Embed Size (px)
Citation preview

Folia Microbiol. 38 (6), 509-514 (1993)
Effects of Photon Flux Density, CO2, Aeration Rate, and Inoculum Density on Growth and Extracellular Polysaccharide Production by Porphyridium cruentum M. IQBAL and S.I. ZAFAR
Biotechnology and Food Research Centre, PaMstan Council o f Scientific and Industrial Research Laboratories Complex, Lahore - 54600, Pakistan
Received March 23, 1993 Revised version July 13, 1993
ABSTRACT. Growth and extracellular polysaccharide production by Porphyridium cruentum were measured as a function of several culture parameters. Photon flux density of 75/zmol m -2 s -1 and CO 2 concentration of 2.5 % were found to be opti- mum for both growth and extracellular polysaccharide production. Interactive studies on these two parameters further con- firmed that at these levels of photon flux density and CO2, when applied together, both growth (5.9 �9 107 cells per mL) and extracellular polysaccharide production (1.9 g/L) were at the maximum. Maximum growth and extracellular polysaccharide production were observed at inoculum density of 106 cells per mL and aeration rate of 500 mL air per min per liter.
Abbreviations used: EP extracellular polysaccharide, PFD photon flux density
The unicellular red alga Porphyridium cruentum produces large amounts of extracellular polysaccharide (EP) which have a number of potential uses in various industries as gelling, thickening, stabilizing and emulsifying agents (Arad et al. 1985). They can also be used as polymers in the fluid systems for forcing oil to the surface in tertiary oil recovery (Savins 1978).
For the achievement of maximum yield of these polysaccharides, coincident with good culture growth, optimization of culture conditions is of essence. A number of studies have been reported on the growth and EP production by P. cruentum as affected by salinity, pH and temperature (Jones et al.
1963; Golueke and Oswald 1962; Sommerfeld and Nichols 1970, Minkova et al. 1987; Gudin and Chaumont 1991) and nitrogen (Arad et al. 1988; Iqbal et al. 1992). The role of incident light as an important growth parameter has also been investigated in terms of photon flux density (Thepenier and Gudin 1985; Lee and Tan 1988; Dermoun et al. 1992). These studies have been, however, limited to either photoperiodic and continuous illumination or the effect of high and low intensity of light. Cells growing under continuous illumination were observed to have a higher growth rate than those exposed to photoperiodic light (von Witsch et al. 1983). Likewise, PFD has been associated with shifts in cell metabolic activity as noted by increased protein level at low PFD and greater EP production at high PFD (Arad et al. 1985), and changes in the proportion of fatty acid production under varying PFD (l~ezanka et al. 1987). Another important input for the optimization of algal culture conditions is the supply of CO2 as the carbon source. P. cruentum cultures were observed to grow much faster when supplied with CO2 than cultures grown without it (Jones et al. 1963). Accordingly, CO2 has been used as the carbon source in studies with P. cruentum, be they physiological, biochemical or biotechnological (Gantt and Lipschultz 1972; Thepenier et al. 1985; l~ezanka et al. 1987; Lee and Vonshak 1988; Cohen and Arad 1989). These studies, however, made no attempt to optimize the concentration of CO2 to be supplied for maximum growth and polysaccharide production. Optimum levels of PFD and CO2 are further dependent on other cultural conditions. Most important among these, having a direct rela- tionship with the availability of light and CO2, are the initial inoculum density and culture aeration (Richmond 1987). The initial inoculum density controls the overall population density whereas the aer- ation rate affects the duration of exposure of each individual cell to the internal cultural environment and facilitates the diffusion of CO2 into the medium (Richmond 1986). However, no information is available on these important cultural parameters in respect of P. cruentum.
In view of the present limited or lacking knowledge on the cited interacting algal culture parameters, a study on the effects of a range of PFD values, CO2 concentration, inoculum density, and aeration rate on the optimization of cultural conditions of P. cruentum for biomass and EP production is reported here.

5"10 M. IQBAL and S.l. ZAFAR Vol. 38
M A T E R I A L S AND M E T H O D S
Porphyridium cmentum cultures were obtained from the Culture Collections for Algae and Protozoa, Cambridge (England) and were grown in artificial sea water medium (Jones et al. 1963). For investigations of the effect of CO2, cells were cultured with air supplemented with different levels of CO2 (2.5, 5.0, 7.5, 10 %) and maintained at a continuous PFD of 50/~mol m -2 s -1. Cultures grown without CO2 served as control. For the determination of the effect of PFD, ceils were cultured under four different intensities (25, 50, 75 and 100/zmol m -2 s -1) and the cultures were aerated with air containing 2.5 % CO2 at a rate of 500 mL min- 1 L- 1. The interactive effects of various PFDs and CO2 concentration were studied in four different sets of combinations: for each of the four PFDs applied (25, 50, 75 or 100p.mol m -2 s-1), four different concentrations of CO2 (2.5, 5.0, 7.5 and 10 %) were investigated; zero CO2 served as control. For investigation of the effects of aeration rate, the algal cells were aerated at five different rates of 125, 250, 500, 750 or 1000 mL min -1 L -1 with air containing 2.5 % CO2. Cultures were maintained under continuous illumination at 75 p.mol m-2 s-1. The effect of inoculum density was measured by transferring 0.5, 1.0, 1.5, 2.0, 2.5 or 3.0 • 109 cells per L, taken from one-week-old cultures, to the artificial sea water medium. The cultures were illuminated continuously at a PFD of 75/zmol m -2 s-1 and aerated with air containing 2.5 % CO2.
Cultures were grown in 2-L custom-built V-shaped flat-sided photobioreactors (Iqbal et al. 1993a) and maintained for 21 d at 25 + 2 ~ The cell concentration was determined in a WSI counting chamber (Weber Scientific International Ltd., England). The level of EP produced in the medium was determined with Alcian blue reagent (Ramus 1977). The growth rate of exponentially growing cells was expressed as relative growth constant (K) and was calculated according to Jones et al. (1963) as
K = (logeN - logeNo)/t = 2.3/t" (loga0N - logl0N0)
where N is the cell concentration at time t, No is the cell concentration at time 0 and t is time in days. Statistical analysis of the data was carried out by Duncan's new multiple-range test a tp < 0.05.
R E S U L T S AND DISCUSSION
Effect of C02 concentration. Carbon dioxide was noted to promote growth at all the concen- trations investigated. However, a wide variation in growth in response to changes in CO2 concentration was observed (Fig. 1, top). The maximum culture density with highest growth rate was observed in cultures grown with 2.5 % CO2. Differences in both the growth rates and culture densities were not significant when cells were grown with 5 % CO2. Cultures supplied with 10 % COz, on the other hand, showed a much slower growth, being approximately 41% less than with 2.5 % CO2. It is evident that the effect of CO2 at lower concentrations was more pronounced than at higher ones.
Table I. Extracellular polysaccharide production (mg/L, xl02) byP. cruentum
CO2 concentration, %/growth rate K
0/0.63 2.5/1.07 5/1.02 7.5/0.73 10/0.67 7.8 14.0 13.5 10.2 8.7
Photon flux density,/.tmol m-2 s - 1/growth rate K
25/0.53 50/1.07 75/1.11 100/0.72 9.9 14.4 18.7 12.8
Aeration rate, mL min- 1 L- 1/growth rate K
125/0.45 250/0.85 500/0.94 750/0.87 1000/0.55 7.8 14.0 18.2 16.7 12.7
The pattern of CO2 effect on EP production was also similar to that for growth (Table I). EP production was observed to be maximum in cells grown with 2.5 % CO2 which, in comparison with cultures grown without CO2 supply, was 80 % greater. The maximum levels of EP after 21 d at

1993 EXTRACELLULAR POLYSACCHARIDE PRODUCTION BY P. cruentum 511
different CO2 concentrations ranged between 792 and 1429 mg/L. EP production at the highest con- centration of 10 % CO2, was significantly less, being only 59 % of that obtained with 2.5 % CO2.
These observations have shown that availability of 6 0 CO2 in the culture medium enhanced both growth and EP
N production. An earlier study on P. cruentum reported higher growth when cultures were supplied with 2 % CO2
0 (Jones et aL 1%3). Other workers have also reported the use of different concentrations of CO2 in their studies on P. cruentum, viz. 2.5 % (Clement-Metral 1976; Iqbal et al.
2 o 1993b), 3.0 % (Arad et al. 1985), 4 % (Cohen and Arad 1989) and 5 % (Vonshak et al. 1985). None of these have, however, attempted to optimize the CO2 concentration for
0 maximum growth and EP production. The present studies 0 suggest that among the CO2 concentrations used
N (2.5-10 %), the level of 2.5 % was most favorable for both cell growth and EP production.
o Effect o f photon f lux density. A significant difference both in growth and in EP production was observed under different PFD (Fig. 1, middle). Maximum growth was
20 achieved by cells grown at 75/zmol m -2 s-1. Culture den- sity at this PFD increased by 80 % as compared with cells
0 grown at 25/zmol m -2 s -1. Growth of cells at the lowest PFD of 25/zmol m -2 s-1 was also the slowest, which is
0 clearly indicated as limiting since stationary phase was not N achieved even after 21 d. The growth rate (K) of cells
3 0 grown at this PFD was only 0.53, which is less than half of that observed for cells grown at 75/zmol m-2s -1. The
20 amount of EP produced under different PFDs ranged from 995 to 1880 mg/L (Table I). Maximum EP produc- tion was achieved at 75/zmol m -2 s -1, which was 89 %
10 greater as compared with that achieved at 25/zmol m -2 s -1. A higher PFD beyond 75/~mol m -2 s -1 was ob- served to have an inhibitory effect resulting in a 32 % de-
0 5 10 15 20 d crease in EP production when the cells were cultured at 100/zmol m- 2 s- 1.
Fig. 1. Growth (culture density N, 1/nL) of Increase in PFD up to 75/xmol m- 2 s- 1 was noted to
P. cruentum; growth rate K - denominators o f numbers at curves; top: effect of CO2 concentra- enhance cell growth which may be regarded as the critical tion (%, numerators); middle: effect of photon light saturation point under the study conditions. The in- flux density 0stool m -2 s -1, numerators); bot- hibitory effect noted at the higher PFD may be attributed tom: effect of aeration rate (mL min -1 L -I , to disruption of chloroplast lamellae caused by exposure numerators), to high light intensities (Brody and Vatter 1959) or the
inactivation of certain enzymes which are involved in CO2 fixation (Codd and Stewart 1980; Anderson 1979). Additional support for these observations is afforded by the studies on Chlorella, which showed that increase in PFD first favors cell division and after the optimum PFD has been attained, any further increase in PFD inhibits cell division (Sorokin and Krauss 1%5). The results presented here suggest that for P. cruentum, the PFD of 75/zmol m-2 S-1 was most favorable for cell division and EP production.
Earlier reports are limited to the expression of high and low light intensities which at best is only relative, as for Arad et al. (1985) the low light refers to 3.8/stool m -2 s -1 and high light to 15gmol m -2 s -1, whereas for Minkova et al. (1987) 15/zmol m -2 s -1 was low light and 60/zmol m -2 s -1 whs high light. The interpretation of their findings based thereon is likewise, relative to the subjective classification of PFDs. On a quantitative scale, however, both values considered high by Arad et aL (1985) as 15/xmol m -z s -1, and by Minkova et al. (1987) as 60/~mol m -2 s -1, were in fact below the optimum saturation point for P. cruentum cultures which was at 75/~moI m-2s -1 as observed in the present studies. This explains why these workers did not observe any inhibitory effect and were able to record a continuous increase in growth and EP production at the PFD that they had used in their studies.
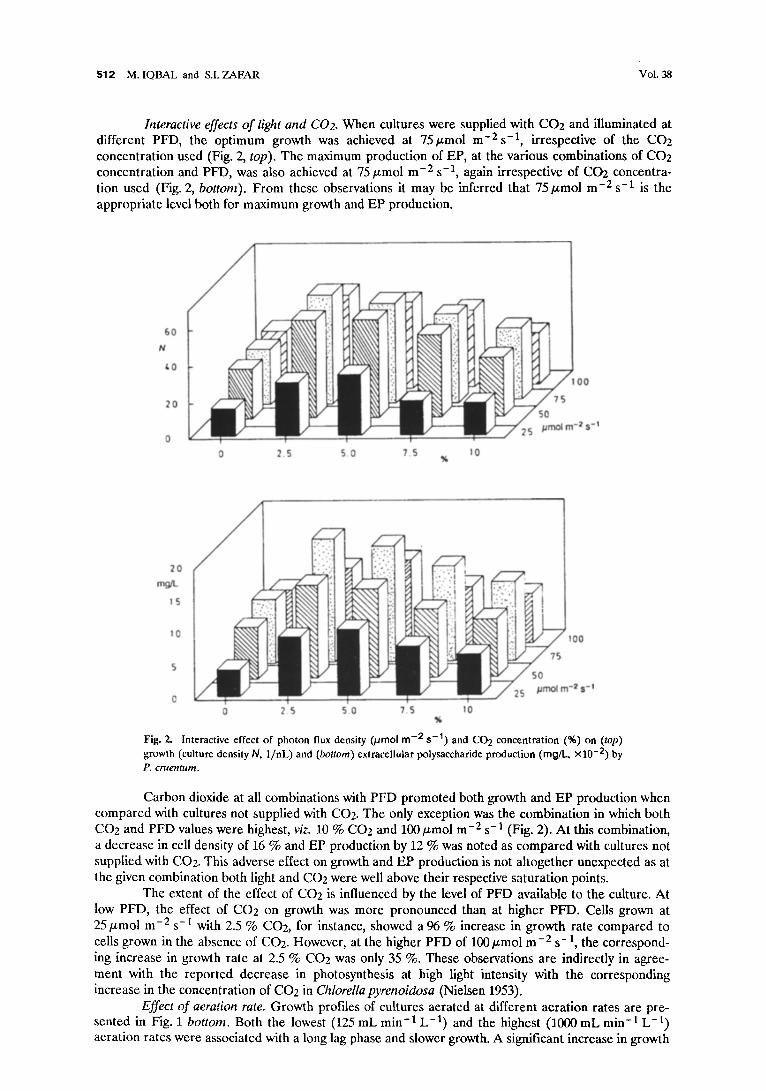
$1 2 M. IQBAL and S.I. ZAFAR Vol. 38
Interactive effects of light and C02. When cultures were supplied with CO2 and illuminated at different PFD, the optimum growth was achieved at 75/zmol m-2s -1, irrespective of the CO2 concentration used (Fig. 2, top). The maximum production of EP, at the various combinations of CO2 concentration and PFD, was also achieved at 75/zmol m -2 s -1, again irrespective of CO2 concentra- tion used (Fig. 2, bottom). From these observations it may be inferred that 75/.tmol m -2 s -1 is the appropriate level both for maximum growth and EP production.
Fig. 2. Interactive effect of photon flux density 0.tmol m -2 s - t ) and CO 2 concentration (%) on (top) growth (culture density h I, 1/nL) and (bottom) extracellular polysaccharide production (mg/I.., x l 0 -2) by P. cruentum.
Carbon dioxide at all combinations with PFD promoted both growth and EP production when compared with cultures not supplied with CO2. The only exception was the combination in which both CO2 and PFD values were highest, viz. 10 % CO2 and 100/zmol m -2 s-1 (Fig. 2). At this combination, a decrease in cell density of 16 % and EP production by 12 % was noted as compared with cultures not supplied with CO2. This adverse effect on growth and EP production is not altogether unexpected as at the given combination both light and CO2 were well above their respective saturation points.
The extent of the effect of CO2 is influenced by the level of PFD available to the culture. At low PFD, the effect of CO2 on growth was more pronounced than at higher PFD. Cells grown at 25/zmol m -2 s -I with 2.5 % CO2, for instance, showed a 96 % increase in growth rate compared to cells grown in the absence of CO2. However, at the higher PFD of 100 ~mol m-2 s-1, the correspond- ing increase in growth rate at 2.5 % CO2 was only 35 %. These observations are indirectly in agree- ment with the reported decrease in photosynthesis at high light intensity with the corresponding increase in the concentration of CO2 in Chlorella pyrenoidosa (Nielsen 1953).
Effect of aeration rate. Growth profiles of cultures aerated at different aeration rates are pre- sented in Fig. 1 bottont. Both the lowest (125 mL min -1 L -1) and the highest (1000 mL min -1 L -1) aeration rates were associated with a long lag phase and slower growth. A significant increase in growth
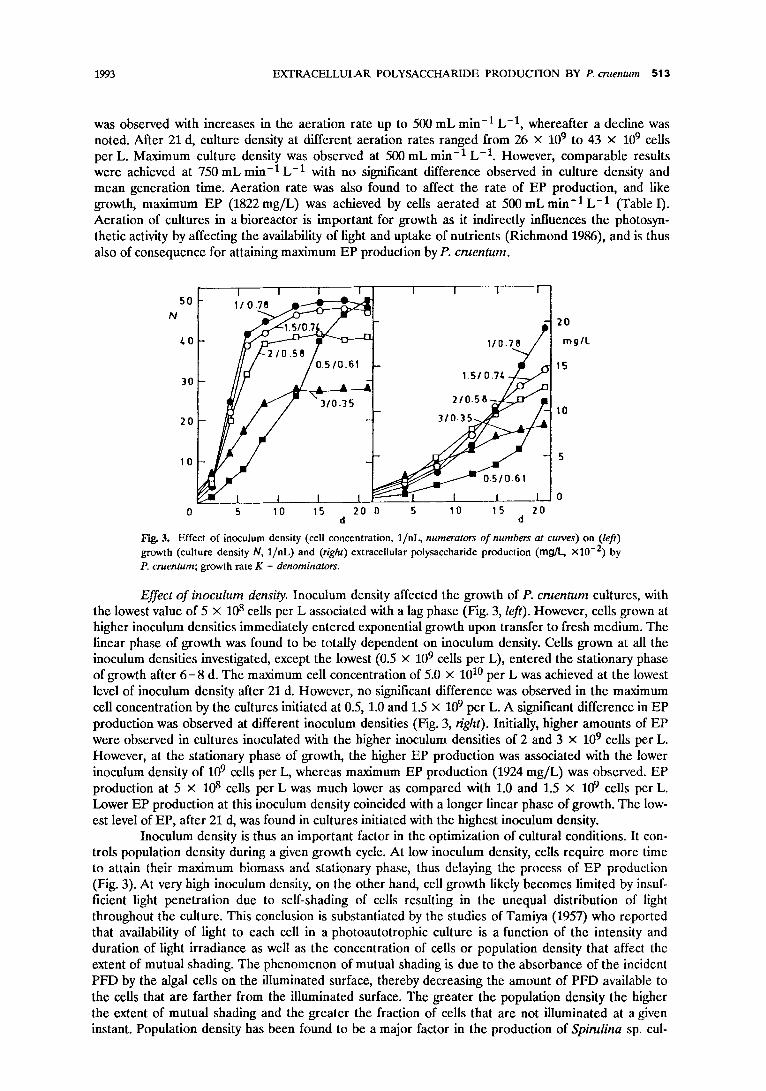
1993 EXTRACELLULAR POLYSACCHARIDE PRODUCTION BY P. cruentum 513
was observed with increases in the aeration rate up to 500 mL min -1 L -1, whereafter a decline was noted. After 21 d, culture density at different aeration rates ranged from 26 • 109 to 43 • 109 cells per L. Maximum culture density was observed at 500 mL min -1 L -1. However, comparable results were achieved at 750 mL min -1 L -1 with no significant difference observed in culture density and mean generation time. Aeration rate was also found to affect the rate of EP production, and like growth, maximum EP (1822 mg/L) was achieved by cells aerated at 500 mLmin -1 L - ] (Table I). Aeration of cultures in a bioreactor is important for growth as it indirectly influences the photosyn- thetic activity by affecting the availability of light and uptake of nutrients (Richmond 1986), and is thus also of consequence for attaining maximum EP production by P. cruentum.
50 N
40
30
20
10
l I I I
y .3/0.35.
5 10 15
I I I I
I/0.7~//
1.5 / 0 .Tt, / [ l
2/0.58~____J~j
20 0 5 10 15 ZO d d
20
mg/L
15
10
5
0
Fig. 3. Effect of inoculum density (cell concentration, 1/nL, numerators o f numbers at curves) on (left) growth (culture density N, 1/nL) and (right) extracellular polysaccharide production (rag/L, xl0 -2) by P. cruentum; growth rate K - denominators.
Effect o f inoculum density. Inoculum density affected the growth of P. cruentum cultures, with the lowest value of 5 • 108 cells per L associated with a lag phase (Fig. 3, left). However, cells grown at higher inoculum densities immediately entered exponential growth upon transfer to fresh medium. The linear phase of growth was found to be totally dependent on inoculum density. Cells grown at all the inoculum densities investigated, except the lowest (0.5 x 109 cells per L), entered the stationary phase of growth after 6 - 8 d. The maximum cell concentration of 5.0 • 1010 per L was achieved at the lowest level of inoculum density after 21 d. However, no significant difference was observed in the maximum cell concentration by the cultures initiated at 0.5, 1.0 and 1.5 x 109 per L. A significant difference in EP production was observed at different inoculum densities (Fig. 3, right). Initially, higher amounts of EP were observed in cultures inoculated with the higher inoculum densities of 2 and 3 • 109 cells per L. However, at the stationary phase of growth, the higher EP production was associated with the lower inoculum density of 109 cells per L, whereas maximum EP production (1924 rag/L) was observed. EP production at 5 x 108 cells per L was much lower as compared with 1.0 and 1.5 x 109 cells per L. Lower EP production at this inoculum density coincided with a longer linear phase of growth. The low- est level of EP, after 21 d, was found in cultures initiated with the highest inoculum density.
Inoculum density is thus an important factor in the optimization of cultural conditions. It con- trols population density during a given growth cycle. At low inoculum density, cells require more time to attain their maximum biomass and stationary phase, thus delaying the process of EP production (Fig. 3). At very high inoculum density, on the other hand, cell growth likely becomes limited by insuf- ficient light penetration due to self-shading of cells resulting in the unequal distribution of light throughout the culture. This conclusion is substantiated by the studies of Tamiya (1957) who reported that availability of light to each cell in a photoautotrophic culture is a function of the intensity and duration of light irradiance as well as the concentration of cells or population density that affect the extent of mutual shading. The phenomenon of mutual shading is due to the absorbance of the incident PFD by the algal cells on the illuminated surface, thereby decreasing the amount of PFD available to the cells that are farther from the illuminated surface. The greater the population density the higher the extent of mutual shading and the greater the fi'action of cells that are not illuminated at a given instant. Population density has been found to be a major factor in the production of Spirulina sp. cul-

514 M. IQBAL and S.I. Z A F A R Vol. 38
tured outdoor through the influence it exerts on the amount of light energy available to individual cells in the culture (Vonshak et al. 1982). Though no similar studies on indoor cultures have been reported, the effect of inoculum density on growth and EP production reported in the present communication is worth considering as a flexible tool for the optimization of any large-scale indoor cultivation studies on P. cruentum.
The general conclusion that may be drawn from the results presented above is that an inter- play of various levels of CO2, PFD, inoculum density, and aeration rate is important in the determina- tion of the optimum set of cultural conditions for maximum growth and EP production. Such a combination has a further significance in any attempt aimed at commercial production of EP by P. cruentum.
REFERENCES
ANDERSON L.E.: Interaction between phytochemistry and activity of enzymes, pp. 271-281 in Photosynthesis, IL Photosynthetic Carbon Metabolism and Related Processes (M. Gibbs, E. Latzko, Eds). Springer-Verlag, Berlin 1979.
ARAD S., ADDA M., COHEN E.: The potential of production of sulphate polysaccharides from Porphyridium. Plant & Soil 89, 117-127 (1985).
ARAD S., FmEDMAM O., ROTEM A.: Effect of nitrogen on polysaccharide production in a Porphyridium sp. AppLEnviron. Microbiol. 54, 2411-2414 (1988).
BRODY M., VATrER A.E.: Observations on cellular structures of Porphyridium cruentum. J.Biophys.Biochem.Cytol. 5, 289-299 (1959).
CLEMENT-METRAL J.D.: Preparation and some properties of protoplasts from red alga Porphyridium cruentum. J.Microsc.Biol.Cell 26, 167-172 (1976).
CODD G.A., STEWCART R.: Photoinactivation of ribulose bisphosphate carboxylase from green algae and cyanobacteria. FEMS Microbiol.Lett. 8, 237-240 (1980).
CO, tEN Z., ARAD S.: A closed system for outdoor cultivation of Porphyridium. Biomass 18, 5 9 - 67 (1989). DERMOUN D., CHAUMONT D., "rHEBAULT J., DAUTA A.: Modelling of growth of Porphyridium cruentum in connection with two
interdependent factors: Light and temperature. Bioresource Technol. 42, 113-117 (1992). GAtcrr E., LWSCHULTZ C.A.: Phycobilisomes of Porphyridium cruentum. ZCell Biol. 54, 313-324 (1972). GOLUEKE C.G., OSWALD W.J.: The mass culture of Porphyridium cruentum. Appl.Microbiol. 10, 102-107 (1962). GUDIN C., CHAUMOter D.: Cell fragility:, the key problem of microalgae mass production in close photobioreaetors. Bioresource
Teclmol. 38, 145 - 151 (1991). IQBAL M., GREY D., STEPAN-SARmSStAN G.: Effect of nitrogen on growth, extracellular polysaccharide and intracellular phyco-
erythrin production by the unicellular red alga Porphyridium cruentum. Acta Microbiol.Pol. 41, 65 - 73 (1992). IOBAL M., GREY D., STEPAN-SARKISSIAN G., FOWLER M.W.: A flat-sided photobioreactor for culturing of microalgae.
Aquacult.Eng. 12, 183-190 (1993a). IOBAL M., GREY D., STEPAN-SARKISSlAN G., FOWLER M.W.: Interactions between the unicellular red alga Porphyridium cruen-
turn and associated bacteria. Eur.J.Phycol. 28, 63 -68 (1993b). JONES R.F., SPEER H.L., KURV W.: Studies on the growth of the red alga Porphyridium cruentum. Physiol.Plant. 16, 636-643
(1963). LEE Y.K., TAN H.M.: Effect of temperature, light intensity and dilution rate on the cellular composition of red alga P o r p ~ -
dium cruentum in light chemostat cultures. MIRCENJ. 4, 231-237 (1988). LEE Y.K., VONSHAK A.: The kinetics of photoinhibition and its recovery in the red alga Porphyridium cruentum. Arch.Microbiol.
150, 529-533 (1988). MINKOVA K.M., GEORGIEV D.I., HOUBAVENS-KA N.B.: Light and temperature dependence of algal biomass and extracellular
polysaccharide production from P. cruentum. BioLPhysiol. 40, 8 7 - 8 9 (1987). NmLSEN E.S.: Carbon dioxide concentration, respiration during photosynthesis and maximum quantum yield of photosynthesis.
Plant Physiol. 40, 87 - 89 (1953). RA~US J.: Alcian Blue: A quantitative aqueous assay for algal acid and sulphated polysaccharides. J.PhycoL 13, 345 -348 (1977). I~,EZANKA W., DOUCHA J., MAREg P., PODOJIL M.: Effect of cultivation temperature and light intensity on fatty acid production
in the red alga Porphyridium cruentum. J.Basic Microbiol. 27, 275 - 278 (1987). RICHMOND A.: Outdoor mass cultures of microalgae, pp. 285-330 in MicroalgalMass Culture (A. Richmond, Ed.). CRC Press,
Boca Raton (Florida) 1986. RICHMOND A.: The challenge confronting industrial microagriculture: High photosynthetic efficiency in large-scale reactors.
Hydrobiologia 151/152, 117-121 (1987). SAVINS J.G.: Oil recovery process employing thickened aqueous driving fluid. US Pat. 4 079 544 (1978). SOMMERFELD M.R., NICHOLS H.W.: Comparative studies on the genus Porphyridium Naeg. J.PhycoL 6, 6 7 - 7 8 (1970). SOROKIN C., KRAUSS R.W.: The dependence of cell division in Chlorella on temperature and light intensity. Am.J.Bot. 52,
331 - 339 (1965). TAMIYA It.: Mass culture of algae. Ann.Rev.Plant Physiol. 18, 309- 334 (1957). THEPENIER C., GUDIN C.: Studies on optimal conditions for polysaccharide production byPorphyridium cruentum. MIRCENJ.
1, 257-268 (1983). THEPENIER C., GUDIN C., THOMAS n.: Immobilization of Porpl~idium cruentum in polyurethane foam for the production of
polysaccharide. Biomass 7, 225-240 (1985). VONSHAK A., COHEN Z., RICHMOND A.: The feasibility of mass cultivation ofPorphyridium. Biomass 8, 13-25 (1983). VONSHAK A., ABELIOVICH A., BOUSIBBA S., ARAD S., RICHMOND A.: Production of Spirulina biomass: Effects of environmental
factors and population density. Biomass 2, 175 - 185 (1982). VON WITSCH H., BOLZE A., HORNUNG J.: Production of biomass and of extracellular polysaccharides in batch cultures of Por-
phyridium aerugineum, Rhodophyceae. Ber.Dtsch.Bot.Ges. 96, 469-481 (1983).





























