatic pressure on algal respiration . Can . J . Bot ., Vol . 52,pp. 2375-2379 .
Scholm, H. 1965. Untersuchungen zur Warmeresistenz vonTiefenalgen . Bot . Mar., Vol. 9, PP. 54-61 .
Taylor, W . R . 1957 . Marine Algae of the Northeastern Coast ofNorth America. University of Michigan Press, Ann Arbor,509 PP .
Thomson, D . A. 1969. 1970 Tide Calendar for the Northern Gulfof California . University of Arizona Press, Tucson, 12 pp .
2 27





























