Embed Size (px)
Citation preview

Neuroscience Letters, 65 (1986) 3540 35 Elsevier Scientific Publishers Ireland Ltd.
NSL 03832
EFFECTS OF C H R O N I C D E H Y D R A T I O N O N A N G I O T E N S I N II R E C E P T O R BINDING IN THE S U B F O R N I C A L ORGAN, P A R A V E N T R I C U L A R HYPOTHALAMIC N U C L E U S A N D A D R E N A L MEDULLA OF LONG-EVANS RATS
BANG H. HWANG l'*, J.-Y. WU 2 and WALTER B. SEVERS 3
Departments ~[" tAnatomy, 2physiology and 3Pharmacology, The Milton S. Hershey Medical Center, The Pennsylvania State University, College of Medicine, Hershey, PA 17033 (U.S.A.)
(Received September 19th, 1985; Revised version received and accepted December 16th, 1985)
Key words: angiotensin lI water deprivation- thirst- subfornical organ adrenal medulla rat
Angiontensin II (AII) is an important peptide known to regulate blood pressure and body fluid. In the present study we used a potent AII antagonist, ~2~I-(Sart,IleS)-AII (~25I-SI-AII), to study AII receptor bind- ing in Long-Evans rats 5 days after water deprivation. Specific structures evaluated include the subfornical organ (SFO) and adrenal gland. With quantitative autoradiography, we have found that there is an in- crease of ~25I-SI-AII binding in the SFO, whereas there is a decrease in AII binding in the adrenal medulla. These observations suggest that central and peripheral AII target tissues are affected differently by dehy- dration. The increase in SI-AII binding in the SFO may indicate participation of this structure during dehydration, as angiotensin stimulation of SFO causes thirst and vasopressin release.
An intracerebroventricular administration of angiotensin II (AII) can exert differ- ent effects including distinct cardiovascular responses, increased vasopressin secre- tion and elicitation of drinking [1, 2, 24, 26, 29], suggesting that AII interactions with AII receptors within the brain are responsible for the above effects. In fact, AII has been localized in the brain by immunocytochemistry [12]. Endogenous renin, angio- tensin converting enzyme and AII receptor binding sites [4, 6, 14, 20] have also been demonstrated within the brain. It is therefore generally accepted that the brain has its own AII system [8, 19, 24, 26].
It is believed that AII is involved in the regulation of body fluid balance [11, 15, 19, 28] through a complicated system in the brain [16, 18, 19] including the subforni- cal organ (SFO). The principal peptides such as AII and vasopressin (VP) responding to dehydration have also been found in the SFO, paraventricular hypothalamic nu- cleus (PVN) and dorsal medulla oblongata [12, 16, 20, 26]. In addition, peripheral organs such as the adrenal gland are also involved in the physiological response to dehydration [19]. In this report, AII receptor binding in the adrenal gland, SFO, PVN and dorsal medulla oblongata were, therefore, studied by quantitative autora- diography using ~25I-Sar~,Ile~-AII (125I-SI-AII) as a specific ligand.
*Author for correspondence.
0304-3940/86/$ 03.50 © 1986 Elsevier Scientific Publishers Ireland Ltd.

36
A d u l t male L o n g - E v a n s rats weighing 251-303 g were used. Two groups o f ani-
mals were studied: (a) 4 rats were depr ived o f d r ink ing water for 5 days with free
access to food [9]; (b) 4 rats had free access to d r ink ing water and food dur ing the
5-day exper iment and served as controls . A t the end o f the 5-day exper iment , the d ry
feces f rom ei ther exper imenta l o r con t ro l s were collect ively weighed. Af te r experi-
ments , an imals were anes thet ized with p e n t o b a r b i t a l (50 mg/kg , i.p.) and perfused
with 300 ml o f ice-cold 0.1% p a r a f o r m a l d e h y d e in phospha te -buf fe red saline (PBS).
The bra ins were removed , frozen and cut into 20-#m sections with a cryosta t . The
sections were t haw-moun ted on subbed glass slides and s tored at least overnight in
freezer at - 2 0 ° C . The in situ incuba t ion for ~25I-SI-AII b ind ing (Kd is a b o u t 0.1 nM)
was s imilar to the p rocedure used previous ly [14]. In brief, the sect ions were incu-
ba ted with 100 p M 125I-SI-AII (spec.act . , 2376 Ci /mmol ) in 0.5 ml PBS con ta in ing
5 m M N a 2 E D T A for 1 h at r o o m tempera ture . Non-specif ic b ind ing was deter-
mined in the presence o f 1/~M non- labe led S I - A I I in the above ~25I-SI-AII solut ion.
Af te r r insing and drying, the sect ions on glass slides were a ppose d to L K B Ul t rof i lms
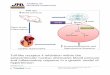
Fig. 1. A: total AII receptor binding in the ADG (black ring) and ADM (m); from a control rat. B: AII receptor binding of the adrenal gland; from a dehydrated rat. m, ADM. C: non-specific binding for the adrenal gland. Similar non-specific binding was obtained in all the brain sections. D: the All binding activ- ities in the medulla oblongata of a control rat. Binding is in the NTS (arrows), DMX (white crosses) and SNV (arrowheads). The area postrema is not at this caudal level of the medulla oblongata. E: the AII receptor binding is seen in the SFO (arrow) and PVN (arrowheads); from a control rat. F: more All receptor binding in the SFO (arrow) can be appreciated in this dehydrated rat as compared to E. Bar = 1 mm (for all figures).

37
in the X-ray cassette for exposure for 5 days at room temperature. The 9 ~25I-SI-AII standards were prepared, cut and collected on the glass slides as we did previously [14] according to Rainbow et al. [21], and then apposed to the LKB films together with the above-mentioned tissue sections. A microcomputer-assisted densitometer was used to determine msI-SI-AII binding activities in fmol/mg protein [3, 14, 21].
Physiological status: The 5-day water deprivation affected the body weight drama- tically. Dehydrated animals lost 25% of their body weight (from 262.4_+2.2 to 196.2_+2.2 g), whereas the control animals gained 12% in body weight (from 300.0_+ 10.0 to 336.8+8.4 g). The dehydrated animals ate and urinated much less than control animals, based upon daily observations. Reduced feeding was indirectly reflected by fecal weight. Dried fecal weight of dehydrated animals was only 16% of controls.
~251-SI-AH receptor binding: in the adrenal gland, no significant difference of SI- AII binding was found in the adrenal glomerulosa (ADG) between dehydrated and control animals. The adrenal medulla (ADM) of dehydrated animals (Fig. IA-C) had a significant reduction (P < 0.01) of SI-AII binding 5 days after water deprivation (Table I). In 5 brain regions (Fig. 1D-F) evaluated, the SFO (Fig. IE, F: Table I) was the only one structure to have a significant increase (P<0.01) in ~25I-SI-AII receptor binding (0.90_+ 0.07 fmol/mg protein) in water-deprived rats as compared to control animals (0.60 + 0.02 fmol/mg protein). There were no significant changes of SI-AII binding in the PVN, nucleus tractus solitarii (NTS), the dorsal motor nu- cleus of the vagus (DMX) and spinal nucleus of the trigeminal (SNV) after chronic dehydration.
This study agrees with the literature [22, 29] that shows water deprivation is asso- ciated with loss of body weight, reduction in food consumption and antidiuresis. The major finding of this study is that chronic dehydration for 5 days results in a 50% increase of SI-AII receptor binding in the SFO of the forebrain. Since the state of
TABLE I
THE CHRONIC EFFECT OF 5-DAY WATER DEPRIVATION ON ANGIOTENSIN I1 RECEPTOR
BINDING IN THE ADRENAL GLAND AND DIFFERENT BRAIN REGIONS OF LONG-EVANS
RATS a
*Significantly different at 0.01 level as evaluated by two-tailed Student's t-test.
Structure Control (n = 4) Dehydrated (n = 4)
SFO 0.60_+0.02 0.90_+0.07*
PVN 0.53 _+ 0.05 0.50 _+ 0.04
NTS 0.73 _+ 0.06 0.73 __+ 0.10
DMX 0.70_+0.04 0.70+_0.07
SNV 0.20 +_ 0.07 0.28 _+ 0.03
ADG 3.75-+0.57 3.38_+0.17
ADM 10.18 -+ 0.62 7.27 -+ 0.37"
aData for each structure were averaged from at least 16 determinations from 4 animals, and given as mean-
+ S.E.M. in fmol/mg protein.

38
chronic dehydration produces countless changes, it is difficult to assess whether the increase of SI-AII receptor binding in the SFO is directly due to dehydration or is a secondary effect. There are several possible mechanisms which may lead to an in- crease of SI-AII receptors. First of all, it may be due to the induction of AI! receptors by dehydration. Secondly, the rate of synthesis of AII receptors may not be altered in the dehydration condition, but rather the turnover/degradation of receptors is reduced, resulting in a net increase of SI-AI! binding in the SFO. Thirdly, it may be due to an enhanced degradation of fodrin which usually masks the receptor bind- ing sites, resulting in an increase of AII receptor density. Such unmasking or uncap- ping has been demonstrated at glutamate receptor binding sites [25, 27]. Further- more, changes of SI-AII receptor binding may also be possible due to differences in the body weight, but this possibility is rather remote because all the animals used at the onset of this study were mature, adult rats. However, our result showing increased AII receptors in the SFO is compatible with the interpretation that dehy- dration activates the AII system [16, 19, 26, 28] in the SFO and evokes the thirst drive and VP release [10, 28, 29]. Other approaches have also shown that the SFO is metabolically activated during dehydration [9]. In general, we support the hypo- thesis that AII elicits thirst by stimulating AII receptors in the SFO. Furthermore, an increase of AII receptor binding capacity could sensitize the SFO to available All, and result in thirst.
Our results also showed that dehydration produces no significant changes in AII receptors in the PVN, NTS, DMX and SNV, areas associated with the cardiovascu- lar control. Other studies have shown that dehydration has little effect on blood pres- sure [22, 29]. Conceivably, AII receptors in these nuclei may therefore not be closely involved in the cardiovascular modulation after 5-day water deprivation. However, alterations of AII receptors in these nuclei during dehydration cannot be ruled out. In addition, it is noteworthy that the PVN, NTS, DMX and SNV possess a blood- brain barrier which is absent in the SFO. Thus, AII receptors in the PVN, NTS, DMX and SNV may not respond in the same manner as the SFO. However, our results showing no changes of AII receptors do not mean that AII receptors are unre- lated to the development of hypertension. For example A II receptors in the VNS [14] and AII in the hypothalamus [8] have been shown to increase significantly in sponta- neously hypertensive rats, suggesting a possible role of the AI! in the hypothalamus and the SNV for genetic hypertension. Collectively, it seems clear that AII systems responsible for the pressor response and thirst are located at least partially in differ- ent neural circuits [16-19].
The adrenal gland possesses the highest AII binding per unit area of structures we have evaluated. In the ADG, there is no difference in AII receptor binding between the dehydrated and control animals, whereas a significant reduction of AII receptor binding in the ADM was detected in the dehydrated animals. The reduction of AII receptor binding is likely to reflect down-regulation of AII receptors which might re- sult in a general decrease of sympathetic function, since AII influences the sympathet- ic nervous system [7, 13]. This may in part explain why dehydration induces increases of the plasma AII and VP [28, 29], but does not affect blood pressure [5, 22, 23, 29].

39
T h i s w o r k w a s s u p p o r t e d in p a r t by f u n d s f r o m A H A - 8 5 1 3 3 4 a n d A H A - P A l 9 8 5 .
J . - Y . W . is s u p p o r t e d b y N S 2 0 9 7 8 a n d N S 2 0 9 2 2 . W e t h a n k M r . A n d r e w G a y d o s , Jr .
fo r t he p h o t o g r a p h i c p r o c e s s i n g a n d Ms . D o r i s L i n e w e a v e r f o r s e c r e t a r i a l a s s i s t ance .
Spec ia l t h a n k s go to D r , J .J . H a r d i n g o f W a s h i n g t o n S t a t e U n i v e r s i t y fo r s u p p l y i n g
t :5I-S1-AII , a n d to D r . B.B. W o l f e fo r a l l o w i n g the a u t h o r to use t he c o m p u t e r -
a s s i s t ed d e n s i t o m e t e r s y s t e m in the D e p a r t m e n t o f P h a r m a c o l o g y , U n i v e r s i t y o f
P e n n s y l v a n i a .
I Andrews~ C.E., Jr. and Brenner, B.M., Relative contributions of arginine vasopressin and angiotensin II to maintenance of systemic arterial pressure in the anesthetized water-deprived rat, Circ. Res., 48 ( 1981 ) 254-258.
2 Bealer, S.L,, Crofton, J.T, and Share, L., Hypothalamic knife cuts after fluid regulation, vasopressin secretion, and natriuresis during water deprivation, Neuroendocrinology, 26 (1983) 364-370.
3 Berck, D.J. and Rainbow, T.C., Microcomputer-assisted densitometer for quantiative receptor autora- diography, J. Neurosci. Meth., 13 (1985) 171 181.
4 Brownfield, M.S., Reid, I.A., Ganten, D. and Ganong, W.F., Differential distribution of immunoreac- live angiotensin and angiotensin-converting enzyme in rat brain, Neuroscience, 7 (1982) 1759 1769.
5 Burnier, M., Bildaz, J., Brunner, D.B. and Brunner, H.R., Blood pressure maintenance in awake dehydrated rats: renin, vasopressin, and sympathetic activity, Am. J. Physiol., 245 (1983) 11203-11209.
6 Chevillard, C., Niwa, M. and Saavedra, J., Angiotensin-converting enzyme in discrete forebrain areas of spontaneously hypertensive rats, Brain Res., 309 (1984) 389-392.
7 Corwin, E.J., Seaton, J.F., Hamaji, M. and Harrison, T.S., Central role for angiotensin in control o1" adrenal catecholamine secretion, Am. J. Physiol., 248 (1985) R363 R370.
8 Ganten, D., Hermann, K., Bayer, C., Unger, Th. and Lang, R.E., Angiotensin synthesis in the brain and increased turnover in hypertensive rats, Science, 221 (1983) 869 871.
9 Gross, P.M., Kadekaro, M., Sokoloff, L., Holcomb, H.H. and Saavedra, J.M., Alterations of local cerebral glucose utilization during chronic dehydration in rats, Brain Res., 330 (1985) 329 336.
10 Haarmann, V.H., Seizinger, B.R. and Herz, A., Levels of dynorphin-(1-13) immunoreactivity in rat neurointermediate pituitaries are concomitantly altered with those of leucine enkephalin and vasopres- sin in response to various endocrine manipulations, Neuroendocrinology, 33 (1981) 333- 339.
1 I Halpcrin, E.S., Summy-Long, J,Y., Keil, L.C. and Severs, W.B., Aspects of salt/water balance after cerebroventricular infusion of angiotensin II, Brain Res., 205 (1981) 219 -221.
12 Healy, D.P., Rettig, R. and Printz, M.P., Evidence for an endogenous angiotensin II system in the rat nucleus tractus solitarius, Hypertension, A6 (1984) 1743-1747.
13 Hughes, J. and Roth, R.H., Enhanced release of transmitter during sympathetic nerve stimulation in the presence of angiotensin, Br. J. Pharmacol., 37 (1969) 516P 517P.
14 llwang, B.H., Harding, J.W., Liu, D.K., Hibbard, L.S., Wieczorek and Wu, J.-Y., Quantitative auto- radiography of ~:sI-(Sar ~, lleS)-angiotensin II binding sites in the brain of spontaneously hypertensive rats. Brain Res. Bull., submitted.
15 Keil, L.C., Summy-Long, J. and Severs, W.B., Release of vasopressin by angiotensin 11. Endocri- nology, 96 (1975) 1063-1065.
16 Lind, R.W. and Johnson, A.K., Subfornical organ-median preoptic connections and drinking and pressor responses to angiotensin It, J. Neurosci., 2 (1982) 1043- 1051.
17 Mcndelsohn. F.A.O., Aguilera, G. and Saavedra, J.M., Characteristics and regulation of angiotensin II receptors in pituitary, circumventricular organs and kidney, Clin. Exp. Hypert., A5 (1983) 1081 - 1097.
18 Miselis, R.R., The efferent projections of the subfornical organ of the rat: a circumventricular organ within a neural network subserving water balance, Brain Res., 230 ( 1981 ) 1-23.
19 Phillips, M.I., Hoffman, W.E. and Bealer, S.L., Dehydration and fluid balance: central effects of angio- tensin, Fed. Proc. Fed. Am. Soc. Exp. Biol., 41 (1982) 2520-2527.
20 Printz, M.P. and Lewicki, J.A., Renin substrate in the CNS: potential significance to central regulatory

40
mechanisms. In J.P. Buckley and C.M. Ferrario (Eds.), Central Actions of Angiotensin and Related Hormones, Pergamon, New York, 1977, pp. 59-73.
21 Rainbow, T,C., Biegon, A. and Berck, D.J., Quantitative receptor autoradiography within tritium-la- beled ligands: comparison of biochemical and densitometric measurements, J. Neurosci, Meth., I 1 (1984)231 241.
22 Rascher, W., Meme, H. and Gross, F., Hemodynamic effects of arginine vasopressin in conscious water-deprived rats, Am. J. Physiol., 249 (1985) H29-H33.
23 Schwartz, J. and Reid, I.A.. Role of vasopressin in blood pressure regulation in conscious water- deprived dogs. Am. J. Physiol., 244 (1983) R74--R 77.
24 Severs, W.B. and Daniels-Severs, A.E., Effects of angiotensin on the central nervous system, Pharma- col. Rev.. 25 (1973) 415M49.
25 Siman, R., Baudry, M. and Lynch, G., Regulation of glutamate receptor binding by the cytoskeletal protein fodrin, Nature (London), 313 (1985) 225-228.
26 Simpson, J.B., Mangiapane, M.L. and Dellmann, HD., Central receptor sites for angiotensin-induced drinking: a critical review, Fed. Proc. Fed. Am. Soc. Exp. Biol., 37 (1978) 2676-2682.
27 Smith, R., Baudry, M. and Lynch, R., Brain fodrin: substrate for calpain I, an endogenous calcium- activated protease. Proc. Natl. Acad. Sci. USA, 81 (1984) 3572-3576.
28 Summy-Long, J.Y., Keil, L.C., Hernandez, L., Emmert, S., Chee, O. and Severs, W.B., Effects of dehydration and renin on vasopressin concentration in the subfornical organ area, Brain Res., 300 (1984) 219-229.
29 Woods, R.L. and Johnston, C.I., Contribution of vasopressin to the maintenance of blood pressure during dehydration. Am. J. Physiol., 245 (1983) F615-F621.