Embed Size (px)
Citation preview

Ann. appl. B i d . (19941, 124, 451-459 Printed in Great Britain 35 I
Effects of beet cryptic virus infection on sugar beet in field trials
By W S XIE, J F ANTONIW, R F WHITE and T H JOLLIFFE*' Plant Pathology Department, A FRC Institute of Arable Crops Research,
Rothamsted Experimental Station, Harpenden, Herts AL5 2JQ) UK *Hilleshog (UK) Ltd, Sugar Beet Breeding Station, Brooke Lodge,
Brooke, Norwich NR1.5 IJG, UK (Accepted 10 January 1994)
Summary The effects of beet cryptic virus (BCV) infection on sugar beet crops were
investigated in field trials in 1990. Two sugar beet breeding stock lines were screened for infection by BCV. Seed lots containing different proportions of seed infected with BCVl & 2 were obtained by crossing the stock lines and used in field trials at five different sites. Five characteristics of the infected plants were assessed. BCV infection appeared to have no significant effects on the sugar beet crop at four locations which suffered from drought stress but significant effects were found at one site where the crop was grown on grade 1 land with good moisture retention properties. Root yield and sugar yield were reduced by up to 17% and 20%, respectively, by BCV infection.
Key words: Sugar beet, beet cryptic virus, yield
Introduction The cryptic virus group contains viruses which consist of small isometric particles, 30-38
nm in diameter, with genomes comprising 2-3 double-stranded RNA (dsRNA) components (Boccardo, Lisa, Luisoni & Milne, 1987; Milne, 1991). These viruses are found in a wide range of plant families, usually occur at very low concentrations in plants, are transmitted only through seed and pollen and have no known vectors (Boccardo et al . , 1987; Antoniw, White & Xie, 1990).
Beet cryptic virus (BCV), first described by Pullen (1968, 1969), has isometric particles of about 30 nm in diameter and is widespread in different cultivars of Beta vulgaris. BCV was found to consist of a mixture of two viruses, BCVl and BCV2, each with a genome of two dsRNA components (Accotto & Boccardo, 1986; Antoniw, Linthorst, White & Bol, 1986; Natsuaki et al., 1986) and which are serologically unrelated (Kuhne, Stanarius & Kleinhempel, 1987). Recently, a third cryptic virus (BCV3) was found in B. vulgaris, which also seems to be serologically distinct from BCVl and BCV2 (Accotto, Brisco & Hull, 1987; Xie, Antoniw & White, 1989; Xie, 1992).
Plants infected with cryptic viruses are usually free of symptoms. However, radish yellow edge virus (another cryptic virus) appears to be associated with yellowing at the leaf edges of some cultivars of radish (Natsuaki, 1985; Natsuaki, Yamashita, Doi & Yora, 1979; Natsuaki et al . , 1983). Also Kassanis, White & Woods (1977) have reported that infection 'Present address: Plant Breeding Station, Sharpes International Seeds Limited, Boothby Graffoe, Lincoln, Lincs LN5 OLF, UK 0 1994 Association of Applied Biologists

452 W S XIE. J F ANTONIW, R F WHITE AND T H JOLLIFFE
of spinach beet with beet cryptic virus may cause yellowing symptoms or severe stunting of the plant. Further investigation showed that the progenies of infected sugar beet plants contained a larger proportion of severely stunted plants than the progenies of virus-free plants. and that young plants derived from infected parent plants showed a transient mild interveinal chlorosis of the leaves. In addition, plants from crosses involving infected plants had smaller roots on average than those derived from crosses between healthy plants (Kassanis, Russell & White, 1978).
Moir (19830, b ) reported that infection with beet cryptic virus adversely affected the early growth stage of sugar beet. In field experiments in 1980 and 1981, plants infected with beet cryptic virus, when compared with uninfected plants, had a decreased sugar yield of 4.9% and 1U.9%. respectively. However, at that time it was not known that BCV is a mixture of BCVl and BCV2. Now that we can assay sugar beet plants to distinguish infection with BCVl and BCV2, we have re-investigated the effects of infection with these viruses.
In this paper, some experiments on the effects of BCV1, BCV2 and mixed BCVl and BCV2 infections on the sugar beet crop are described. The seedlings from two commercial sugar beet parent lines were screened to obtain seedlings which were: BCV1-, BCV2-, BCVl+2-iniected or healthy. These plants were crossed, and the hybrid seed produced then screened for virus infection before being used in field trials.
Materials and Methods
BCVl and BCV2 infection in seedlings of different sugar beet lines A common sugar beet breeding method involves crossing two monogerm diploid parents
to obtain hybrids (F,) (male sterile). which are subsequently pollinated with a tetraploid pollinator to produce triploid hybrid seed which is used by growers for planting crops. In this experiment, plants of both the male sterile F, hybrid monogerm parent and the tetraploid multigerm pollinator parent (Hilleshog breeding lines HUL88 and SYH88 respectively) were screened for infection with BCVl and BCV2. The object was to find parent plants that when crossed would give four seed types: virus-free, BCVI-, BCV2- and BCVl +Zinfected. ‘To minimise the genetic variability of the seed produced, 20 parent plants of each type were identified.
The seedlings of the sugar beet F, hybrid line and a pollinator line were assayed for BCVl and BCV2 at the third and fourth leaf stage (the first and second true leaves). One of the cotyledons of each seedling was taken, total nucleic acid extracted in microcentrifuge tubes, electrophoresed in 1 % agarose denaturing gels, and transferred onto nylon membranes (Hybond-N, Amersham). These Northern blots were hybridised with 32P-labelled inserts from clone 5B (BCV1-specific) and clone 53 (BCV2-specific) by nick translation as described by Xie et al. (1989).
Screening of BCVl and BCV2 infection in the seed lots obtained from crossing the parent sugar beet lines
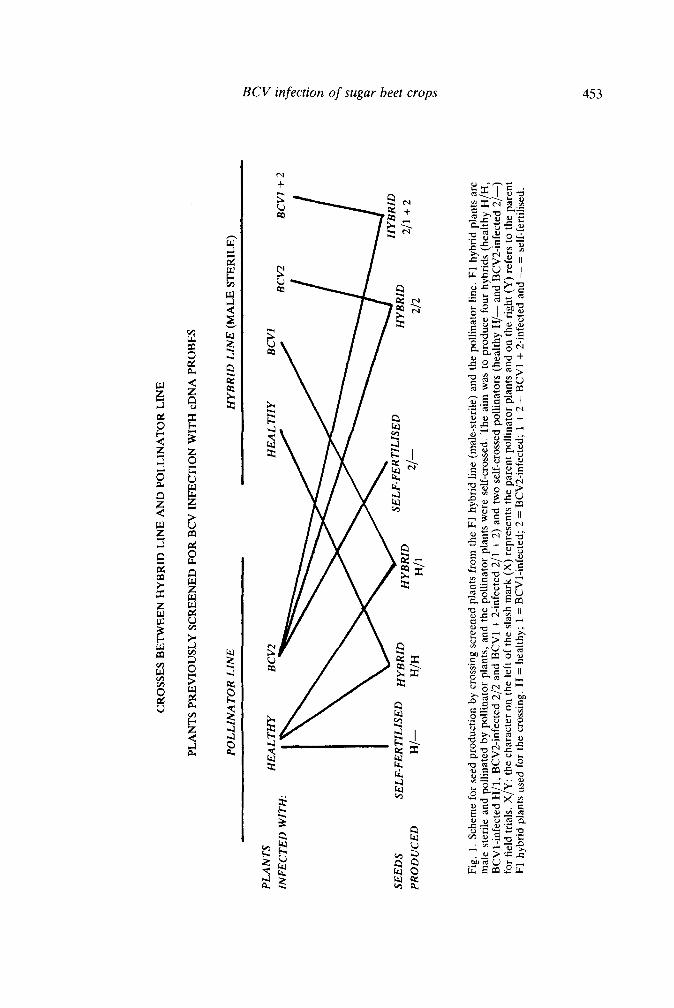
The F, hybrid line plants (male sterile) were pollinated with the corresponding pollinator line (Fig. 1) to produce the seed to be used in the field trials. All of the seed was produced at Brooke near Norwich. Six different seed lots were obtained (0.5-1.0 kg seed/lot), four of which were tripIoid hybrid seeds (healthy H/H, BCV1-infected H/1, BCV2-infected 2/ 2 and BCV1+2 infected 2/1+2) and two lots resulted from self-crossing of the pollinator line plants (healthy H/- and BCV2-infected 2/-). The triploid hybrid and self-crossed seed
CR
OSS
ES
BE
TW
EE
N H
YB
RID
LIN
E A
ND
PO
LL
INA
TO
R L
INE
PLA
NT
S PR
EV
IOU
SLY
SC
RE
EN
ED
FO
R B
CV
IN
FEC
TIO
N W
ITH
cD
NA
PR
OB
ES
POLL
INA
TOR
LIN
E
HY
BR
ID L
INE
(M
AL
E S
TE
RIL
E)
HE
ALT
HY
BC
V2
HE
ALT
HY
BC
Vl
PLA
NTS
IN
FE
CTE
D W
ITH
: B
CV
l +
2
HYB
RID
SE
ED
S PR
OD
UC
ED
Fig.
1. S
chem
e fo
r se
ed p
rodu
ctio
n by
cro
ssin
g sc
reen
ed p
lant
s fr
om th
e F1
hyb
rid l
ine
(mal
e-st
erile
) an
d th
e po
llina
tor l
ine.
F1
hybr
id p
lant
s ar
e m
ale
ster
ile a
nd p
ollin
ated
by
polli
nato
r pl
ants
, and
the
pol
linat
or p
lant
s w
ere
self-
cros
sed.
The
aim
was
to
prod
uce
four
hyb
rids
(hea
lthy
H/H
, B
CV
1-in
fect
ed H
/1,
BC
V2-
infe
cted
2/2
and
BC
Vl +
2-in
fect
ed 2
/1 +
2) a
nd tw
o se
lf-cr
osse
d po
llina
tors
(he
alth
y €
f-
and
BC
V2-
infe
cted
2/-)
fo
r fie
ld tr
ials
. X/Y
: th
e ch
arac
ter o
n th
e le
ft of
the
sla
sh m
ark
(X) r
epre
sent
s th
e pa
rent
pol
linat
or p
lant
s an
d on
the
right
(Y
) ref
ers
to th
e pa
rent
F1
hyb
rid p
lant
s us
ed f
or t
he c
ross
ing.
H =
hea
lthy;
1 =
BC
V1-
infe
cted
; 2
= B
CV
2-in
fect
ed;
1 + 2
= B
CV
l + 2
-infe
cted
and
- =
sel
f-fe
rtilis
ed.
Q- m 2
P
m
w

454 W S XIE, J F ANTONIW, R F WHITE AND T H JOLLIFFE
lots were sown in seed trays and the plants grown in a glasshouse. For each seed lot, total nucleic acids were extracted from one cotyledon leaf of each of 20 individual seedlings, denatured and then slot-blotted onto Hybond-N membranes. The blots were hybridised with inserts from either clone SB (BCV1-specific) or clone 53 (BCV2-specific) by nick translation as described above.
Field trials The six seed lots obtained from crossing the F1 hybrid and the pollinator line plants and
self-crossing within the pollinator line plants were used in trials at five different locations, each replicated four times in a randomised block. Four locations were in normal yield trials, kept isolated and free of other diseases. The fifth was in an area where there was a likelihood of high levels of natural BMYV and BYV infection.
The five sites had the following characteristics: CL/D was on a clay-loam soil type and was affected by drought; FCL/D was on a very fine clay loam-soil type and was affected by drought; HC/ED was on a very heavy clay soil type and suffered extreme drought stress with natural BYV and BMYV infection; MS/- was on a marshland soil type and suffered no moisture stress and S/D was on a light sand soil type and was affected by drought. The sites were visited frequently during the experiment and on many occasions all of the sites except MS/- showed the effects of drought characterised by severely flagging leaves.
In these experiments, plants were grown at a density of 70 000-80 000 per hectare. The trial design was randomised blocks of plots 6 m long by three rows set at 50 cm spacings. The trial margins were guarded but within the trial there were no guards. All other treatments were in accordance with the standard agronomic practices of the area, i.e. plant nutrition (Pocock, Milford & Armstrong, 1988), plant protection (Dewar, 1988) and weed control (Proctor, 1988). To guard against weak emergence the trial was not precision drilled, but was ‘singled’ by hand hoeing to final stands at the 2-4 leaf stage. Drilling was during the normal commercial period of late-March to late-April and the plants were harvested between mid-October and mid-November using modified commercial machinery. The following characters were assessed: sugar content (SC), root yield (RY), potassium content (K) , sodium content (Na) and amino-nitrogen content (AMN) which were measured according to Jolliffe (1990).
Results
BCVl and BCV2 infection in seedlings of different parent sugar beet lines Plants of both the male sterile F, hybrid monogerrn parent and the tetraploid multigerm
pollinator parent were screened for infection with BCVl and BCV2. Of 100 F, hybrid line seedlings analysed, 19% were infected with BCVl alone, 26% with BCV2 alone, 23% contained both viruses and 32% contained neither. Of 105 pollinator line seedlings analysed, none was infected with BCVl alone, 64% contained BCV2 alone, 1 % had both viruses and 35% seemed to be virus-free.
The healthy seedlings from both the F, hybrid line and the pollinator line were re- examined using Northern blot analysis with both positive and negative controls (data not shown). Leaf extracts from the F, hybrid line healthy seedlings did not hybridise with either the BCVl or BCV2 probe, confirming that these seedlings were free of both BCVl and BCV2. However, some of the seedlings of the pollinator line shown to be healthy in the first screen contained BCV2 when re-examined. Therefore, the remaining healthy seedlings of this pollinator line were screened for a third time. However, few of these seedlings were
BCV infection of sugar beet crops 455
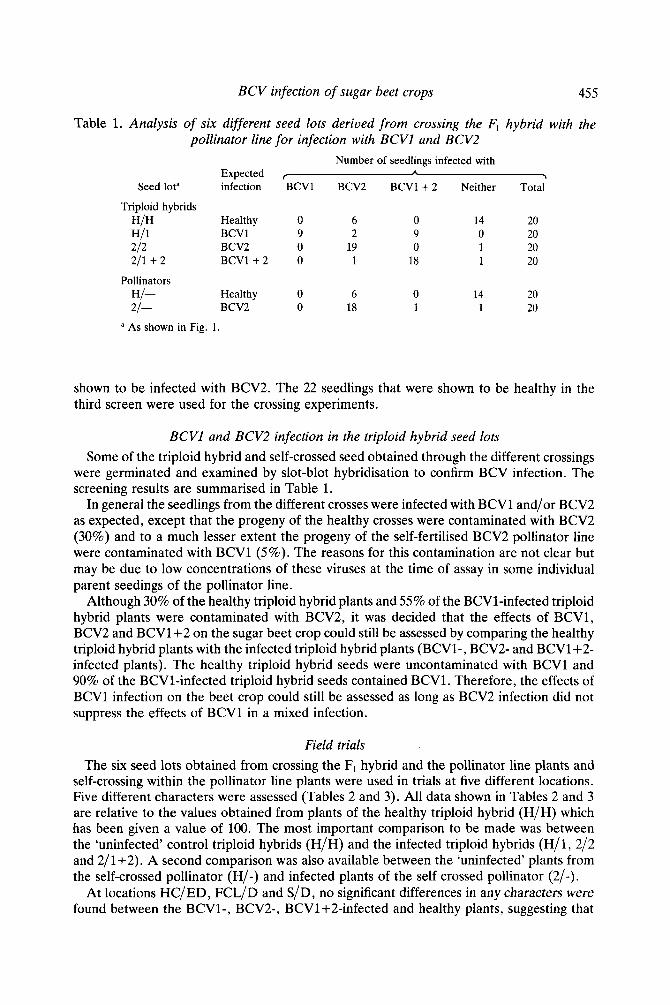
Table 1. Analysis of six different seed lots derived from crossing the Fl hybrid with the pollinator line for infection with BCVl and BCV2
Number of seedlings infected with Expected L \
Seed lota infection BCVl BCV2 BCVl + 2 Neither Total
Triploid hybrids HIH Healthy 0 6 0 14 20 HI1 BCVl 9 2 9 0 20 212 BCV2 0 19 0 1 20 211 + 2 BCV1+2 0 1 18 1 20
HI- Healthy 0 6 0 14 20 21- BCV2 0 18 1 1 20
Pollinators
a As shown in Fig. 1.
shown to be infected with BCV2. The 22 seedlings that were shown to be healthy in the third screen were used for the crossing experiments.
BCVl and BCV2 infection in the triploid hybrid seed lots Some of the triploid hybrid and self-crossed seed obtained through the different crossings
were germinated and examined by slot-blot hybridisation to confirm BCV infection. The screening results are summarised in Table 1.
In general the seedlings from the different crosses were infected with BCVl and/or BCV2 as expected, except that the progeny of the healthy crosses were contaminated with BCV2 (30%) and to a much lesser extent the progeny of the self-fertilised BCV2 pollinator line were contaminated with BCVl (5%) . The reasons for this contamination are not clear but may be due to low concentrations of these viruses at the time of assay in some individual parent seedings of the pollinator line.
Although 30% of the healthy triploid hybrid plants and 55% of the BCV1-infected triploid hybrid plants were contaminated with BCV2, it was decided that the effects of BCV1, BCV2 and BCVl+2 on the sugar beet crop could still be assessed by comparing the healthy triploid hybrid plants with the infected triploid hybrid plants (BCV1-, BCV2- and BCV1+2- infected plants). The healthy triploid hybrid seeds were uncontaminated with BCVl and 90% of the BCV1-infected triploid hybrid seeds contained BCVl . Therefore, the effects of BCVl infection on the beet crop could still be assessed as long as BCV2 infection did not suppress the effects of BCVl in a mixed infection.
Field trials The six seed lots obtained from crossing the F1 hybrid and the pollinator line plants and
self-crossing within the pollinator line plants were used in trials at five different locations. Five different characters were assessed (Tables 2 and 3). All data shown in Tables 2 and 3 are relative to the values obtained from plants of the healthy triploid hybrid (H/H) which has been given a value of 100. The most important comparison to be made was between the ‘uninfected’ control triploid hybrids (H/H) and the infected triploid hybrids (H/l, 2/2 and 2/1+2). A second comparison was also available between the ‘uninfected’ plants from the self-crossed pollinator (H/-) and infected plants of the self crossed pollinator (2/-).
At locations HC/ED, FCL/D and S/D, no significant differences in any characters were found between the BCV1-, BCV2-, BCV1+2-infected and healthy plants, suggesting that
456 W S XIE, J F ANTONIW, R F WHITE AND T H JOLLIFFE
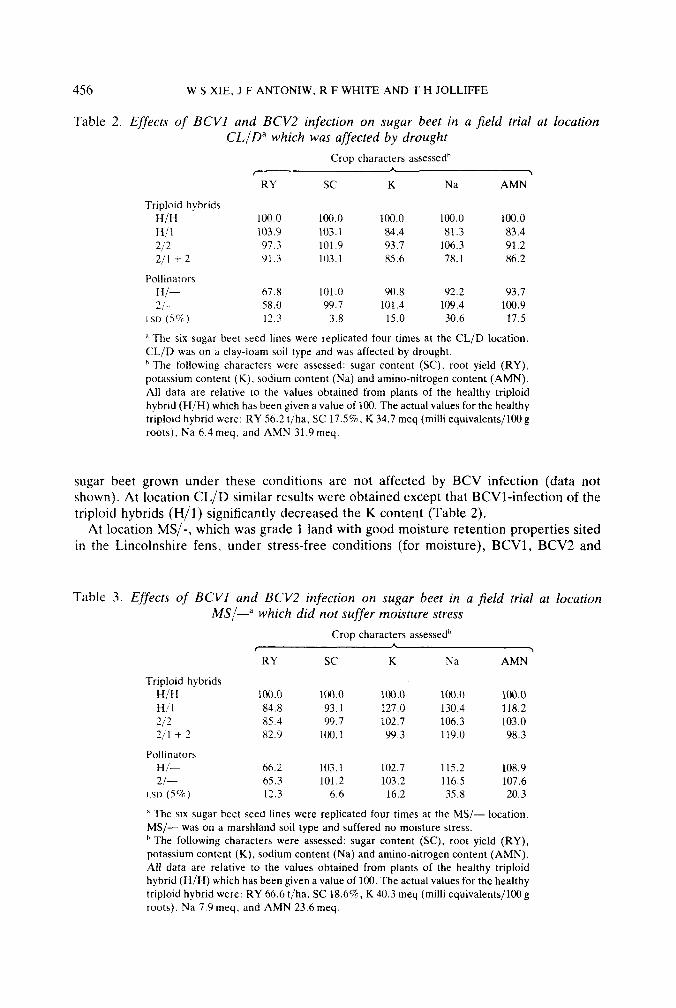
Table 2. Effects of BCVl and BCV2 infection on sugar beet in a field trial at location CL,/D” which was affected by drought
Crop characters assessedb
RY sc K Na AMN
H/H 100.0 100.0 100.0 100.0 100.0 H/1 103.9 103.1 84.4 81.3 83.4 212 97.3 101.9 93.7 106.3 91.2 211 + 2 91.3 103.1 85.6 78.1 86.2
r A 3
Triploid hybrids
Pollinators H/- 67.8 101.0 90.8 92.2 93.7 2/- 58.0 99.7 101.4 109.4 100.9
LSD (5%) 12.3 3.8 15.0 30.6 17.5
A The six sugar beet seed lines were replicated four times at the CL/D location. CL/D was on a clay-loam soil type and was affected by drought.
The following characters were assessed: sugar content (SC), root yield (RY), potassium content (K). sodium content (Na) and amino-nitrogen content (AMN). All data are relative to the values obtained from plants of the healthy triploid hybrid (H/H) which has been given a value of 100. The actual values for the healthy triploid hybrid were: RY 56.2 t/ha, SC 17.5%. K 34.7 meq (milli equivalents/100 g roots), Na 6.4 meq, and AMN 31.9 meq.
sugar beet grown under these conditions are not affected by BCV infection (data not shown). At location CL/D similar results were obtained except that BCV1-infection of the triploid hybrids (H/I) significantly decreased the K content (Table 2).
At location MS/-, which was grade 3 land with good moisture retention properties sited in the Lincolnshire fens, under stress-free conditions (for moisture), BCV1, BCV2 and
Table 3 . Effects of BCVl and BCV2 infection on sugar beet in a field trial at location MS/-d which did not suffer moisture stress
Crop characters assessedb
RY sc K Na AMN
Triploid hybrids H/H 100.0 100.0 100.0 100.0 100.0 HI 1 84.8 93.1 127.0 130.4 118.2 212 85.4 99.7 102.7 106.3 103.0 211 + 2 82.9 100. 1 99.3 119.0 98.3
Pollinators HI- 66.2 103.1 102.7 115.2 108.9 21- 65.3 101.2 103.2 116.5 107.6
LSD (5%) 12.3 6.6 16.2 35.8 20.3
a The six sugar beet seed lines were replicated four times at the MS/- location. MS/- was on a marshland soil type and suffered no moisture stress.
The following characters were assessed: sugar content (SC), root yield (RY), potassium content ( K ) , sodium content (Na) and amino-nitrogen content (AMN). All data are relative to the values obtained from plants of the healthy triploid hybrid (H/H) which has been given a value of 100. The actual values for the healthy triploid hybrid were: RY 66.6 t/ha, SC 18.696, K 40.3 meq (milli equivalents/100 g roots). Na 7.9 meq, and AMN 23.6 meq.
BCV infection of sugar beet crops 457
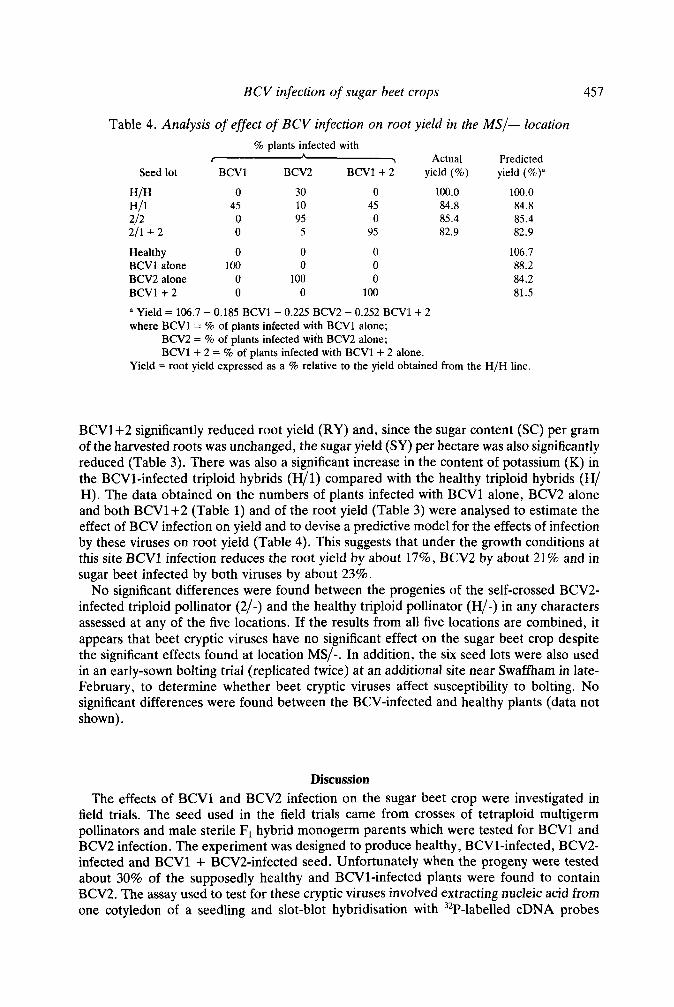
Table 4. Analysis of effect of BCV infection on root yield in the MS/- location % plants infected with
r h > Actual Predicted Seed lot BCVl BCV2 BCVl + 2 yield (%) yield (%)”
H/H 0 30 0 100.0 100.0
212 HI1 45 10 45 84.8 84.8
0 95 0 85.4 85.4 211 + 2 0 5 95 82.9 82.9
Healthy 0 0 0 BCVl alone 100 0 0 BCV2 alone 0 100 0 BCVl + 2 0 0 100
106.7 88.2 84.2 81.5
a Yield = 106.7 - 0.185 BCVl - 0.225 BCV2 - 0.252 BCVl + 2 where BCVl = % of plants infected with BCVl alone;
BCV2 = % of plants infected with BCV2 alone; BCVl + 2 = % of plants infected with BCVl + 2 alone.
Yield = root yield expressed as a % relative to the yield obtained from the H/H line.
BCV1+2 significantly reduced root yield (RY) and, since the sugar content (SC) per gram of the harvested roots was unchanged, the sugar yield (SY) per hectare was also significantly reduced (Table 3). There was also a significant increase in the content of potassium (K) in the BCV1-infected triploid hybrids (H/1) compared with the healthy triploid hybrids (H/ H). The data obtained on the numbers of plants infected with BCVl alone, BCV2 alone and both BCVl+2 (Table 1) and of the root yield (Table 3) were analysed to estimate the effect of BCV infection on yield and to devise a predictive model for the effects of infection by these viruses on root yield (Table 4). This suggests that under the growth conditions at this site BCVl infection reduces the root yield by about 17%, BCV2 by about 21% and in sugar beet infected by both viruses by about 23%.
No significant differences were found between the progenies of the self-crossed BCV2- infected triploid pollinator (2/-) and the healthy triploid pollinator (H/-) in any characters assessed at any of the five locations. If the results from all five locations are combined, it appears that beet cryptic viruses have no significant effect on the sugar beet crop despite the significant effects found at location MS/-. In addition, the six seed lots were also used in an early-sown bolting trial (replicated twice) at an additional site near Swaffham in late- February, to determine whether beet cryptic viruses affect susceptibility to bolting. No significant differences were found between the BCV-infected and healthy plants (data not shown).
Discussion The effects of BCVl and BCV2 infection on the sugar beet crop were investigated in
field trials. The seed used in the field trials came from crosses of tetraploid multigerm pollinators and male sterile F1 hybrid monogerm parents which were tested for BCVl and BCV2 infection. The experiment was designed to produce healthy, BCV1-infected, BCV2- infected and BCVl + BCVZinfected seed. Unfortunately when the progeny were tested about 30% of the supposedly healthy and BCV1-infected plants were found to contain BCV2. The assay used to test for these cryptic viruses involved extracting nucleic acid from one cotyledon of a seedling and slot-blot hybridisation with 32P-labelled cDNA probes

458 W S XIE, J F ANTONIW, R F WHITE AND T H JOLLIFFE
specific for BCVl and BCV2. This assay is very sensitive and the parental plants were tested twice, so it was surprising to find this level of contamination. A possible explanation for this is that some infected seedlings may contain very low levels of cryptic virus, below the level of detection by cDNA hybridisation. It is also possible that the concentration of virus may vary in different parts of an infected plant. Whatever the reason it is clear that great care has to be taken in testing and selecting the breeding stocks. Although the contamination with BCV2 made the results more difficult to interpret and less sensitive, it was still possible to make useful comparisons of the field data.
None of the pollinators showed any effect due to BCV2 infection at any of the sites, suggesting that under these growth conditions plants with the genetic background of these tetraploid multigerm pollinators are not adversely affected by BCV2. Also at four of the five sites there was no apparent effect of BCV infection on root yield in the higher yielding triploid hybrids. However all four of these sites suffered from drought. But at the fifth site, located on marshland type soil, where the beet did not suffer any moisture stress, cryptic virus infection seemed to have a significant effect on root yield and sugar yield. The predictive model shown in Table 4 suggests that at this site, under these growth conditions, infection with either BCVl or BCV2 is associated with a reduction of about 20% in root yield and sugar yield. When beet cryptic viruses had an effect in these experiments, it seemed to be on crops not suffering from moisture stress so it is possible that the full effects of cryptic virus infection on the crop are only seen when the plants are grown under stress- free conditions. As growers strive to maximise the yield of their crop by growing it under the best possible conditions, a limiting factor could be the degree of infection of the crop with cryptic viruses.
Although cryptic viruses do not normally induce symptoms, they might synergistically interact with another pathogenic virus in a mixed infection. Wild beet sources (e.g. Beta maritirna) are being increasingly used to breed new beet resistant to plant viruses (e.g. yellowing viruses and rhizomania) (Jolliffe, 1988; Kimber, 1992). As already reported, the wild beet, B. maritima contains BCVl (Xie et a f . , 1989), and BCV2 which was found in B. muriiirna from Denmark (Xie, 1992). Hammer, Stanarius & Kuhne (1990) also reported that several wild beet and primitive cultivars were infected by either BCVl, BCV2 or both BCVl and BCV2. Thus the presence of beet cryptic viruses in these beet should be considered carefully in breeding programmes in case beet cryptic viruses cause any potential economic problems.
Acknowledgements We thank Professors K W Buck and R T Plumb for helpful discussions, and the British
Council and Hilleshog Sugar Beet Breeding Station for financial support. The field trials and their analysis were done at, and in collaboration with, the Hilleshog Sugar Beet Breeding Station in Norwich, UK.
References Accotto G P, Boccardo G . 1986. The coat proteins and nucleic acids of two beet cryptic viruses.
Accotto G P, Brisco M J, Hull R. 1987. In uitro translation of the double-stranded RNA genome from Journal of General Virology 61:363-366.
beet cryptic virus 1 . Journal of General Virology 68:1417-1422.

BCV infection of sugar beet crops 459
Antoniw J F, White R F, Xie W S. 1990. Cryptic viruses of beet and other plants. In Recognition and Response in Plant-Virus interaction, pp. 273-286. Ed. R S S Fraser. NATO AS1 Series, Vol. H41. Berlin, Heidelberg: Springer-Verlag.
Antoniw J F, Linthorst H J M, White R F, Bol J F. 1986. Molecular cloning of the double-stranded RNA of beet cryptic viruses. Journal of General Virology 67:2047-2051.
Boecardo G, Lisa V, Luisoni E, Milne R G. 1987. Cryptic plant viruses. Advances in Virus Research
Dewar A. 1988. Rational pesticide use in sugar beet. British Sugar Beet Review 56(3):&8. Hammer K, Stanarius A, Kuhne T. 1990. Differential occurrence of beet cryptic viruses - a new tool
for germplasm characterization and evolutionary studies in beets? Euphytica 45:23-27. Jolliffe T H. 1988. Breeding for tolerance/resistance to virus diseases in sugar beet. Aspects of Applied
Biology 17, Environmental Aspects of Applied Biology, pp. 115-123. J o l l i e T H. 1990. Genetical studies in relation to breeding objectives in sugar beet. Ph.D. Thesis,
University of East Anglia. Kassanis B, Russell G E, White R F. 1978. Seed and pollen transmission of beet cryptic virus in sugar
beet plants. Phytopathologische Zeitschrift 91:7&79. Kassanis B, White R F, Woods R D. 1977. Beet cryptic virus. Phytopathologische Zeitschrift 90:35&
360. Kimber D. 1992. Breeding for disease resistance in sugar beet. British Sugar Beet Review 60(1):3@
31. Kuhne T, Stanarius A, Kleinhempel H. 1987. Beet cryptic virus - purification and serological evidence
in four varieties of Beta vulgaris L. Archiv Phytopathologie Pjlanzenschutz, Berlin 23:95-104. Milne R G. 1991. Cryptic virus group. In Classification and nomenclature of viruses, pp. 212-215. Eds
R I B Francki, C M Fauquet, D L Knudson and F Brown. Fifth report of the International Committee on Taxonomy of Viruses. New York: Springer-Verlag Wien. Archives of Virology, Supplementum 2.
Moir D L. 1983~. Effects of beet cryptic virus on sugar beet. In Diseases and pests, Rothamsted Station Report for 1982, Part I , pp. 80-81.
Moir D L. 1983b. Some effects of beet cryptic virus in sugar beet. Ph.D. Thesis, University of Newcastle- Upon-Tyne.
Natsuaki T. 1985. Radish yellow edge virus. Association of Applied Biologists Descriptions of Plant Viruses No. 298, 4 pp.
Natsuaki T, Yamashita S, Doi Y, Yora K. 1979. Radish yellow edge virus. Annals of the Phy- topathological Society of Japan 45313-320.
Natsuaki T, Yamashita S, Doi Y, Okuda S, Teranaka M. 1983. Radish yellow edge virus, a seed-borne virus with double-stranded RNA, of a possible new group. Annals of the Phytopathological Society of Japan 49:59>599.
Natsuaki T, Natsuaki K T, Okuda S, Teranaka M, Mihe R G, Boccardo G, Luisoni E. 1986. Relationship between the cryptic and temperate viruses of alfalfa, beet and white clover. Znter- virology 2559-75.
Pocock T, Milford G, Armstrong M. 1988. The nitrogen nutrition of sugar beet. British Sugar Beet Review 56(3):41-44.
Proctor G. 1988. Weed beet control - British Sugar’s bolter cutter project. British Sugar Beet Review
Pullen M E. 1968. Virus diseases in root crops - ,ugar beet. In The incidence and spread of viruses.
Pullen ME. 1969. Virus-like particles in sugar beet. In Virus diseases of grasses and cereals. Rothamsted
Xie W S. 1992. Studies on cryptic viruses in sugar beet and ryegrass. Ph.D. Thesis, University of
Xie W S, Antoniw J F, White R F. 1989. Detection of beet cryptic viruses 1 and 2 in a wide range of
32~171-214.
56(1):9-10.
Rothamsted Experimental Station Report for 1967, Part I , pp. 124-125.
Experimental Station Report for 1968, Part I , p. 127.
London.
beet plants using cDNA probes. Plant Pathology 38527-533.
(Received 28 June 1993)





























