Embed Size (px)
Citation preview

Effects of Plant Age on Photoperiodic Induction and Development of Flowers and Fruits in Abelmoschus esculentus (L.) Moench.
F.1. o. NWOKE
Department of Botany, University of Nigeria, Nsukka, Nigeria
Received September 9, 1982 . Accepted March 14, 1983
Summary
Photoperiodic induction and development of flower buds and fruits depend on the age at which short-day (SD) tratment is started. The minimum number of SD, which is required, decreases from 6 to 4 with increasing age up to 10 days. The time from the start of SD treatment to appearance of the first flower buds is constant in all conditions tested except if induction is performed by only 4 SD; in this case, the appearance time is delayed by 6 days. The number of flower buds increased with the number of SD cycles up to 15, and with the age up to 10 days. Likewise, the efficiency of the photoperiodic induction on flower and fruit development increased with the number of SD cycles and with the age of the plant.
Key words: Abelmoschus esculentus (L.) Moench. Malvaceae, short·day plant, flowering, fruiting, photoperiod, plant age.
Introduction
In photoperiodically sensitive plants, it has been established that the plant age, or the developmental stage, at the time of induction profoundly affects the initiation and subsequent development of flowers (Lang, 1965; Hillman, 1969). In many species a «juvenile phase» exists during which the plants are unable to flower even if subjected to inductive photoperiods. In trees, this phase usually lasts for a few years and can sometimes reach up to 40 years; but in herbaceous plants much shorter juvenile phases are found (Lang, 1965; Robinson and Wareing, 1969). Moreover, after the termination of the juvenile phase, the responsiveness to photoperiodic induction may increase with age (Borthwick and Parker, 1938; Cumming, 1959; Evans, 1960):
On the other hand, some herbaceous plants have little or no juvenile phase and can therefore be induced to flower in the seedling stage right after germination. Among those plants are SD species such as Chenopodium rubrum (Cumming, 1959) and Pharo bitis nil (Kujirai and Imamura, 1958), and LD species like Anagallis arvensis (Ballard and Grant Lipp, 1964) and Lolium temulentum (Evans, 1960). In these cases, however, a quantitative effect of age may exist, with the sensitivity to photoperiodic induction
Abbreviations: alt, alternate; cot, cotyledon; exp, expanded; lv, leaf (leaves); LD, long day, daylength 16 h; SD, short day, daylength 10 h.
z. Pjlanzenphysiol. Bd. 110. S.393-400. 1983.

394 F. I. O. NWOKE
increasing with age. There is usually a minimum number of leaves or nodes that must be produced before the appearance of the first flower bud (Borthwick and Parker, 1938; Kujirai and Imamura, 1958; Cumming, 1959; Evans, 1960; Lang, 1965).
Finally, in exceptional cases photoperiodic sensitivity can decrease with age (Ballard and Grant Lipp, 1964; Seidlova and Krekule, 1973).
Early okra, an early flowering and maturing variety of A. esculentus, is a short day plant with a critical photoperiod of 12.5 h (Njoku, 1958). In a previous study (Nwoke, 1980), early okra was shown to require a minimum of 6 SD cycles (10 h, natural daylight) for floral bud initiation. Nwoke (1980) showed also that the number of SD cycles influenced both flower and fruit development as more than 20 SD cycles were needed for the flower buds to develop till anthesis as well as for the growth and development of fruits. These results, however, have been obtained if the inductive treatment was started at the time of cotyledon release. It may well be that these minimum numbers of SD could be reduced if induction were started at a later stage in development. The present study, therefore, investigates the effect of plant age on the photoperiodic induction and development of flowers and fruits in the early okra.
Materials and Methods Four early okra seeds were planted in black plastic bags (20 x 12 em, and 0.004 em thick) fill
ed with soil, heap compost and dung (3:1:0.1 by vol.) and kept under 16h long days in a glasshouse. After germination, the plants were thinned to one per bag and watered twice each day. Starting from the secon.d week, the bags were flooded once each week with complete nutrient solution of Hewitt (1966) and the excess allowed to drain through. In an experiment on the effects of plant age on the induction and development of flower buds, 13 replicate plants each were moved from LD to continuous SD at the ages of 0 (at the time of cotyledon release), 5, 10, 20 and 30 days respectively. In other experiments, the number of SD had to be varied, in order to test its effect on formation and development of flowers and fruits on plants of various age and to find out the minimum number of SD required for flower bud formation. In these experiments, six sets of 13 plants each were moved from LD to SD at ages, 0, 5, 10,20 and 30 days, respectively, and then one of the six sets in each group was returned to LD after 1, 5, 10, 15,20 and 30 SD cycles or after 1 to 6 SD cycles each.
The 10h natural daylight was achieved by raising a hood of thick black cloth at 8h and lowering it again over a wooden frame constructed above the plants at 18 h. For the 16 h photoperiod, the natural daylight was extended with four 40 W Atlas tropical daylight fluorescent tubes between 18h and 22h by means of an automatic time switch and again between 6h and 8 h to allow for differences in the times of sunrise. This light source gave an irradiance of 450 Ix at bench level. These experiments were carried out during the months of June to October when the mean monthly maximum and minimum temperatures in the glasshouse were 28±2 °C and 23 ± 2°C respectively while the mean monthly relative humidity was 85 ± 2 %. The temperature under the thick black cloth shades was usually 2 °C above the minimum night temperature. All experiments were repeated at least twice.
In each experiment, observations were made on the number of days from the start of SD treatment to the time of macroscopic appearance of the first flower bud and to the time of first flower opening exposing corolla colour. The number of flower buds and fruits as well as the number of mature fruits produced on each plant were recorded. Sixty days after the start of the SD treatment, the dry weights of the fruits were obtained by drying in a forced air oven at
Z. Pjlanzenphysiol. Ed. 110. S.393-400. 1983.
Plant age effect on flowering and fruiting of Abelmoschus 395
100°C for 48 h. The fruits on each plant were dissected and the total number of seeds was counted and their dry weights as well as the 100 seed weights obtained.
Results
Flower induction and development
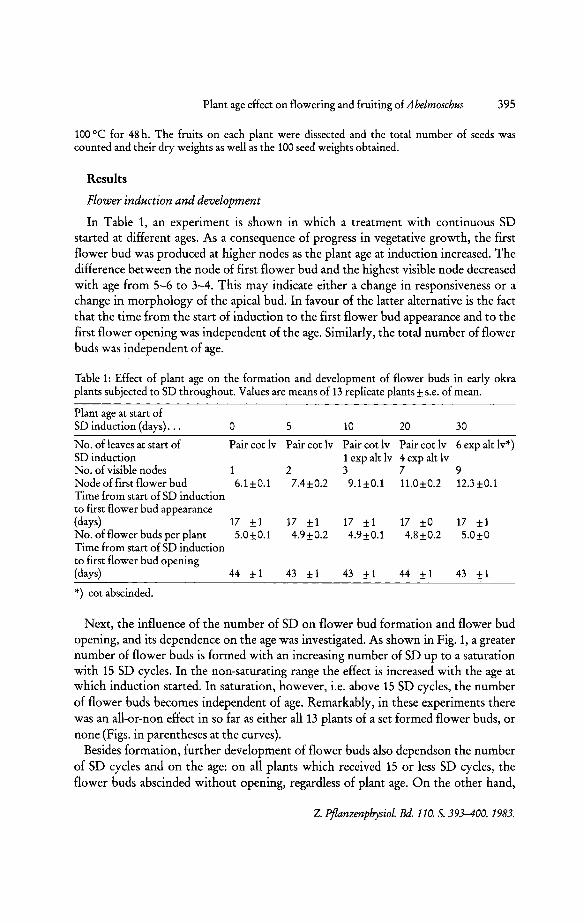
In Table 1, an experiment is shown in which a treatment with continuous SD started at different ages. As a consequence of progress in vegetative growth, the first flower bud was produced at higher nodes as the plant age at induction increased. The difference between the node of first flower bud and the highest visible node decreased with age from 5-6 to 3-4. This may indicate either a change in responsiveness or a change in morphology of the apical bud. In favour of the latter alternative is the fact that the time from the start of induction to the first flower bud appearance and to the first flower opening was independent of the age. Similarly, the total number of flower buds was independent of age.
Table 1: Effect of plant age on the formation and development of flower buds in early okra plants subjected to SD throughout. Values are means of 13 replicate plants ± s.e. of mean.
Plant age at start of SD induction (days) ... 0 5 10 20 30
No. of leaves at start of Pair cot Iv Pair cot Iv Pair cot Iv Pair cot Iv 6 exp alt Iv*) SD induction 1 exp alt Iv 4 exp alt Iv No. of visible nodes 1 2 3 7 9 Node of first flower bud 6.1±0.1 7.4±0.2 9.1 ±0.1 11.0±0.2 12.3 ±0.1 Time from start of SD induction to first flower bud appearance (days) 17 ±1 17 ±1 17 ±1 17 ±O 17 ±1 No. of flower buds per plant S.O±O.l 4.9±0.2 4.9±0.1 4.8±0.2 S.O±O Time from start of SD induction to first flower bud opening (days) 44 ±1 43 ±1 43 ±1 44 ±1 43 ±1
*) cot abscinded.
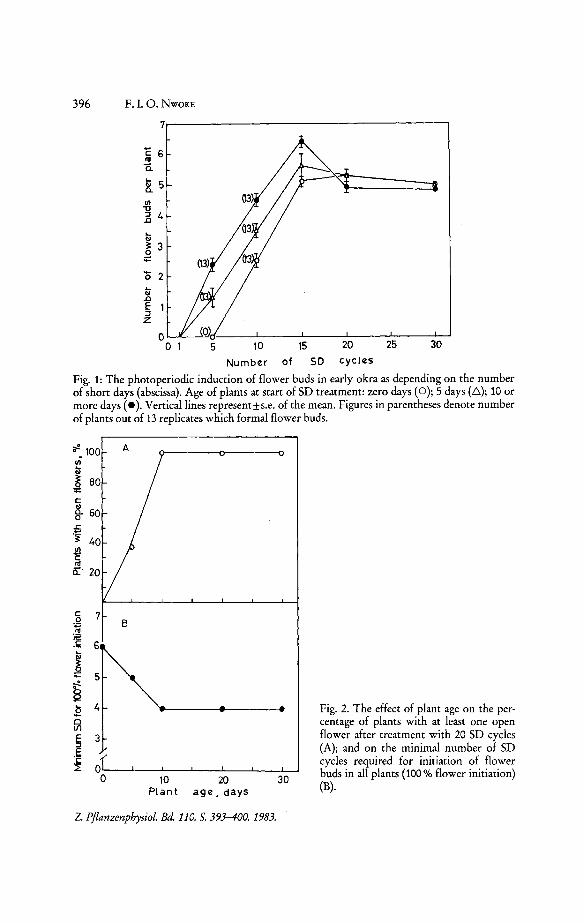
Next, the influence of the number of SD on flower bud formation and flower bud opening, and its dependence on the age was investigated. As shown in Fig. 1, a greater number of flower buds is formed with an increasing number of SD up to a saturation with 15 SD cycles. In the non-saturating range the effect is increased with the age at which induction started. In saturation, however, i.e. above 15 SD cycles, the number of flower buds becomes independent of age. Remarkably, in these experiments there was an all-or-non effect in so far as either alll3 plants of a set formed flower buds, or none (Figs. in parentheses at the curves).
Besides formation, further development of flower buds also dependson the number of SD cycles and on the age: on all plants which received 15 or less SD cycles, the flower buds abscinded without opening, regardless of plant age. On the other hand,
Z. Pjlanzenphysiol. Ed. 110. S.393-400. 1983.
396 F. 1. O. NWOKE
7
'E 6 III a. ~5 III "0 .E 4
... ... ~ 3 .g a 2 ... ...
..Q
E ::J Z
0 0 5 10 15 20 25 30
Number of SD cycles
Fig. I: The photoperiodic induction of flower buds in early okra as depending on the number of short days (abscissa). Age of plants at start of SD treatment: zero days (0 ); 5 days (6 ); 10 or more days (e). Vertical lines represent±s.e. of the mean. Figures in parentheses denote number of plants out of 13 replicates which formal flower buds.
III ~
~ c: a 0 ..c: .1: ~
£I c: III ;r
c: .Q ;; :-e .!; ...
~ 5 . .... s ~ 4 0 III
5 3
~ 0 0 10 20
Plant age. days
Z. Pjlanzenphysiol. Bd. 110. S.393-400. 1983.
30
Fig. 2. The effect of plant age on the percentage of plants with at least one open flower after treatment with 20 SD cycles (A); and on the minimal number of SD cycles required for initiation of flower buds in all plants (100% flower initiation) (B).

Plant age effect on flowering and fruiting of Abelmoschus 397
on all plants with 30 SD cycles the first 3 out of the 5 buds developed and opened, while the 2 later-formed buds abscinded, again regardless of age. However, after 20 SD cycles the number of viable flower buds became age-dependent. As can be seen in Fig. 2 A, the first out of the 5 buds developed and opened in an increasing percentage of plants, with saturation reached at the age of 10 days. In this saturation range, occasionally also a second bud opened.
In Fig. 1 it can be observed that all plants aged 5 or 10 days at induction formed flower buds in 5 SD while zero day-old plants did not. Therefore, the effect of plant age on the minimum number of SD cycles required for flower bud formation in all plants (100% flower initiation) was determined. As can be seen in Fig. 2B, the minimum number of SD required decreases from 6 to 4 cycles with increasing age. But beyond an age of 10 days no further reduction is possible. In those plants, however, which are induced to flower bud formation by 4 SD only, the time to the appearance of the first bud was delayed by 6 days (Table 2).
Taking together the data in Figs. 1, 2A and 2B, an age of about 10 days seems to be a saturation point beyond which age does no longer influence the photoperiodic response, as measured by the number of flower buds, flowerbud development, and photoperiodic sensitivity.
Fruit formation and development
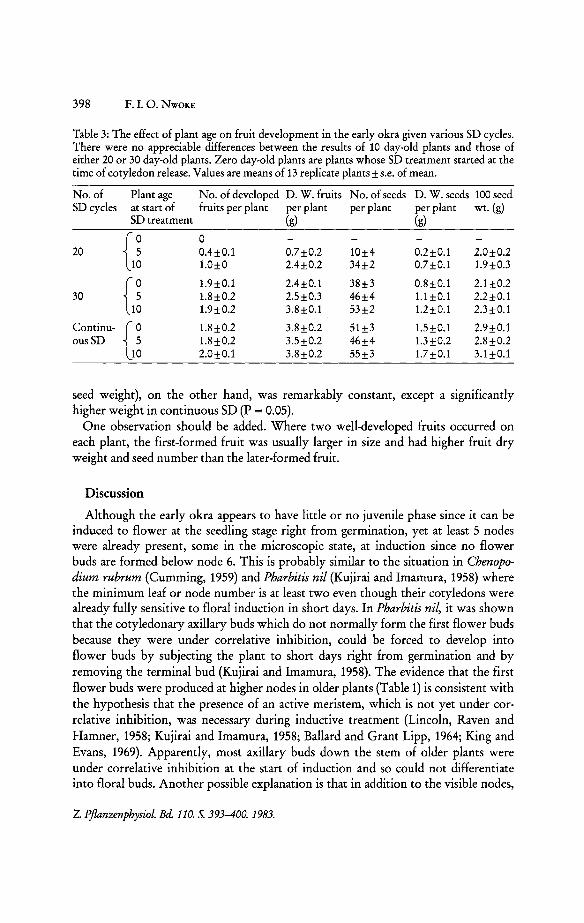
As had been shown in Fig. 2 A, after 20 SD cycles the number of buds which developed to open flowers depended on the age. Accordingly, the number of developed fruits ranged from zero to about one per plant for increasing ages from 0 to 10 days (Table 3). After 30 SD or in continuous SD, however, the fruit number became independent of the age: Out of three fruits produced per plant, two developed to ripeness (Table 3), whereas one usually abscinded in an early stage. Yet, there is some influence of age on the dry weight of total fruits per plant: maximum is reached with 30 SD for the age-l0 plants, but it requires continuous SD for the younger plants.
Both seed number and seed dry weight per plant increased as the number of SD cycles increased in all age groups (Table3). However, differences between the age groups are hardly significant. The average weight of the seeds (as measured by the 100
Table 2: Effect of plant age on the number of days from start of SD induction to first flower bud appearance in early okra plants subjected to various SD cycles. Values are means of 13 plants ± s.e. of mean.
Plant age No. ofSD cycles
(days) 4 5 6
0 17 ± 1 5 17±1 17±1
100r more 23±1 17±O 17±1
Z. Pjlanzenphysiol. Ed. 110. S.393-400. 1983.
398 F. I. O. NwoKE
Table 3: The effect of plant age on fruit development in the early okra given various SD cycles. There were no appreciable differences between the results of 10 day-old plants and those of either 20 or 30 day-old plants. Zero day-old plants are plants whose SD treatment started at the time of cotyledon release. Values are means of 13 replicate plants ± s.e. of mean.
No. of Plant age No. of developed D. W. fruits No. of seeds D. W. seeds 100 seed SD cycles at start of fruits per plant per plant per plant per plant wt. (g)
SD treatment (g) (g)
{J 0 20 0.4±0.1 0.7±0.2 10±4 0.2±0.1 2.0±0.2
1.0±0 2.4±0.2 34±2 0.7±0.1 1.9±0.3
{J 1.9±0.1 2.4±0.1 38±3 0.8±0.1 2.1±0.2 30 1.8±0.2 2.5±0.3 46±4 1.1 ±0.1 2.2±0.1
1.9±0.2 3.8±0.1 53±2 1.2±0.1 2.3±0.1
Continu-
{~ 1.8±0.2 3.8±0.2 51±3 1.5±0.1 2.9±0.1
ousSD 1.8±0.2 3.5±0.2 46±4 1.3±0.2 2.8±0.2 2.0±0.1 3.8±0.2 55±3 1.7 ±0.1 3.1 ±0.1
seed weight), on the other hand, was remarkably constant, except a significantly higher weight in continuous SD (P = 0.05).
One observation should be added. Where two well-developed fruits occurred on each plant, the first-formed fruit was usually larger in size and had higher fruit dry weight and seed number than the later-formed fruit.
Discussion
Although the early okra appears to have little or no juvenile phase since it can be induced to flower at the seedling stage right from germination, yet at least 5 nodes were already present, some in the microscopic state, at induction since no flower buds are formed below node 6. This is probably similar to the situation in Chenopodium rubrum (Cumming, 1959) and Pharbitis nil (Kujirai and Imamura, 1958) where the minimum leaf or node number is at least two even though their cotyledons were already fully sensitive to floral induction in short days. In Pharbitis nil, it was shown that the cotyledonary axillary buds which do not normally form the first flower buds because they were under correlative inhibition, could be forced to develop into flower buds by subjecting the plant to short days right from germination and by removing the terminal bud (Kujirai and Imamura, 1958). The evidence that the first flower buds were produced at higher nodes in older plants (Table 1) is consistent with the hypothesis that the presence of an active meristem, which is not yet under correlative inhibition, was necessary during inductive treatment (Lincoln, Raven and Hamner, 1958; Kujirai and Imamura, 1958; Ballard and Grant Lipp, 1964; King and Evans, 1969). Apparently, most axillary buds down the stem of older plants were under correlative inhibition at the start of induction and so could not differentiate into floral buds. Another possible explanation is that in addition to the visible nodes,
Z. Pjlanzenphysiol. Bd. 110. S.393-400. 1983.

Plant age effect on flowering and fruiting of Abelmoschus 399
which increase with age, there are several other nodes in the microscopic state prior to induction; but only the axillary buds of these leaves which are not yet visible are able to be determined to become flower buds. Consequently, the more nodes that are already developed before induction, the higher the node with the first responsive axillary bud.
The data presented in Figs. 1 and 2 B show that the sensitivity of the early okra to photoperiodic induction of flowers increased with increase in plant age and that maximum sensitivity was achieved by plants that were at least 10 days old. This is consistent with the general observation that photoperiodic plants usually require a lower number of inductive cycles, as they get older (Lang, 1965); although there is no general agreement whether the changes in sensitivity concern the stimulus-perceiving leaves or the responding apex (Zeevaart, 1958, 1969; Evans, 1960).
It has been shown that the development of flowers and fruits of the early okra is photoperiodic and controlled by the number of inductive SD cycles applied (Nwoke, 1980). The data presented here indicate that the minimum number of cycles needed for both flower and fruit development was less in older plants. Previous data have established that photoperiod influences the growth and development of flowers and fruits through its effect on the concentrations of various growth substances (promoters and inhibitors) produced by the leaves, flower buds and young fruits (Nitsch, 1971; Bentley et al., 1975; Coombe, 1976). However, the role of plant age in modifying either the production or the concentrations of growth substances in older plants so that lower minimal number of SD cycles would be required for better flower and fruit development is still not clear.
In some SD plants it has been shown that the increase of photoperiodic sensitivity with age is reflected also by a longer critical daylength (shorter critical dark period) of older plants (Long, 1939; Imamura and Takimoto, 1955; Imamura et aI., 1966). If the photoperiodic behaviour of a SD plant is to be characterized, it seems reasonable, therefore, to use plants which have reached their maximum photoperiodic sensitivity. For early okra, in consequence, the critical daylength of 12.5 h, as determined in just germinating plants, has to be considered as preliminary; it has to be reinvestigated in plants that are at least 10 days old.
Acknowledgements
I thank Mr. R. Abonyi and other staff of the Botanical Garden, University of Nigeria, Nsukka, for helping to maintain the plants and operate the dark chambers.
References
BALLARD, L. A. T. and A. E. GRANT LIpp: Juvenile photoperiodic sensitivity in Anagallis arve· nesis L. Subsp. Feemina (MilL) Schinz and Theil. Aust. J. BioL Sci. 17, 323-337 (1964).
BENTLEY, B., C. B. MORGAN, D. G. MORGAN, and F. A. SAAD: Plant growth substances and effects of photoperiod on flower bud development in Phaseolus vulgaris. Nature 256, 121-122 (1975).
Z. Pjlanzenphysiol. Bd. 110. S.393-400. 1983.

400 F. I. O. NwoKE
BORTHWICK, H. A. and M. W. PARKER: Effectiveness of photoperiodic treatments of plants of different age. Bot. Gaz. 100, 245-249 (1938).
COOMBE, B. G.: The development of fleshy fruits. A. Rev. PI. Physiol. 27, 207-288 (1976). CUMMING, B. G.: Extreme sensitivity of germination and photoperiodic reaction in the genus
Chenopodium (Tourn.) L. Nature 184,1044-1045 (1959). EVANS, L. T.: Inflorescence initiation in Lolium temulentum. 1. Effect of plant age and leaf area
on sensitivity to photoperiodic induction. Aust. J. BioI. Sci. 13, 123-131 (1960). HEWITT, E. J.: Sand and water culture methods used in the study of plant nutrition. 2nd edn.
Farnham Royal, Bucks: Commonwealth Agricultural Bureaux (1966). HILLMAN, W. S.: Photoperiodism and vernalization. In: M. B. WILKINS (ed.): Physiology of
plant growth and development, 559-601. McGraw-Hill, London, 1969. IMAMURA, S. and A. TAKIMOTO: Photoperiodic responses in Japanese morning glory, Pharbitis
nil Chois., a sensitive short day plant. Bot. Mag. Tokyo 68, 235-241 (1955). IMAMURA, S., M. MURAMATSU, S. 1. KITAJO, and A. TAKIMOTO: Verietal difference in photo
periodic behaviour of Pharbitis nil Chois. Bot. Mag. Tokyo 79, 714-721 (1966). KING, R. W. and L. T. EVANS: Timing of evocation and development of flowers in Pharbitis nil.
Aust. J. BioI. Sci. 22, 559-572 (1969). KUJIRAI, C. and S. IMAMURA: Dber die photoperiodische Empfindlichkeit der Kotyledonen von
Pharbitis nil Chois. Bot. Mag. Tokyo 71, 408-416 (1958). LANG, A.: Physiology of flower initiation. In: W. H. RUHLAND (ed.): Handbuch der Pflanzen
physiologie. Vol. 15, pt. 1, 1380-1536. Springer-Verlag, Berlin, 1965. LiNCOLN, R. G., K. A. RAVEN, and K. C. HAMNER: Certain factors influencing expression of the
flowering stimulus in Xanthium. II. Relative contribution of buds and leaves to effectiveness of inductive treatment. Bot. Gaz. 119,179-191 (1958).
LONG, E. M.: Photoperiodic induction as influenced by environmental factors. Bot. Gaz., 101, 168-188 (1939).
NITSCH, J. P.: Perenation through seeds and other structures: Fruit development. In: F. C. STEWARD (ed.): Plant Physiology - a treatise. Vol. VIA, 413-501. Academic Press, London and New York, 1971.
NJoKu, E.: The photoperiodic response of some Nigerian plants. JI. W. Afr. Sci. Ass. 4, 99-111 (1958).
NwoKE, F. I. 0.: Effect of number of photoperiodic cycles on flowering and fruiting in early and late varieties of okra [Abelmoschus esculentus (L.) Moench.]. J. expo Bot. 31, 1657-1664 (1980).
ROBINSON, L. W. and P. F. WAREING: Experiments on juvenile-adult phase change in some woody species. New Phytologist 68, 67-78 (1969).
SEIDLOV A, F. and J. KREKULE: The negative response of photoperiodic floral induction in Chenopodium rubrum L. to preceding growth. Ann. Bot. 37, 605-614 (1973).
ZEEVAART, J. A. D.: Flower formation as studied by grafting. Meded. Landbouwhogesch. Wageningen 58, 1-88 (1958).
ZEEVAART, J. A. D.: Perilla. In: L. T. EVANS (ed.): The induction of flowering: some case histories, 115-155. Macmillan of Australia, Melbourne, 1969.
Z. Pjlanzenphysiol. Bd. 110. S.393-400. 1983.








![Heterosis studies in okra [Abelmoschus esculentus (L ... · Okra [Abelmoschus esculentus (L.) Moench] is one of the most important vegetable crops grown extensively throughout the](https://img.pdfslide.net/doc/110x75/5e4c467e06af562e105d3a03/heterosis-studies-in-okra-abelmoschus-esculentus-l-okra-abelmoschus-esculentus.jpg)
![Physicochemical Properties of Abelmoschus esculentus seed oil · The acid value was determined following established procedure of AOAC [3]. Iodine value was estimated by applying](https://img.pdfslide.net/doc/110x75/61281bb19a00f468d360d1e2/physicochemical-properties-of-abelmoschus-esculentus-seed-oil-the-acid-value-was.jpg)



















