Embed Size (px)
Citation preview

J Physiol 560.1 (2004) pp 207–217 207
Effects of cortical stimulation on auditory-responsivethalamic neurones in anaesthetized guinea pigs
Ying Xiong1,3, Yan-Qin Yu1, Ying-Shing Chan2 and Jufang He1
1Department of Rehabilitation Sciences, The Hong Kong Polytechnic University, Hung Hom, Kowloon, Hong Kong, China2Department of Physiology, Faculty of Medicine, The University of Hong Kong, Hong Kong, China3Laboratory of Neural Plasticity, Institute of Neuroscience, Chinese Academy of Sciences, Shanghai, China
In the present study, we investigated neuronal responses to acoustic stimuli and corticalstimulation in the medial geniculate body (MGB) through in vivo intracellular recordingsin anaesthetized guinea pigs. Of the 54 neurones examined with acoustic stimuli, 36 showedexcitatory postsynaptic potential (EPSP) responses and 19 showed inhibitory postsynapticpotential (IPSP) responses to acoustic stimuli. Of the 36 EPSP neurones examined withcorticofugal modulation, 29 received corticofugal depolarization, 3 corticofugal inhibition,and 4 showed no effect. Of the 19 IPSP neurones, 17 received corticofugal inhibition and2 were not affected. The mean amplitude of the EPSPs evoked by acoustic stimuli was similarto that evoked by the electrical cortical stimulation (9.19 ± 5.55 mV versus 9.22 ± 5.16 mV).There was a significant correlation between the parameters of the EPSPs evoked by anacoustic stimulus and those evoked by cortical stimulation. The mean amplitude of theIPSP evoked by electrical cortical stimulation was significantly greater than that evoked byacoustic stimuli (11.6 ± 3.8 mV versus 9.1 ± 3.7 ms, P < 0.05). Seven auditory EPSP and 7 IPSPneurones were examined with corticofugal modulation and labelled with Neurobiotin.Of the 7 EPSP neurones, 5 showed excitatory responses to cortical stimulation and 2demonstrated no effects. Four of the 5 neurones that received corticofugal depolarizationwere located in the lemniscal MGB and 1 in the non-lemniscal MGB; of the remaining 2, 1was located in the lemniscal and the other in the non-lemniscal MGB. Of the 7 IPSP neurones,1 received an excitatory corticofugal input followed by an inhibitory input and 4 received onlyan inhibitory corticofugal input, while the remainder demonstrated no corticofugal effects.All 7 neurones were located in the non-lemniscal MGB. The result that both ascending anddescending inputs caused similarly shaped EPSPs reflects a neuronal endogenous characteristicirrespective of the physical locations of the synapses. The IPSP responses to both acousticstimuli and electrical cortical stimulation are likely to be caused by feedback from the thalamicreticular nucleus.
(Resubmitted 4 May 2004; accepted after revision 21 July 2004; first published online 22 July 2004)Corresponding author J. He: Department of Rehabilitation Sciences, The Hong Kong Polytechnic University, HungHom, Kowloon, Hong Kong, China. Email: [email protected]
In the accompanying paper (Yu et al. 2004b), weinvestigated the auditory responses of medial geniculateneurones through in vivo intracellular recordings inanaesthetized guinea pigs. We categorized the neuronesto the following types based on the response patternsto acoustic stimuli: the excitatory postsynaptic potential(EPSP) type, the pure inhibitory postsynaptic potential(IPSP) type, and to an IPSP preceded by an EPSP(EPSP–IPSP) type. Some of the neurones showing the
Y. Xiong and Y.-Q. Yu contributed equally to this work.
above response patterns were anatomically labelled andlocalized. The EPSP neurones are located in both thelemniscal and non-lemniscal MGB, and the IPSP neuronesare located only in the non-lemniscal MGB. By injectingan electrical current, we were able to manipulate themembrane potential and examine the dependence ofthe EPSP and IPSP on the resting membrane potential.The responses of low-threshold calcium spikes (LTS)/LTSbursts to acoustic stimuli were elicited from both EPSP andIPSP neurones when their membrane potential was hyper-polarized to below −74 mV. Given that non-lemniscalneurones are multisensory and involved in adjusting the
C© The Physiological Society 2004 DOI: 10.1113/jphysiol.2004.067686

208 Y. Xiong and others J Physiol 560.1
global activity of the auditory cortex, we proposed thatselective inhibition in the non-lemniscal MGB enables theascending pathway to maintain the auditory cortex in acondition for receiving subsequent auditory information(Yu et al. 2004b).
The reciprocal projection from the cortex to the MGBis much stronger than the ascending thalamocorticalprojection (Andersen et al. 1980; Winer & Larue, 1987).It has been suggested that this corticofugal projectionprovides a gating or gain-control mechanism in thetransmission of information from the periphery tothe cortex through the corticofugal facilitatory and/orinhibitory modulation of the thalamic relay neurones,either directly via stimulated neurones or indirectly viaa polysynaptic path (Watanabe et al. 1966; Ryugo &Weinberger, 1976; Deschenes & Hu, 1990; Villa et al. 1991;He, 1997, 2003a; Suga et al. 1997; Zhou & Jen, 2000; Heet al. 2002; Yu et al. 2004a). It is also well known that thecorticofugal system has a powerful modulating effect onthalamic oscillation (Golshani & Jones, 1999; Blumenfeld& McCormick, 2000; Steriade, 2000, 2001b; He,2003b).
Recently, we observed a mostly inhibitory effect on the‘on’ responses of non-lemniscal MGB neurones followingcortical activation (He, 2003a; Yu et al. 2004a). Weproposed that the strong corticofugal inhibition is createdvia the cotico-reticulo-thalamic pathway (Golshani et al.2001; He, 2003a). In the accompanying paper, we foundthat the IPSP-type neurones were mainly located in thenon-lemniscal MGB. The question that arises here iswhether there is any correlation between the neuronalresponse patterns to acoustic stimuli and the corticofugalmodulation.
In the present intracellular study in anaesthetized guineapigs, we investigated the responses of MGB neurones toacoustic stimuli and cortical stimulation. The circuitrywas analysed with respect to the membrane responses ofMGB neurones to acoustic stimuli, spontaneous spikes,and electrical stimulation of the auditory cortex.
Methods
Animal preparation for the intracellular recordings
Thirty-seven guinea pigs were used for the presentintracellular recording study. A large proportion of theexperimental animals were the same as those usedin the accompanying paper (Yu et al. 2004b). Theprocedures involved in the preparation of the animals,acoustic stimulus, and anatomical confirmation werethe same as those described in the accompanyingpaper. Anaesthesia was initially induced with pento-barbital sodium (Nembutal; Abott; 35 mg kg−1; i.p.) andmaintained by supplemental doses of the same anaesthetic(∼5–10 mg kg−1 h−1) during the surgical preparation andrecording. They were killed by overdose of anaesthetic after
the experiment. All of the procedures were approved by theAnimal Subjects Ethics Sub-Committee of The Hong KongPolytechnic University.
Electrical stimulation
We performed several mapping studies of the auditorycortex prior to the present study and found that thesite of stimulation only made a quantitative differencein the corticofugal modulation of thalamic neurones inthe guinea pig (He et al. 2002; He, 2003a). In the presentstudy, we omitted the mapping procedure to save timefor conducting the intracellular recordings. An electrodearray consisting of three parallel electrodes was implantedinto the auditory cortex (the anterior and dorsocaudalauditory fields; Redies et al. 1989; He et al. 2002). Weused electrical current pulse trains (0.1 ms or 0.2 ms inwidth, 50 Hz or 200 Hz in frequency, and 5–20 pulses) toactivate the auditory cortex (Edeline et al. 1994; He, 1997;He et al. 2002). The number of pulses was examined as aparameter in the present study. Electrical currentsof 50–200 µA were applied to the auditory cortex,ipsilaterally to the recording thalamus, through eithera mono- or bipolar low impedance electrode array.Following a delay of 100 ms after the end of the corticalstimulation, a sound stimulus was delivered to thecontralateral ear of the recording thalamus (He, 1997,2003a,b).
Data acquisition and analysis
A negative membrane potential was recorded when theglass recording pipette penetrated the membrane of acell. After amplification, the membrane potential with anartefact of electrical stimulation, as well as the auditorystimulus, was stored in the computer with the aid ofcommercial software (AxoScope, Axon). In the presentstudy, the amplitude, rise time, decay time, and durationof the EPSPs and the IPSPs were measured. The beginningand end of an EPSP and IPSP were defined at a thresholdof 10% of their amplitude. The rise time was defined asthe period between 10% and 90% of their amplitude,and the decay time was defined as the period between90% and 10% of their amplitude. To standardize theexcitatory and inhibitory effect across the neurones,measurements were made of the neuronal responses whenthe membrane potential was as close to −60 mV aspossible. No manipulations of membrane potentials weremade on the data presented here.
The level of the direct current of the recordingelectrode was frequently checked and set to zero duringthe experiments. The level of the direct current after arecording was used to compensate for the membranepotential of some neurones, especially for those with along recording time.
C© The Physiological Society 2004

J Physiol 560.1 MGB responses to cortical stimulation 209
Figure 1. Electrical cortical stimulation potentiates themembrane potential of three MGB neurones thatshowed EPSPs to acoustic stimuliThe left and right panels show auditory responses withoutand with cortical stimulation, respectively. The neurone inA had a resting membrane potential of −57 mV, and that inB, −60 mV. The neurone in A was examined with 1 electricalpulse and 20 electrical pulses in the auditory cortex. Thestimulation current was 100 µA.
Numerical results are expressed as means ± standarddeviation (s.d.). Student’s paired t test was used toexamine the differences in the shapes of the postsynapticpotentials (EPSP and IPSP) between those evoked byacoustic stimuli and those by electrical stimulation of theauditory cortex. The confidence level was taken as 95%(P < 0.05).
Results
The present results were sampled from 54 neurones in37 animals. Of the 54 neurones examined with acousticstimuli, 36 showed EPSPs to an acoustic stimulus and 19showed IPSPs to an acoustic stimulus as defined in theaccompanying paper (Yu et al. 2004b). Of the 36 auditoryEPSP neurones examined with corticofugal modulation,29 demonstrated corticofugal depolarization, 3 cortico-fugal inhibition, and 4 demonstrated no effects. Of the19 auditory IPSP neurones, 17 demonstrated corticofugalinhibitory effects and 2 demonstrated no effects.
Figure 2. An MGB neurone received corticofugaldepolarization in its membrane potentialA, the MGB neurone responded to an acoustic stimulus witha spike and EPSP. B, the membrane potential wasdepolarized by cortical stimulation. Scale bars apply to bothA and B. C, the neurone showed a frequency preference.Five trials of responses to pure-tone stimuli at 60 dB soundpressure level are shown at each of the followingfrequencies: 1, 4, 6 and 14 kHz.
Corticofugal depolarization of auditoryEPSP neurones
Three neurones in Fig. 1 showed an EPSP and a spikeor spike burst in response to an acoustic stimulus,and were depolarized when the auditory cortex waselectrically stimulated with a pulse or a train of pulses. Thecorticofugal depolarization on the neurone in Fig. 1A was11 mV and that on the neurone in Fig. 1B was 18 mV.
The neurone in Fig. 2 showed an auditory responsesimilar to those in Fig. 1A with an EPSP and aspike. With cortical stimulation, this neurone receiveda depolarization in its membrane potential (Fig. 2A).This neurone showed a low-frequency preference ofbetween 1 and 4 kHz in its response to pure-tone stimuli(Fig. 2C).
Corticofugal inhibition of auditory IPSP neurones
Most auditory IPSP neurones were hyperpolarized whenthe auditory cortex was activated with an electrical pulse
C© The Physiological Society 2004
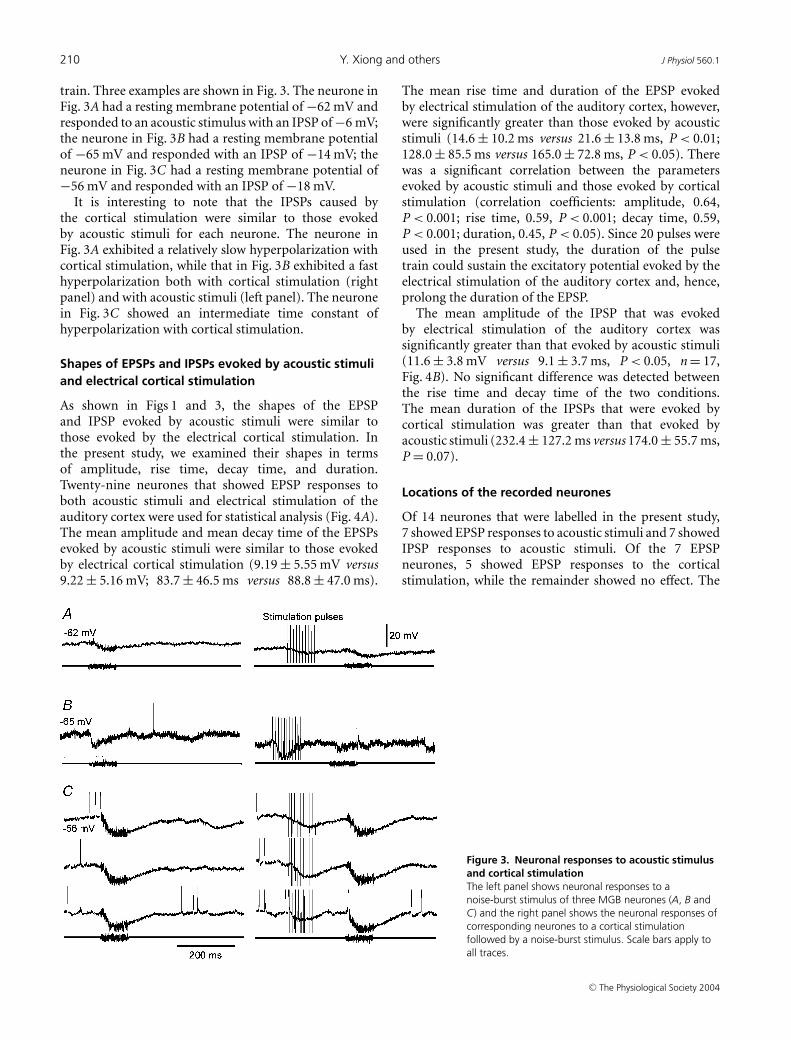
210 Y. Xiong and others J Physiol 560.1
train. Three examples are shown in Fig. 3. The neurone inFig. 3A had a resting membrane potential of −62 mV andresponded to an acoustic stimulus with an IPSP of −6 mV;the neurone in Fig. 3B had a resting membrane potentialof −65 mV and responded with an IPSP of −14 mV; theneurone in Fig. 3C had a resting membrane potential of−56 mV and responded with an IPSP of −18 mV.
It is interesting to note that the IPSPs caused bythe cortical stimulation were similar to those evokedby acoustic stimuli for each neurone. The neurone inFig. 3A exhibited a relatively slow hyperpolarization withcortical stimulation, while that in Fig. 3B exhibited a fasthyperpolarization both with cortical stimulation (rightpanel) and with acoustic stimuli (left panel). The neuronein Fig. 3C showed an intermediate time constant ofhyperpolarization with cortical stimulation.
Shapes of EPSPs and IPSPs evoked by acoustic stimuliand electrical cortical stimulation
As shown in Figs 1 and 3, the shapes of the EPSPand IPSP evoked by acoustic stimuli were similar tothose evoked by the electrical cortical stimulation. Inthe present study, we examined their shapes in termsof amplitude, rise time, decay time, and duration.Twenty-nine neurones that showed EPSP responses toboth acoustic stimuli and electrical stimulation of theauditory cortex were used for statistical analysis (Fig. 4A).The mean amplitude and mean decay time of the EPSPsevoked by acoustic stimuli were similar to those evokedby electrical cortical stimulation (9.19 ± 5.55 mV versus9.22 ± 5.16 mV; 83.7 ± 46.5 ms versus 88.8 ± 47.0 ms).
Figure 3. Neuronal responses to acoustic stimulusand cortical stimulationThe left panel shows neuronal responses to anoise-burst stimulus of three MGB neurones (A, B andC) and the right panel shows the neuronal responses ofcorresponding neurones to a cortical stimulationfollowed by a noise-burst stimulus. Scale bars apply toall traces.
The mean rise time and duration of the EPSP evokedby electrical stimulation of the auditory cortex, however,were significantly greater than those evoked by acousticstimuli (14.6 ± 10.2 ms versus 21.6 ± 13.8 ms, P < 0.01;128.0 ± 85.5 ms versus 165.0 ± 72.8 ms, P < 0.05). Therewas a significant correlation between the parametersevoked by acoustic stimuli and those evoked by corticalstimulation (correlation coefficients: amplitude, 0.64,P < 0.001; rise time, 0.59, P < 0.001; decay time, 0.59,P < 0.001; duration, 0.45, P < 0.05). Since 20 pulses wereused in the present study, the duration of the pulsetrain could sustain the excitatory potential evoked by theelectrical stimulation of the auditory cortex and, hence,prolong the duration of the EPSP.
The mean amplitude of the IPSP that was evokedby electrical stimulation of the auditory cortex wassignificantly greater than that evoked by acoustic stimuli(11.6 ± 3.8 mV versus 9.1 ± 3.7 ms, P < 0.05, n = 17,Fig. 4B). No significant difference was detected betweenthe rise time and decay time of the two conditions.The mean duration of the IPSPs that were evoked bycortical stimulation was greater than that evoked byacoustic stimuli (232.4 ± 127.2 ms versus 174.0 ± 55.7 ms,P = 0.07).
Locations of the recorded neurones
Of 14 neurones that were labelled in the present study,7 showed EPSP responses to acoustic stimuli and 7 showedIPSP responses to acoustic stimuli. Of the 7 EPSPneurones, 5 showed EPSP responses to the corticalstimulation, while the remainder showed no effect. The
C© The Physiological Society 2004

J Physiol 560.1 MGB responses to cortical stimulation 211
5 neurones that demonstrated EPSP responses to corticalstimulation were located in the ventral nucleus (MGv) ofthe MGB. Of the remaining 2 that showed no corticofugaleffects, 1 was located in the MGv and the other in thenon-lemniscal MGB.
Of the 7 IPSP neurones, 1 received an excitatory cortico-fugal input followed by an inhibitory input, 4 receivedonly an inhibitory corticofugal input, and 2 received nocorticofugal effects. All 7 neurones were located in thenon-lemniscal MGB.
Figure 5 shows the anatomical locations of six MGBneurones, which exhibited EPSP and IPSP responsesto acoustic stimuli and to electrical stimulation of theauditory cortex. The neurones in Fig. 5A–C respondedto acoustic stimuli and cortical stimulation with adepolarization of their membrane potential. The neuronesexhibited variety in their shapes. Since the number oflabelled neurones was still small, no further analysis oftheir morphology was conducted in the present study. Twoof the neurones (Fig. 5A and B) were located in the ventralnucleus, the lemniscal MGB, while the third (Fig. 5C) waslocated in the shell nucleus (MGs) of the MGB.
The neurones in Fig. 5D–F responded to electricalcortical stimulation with hyperpolarization of theirmembrane potential. Of these, two neurones (Fig. 5D andF) showed an IPSP response to acoustic stimuli. Theneurone in Fig. 5E showed no response to acoustic stimuli.All of these neurones had a larger soma size than theneurones that showed EPSP responses to acoustic stimuliand cortical stimulation as illustrated in Fig. 5A–C. Theneurone in Fig. 5D was located in the posterior nucleusof the thalamus, where neurones respond to auditorystimuli. The neurone in Fig. 5E was located in the MGs.This neurone did not respond to the noise-burst stimulus.The neurone in Fig. 5F showed an IPSP to the acousticstimulus and to the cortical stimulation. The neuronewas located in the caudomedial nucleus of the MGB. Insummary, all of the neurones in Fig. 5D–F were located inthe non-lemniscal MGB.
IPSP neurones
Figure 6 shows the auditory responses, spontaneousresponses, and responses to cortical stimulation of anauditory thalamic neurone. The neurone responded tothe acoustic stimulus with an IPSP (Fig. 6A). This neuroneshowed an IPSP after each spontaneous spike (Fig. 6B) andresponded to a pulse train of cortical stimulation with thesame shape of IPSP as that found after a spontaneous spike(Fig. 6C).
The neurone in Fig. 7 responded to the acoustic stimuluswith a burst of spikes that was followed by a prolongedIPSP (Fig. 7A). The neurone showed spontaneous firings,which consisted of a single spike and a burst of twospikes with an interval of about 10 ms inbetween (Fig. 7B).
Again, a prolonged IPSP immediately followed the burstof spikes. Interestingly, the neurone showed the sameresponse of a spike burst and an IPSP to the corticalstimulation (Fig. 7C) as to the acoustic stimulus. Theresponses to acoustic stimuli were, however, different atdifferent times after the neurone’s spontaneous firing.This neurone responded with (a) a burst of two spikesto the acoustic stimulus when the acoustic stimulus waspresented well after the spontaneous firing, (b) one spiketo the same acoustic stimulus when the acoustic stimuluscame at about 80 ms after the spontaneous firings, and (c)an EPSP only when the acoustic stimulus came at about20 ms after the spontaneous firings (Fig. 7D).
Effect of the number of pulsesin the electrical stimulation
The neurone in Fig. 8 showed a strong IPSP of > 10 mVto the acoustic stimulus. A brief EPSP was detected beforethe IPSP (Fig. 8A). The neurone responded to the corticalstimulation with a strong IPSP of > 10 mV (Fig. 8B). AnEPSP was detected between the first and second electrical
Figure 4. Mean amplitudes, rise times, decay times anddurations of the postsynaptic potentials of the MGB neuronesevoked by an auditory stimulus and by cortical stimulationA, the means of the EPSPs evoked by the auditory stimuli (A Evoked,open columns) and by electrical cortical stimulation (E-Cx Evoked, greycolumns). B, the means of the A-Evoked and E-Cx-Evoked IPSPs.Comparisons were made between the means of the A-Evoked andE-Cx-Evoked IPSPs using Student’s paired t test. ∗ P < 0.05;∗∗ P < 0.01.
C© The Physiological Society 2004

212 Y. Xiong and others J Physiol 560.1
Figure 5. The anatomical locations of the MGB neurones showed varied response patterns to electricalcortical stimulationThe neurones in A–C showed EPSP responses to the acoustic stimulus and EPSP responses to electrical corticalstimulation. The neurones in (D–F) showed inhibitory responses to electrical cortical stimulation. The leftpanels show physiological responses to acoustic stimuli and to electrical stimulation; the centre panelsshow the morphologies of the recorded neurones that were labelled with Neurobiotin; the right panels show lower
C© The Physiological Society 2004

J Physiol 560.1 MGB responses to cortical stimulation 213
pulses. The duration of the IPSP evoked by the corticalstimulation of 5 pulses was greater than that evoked bythe acoustic stimulus. However, the IPSP evoked by thecortical stimulation was shortened when the number ofelectrical pulses was reduced from five to one, as shownin Fig. 8C. This neurone was located in the non-lemniscalMGB at the caudomedial nucleus (Fig. 8D).
For some neurones, the number of pulses in theelectrical stimulation made no difference to corticofugalmodulation, as seen in the example shown in Fig. 1A. Theneurone showed an EPSP to the acoustic stimulus, andresponded to the electrical stimulation of one pulse witha spike(s) and an EPSP. The EPSP evoked by a corticalstimulation of 20 pulses was similar to that evoked by onepulse.
The corticofugal hyperpolarization of the membranepotential induced a low-threshold calcium spikeresponse to an acoustic stimulus
Figure 9 illustrates a representative example of burstingauditory response ‘conditioned’ by a train of corticalstimulation. In this neurone, which had a restingmembrane potential of −62 mV, cortical stimulationinduced a large membrane hyperpolarization (> 15 mV).The acoustic stimulation delivered during the returningphase of the IPSP subsequently evoked an LTS burst.With a further gradual decrease in the amplitude of theIPSP, i.e. to 200–300 ms after the cortical stimulation, theneurone returned to single-spike mode of spontaneousfiring, suggesting that cortically induced IPSPs may play acritical role in inducing a burst auditory response.
Discussion
Corticofugal depolarizationon auditory-EPSP neurones
Most auditory-EPSP neurones receive an excitatory inputfrom the cortex. Previous extracellular studies haveindicated that cortical stimulation has a facilitatory effecton the functionally matched sites in the thalamus and inthe inferior colliculus (IC), especially in their principlenuclei (He, 1997; Gao & Suga, 1998; Zhou & Jen, 2000; Heet al. 2002). Corticofugal modulation has a very broadfacilitatory effect on the thalamus of the guinea pig,especially on the lemniscal MGB (He et al. 2002; Yuet al. 2004a). Five of the seven auditory-EPSP neurones
magnifications for the sections after they were counterstained with neutral red. Recorded neurones injected withNeurobiotin were labelled in dark brown after the DAB reaction, as shown in the centre panels. The neurones inthe middle panels of (A–C and E) were photographed before they were further processed with neutral red staining,as shown in the right panels. The subdivisions of the MGB were parcelled based on the neural architectures ofneutral red staining. v, Ventral nucleus; s, shell nucleus; cm, caudomedial nucleus; rm, rostromedial nucleus of theMGB. Scale bars: centre panels, 100 µm; right panels, 1000 µm.
were located in the lemniscal MGB. Cortical stimulationdepolarized the membrane potential of five neurones,while the remaining neurones showed no effect. Thepresent study once again supports the finding that cortico-fugal modulation has a mainly facilitatory effect on thelemniscal MGB and that a modulation from any site of theauditory cortex has a wide coverage of the lemniscal MGB.
In the present study, we found that the shapes ofthe EPSPs caused by the acoustic stimulus and corticalstimulation were similar (Figs 1, 2, 4 and 5). Previousstudies showed that about half of the synapses on athalamic relay neurone are RS terminals (small profileswith rounded vesicles, as defined by Guillery, 1969 andRalston et al. 1988) (Jones & Powell, 1969a,b; Liu et al.1995a). The majority of RS terminals appear to be derivedfrom corticothalamic fibres (Jones & Powell, 1969a).The RS terminals have their main contact on the distaldendrites, while the ascending afferent terminals (RLtypes) end mainly on the proximal and intermediatedendrites (Liu et al. 1995b; Bartlett et al. 2000). We believethat the EPSPs evoked by acoustic stimuli were causedby the ascending afferent terminals, i.e. the RL terminals;and those evoked by cortical stimulation were causedby the descending corticothalamic terminals, i.e. theRS terminals.
The present report is the first to quantitatively examineboth EPSPs evoked by acoustic stimuli and those evokedby electrical cortical stimulation. Surprisingly, both theamplitudes and durations of the two EPSPs were foundto be similar. Based on the above discussion, the presentresults suggest that the shape and persistence of the EPSPdepend on the intrinsic characteristics of the neurone, noton the physical location of the synapses on the neurone.
Auditory IPSP neurones receive corticofugal inhibition
Most of the neurones that showed IPSPs to acousticstimuli were hyperpolarized by cortical stimulation. Noneof them showed a pure excitatory effect from cortico-fugal modulation. The absence of direct cortical excitationis puzzling, as anatomical studies have demonstrated arobust and direct glutamatergic innervation from layer 5and 6 cortical neurones to the dorsal division of the MGB(MGd) (Ojima, 1994; Bajo et al. 1995; Winer et al. 1999). Itis possible that a large proportion of the EPSPs are maskedby the strong IPSPs as shown in the ventrolateral thalamus(Deschenes & Hu, 1990). Further experiments are requiredto address this issue.
C© The Physiological Society 2004

214 Y. Xiong and others J Physiol 560.1
Figure 6. Response of the membrane potential to an auditorystimulus, spontaneous spike, and cortical stimulation of anMGB neuroneA, the neurone responded to repeated noise-burst stimuli with IPSPs.B, changes in the membrane potential after a spontaneous spike.C, reponses to cortical stimulations. The scale bars of the membranepotential and time apply to all traces.
Figure 7. Responses of the membranepotential to auditory stimuli, spontaneousspikes and cortical stimulations of an MGBneuroneA, the neurone responded to a noise-burst stimuluswith two spikes and an IPSP. B, change in themembrane potential after spontaneous spike trains.The inset shows the indicated area at a highermagnification. C, response to cortical stimulation.D, responses to auditory stimuli in variouspostspontaneous periods. The scale bars ofmembrane potential and time apply to all tracesexcept that in the inset of B.
When comparing the shapes of the IPSPs evoked byacoustic stimuli and by electrical stimulation of theauditory cortex, it is apparent that they show similar riseand decay times, but that the IPSPs evoked by corticalstimulation have a significantly greater mean durationthan those evoked by acoustic stimuli. The longer durationof the IPSPs evoked by the electrical stimulation waspartly caused by the multiple pulses (Fig. 8C). In arecent extracellular study, He (2003a) observed a mostlyinhibitory effect on the ‘on’ responses of non-lemniscalMGB neurones following cortical activation. In someextreme cases, the ‘on’ response was switched off bythe cortical stimulation. The corticofugal inhibitionwas widespread (He, 2003a). The strong corticofugalinhibition was thought to have been created via thecortico-reticulo-thalamic pathway (Golshani et al. 2001;He, 2003a; Yu et al. 2004a). In a thalamocortical slicepreparation, MGd neurones responded to thalamicradiation stimulation with a long-duration IPSP of over200 ms, suggesting that the stimulation activated thethalamic reticular nucleus (TRN) fibres, which in turninhibited the MGB neurones through GABAB receptors(Bartlett & Smith, 1999).
C© The Physiological Society 2004

J Physiol 560.1 MGB responses to cortical stimulation 215
Figure 8. Anatomical location and responsesof membrane potential to acoustic stimuli andcortical stimulations of an MGB neuroneA, responses to repeated acoustic stimuli. B and C,responses to electrical stimulations of one and fourpulses. Scale bars in C apply to A–C. D, anatomicallocation of the recorded neurone. The neurone waslabelled with Neurobiotin and counterstained withneutral red. The inset at the top right shows theindicated area at a higher magnification. Scale bar,1000 µm.
The following links can now be made: auditory-IPSPneurones are located in the non-lemniscal MGB (Yu et al.2004b; Fig. 5); neurones in the non-lemniscal MGB receivecorticofugal inhibition (He, 2003a; Yu et al. 2004a); andauditory-IPSP neurones receive corticofugal inhibition(present results).
An interesting result found in the present study was thatthe IPSPs caused by both auditory stimuli and corticalstimulation had a similar waveform. This result impliesthat the two inhibitions were possibly from the samesource, the TRN. In other words, some of the acousticallyevoked IPSPs that were observed in the present studymight be caused by the recurrent inhibition from theTRN. An acoustic stimulus evokes spikes in lemniscalMGB neurones with a short latency of < 10 ms (He, 2002).The lemniscal MGB neurones innervate neurones in theauditory cortex and GABAergic neurones in the TRN. TheTRN neurones, in turn, inhibit the non-lemniscal MGB,causing a strong IPSP with a longer latency, as observed inthe present study. The neurones that responded to acousticstimuli with an EPSP or spikes before an IPSP (Fig. 7A, andthe accompanying paper, Yu et al. 2004b) might receive anexcitatory input from the ascending pathway before theTRN inhibition.
The TRN receives inputs from both the thalamus andthe cortex (Jones, 1975). Recent physiological resultsindicate that the TRN terminals have a very stronginhibitory effect on the thalamic relay neurones (Bartlettet al. 2000; Golshani et al. 2001). We observed that(a) an auditory stimulus evoked an IPSP (Fig. 6A), (b)a spontaneous spike was always followed by an IPSP witha short time delay of about 7 ms (Fig. 6B), and (c) cortical
stimulation resulted in a similar shape of IPSP (Fig. 6C).The spontaneous spike of this neurone would activate theTRN neurone(s), which in turn would inhibit a groupof thalamic neurones, including the neurone in Fig. 6.The fact that similar IPSPs can be evoked by an auditorystimulus, a spontaneous spike, and cortical stimulationfurther supports the view that the TRN is the source of theinhibition.
Corticofugal inhibition causes low-thresholdcalcium spikes
Electrical activation of the auditory cortex causeda hyperpolarization of the membrane potential for
Figure 9. Neuronal responses to a combination of electricalstimulation and an auditory stimulusThe resting membrane potential was −62 mV and the scale bars applyto both traces.
C© The Physiological Society 2004

216 Y. Xiong and others J Physiol 560.1
most non-lemniscal MGB neurones. After the cortico-fugal hyperpolarization, thalamic neurones responded tosubsequent acoustic stimulus with an LTS burst (Fig. 9). Toour knowledge, this is the first time that such a mechanismof the natural auditory burst response has been shown inthe thalamus.
Although the functional implications of the LTS burstsare still debatable (Swadlow & Gusev, 2001; Sherman,2001; Steriade, 2001a,b), our results suggest that cortico-fugal inhibition could have two opposite effects onthalamic neurones, depending on their resting membranepotentials. While the corticofugal inhibition suppressedthe neuronal responses when the membrane potentialwas > −74 mV, it evoked LTS spikes/bursts when themembrane potential was < −74 mV. These results areconsistent with the finding in a recent report by Mooneyet al. (2004), that neuromodulatory inputs may lead todifferent patterns of synaptic output depending upon thelevel of their resting membrane potential.
Concluding remarks
The corticofugal modulation of MGB neurones thatshowed EPSP responses to acoustic stimuli could be bothfacilitatory and inhibitory, although the majority of suchneurones exhibited a facilitatory effect. The majority of theneurones that showed an EPSP both to acoustic stimuliand to the electrical stimulation of the auditory cortex arelocated in the lemniscal MGB. These two types of EPSPhave a similar shape in terms of amplitude, rise time, decaytime, and duration, implying that the shape of the EPSPdepends on the intrinsic properties of the cell membrane,not on the physical location of the synapses.
The neurones that showed an IPSP or an IPSPpreceded by an EPSP/spike(s) exhibited an IPSP toelectrical stimulation of the auditory cortex and werelocated in the non-lemniscal MGB. Some of these neuronesshowed a similar shape of IPSP after a spontaneous spike.This evidence supports the notion in the accompanyingpaper that the IPSPs are caused by the TRN feedbackinhibition to the thalamus (Yu et al. 2004b). A spontaneousspike of MGB neurone(s) might trigger the ‘big guardian’,i.e. the TRN neurone(s), in generating action potentials.This, in turn, will inhibit a group of non-lemniscal MGBneurones, resulting in IPSPs.
References
Andersen RA, Knight PL & Merzenich MM (1980). Thethalamocortical and corticothalamic connections of AI, AII,and the anterior auditory field (AAF) in the cat: Evidence fortwo largely segregated systems of connections. J Comp Neurol194, 663–701.
Bajo VM, Rouiller EM, Welker E, Clerke S, Villa AEP,de Ribaupierre Y & de Ribaupierre F (1995). Morphologyand spatial distribution of corticothalamic terminalsoriginating from cat auditory cortex. Hear Res 83, 161–174.
Bartlett EL & Smith PH (1999). Anatomic, intrinsic, andsynaptic properties of dorsal and ventral division neurons inrat medial geniculate body. J Neurophysiol 81, 1999–2016.
Bartlett EL, Stark JM, Guillery RW & Smith PH (2000).Comparison of the fine structure of cortical and collicularterminals in the rat medial geniculate body. Neuroscience100, 811–828.
Blumenfeld H & McCormick DA (2000). Corticothalamicinputs control the pattern of activity generated inthalamocortical networks. J Neurosci 20, 5153–5162.
Deschenes M & Hu B (1990). Electrophysiology andpharmacology of corticothalamic input to lateral thalamusnuclei: an intracellular study in the cat. Eur J Neurosci 2,140–152.
Edeline JM, Hars B, Maho C & Hennevin E (1994). Transientand prolonged facilitation of tone-evoked responses inducedby basal forebrain stimulations in the rat auditory cortex.Exp Brain Res 97, 373–386.
Gao E & Suga N (1998). Experience-dependent corticofugaladjustment of midbrain frequency map in bat auditorysystem. Proc Natl Acad Sci U S A 95, 12663–12670.
Golshani P & Jones EG (1999). Synchronized paroxysmalactivity in the developing thalamocortical network mediatedby corticothalamic projections and ‘silent’ synapses.J Neurosci 19, 2865–2875.
Golshani P, Liu X-B & Jones EG (2001). Differences in quantalamplitude reflect GluR4-subunit number at corticothalamicsynapses on two populations of thalamic neurons. Proc NatlAcad Sci U S A 98, 4172–4177.
Guillery RW (1969). The organization of synapticinterconnection in the laminae of the dorsal lateral geniculatenucleus of the cat. Z Zellforsh Mikrosk Anat 96, 1–38.
He J (1997). Modulatory effects of regional cortical activationon the onset responses of the cat medial geniculate neurons.J Neurophysiol 77, 896–908.
He J (2002). OFF responses in the auditory thalamus of theguinea pig. J Neurophysiol 88, 2377–2386.
He J (2003a). Corticofugal modulation on both ON and OFFresponses in the non-lemniscal auditory thalamus of theguinea pig. J Neurophysiol 89, 367–381.
He J (2003b). Slow oscillation in non-lemniscal auditorythalamus. J Neurosci 23, 8281–8290.
He J, Yu YQ, Xiong Y, Hashikawa T & Chan YS (2002).Modulatory effect of cortical activation on the lemniscalauditory thalamus of the guinea pig. J Neurophysiol 88,1040–1050.
Jones EG (1975). Some aspects of the organization of thethalamic reticular complex. J Comp Neurol 162,285–308.
Jones EG & Powell TPS (1969a). Electron microscopy ofsynaptic glomeruli in the thalamic relay nuclei of the cat.Proc R Soc Lond B Biol Sci 172, 153–171.
Jones EG & Powell TPS (1969b). An electron microscopic studyof the mode of termination of corticothalamic fibers in thethalamic relay nuclei of the cat. Proc R Soc Lond B Biol Sci172, 173–185.
Liu X, Honda CN & Jones EG (1995a). Distribution of fourtypes of synapse on physiologically identified relay neuronsin the ventral posterior thalamic nucleus of the cat. J CompNeurol 352, 69–91.
C© The Physiological Society 2004

J Physiol 560.1 MGB responses to cortical stimulation 217
Liu X, Warren RA & Jones EG (1995b). Synaptic distribution ofafferents from reticular nucleus in ventroposterior nucleus ofcat thalamus. J Comp Neurol 352, 187–202.
Mooney DM, Zhang L, Basile C, Senatorov VV, Ngsee J, OmerA & Hu B (2004). Distinct forms of cholinergic modulationin parallel thalamic sensory pathways. Proc Natl Acad SciU S A 101, 320–324.
Ojima H (1994). Terminal morphology and distribution ofcorticothalamic fibers originating from layers 5 and 6 of catprimary auditory cortex. Cereb Cortex 4, 646–663.
Ralston HJ III, Ohara PT, Raston DD & Chazal G (1988). Theneuronal and synaptic organization of the cat and primatesomatosensory thalamus. In Cellular Thalamic Mechanisms,ed. Bentivoglio M & Spreafico R, pp. 127–141. Elsevier,Amsterdam.
Redies H, Sieben U & Creutzfeldt OD (1989). Functionalsubdivisions in the auditory cortex of the guinea pig. J CompNeurol 282, 473–488.
Ryugo DA & Weinberger NM (1976). Corticofugal modulationof the medial geniculate body. Exp Neurol 51, 377–391.
Sherman SM (2001). A wake-up call from the thalamus. NatNeurosci 4, 344–346.
Steriade M (2000). Corticothalamic resonance, states ofvigilance and mentation. Neuroscience 101, 243–276.
Steriade M (2001a). To burst, or rather, not to burst. NatNeurosci 4, 671.
Steriade M (2001b). Impact of network activities on neuronalproperties in corticothalamic systems. J Neurophysiol 86,1–39.
Suga N, Zhang Y & Yan J (1997). Sharpening of frequencytuning by inhibition in the thalamic auditory nucleus of themustached bat. J Neurophysiol 77, 2098–2114.
Swadlow HA & Gusev AG (2001). The impact of ‘bursting’thalamic impulses at a neocortical synapse. Nat Neurosci 4,402–408.
Villa AEP, Rouiller EM, Zurita SP, de Ribaupierre Y & deRibaupierre F (1991). Corticofugal modulation of theinformation processing in the auditory thalamus of the cat.Exp Brain Res 86, 506–517.
Watanabe T, Yanagisawa K, Kanzaki J & Katsuki Y (1966).Cortical efferent flow influencing unit responses of medialgeniculate body to sound stimulation. Exp Brain Res 2,302–317.
Winer JA & Larue DT (1987). Patterns of reciprocity inauditory thalamocortical and corticothalamic connections:Study with horseradish peroxidase and autoradiographicmethods in the rat medial geniculate body. J Comp Neurol257, 282–315.
Winer JA, Larue DT & Huang CL (1999). Two systems of giantaxon terminals in the cat medial geniculate body:convergence of cortical and GABAergic inputs. J CompNeurol 413, 181–197.
Yu YQ, Xiong Y, Chan YS & He JF (2004a). Corticofugal gatingof auditory information in the thalamus: an in vivointracellular recording study. J Neurosci 24, 3060–3069.
Yu YQ, Xiong Y, Chan YS & He JF (2004b). In vivo intracellularresponses of the medial geniculate neurones to acousticstimuli in anaesthetized guinea pigs. J Physiol 560, 191–205.
Zhou X & Jen PHS (2000). Brief and short-term corticofugalmodulation of subcortical auditory responses in the bigbrown bat, Eptesicus fuscus. J Neurophysiol 84, 3083–3087.
Acknowledgements
This study was supported by grants from the Hong KongResearch Grants Council (CERG PolyU5211/99M and CERGPolyU5407/03M). We thank Mr S. S. M. Chan of The Universityof Hong Kong for his excellent technical assistance in theexperiments, and Dr L. Kitzes of the University of Californiaat Irvine and Dr B. Hu of the University of Calgary for criticallyreviewing an earlier version of the manuscript.
Author’s present address
Y.-Q. Yu: Department of Physiology, The University of ZhejiangSchool of Medicine, Hangzhou, Zhejiang, China.
C© The Physiological Society 2004





























