Embed Size (px)
Citation preview

Proc. Nati. Acad. Sci. USAVol. 91, pp. 4150-4154, May 1994Neurobiology
Effects on protein synthesis produced by pairing depolarizationwith serotonin, an analogue of associative learning in Aplysia
(associative plasticity/two-dlmensional polyacrylamide gel electrophoresis/abdominal ganglia/stringent starvation protein/glutathloneS-transferase)
F. NOEL*, C. KOUMENISt, M. NUNEZ-REGUEIROt, U. RAJUtt, J. H. BYRNE*, AND A. ESKINt§*Department of Neurobiology and Anatomy, University of Texas Medical School, Houston, TX 77225; and tDepartment of Biochemical and BiophysicalSciences, University of Houston, Houston, TX 77204
Communicated by Eric R. Kandel, January 10, 1994
ABSTRACT A form of associative plasticity in Aplysia,activity-dependent neuromodulation, involves the convergenceofneuronal activity and the effects ofa modulatory tansmitter.To investigate the role of protein synthesis in assoiativeplasticity, we examined the effects of a biochemical anlgue ofactivity-dependent neuromodulation on the level of incorpo-ration oflabeled amino acid into proteins. To mimic assoiativetraining, abdominal ganglia were exposed to paired treatmentsof a depolarizing agent, elevated potassium, and a modulatorytransmitter, serotonin. The effects of elevated potassium andserotonin applied alone were also exmined. At least twoproteins (nos. 9 and 17) were affected in a nonadditive way bythe paired procedure. Incorporation of label into protein 9 wasincreased by the paired procedure but was not affected byeither elevated potassium or serotonin. Incorporation of labelinto protein 17 was sificantly affected by elevated potassiumor serotonin, but the effect of the paired procedure wassignfeantly less than the summed effects ofelevated potassiumand serotonin applied alone. These results indicate that changesin protein synthesis may be important in the induction ofassociative plasticities. Amino acid sequences of two peptidesderived from protein 9 were obtained. Then, a partial cDNAclone for protein 9 was obtained by performing PCR withdegenerate primers corresponding to portions of the sequencesof the two jeptides. The sequence of protein 9 is related tosequences prewiously reported for a family of genes comprisingthe strinnt starvation protein of Escherichia coli, auxin-induced proteins of plants, and glutathione S-transferases of anumber of organisms.
A cellular analogue of classical conditioning produces anenhancement of the connections between sensory neuronsand motor neurons in Aplysia (1, 2). Sensory-motor synaptictransmission involved in the tail and siphon withdrawalreflexes of Aplysia is enhanced by pairing spike activity in asensory neuron (conditioned stimulus) with electrical stimu-lation of the tail or peripheral nerves (unconditioned stimu-lus). This type of associative plasticity, called activity-dependent neuromodulation, has been observed in a short-term form lasting minutes (1, 2) and a long-term form lasting24 hr (3). This short-term associative plasticity in Aplysia isbelieved to involve the convergent action of two secondmessengers, Ca2+ and cAMP (4-8). Convergent actions oftwo second-messenger pathways may also produce the long-term form of the associative plasticity by modifying tran-scription and protein synthesis. Although the importance oftranscription and protein synthesis in some forms of nonas-sociative plasticity have been well established, their impor-tance in associative plasticity is just beginning to be deter-mined (ref. 49; for review, see refs. 8-14).
In the present study we examined whether an analogue ofactivity-dependent neuromodulation induced changes in pro-tein synthesis in cells of Aplysia abdominal ganglia. Theanalogue consisted of exposure of abdominal ganglia topaired treatments of (i) high K+ and (ii) serotonin (5-hydroxytryptamine, 5-HT). Depolarization by high K+ wasused to mimic spike activity. 5-HT, which mediates manyeffects of sensitizing stimuli (ref. 15; for review, see ref. 16),was used to mimic the effects of sensory stimulation. Theeffects of the paired procedure were compared with theeffects of high K+ and 5-HT applied individually. We foundthat pairing high K+ with 5-HT produced effects on proteinsthat were different from a summation of the effects producedby each stimulus applied alone. Furthermore, we obtained apartial amino acid sequence and then a partial cDNA clone ofone of the proteins affected by the paired treatment.
MATERIALS AND METHODSAplysia californica were obtained from Alacrity MarineBiological (Redondo Beach, CA), Marinus (Long Beach,CA), and Sea Life Supply (Sand City, CA) and maintained inartificial seawater (Instant Ocean, Aquarium Systems, Men-tor, OH) at 15'C. Two days or more after their arrival,animals were anesthetized by injection of an amount ofisotonic MgCl2 equivalent to approximately halfoftheir bodyweight. Abdominal ganglia were removed and placed intofour separate chambers (three ganglia per chamber) contain-ing buffered filtered seawater [BFSW: artificial seawatercontaining 30 mM Hepes, streptomycin sulfate (BioWhit-taker) (100 ,ug/ml), and penicillin G (100 units/ml) at pH7.65]. The ganglia remained in BFSW at 15'C for at least 2 hrbefore exposure to experimental treatments. Each group ofganglia was exposed to one of the following treatments: (i)high K+ (80mM KCl) applied alone for 1 hr; (ii) 5-HT (5 AuM)applied alone for 2 hr; (iii) high K+ for 1 hr paired with 5-HTfor 2 hr, with the two treatments overlapping during the last0.5 hr ofthe high K+ treatment and the first 0.5 hr ofthe 5-HTtreatment; or (iv) no treatment. Elevation of the K+ concen-tration in the medium is a well-established method for pro-ducing membrane depolarization of molluscan neurons (6,17-21). Two hours of training or 5 uM 5-HT are commonlyused to elicit long-term plasticity in Aplysia (22-26). The fourchambers were thoroughly washed with BFSW at the end ofeach treatment. The ganglia were metabolically labeled with[3H]leucine (0.1 mCi/ml; 1 mCi = 37 MBq) for 2 hr starting5 hr after the end of 5-HT treatments or at times of other
Abbreviations: 5-HT, 5-hydroxytryptamine (serotonin); SSP, strin-gent-starvation protein; GST, glutathione S-transferase; AIP, auxin-induced protein.tPresent address: Department of Pediatrics, Baylor College of Med-icine, Houston, TX 77030.§To whom reprint requests should be addressed.
4150
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.

Proc. Natl. Acad. Sci. USA 91 (1994) 4151
treatments corresponding to the projected end of the 5-HTtreatment (see Fig. 1).The ganglia were homogenized on ice and their proteins
were separated by two-dimensional PAGE as described byEskin et al. (27). In this procedure, minigels (Idea Scientific,Corvallis, OR) were used in the second dimension. Equalamounts of trichloroacetic acid-precipitable radioactivitywere loaded on the gels. To reduce variations, the fourexperimental and control samples of an experiment werealways run on the same batch of isoelectric focusing andSDS/polyacrylamide gels and then exposed to film at thesame time. Gels were exposed to films for different times toensure that fluorographs were not saturated for certain pro-teins.The results of each experiment were determined by com-
paring the treated group with an untreated control group runin the same experiment. The effects of treatments on incor-poration of amino acid into specific proteins were firstanalyzed visually. Densities of experimental proteins ontwo-dimensional gels werejudged as increased, decreased, ornot changed relative to the same protein from the control gel.Twenty-four proteins that appeared to be affected by at leastone of the experimental treatments were selected for furtheranalysis. As a second step, optical densities (ODs) of theselected proteins were determined by computerized imageanalysis (DNA Proscan, Nashville, TN). To normalize theODs for any discrepancies in the amount of radioactivityloaded on the gels, a percent OD was calculated for eachselected protein by dividing the OD of the selected protein bythe summed ODs of 5 proteins that were unaffected by thetreatment. An example of percent ODs ± SEM (n = 13) forprotein 9 is as follows: 7.73 ± 1.93% (5-HT alone), 8.97 +1.94% (high K+ alone), 8.27 ± 1.14% (control), and 12.48 +2.27% (paired 5-HT and high K+). To determine whetherproteins were significantly affected by one of the treatments,a Wilcoxon ranked sign test was performed between eachexperimental and control group (28). A Wilcoxon test wasalso used to determine whether the effect of the pairedprocedure was significantly different from the sum of theeffects of the individual treatments applied alone.To obtain amino acid sequences of proteins affected by
paired treatments, proteins ofinterest were first purified withpreparative two-dimensional gels. Protein purification, diges-tion with Staphylococcus aureus V8 protease, and peptidesequencing were carried out as described (29) with somemodifications (30). Protein 9 was first purified from abdom-inal ganglia. About 500 ,ug of protein from abdominal gangliawas loaded on each gel. After Coomassie brilliant blue R-250staining of the gel, protein 9 was cut out of 26 preparativetwo-dimensional gels. Protein 9 was digested in situ with S.aureus V8 protease. The peptides derived from protein 9were separated by SDS/15% PAGE and transferred onto apoly(vinylidene difluoride) membrane (Problott, AppliedBiosystems) and stained with Coomassie brilliant blue R-250.Protein 9 was purified a second time from abdominal andpleural-pedal tissue and digested in situ with cyanogenbromide (31). Peptide bands were cut from the Problottmembrane and sequenced with a 477A pulsed-liquid proteinsequencer (Applied Biosystems). Peptide sequence analysiswas performed at the National Center for BiotechnologyInformation with the BLAST network system (32).To obtain a partial cDNA clone for protein 9, poly(A)+
mRNA was first isolated from Aplysia abdominal and headganglia. Total RNA was obtained by homogenization ofganglia in a phenol/SDS mixture at 650C, followed by twoextractions with phenol/chloroform, 1:1 (vol/vol), and thenprecipitation with 70o ethanol in 0.1 M NaCl. The ethanolprecipitate was reconstituted in water and precipitated with2 M sodium acetate (pH 6.0). Poly(A)+ RNA was isolatedfrom total RNA on an oligo(dT) column (33). cDNA was then
synthesized from 7 ,g ofRNA by using a cDNA synthesis kit(United States Biochemical) with random hexamers as prim-ers. Degenerate primers containing deoxyinosine (Genosys,The Woodlands, TX) and corresponding to regions ofthe twopeptides obtained from protein 9 were then designed andsynthesized (see Fig. 3).PCRs were performed (Perkin-Elmer/Cetus kit) on 40 ng
of Aplysia ganglion cDNA with 10 ,pM each primer, 10 01 of1Ox reaction buffer, 200 ,uM dNTPs, and 2.5 units of Am-pliTaq enzyme in 100 sul. Reaction conditions were optimizedas follows: initial (first cycle only) denaturation at 940C for 2min, subsequent denaturations at 920C for 1 min, annealing at420C for 1 min, extension at 720C for 1 min. Thirty cycles wererun with the PTC-100 programmable thermal controller (MJResearch, Waltertown, MA).The PCR fragment of interest was cloned into a vector with
the TA cloning system (Invitrogen). Plasmid preparationswere performed on 10 colonies with Magic Minipreps(Promega). Insert-containing clones were identified by cleav-age with EcoRI restriction endonuclease (1 unit/100 ng ofDNA). The sequence of the 180-bp insert was verified bydideoxy sequencing (United States Biochemical Sequenasekit); the sequencing reaction products were run on a Sequi-Gen apparatus (Bio-Rad).
RESULTSIn each experiment, groups of three abdominal ganglia wereexposed to high K+, 5-HT, paired application of high K+ and5-HT, or no treatment (Fig. 1). Thirteen independent exper-iments with 12 animals per experiment were performed.Visual analysis of the gels from these experiments yielded 24proteins that appeared to be affected by at least one of theexperimental treatments. This group of 24 proteins wasexamined statistically for proteins affected by one of thetreatments. One protein, no. 9, was significantly increased bythe paired procedure but not by high K+ or 5-HT appliedalone (Wilcoxon test). To determine whether the results ofthe paired application of high K+ and 5-HT could be ex-plained by addition ofthe effects ofhigh K+ and 5-HT appliedalone, the effects on protein synthesis produced by pairedhigh K+ and 5-HT were compared statistically with a sum-mation of the changes produced by high K+ and 5-HT appliedalone (Wilcoxon test). For protein 9, the effect of pairedapplication of high K+ and 5-HT was significantly greaterthan the summed results of high K+ and 5-HT applied alone(Fig. 2A). We did observe some cases-e.g., protein 5-inwhich the effect ofthe paired application ofhigh K+ and 5-HTdid not differ significantly from the summed results of highK+ and 5-HT applied alone (Fig. 2B).Another interesting nonadditive type of effect of the paired
treatment was observed. For example, protein 17 was signif-icantly affected by high K+ and 5-HT applied alone. However,the effects of paired high K+ and 5-HT were significantly lessthan the summed effects of high K+ and 5-HT applied alone(Fig. 2C).More precise study of the role of these proteins in memory
formation requires identification of the proteins and deter-mination of their cellular function. To do this, we initiatedexperiments to obtain a partial amino acid sequence ofprotein 9. Protein 9 was purified from abdominal and pleural-pedal ganglia by preparative two-dimensional PAGE. Spotsof protein 9 were removed from the gels and digested withcyanogen bromide or with S. aureus V8 protease. Thepeptides were then separated in one-dimensional SDS/polyacrylamide gels, blotted onto poly(vinylidene difluoride)membranes, and stained with Coomassie brilliant blue. Asequence of 36 aa was obtained by cyanogen bromide diges-tion and another non-overlapping sequence of 37 aa wasobtained by V8 protease digestion (Fig. 3A).
Neurobiology: Noel et al.

Proc. Nati. Acad. Sci. USA 91 (1994)
!D14.6 5.1
AHigh K+ + 5-HT
High K.
5-HT
1 hr
3H-leucine
I.I
2 hr 5 hr 2 hr
B5-HT
5-HT 3H-leucine
0.5 hr 2 hr 5 hr 2 hr
CHigh Ki
High K" 3H-leucine
1 hr 6.5 hr 2 hr
Control
3H-Ieucine-h9,
7.5 hr 2 hr
-. IL *
0. IO
S.04. *
t~~~~~A_ A_...k
A&
-~~~~~~~At
1.,p0w
:f *a t.
5.8
98
65
43 kDa
31
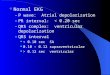
FIG. 1. Proteins affected by thepaired application of high K+ and 5-HT.Fluorograms ofexperimental and controltwo-dimensional gels are shown. Abdom-inal ganglia were exposed either to pairedtreatments of high Ki (80 mM) for 1 hrand 5-HT (5 AM) for 2 hr so that thetreatments overlapped over a 30-min pe-riod (A), to 5-HT alone (B), or to high K+alone (C). Control ganglia received notreatment (D). After 5 hr, the gangliawere labeled with [3H]leucine for 2 hr.Proteins 9 and 17 were affected in anonadditive fashion by the paired appli-cation of high K+ and 5-HT (see Fig. 2Aand C). The effect of the paired applica-tion ofhigh K+ and 5-HT on protein S didnot differ from the summed effects ofhigh K+ and 5-HT applied alone (see Fig.2B).
To align the two peptides from protein 9 and obtainadditional sequence, degenerate primers with deoxyinosinesat the points of highest degeneracy were synthesized corre-sponding to the two peptides and used to amplify cDNAsynthesized from poly(A)+ mRNA from Aplysia ganglia. Aband of %180 bases was obtained and purified from a poly-acrylamide gel. After cloning and sequencing, this band wasshown to contain sequences that corresponded to the tar-geted regions of the two peptides. In addition, a sequence of19 aanbetween the two peptides was obtained. Therefore, acombined sequence of92 aa was obtained from protein 9 (Fig.3A).The BLAST program (32) was used to compare the 92-aa
Aplysia peptide to sequences of known proteins (search wasperformed on November 20, 1993). This long peptide wasfound to have 34% identity over an 88-aa region to the SSPofE. coli (34), 43% identity over a 59-aa region to a GST fromD. caryophyllus (35), and 40%o identity over a 46-aa region toan AIP from N. tabacum (36) (Fig. 3A). The similaritiesbetween protein 9 and SSP, GST, and AIP are highly signif-icant because the probabilities to obtain such matches bychance are extremely low (Fig. 3B). In addition, the SSPs andGSTs are related (37). An AIP from Nicotiana plumbagini-folia that is very similar to the AIP mentioned above also hassignificant similarity to the SSP and GST families (38).Finally, the sequence ofour peptide from protein 9 was found
to have significant similarity to a cosmid encoding a Cae-norhabditis elegans protein of unidentified function (39).
DISCUSSIONOur finding that paired stimulation produced nonadditiveeffects on the incorporation of label into proteins suggeststhat translation, and perhaps transcription as well, may beinvolved in generating some forms of associative plasticity.Similar conclusions emerged from experiments on PC12 ratpheochromocytoma cells, a neuronal cell line in which cAMPand Ca2+ appear to interact in a nonadditive or synergisticfashion in the regulation of transcription (40, 41). Nonaddi-tive effects of stimuli imply convergence ofthe inputs at somelevel of the signaling pathway. In our case, the inputs mustconverge upon some molecular event of the informationprocessing pathways. The identification of molecular eventsthat may act as coincident detectors in neurons is an impor-tant issue (e.g., refs. 7, 42, and 43), and several such detectors(e.g., adenylate cyclases and cAMP response element-binding proteins; refs. 4, 41, and 44) have been under study.The identification of proteins 9 and 17, whose synthesisappears to be governed by such coincident detectors, shouldallow us to work backwards to identify the mechanismsresponsible for coincident detection in Aplysia.A critical issue raised by our results is the role of these
specific proteins in generating behavioral plasticity in Aply-
4152 Neurobiology: Noel et aL

Proc. Natl. Acad. Sci. USA 91 (1994)
B(f)l
-Il
protein']11 3
C >3-H1'
C]-. 11i',1 K' a i" 1-HLliiirllK 1 '- .-{ I
*0 IK_] 7'i;|
iL! t**
[IIN()lti 1lFIG. 2. Changes in incorporation ofamino acid into proteins produced by paired high K+ and 5-HT compared with a summation of the effects
of high K+ and 5-HT applied alone (high K+ plus 5-HT). Percent differences of integrated ODs of experimental proteins relative to controlproteins are shown. (A) Incorporation into protein 9 was increased by paired high K+ and 5-HT but did not appear to be affected by either highK+ or 5-HT applied alone. The effect of paired high K+ and 5-HT was significantly different from the summed effects of high K+ and 5-HTapplied alone. (B) Incorporation into protein 5 was decreased by paired high K+ and 5-HT or by high K+ or 5-HT applied alone. The effect ofthe paired application of high K+ and 5-HT did not differ significantly from the summed results of high K+ and 5-HT applied alone. (C)Incorporation into protein 17 was decreased by high K+ or 5-HT applied alone but was not affected by the paired treatment. Paired high K+and 5-HT produced a significantly different effect from the summed effects of high K+ and 5-HT applied alone. Level of significant differencebetween the paired high K+ and 5-HT results and the summed results of high K+ and 5-HT applied alone: ns, nonsignificant; *, P < 0.05; **,P < 0.01.
sia. It will be important to determine whether the types ofpairing specific changes we observed depend on a particulartemporal relationship of the two stimuli. Also, it will beimportant to investigate whether the types of changes weobserved occur in neurons known to be responsible forspecific types of learning. Protein 9 has not been identified inprevious studies examining learning-related changes in pro-teins in Aplysia (13, 22, 25, 27, 29, 31, 45, 46). This is not
surprising, however, since those studies examined effects ofanalogues of nonassociative learning on proteins. Finally, thecellular functions of proteins 9 and 17 need to be determined.The 92-aa sequence obtained from protein 9 by microse-
quencing and PCR represents about one-third of the totalprotein, since protein 9 is about 31 kDa. This derived aminoacid sequence had significant similarity to the SSP from E.coli, GSTs from a number of organisms, and an AIP from a
Aplysia Peptide
SSP
AIP
RTCPYQRA.II IIISDLVNNDLNLNPSLIAAEYLEEAPbPPLHAK
21 YSHQVnJVL F IEH ELITLI YEP RLPP VPV AIkrERLYWHRIEI l(S
XFL LNP PVLVGDDSL XIMYLE=PENPLLPQlLQKRA 98
Y)YKKVPVUNGXVESM I LEYIDGPTLPDPYD FW 9
3
Identitvto .-ApvsEia peptide
-P lue
551'WSIL U,li.
G SI1!.A. CI ')L:J:I !'.&.
.lt(4 {
l .\Il () 2.()x\ ()
FIG. 3. Comparison of amino acid sequences derived from Aplysia protein 9, stringent starvation protein (SSP) of Escherichia coli (34),glutathione S-transferase (GST) from Dianthus caryophyllus (35), and auxin-induced protein (AIP) from Nicotiana tabacum (36). (A) Theunderlined amino acids in the Aplysia peptide sequence are the amino acids obtained by PCR. To the left of this underlined region is the aminoacid sequence obtained by cyanogen bromide digestion, while the one to the right of the underlined region is the sequence obtained by S. aureusV8 protease digestion. The regions in each peptide to which corresponding degenerate primers were synthesized for PCR are indicated by arrows.The two primers were as follows: 5'-AA(R-) AA(R-) CCI-GA(Y-) CA(Y-) TT(Y-) TT(Y-) GA-3' (a) and 5'-TG(R-) TT(R-) AA(R-)TAI-ATI-C(KY-)TC (b), where I is deoxyinosine. To show the similarity among the four peptides, the amino acids which are identical in at leasttwo of the sequences are included in shaded boxes. (B) The percent identities of the Aplysia peptide to each of the other three peptides, andthe P values for those sequence identities, were obtained with the BLAST program (32). P values are probabilities that matches as good or betterthan those found would occur by chance.
A
Siz
-j
J
Z-
A
Neurobiology: Noel et A 4153
.tl
It
.1. !i
*.-.
(,.I
protetill Q

Proc. Natl. Acad. Sci. USA 91 (1994)
plant. The E. coli SSP may interact with RNA polymerase toregulate its function (47). An AIP with similarity to the E. coliSSP has been found in N. plumbaginifolia (38). This plantprotein may also have a regulatory effect on transcription. Itwas proposed that the family of genes coding for this familyof plant proteins be called multiple-stimulus response genes(38). GSTs comprise a large family of proteins which areinvolved in cell protective mechanisms (for review, see ref.48). Although protein 9 had significant similarity to an SSP,an AIP, and a GST, the function of protein 9 in Aplysiaremains to be determined. Our results suggest that protein 9may have a specific regulatory role in events associated withcoincident input signals. Further study ofprotein 9 may allowus to determine how paired treatments alter protein synthesisand how protein synthesis is involved in the induction ofassociative forms of memory.
We wish to thank Dr. R. Cook for the determination ofamino acidsequences and Z. Chen, C. Wang, B. Wells, and Haixu Chen forassisting with experiments and data analysis. We thank Dr. D.Davison for helping us with the analysis of sequence comparisons.We thank Dr. T. Kennedy for helping us develop the procedure forcyanogen bromide digestion, R. Homayouni for assisting in thedevelopment of PCR and cloning techniques, and R. Zwartjes forisolation of poly(A)+ mRNA. This work was supported by theFondation Fyssen (F.N.), National Institutes of Health GrantNS28462 and Armed Forces Office of Scientific Research GrantF49620-92-J-0494 (A.E.), and National Institutes of Health GrantNS19895 and National Institute of Mental Health Award MH00649(J.H.B.).
1. Hawkins, R. D., Abrams, T. W., Carew, T. J. & Kandel, E. R.(1983) Science 219, 400-405.
2. Walters, E. T. & Byrne, J. H. (1983) Science 219, 405-408.3. Buonomano, D. V. & Byrne, J. H. (1990) Science 249, 420-
423.4. Abrams, T. W., Karl, K. A. & Kandel, E. R. (1991) J. Neu-
rosci. 11, 2655-2665.5. Kandel, E. R., Abrams, T. W., Bernier, L., Carew, T. J.,
Hawkins, R. D. & Schwartz, J. H. (1983) Cold Spring HarborSymp. Quant. Biol. 48, 821-830.
6. Ocorr, K. A., Walters, E. T. & Byrne, J. H. (1985) Proc. Nati.Acad. Sci. USA 82, 2548-2552.
7. Abrams, T. W. & Kandel, E. R. (1988) Trends Neurosci. 11,128-136.
8. Byrne, J. H., Zwartjes, R., Homayouni, R., Critz, R. D. &Eskin, A. (1993) Adv. Second Messenger Phosphoprotein Res.27, 47-108.
9. Abraham, W. C., Dragunow, M. & Tate, W. P. (1992) Mol.Neurobiol. 5, 297-314.
10. Bliss, T. V. P. & Collingridge, G. L. (1993) Nature (London)361, 31-39.
11. Davis, H. P. & Squire, L. R. (1984) Physiol. Bull. 96, 518-559.12. Hawkins, R. D., Kandel, E. R. & Siegelbaum, S. A. (1993)
Ann. Rev. Neurosci. 16, 625-665.13. Kennedy, T. E., Hawkins, R. D. & Kandel, E. R. (1992) in
Neuropsychology ofMemory, eds. Squire, L. R. & Butters, N.(Guilford Press, New York), pp. 557-574.
14. Rose, S. P. R. (1991) Trends Neurosci. 14, 390-397.15. Glanzman, D. L., Mackey, S. L., Hawkins, R. D., Dyke,
A. M., Lloyd, P. E. & Kandel, E. R. (1989) J. Neurosci. 19,4200-4212.
16. Cleary, L. J., Baxter, D. A., Nazif, F. & Byrne, J. H. (1991)Biol. Bull. Woods Hole Mass. 180, 252-261.
17. Carpenter, D. 0. & Alving, B. 0. (1968) J. Gen. Physiol. 52,1-21.
18. Curtis, H. J. & Cole, K. S. (1942) J. Cell Comp. Physiol. 19,135-144.
19. Gorman, A. L. F. & Marmor, M. F. (1970) J. Physiol. (Lon-don) 210, 897-917.
20. Hodgkin, A. L. & Keynes, R. D. (1955) J. Physiol. (London)128, 61-88.
21. Kerkut, G. A. & Meech, R. W. (1967) Comp. Biochem. Phys-iol. 20, 411-429.
22. Barzilai, A., Kennedy, T. E., Sweatt, J. D. & Kandel, E. R.(1989) Neuron 2, 1577-1586.
23. Glanzman, D. L. & Kandel, E. R. (1990) Science 249,799-802.24. Montarolo, P. G., Goelet, P., Castellucci, V. F., Morgan, J.,
Kandel, E. R. & Schacher, S. (1986) Science 234, 1249-1254.25. Noel, F., Eskin, A. & Byrne, J. H. (1991) Brain Res. 568,
67-75.26. Sweatt, J. D. & Kandel, E. R. (1989) Nature (London) 339,
51-54.27. Eskin, A., Garcia, K. S. & Byrne, J. H. (1989) Proc. Natl.
Acad. Sci. USA 86, 2458-2462.28. Zar, J. H. (1984) Biostatistical Analysis (Prentice-Hall, Engle-
wood Cliffs, NJ), pp. 153-156.29. Kennedy, T. E., Gawinowicz, M. A., Barzilai, A., Kandel,
E. R. & Sweatt, J. D. (1988) Proc. Natl. Acad. Sci. USA 85,7008-7012.
30. Raju, U., Nunez-Regueiro, M., Cook, R., Kaetzel, M. A.,Yeung, S. J. & Eskin, A. (1994) J. Neurochem. 61, 1236-1245.
31. Kennedy, T. E., Kuhl, D., Barzilai, A., Sweatt, J. D. &Kandel, E. R. (1992) Neuron 9, 1013-1024.
32. Altschul, S. F., Gish, W., Miller, W., Myers, E. W. & Lipman,D. J. (1990) J. Mol. Biol. 215, 403-410.
33. Sambrook, J., Fritsch, E. F. & Maniatis, T. (1989) MolecularCloning: A Laboratory Manual (Cold Spring Harbor Lab.Press, Plainview, NY), 2nd Ed., pp. 7.26-7.29.
34. Serizawa, H. & Fukuda, R. (1987) Nucleic Acids Res. 15,1153-1163.
35. Meyer, R. C., Jr., Goldsbrough, P. B. & Woodson, W. R.(1991) Plant Mol. Biol. 17, 277-281.
36. Van Der Zaal, E. J., Droog, F. N. J., Boot, C. J. M., Hen-skens, L. A. M., Hoge, J. H. C., Schilperoort, R. A. &Libbenga, K. R. (1991) Plant Mol. Biol. 16, 983-988.
37. Toung, Y. & Tu, C. D. (1992) Biochem. Biophys. Res. Com-mun. 182, 355-360.
38. Dominov, J. A., Stenzler, L., Lee, S., Schwartz, J. J., Leisner,S. & Howell, S. H. (1992) Plant Cell 4, 451-461.
39. Sulston, J., Du, Z., Thomas, K., Wilson, R., Hillier, L.,Staden, R., Halloran, N., Green, P., Thierry-Mieg, J., Qiu, L.,Dear, S., Coulson, A., Craxton, M., Durbin, R., Berks, M.,Metzstein, M., Hawkins, T., Ainscough, R. & Waterston, R.(1992) Nature (London) 356, 37-41.
40. Ginty, D. D., Bading, H. & Greenberg, M. E. (1992) Curr.Opin. Neurobiol. 2, 312-316.
41. Sheng, M., McFadden, G. & Greenberg, M. E. (1990) Neuron4, 571-582.
42. Byrne, J. H. (1987) Physiol. Rev. 67, 329-439.43. Bourne, H. R. & Nicoll, R. (1993) Neuron 10, 65-75.44. Andrade, R. (1993) Neuron 10, 83-88.45. Noel, F., Nunez-Regueiro, M., Cook, R., Byrne, J. H. &
Eskin, A. (1993) Mol. Brain Res. 19, 203-210.46. Castellucci, V. F., Kennedy, T. E., Kandel, E. R. & Goelet, P.
(1988) Neuron 1, 321-328.47. Ishihama, A. & Saitoh, T. (1979) J. Mol. Biol. 129, 517-530.48. Pickett, C. B. & Lu, A. Y. H. (1979) Annu. Rev. Biochem. 58,
743-764.49. Crow, T. & Forrester, J. (1990) Proc. Natl. Acad. Sci. USA 87,
4490-4494.
4154 Neurobiology: Noel et al.