Embed Size (px)
Citation preview

Institut für Tierernährung und Ernährungsphysiologie
(Direktor: Prof. Dr. K. Eder)
Einfluss oxidierter Fette auf zelluläre Signalwege im Modelltier
Dissertation zur Erlangung des akademischen Grades des
Doktors der Ökotrophologie (Dr. oec. troph.)
eingereicht im
Fachbereich Agrarwissenschaften, Ökotrophologie und Umweltmanagement
Justus-Liebig-Universität Gießen
(Dekan: Prof. Dr. Peter Kämpfer)
vorgelegt von
Diplom-Ernährungswissenschaftlerin
Juliane Varady
Gießen, 2013
1. Gutachter: Prof. Dr. K. Eder
2. Gutachter: Prof. Dr. U. Wenzel

I
Inhaltsverzeichnis
II. Abkürzungsverzeichnis
1. Einleitung ..................................................................................................................... 1
2. Zielstellung ................................................................................................................. 6
3. Originalarbeiten ....................................................................................................... 10
3.1 Studie 1: Dietary oxidized fat activates the oxidative stress-responsive
transcription factors NF-κB and Nrf2 in intestinal mucosa of mice…..…….……….11
3.2 Studie 2: Dietary moderately oxidized oil activates the Nrf2 signaling
pathway in the liver of pigs……………………………………………………………………………………….23
3.3 Studie 3: Dietary moderately oxidized oil induces expression of
fibroblast growth factor 21 in the liver of pigs……………………………………………………….32
4. Diskussion ................................................................................................................ 38
4.1 Einfluss oxidierter Fette auf den Nrf2........................................................................... 38
4.2 Einfluss oxidierter Fette auf den NF-κB ...................................................................... 42
4.3 Einfluss eines oxidierten Fettes auf den FGF21 ........................................................ 45
4.4 Schlussfolgerungen ............................................................................................................ 49
5. Zusammenfassung .................................................................................................. 53
6. Summary ................................................................................................................... 56
7. Literaturverzeichnis ............................................................................................... 58
A Eidesstattliche Erklärung
B Danksagung

II
Abkürzungsverzeichnis
Neben den Abkürzungen der Deutschen Rechtschreibung laut Duden wurden folgende Abkürzungen
verwendet:
ACO Acyl-CoA Oxidase
ARE antioxidant response element bZIP basischer Leucin-Zipper
CED chronisch-entzündliche Darmerkrankung
CFAM Cyclic fatty acid monomers; zyklische Fettsäuremonomere COX cytochrome c oxidase CPT Carnitin-Palmitoyltransferase
CRB CREB binding protein DGF Deutsche Gesellschaft für Fettwissenschaft e.V.
DNA desoxyribonucleic acid; Desoxyribonukleinsäure FGF Fibroblast Growth Factor GLUT Glukosetransporter
GPX Glutathionperoxidase
HDL high density lipoprotein HMGCoAS2 mitochondriale 3-Hydroxy-3-Methylglutaryl-CoA Synthase
HNE Hydroxynonenal
HO Hämoxygenase
HODE Hydroxy octadecadienoic acid; Hydroxy-octadecadiensäure HPODE Hydroperoxy octadecadienoic acid; Hydroperoxy-octadecadiensäure IKK IκB-Kinase-Komplex IL Interleukin
iNOS inducible nitric oxide synthase Keap1 Kelch ECH Associating Protein 1 LDL low density lipoprotein mRNA messenger ribonucleic acid NEMO NF-κB essential modulator NF-κB Nuclear factor 'kappa-light-chain-enhancer' of activated B-cells NQO NAD(P)H-Quinon-Oxidoreduktase
Nrf2 Nuclear factor-erythroid 2-related factor 2 OCTN Novel organic cation transporter PPAR peroxisome proliferator-activated receptor PPRE peroxisome proliferator response element ROS Reactive oxygen species, reaktive Sauerstoffspezies RXR retinoid X receptor; 9-cis-Retinsäurerezeptor SCD Stearoyl-CoA Desaturase
SOD Superoxiddismutase
T3 Trijodothyronin
T4 Thyroxin
TNF Tumor necrosis factor TXNR Thioredoxinreduktase
UGT UDP-Glukuronosyltransferase
VLDL very low density lipoproteins

Einleitung
1
1. Einleitung
Oxidierte Fette sind chemisch veränderte Fette, wie sie in erhitzten und unsachgemäß gelagerten
Lebensmitteln zu finden sind. Insbesondere während des Frittierens bilden sich große Mengen an
Oxidationsprodukten im Frittieröl. Das Frittieren von Lebensmitteln ist eine weltweit verbreitete
und sehr populäre Art der Zubereitung von Lebensmitteln. Kennzeichnend für frittierte Lebensmittel
sind ein typisches Frittieraroma, eine goldbraune Färbung und knusprige Oberfläche. Beim Frittieren
werden Lebensmittel für kurze Zeit in 160 °C bis 190 °C heißes Öl vollständig eingetaucht und somit
gegart. Während des Frittiervorgangs bildet sich im Öl ein Mix aus verschiedensten Substanzen, die
teilweise vom Frittiergut absorbiert und somit beim Verzehr aufgenommen werden. So liegt der
Fettgehalt nach dem Frittieren in Kartoffelchips durchschnittlich zwischen 33-38 % und in Pommes
frites zwischen 10-15 % (zit. nach Choe und Min, 2007). Die während dieses Prozesses ablaufenden
Reaktionen bestimmen maßgeblich die physikalischen, chemischen und nutritiven Eigenschaften
sowohl des Frittierguts als auch des Frittieröls. Die bedeutendsten dabei ablaufenden Prozesse sind
die Hydrolyse, die Oxidation und die Polymerisation. Sie sind u. a. abhängig von der Art und
Zusammensetzung des Frittieröls, der Frittiertemperatur und -dauer, dem Frittiergut, dem Gehalt an
Antioxidantien und der Art der Fritteuse. Durch die Hydrolyse von Triglyceriden steigt der Gehalt an
freien Fettsäuren, Mono- und Diglyceriden und Glycerin im Öl an (Choe und Min, 2007). Die
Oxidation der Fettsäuren ist eine radikalische Kettenreaktion, bei welcher primäre und sekundäre
Lipidperoxidationsprodukte gebildet werden. Bedeutende Oxidationsprodukte der Linolsäure sind
die Hydroxylderivate 9- und 13-Hydroxy-octadecadiensäure (9-HODE und 13-HODE) und 13-
Hydroperoxy-octadecadiensäure (13-HPODE). Aus der Linolsäure und α-Linolensäure können sich
bei Temperaturen über 200°C auch zyklische Fettsäuremonomere (CFAM) bilden. Außerdem
entstehen eine Reihe flüchtiger Substanzen, wie Aldehyde, Ketone, Carbonsäuren und kurzkettige
Alkane und Alkene.
Aus Untersuchungen im Menschen und in verschiedenen Tiermodellen, wie Maus, Ratte und
Schwein, ist bekannt, dass die Aufnahme oxidierter Fette verschiedenste biologische Effekte im
Organismus hervorrufen kann (Cohn, 2002; Ringseis und Eder, 2011; Staprans et al., 2005).
Grundsätzlich wird die Aufnahme oxidierter Fette als ungünstig für den Organismus angesehen, da
sie mit einer Beeinträchtigung der Glukosetoleranz, Beeinflussung der Schilddrüsenfunktion und
proatherogenen Wirkungen in Verbindung gebracht werden (Chao et al., 2007; Eder und Stangl,
2000; Skufca et al., 2003; Staprans et al., 2005). Es können jedoch auch günstige Wirkungen im
Stoffwechsel vermittelt werden. Dazu zählt die Reduktion der Triglycerid- und
Cholesterinkonzentrationen im Plasma, in very low density lipopoteins (VLDL) und in der Leber (Eder
und Kirchgessner, 1998; Huang et al., 1998; Sülzle et al., 2004). Erhöhte Plasmalipide sind ein
bekannter Risikofaktor in der Pathogenese von Herz-Kreislauf-Erkrankungen, wie der
Arteriosklerose. Weiterhin gibt es Hinweise darauf, dass oxidierte Fette auch antiadipogene und

Einleitung
2
antiatherogene Wirkungen hervorrufen und der Entwicklung einer Alkohol-induzierten Fettleber
vorbeugen können (Chao et al., 2007; Kämmerer et al., 2011; Ringseis et al., 2007a).
Eine der wichtigsten metabolischen Wirkungen oxidierter Fette ist jedoch die Induktion von
oxidativem Stress. Kennzeichnend für oxidativen Stress ist die intrazelluläre Akkumulation von
reaktiven Sauerstoffspezies (ROS) [freie Radikale (wie Hydrogenperoxide, Superoxide,
Hydroxylradikale) und reaktive Metabolite], aufgrund einer verstärkten Bildung von ROS durch
prooxidative Prozesse und/oder einer verminderten Inaktivierung von ROS durch antioxidative
Prozesse, wodurch Schäden an Biomolekülen verursacht oder Redoxsignalwege gestört werden
(Bergamini et al., 2004; Halliwell, 2007). Die Ursachen für die Induktion von oxidativem Stress durch
oxidierte Fette liegen einerseits in der Absorption von Lipidperoxidationsprodukten, wie
Hydroperoxiden, aus dem oxidierten Fettgemisch, welche im weiteren Verlauf die Autoxidation
endogener ungesättigter Fettsäuren verursachen können, und andererseits in der Generierung von
ROS über Cytochrom-P450-Enzyme, welche durch oxidierte Fette induziert werden können (Chao et
al., 2001). Oxidativer Stress äußert sich in Tieren, welchen oxidierte Fette verabreicht wurden, durch
erhöhte Konzentrationen an Lipidperoxidationsprodukten, reduzierte Konzentrationen an exogenen
(z.B. Tocopherole, Ascorbinsäure) und endogenen (z.B. Glutathion) Antioxidantien und einem
verminderten Verhältnis von reduziertem zu oxidiertem Glutathion im Plasma und in Geweben
(Brandsch et al., 2004; Liu und Huang, 1996; Liu und Huang, 1995; Srinivasan und Pugalendi, 2000).
ROS und andere reaktive Metabolite werden im aerob lebenden Organismus, z.B. im Rahmen der
Atmungskette, permanent gebildet und können in aller Regel durch zelluläre antioxidative
Abwehrmechanismen effektiv eliminiert werden (Michiels et al., 1994; Turrens, 2003). Erst wenn die
gebildeten ROS die Kapazität dieser Mechanismen übersteigen, kommt es zur Entstehung von
oxidativem Stress und Störungen im zellulären Redoxstatus. In diesem Fall können ROS als wichtige
Signalmoleküle zur Aktivierung redox-sensitiver Transkriptionsfaktoren fungieren, welche die
Expression von Genen steuern, die in die zelluläre Abwehr, in inflammatorische Prozesse und bei der
Adaptation auf oxidativen Stress involviert sind (Sun und Oberley, 1996; Surh et al., 2005). Zwei sehr
bedeutende Transkriptionsfaktoren, welche durch Veränderungen im zellulären Redoxstatus
aktiviert werden, sind der Nrf2 (Nuclear factor-erythroid 2-related factor 2) und der NF-κB (Nuclear
factor 'kappa-light-chain-enhancer' of activated B-cells) (Jaiswal, 2004; Michiels et al., 2002).
Dem Nrf2, welcher zusammen mit Nrf1, Nrf3, Bach1 und Bach2 zur Cap´N´Collar Familie der
basischen Leucin-Zipper-Transkriptionsfaktoren (bZIP) zählt, kommt eine zentrale Rolle im
zellulären Abwehrsystem zu, da er sowohl die basale als auch die induzierbare Expression einer
hohen Anzahl zytoprotektiver Gene kontrolliert (Itoh et al., 1997; Lee et al., 2004). Unter
physiologischen bzw. nicht-stimulierten Bedingungen liegt der Nrf2 im Zytosol der Zelle gebunden
an das Aktin-verankerte Inhibitorprotein Kelch ECH Associating Protein 1 (Keap1) vor, welches die
konstitutive Ubiquitinierung und proteasomale Degradierung des Nrf2 reguliert (Itoh et al., 1999;
McMahon et al., 2003). Als Reaktion auf elektrophilen oder oxidativen Stress kommt es zur

Einleitung
3
Modifizierung von Cysteinresten an Keap1 - den „Redoxsensoren“ - und somit zur
Konformationsänderung, woraufhin Nrf2 freigesetzt und seine nukleäre Translokation erhöht wird
(Itoh et al., 1999). Im Nukleus heterodimerisiert Nrf2 mit anderen bZIP-Proteinen, wie den small
MAFs, um an bestimmte regulatorische Regionen der DNA, den sogenannten antioxidant response
elements (ARE) zu binden (Rushmoore et al., 1991). Der Nrf2/MAF-Komplex rekrutiert zusammen mit
den Cofaktoren CREB binding protein (CRB) und p300 die Histonacetyltransferasen und RNA-
Polymerasen (Vo und Goodman, 2001; Zhu und Fahl, 2001). Dadurch wird die Transkription von
Nrf2-Zielgenen verstärkt, zu welchen u. a. Enzyme der Phase II des Fremdstoffmetabolismus,
antioxidative Enzyme, wie Enzyme des Glutathionmetabolismus, verschiedene Transportproteine
und proteasomale Proteine zählen (Lee et al., 2005). Aufgrund seiner großen Bedeutung in der
zellulären Abwehr wird der Nrf2 nahezu ubiquitär im Organismus exprimiert. Hohe
Expressionsraten finden sich in der Leber als zentralem Organ des Fremdstoffmetabolismus, aber
auch im Gastrointestinaltrakt als Eintrittspforte für Fremdstoffe (Moi et al., 1994; Petrick und
Klaassen, 2007).
Der nahezu ubiquitär exprimierte Transkriptionsfaktor NF-κB spielt eine bedeutende Rolle in der
induzierbaren Expression einer großen Anzahl von stressresponsiven Genen im Rahmen
verschiedener zellulärer Prozesse, wie der Immun- und Stressantwort und der Inflammation. Beim
Säuger umfasst der NF-κB fünf verschiedene Proteine, welche homo- bzw. heterodimerisieren können
und sich somit in ihrer DNA-Bindungsspezifität und ihrem transkriptionellen Aktivierungspotential
unterscheiden. Dazu zählen NF-κB1 (= p50) und NF-κB2 (= p52) mit ihren Präkursorproteinen p105
bzw. p100, sowie RelA (p65), c-Rel und RelB. Die dominante und klassische Konstellation stellt dabei
das Heterodimer aus p50 und p65 dar. In den meisten Zelltypen und unter nicht-stimulierten
Bedingungen liegt der NF-κB im Zytosol der Zelle gebunden an spezifische Inhibitorproteine (IκBα,
IκBβ, IκBε, IκBγ), welche zur IκB-Familie zählen, vor. Induziert wird die Aktivierung des NF-κB über
den IκB-Kinase-Komplex (IKK), welcher aus zwei katalytischen Untereinheiten (IKKα und IKKβ)
und dem regulatorischen Protein NF-κB essential modulator (NEMO) besteht. Nach Aktivierung durch
verschiedenste Stimuli, wie proinflammatorische Zytokine, virale und bakterielle Bestandteile,
physikalischen oder oxidativen Stress, werden die IκB-Proteine durch den IKK-Komplex an
spezifischen Serinresten phosphoryliert und somit der Ubiquitinierung und anschließenden
proteasomalen Degradierung zugeführt. Der so freigesetzte NF-κB kann daraufhin in den Zellkern
translozieren und durch Bindung an κB-responsive Elemente der DNA die Transkription seiner
Targetgene aktivieren (Barnes et al., 1997; Hayden und Ghosh, 2004; Luedde et al., 2007; Li und Stark,
2002).
Aus verschiedenen Untersuchungen ist bekannt, dass oxidierte Fette die Transkriptionsfaktoren
peroxisome proliferator-activated receptor (PPAR)α und PPARγ aktivieren und so einen Teil ihrer
metabolischen Wirkungen vermitteln können (Chao et al. 2001; Nagy et al., 1998; Schild et al., 2002;

Einleitung
4
Sülzle et al. 2004). Die potentiellen Liganden bzw. Aktivatoren dieser Transkriptionsfaktoren sind
wahrscheinlich Hydroxy- und Hydroperoxyfettsäuren (9-HODE, 13-HODE, 13-HPODE) und CFAM
(Delerive et al., 2000; König und Eder, 2006; Martin et al., 2000). Die PPARs, zu denen auch die
Isoform PPAR β/δ zählt, gehört zur Superfamilie der nukleären Hormonrezeptoren. Nach
Ligandenbindung und Heterodimerisierung mit dem 9-cis-Retinsäurerezeptor (RXR) binden sie im
Zusammenspiel mit Corepressoren und Coaktivatoren an spezifische DNA-Sequenzen, den peroxisome
proliferator response elements (PPREs) in der Promotorregion ihrer Zielgene. Die PPARs übernehmen im
Stoffwechsel zahlreiche wichtige Funktionen. So sind sie in die Regulation des Lipid- und
Lipoproteinstoffwechsels, der Glukosehomöostase, inflammatorischer Prozesse und der
Zellproliferation und -differenzierung involviert (Delerive et al., 2001; Gulick et al., 1994; Rosen et al.,
1999).
Der PPARα als zentraler Regulator des Energie- und Lipidstoffwechsels kontrolliert die Expression
von Genen, welche in die zelluläre Fettsäureaufnahme und den intrazellulären Fettsäuretransport, die
mitochondriale und peroxisomale Fettsäureoxidation und die Bildung von Ketonkörpern, den
Aminosäurestoffwechsel, die Glukoneogenese und die Lipogenese involviert sind (Mandard et al.,
2004). Hohe Expressionsraten sind vorwiegend in der Leber, aber auch im Herz- und Skelettmuskel,
in der Niere, im braunen Fettgewebe und in der Darmmukosa zu finden (Braissant et al., 1996).
PPARα und PPARγ fungieren als bedeutende Modulatoren bei inflammatorischen Reaktionen. Ihre
anti-inflammatorischen Wirkungen vermitteln sie über die Transrepression anderer
Transkriptionsfaktoren, wie dem NF-κB (Okayasu et al., 2008; Wang et al., 2002). Der PPARγ ist
außerdem in die Regulation der Adipogenese, Insulinsensitivität und Glukosehomöostase involviert
(Leonardini et al., 2009; Picard und Auxwerd, 2002; Tontonoz et al., 1994). Vom PPARγ existieren
zwei verschiedene Proteinisoformen, wobei hohe Expressionsraten an PPARγ2 im weißen und
braunen Fettgewebe zu finden sind, während PPARγ1 in den meisten Geweben in geringen Raten
exprimiert wird (Rosen und Spiegelman, 2001).
Kürzlich wurde der Fibroblast Growth Factor (FGF)21 als neuartiger hormonähnlicher Regulator im
Stoffwechsel identifiziert. FGF21 zählt zusammen mit FGF15/19 und FGF23 zur Unterfamilie 19 der
FGF-Superfamilie. Ihnen drei gemeinsam ist das Fehlen der heparin-bindenden Domäne, wodurch ein
schneller Übertritt vom Syntheseort in die Zirkulation gewährleistet ist (Kharitonenkov, 2009).
FGF21 vermittelt seine Effekte als endokriner Faktor über spezielle FGF-Rezeptoren an der
Oberfläche von Zellen vermutlich im Komplex mit dem Corezeptor β-Klotho (Kharitonenkov et al.,
2008; Ogawa et al., 2007). Hohe Expressionsraten an FGF21 sind in der Leber, im Fettgewebe und im
Pankreas zu finden, wo z. T. auch auto- bzw. parakrine Funktionen des FGF21 beschrieben sind
(Muise et al., 2008; Nishimura et al., 2000; Wente et al., 2006). Die Regulation seiner Expression
erfolgt über die nukleären Rezeptoren PPARα und PPARγ, aber auch über den AKT1-Signalweg
(Badman et al., 2007; Izumiya et al., 2008; Moyers et al., 2007). Seine physiologische Bedeutung liegt
in der Kontrolle der Lipid- und Glukosehomöostase, indem er die Glukoneogenese, die Ketogenese,

Einleitung
5
die Fettsäureoxidation und den Torpor - ein Starrezustand in kleinen Warmblütlern zur
Energieeinsparung - induziert und das somatische Wachstum inhibiert. Die hepatische Induktion des
FGF21 unterliegt der direkten Kontrolle des PPARα (Lundåsen et al., 2007). So konnte gezeigt
werden, dass durch Fasten und synthetische PPARα-selektive Agonisten die hepatische FGF21-
Expression induziert wird und zu einer Erhöhung der Plasmaspiegel an FGF21 führt, während in
PPARα knock out-Mäusen keine fasteninduzierte FGF21-Expression in der Leber ermittelt werden
konnte (Badman et al., 2007; Inagaki et al., 2007; Lundåsen et al., 2007).

Zielstellung
6
2. Zielstellung Ziel der vorliegenden Arbeit war die Aufklärung molekularer Mechanismen, über die oxidierte Fette
ihre Wirkungen im Organismus vermitteln.
Aus zahlreichen vorangegangen Untersuchungen an der Maus, der Ratte und dem Schwein ist
bekannt, dass die Aufnahme oxidierter Fette vielfältigste biologische Wirkungen im Organismus
hervorrufen kann (Chao et al., 2007; Skufca et al., 2003; Staprans et al., 2005; Sülzle et al., 2004). Eine
der Bedeutendsten ist die Induktion von oxidativem Stress (Eder et al., 2003a; Eder et al., 2003b;
Keller et al., 2004; Liu und Huang, 1996; Liu und Huang 1995), welcher zur Aktivierung der
antioxidativen Abwehr, detoxifizierender Mechanismen und auch proinflammatorischer Prozesse
führen kann.
So führte im Dünndarm der Ratte die Verabreichung eines oxidierten Fettes zu einer Erhöhung der
Expression antioxidativer Enzyme und proinflammatorischer Zytokine, welches als adaptative
Reaktion auf den durch die aufgenommenen Lipidperoxidationsprodukte verursachten oxidativen
Stress zurückgeführt wurde (Oliviero et al., 2010). Auch in der Leber der Ratte konnte durch die
Aufnahme eines oxidierten Fettes ein starker Anstieg der Phase I- und Phase II-Enzyme des
Fremdstoffmetabolismus beobachtet werden (Chen et al., 2005). Im Schwein resultierte die
Aufnahme eines oxidierten Fettes in einer Aktivierung der zellulären Stressantwort, wie an einer
gesteigerten hepatischen Genexpression und Aktivität zytoprotektiver Enzyme zu erkennen war
(Luci et al., 2007).
Die molekularen Mechanismen, die diesen Beobachtungen zugrunde liegen, sind bis heute jedoch
nicht vollständig geklärt. Da sowohl der Nrf2 als auch der NF-κB durch oxidativen Stress aktiviert
werden können und in die Regulation der zellulären Stressantwort involviert sind, sollte im ersten
Teil dieser Arbeit die Hypothese untersucht werden, dass die Aufnahme oxidierter Fette zu einer
Aktivierung dieser redox-sensitiven Transkriptionsfaktoren im Organismus führt (Studie 1 und 2).
In den beiden dieser Arbeit zugrundeliegenden Versuchen in den Modelltieren Maus und Schwein,
wurden Sonnenblumenöl (Studie 1) und Rapsöl (Studie 2 und 3) als Versuchsfette verwendet. Diese
Öle kommen in der Humanernährung vielfältig zum Einsatz. Aufgrund ihrer positiven Eigenschaften,
wie einer relativ hohen Oxidationsstabilität und Geschmacksneutralität und einem geringen Gehalt
an gesättigten Fettsäuren, werden sie häufig zum Erhitzen von Lebensmitteln, insbesondere dem
Frittieren, verwendet. Die Erhitzungsdauer (48 h bzw. 72 h) und -temperatur (190 °C bzw. 175 °C)
der Behandlungsfette orientierte sich an den Bedingungen zum Frittieren von Lebensmitteln in der
Humanernährung. Somit zeigen die in diesen Untersuchungen eingesetzten Versuchsfette eine hohe
Praxisrelevanz.

Zielstellung
7
Studie 1:
Um der aufgestellten Hypothese nachzugehen, wurde der erste Versuch in der Maus durchgeführt, da
sich diese als grundlagenorientiertes Modelltier zur Erforschung von molekularen Mechanismen
eignet. Zudem erwies sie sich bereits in früheren Fragestellungen zur Untersuchung antioxidativer
und inflammatorischer Signalwege im Darm als geeignetes in vivo-Labormodell (Khor et al., 2006). Des
Weiteren gestaltet sich die kontrollierte Verabreichung von Versuchsfetten über spezielle
Schlundsonden in der Maus als problemlos.
Um eine potentielle Aktivierung des NF-κB- und des Nrf2-Signalweges durch oxidiertes Fett
(Sonnenblumenöl) in der Dünndarmmukosa der Maus zu detektieren, wurden die nukleären
Konzentrationen an NF-κB/p50 und Nrf2 sowie die Genexpression oxidativer Stress-responsiver
Zielgene gemessen. Zusätzlich sollte der Einfluss des oxidierten Fettes auf die Aktivierung der
Transkriptionsfaktoren PPARα und PPARγ und der PPAR-responsiven Genexpression sowie eine
mögliche Interferenz mit dem redox-sensitiven NF-κB-Signalweg ermittelt werden.
Eine detaillierte Beschreibung von Material und Methodik, sowie Ergebnisse und Diskussion sind der
nachfolgend angeführten Publikation zu entnehmen (Studie 1).
Varady J, Eder K, Ringseis R. Dietary oxidized fat activates the oxidative stress-responsive
transcription factors NF-κB and Nrf2 in intestinal mucosa of mice. Eur J Nutr. 2011;50(8):601-9.
Reproduced with permission of Springer.
Studie 2:
Um zu überprüfen, ob die in der Maus erzielten Ergebnisse der Studie 1 sich auch in einer Spezies
nachweisen lassen, welche dem Menschen näher verwandt ist und daher Rückschlüsse auf diesen
zulässt, wurde eine weitere Untersuchung im Schwein durchgeführt.
Das Schwein zeichnet sich durch zahlreiche genetische, anatomische und stoffwechselphysiologische
Ähnlichkeiten und Gemeinsamkeiten zum Menschen, insbesondere der Physiologie des Magen-Darm-
Traktes, des Energie- und Lipidstoffwechsels und des hepatischen Fremdstoffmetabolismus aus
(Swindle et al., 2012; Bendixen et al., 2010; Bode et al., 2010; Barth et al., 1990). Bezüglich der
Expression an PPARα zählt das Schwein, ebenso wie Mensch und Affe, zu den nicht-proliferierenden
Spezies und steht somit im Gegensatz zu den Nagern, welche der proliferierenden Spezies angehören.
In Nagern führt die Aktivierung des hepatischen PPARα zur Induktion der Peroxisomenproliferation,
welche mit einer gesteigerten Expression von peroxisomalen Enzymen, wie der Acyl-CoA Oxidase
(ACO), einhergeht (Roberts et al., 2000). Begründet ist dies in den generell höheren Expressionsraten
des hepatischen PPARα im Vergleich zur nicht-proliferierenden Spezies und damit verbunden

Zielstellung
8
stärkeren Reaktion auf eine Aktivierung des PPARα (Cheon et al., 2005; Tugwood et al., 1998). Des
Weiteren erwies sich das Schwein zur Untersuchung von Fragestellungen im Zusammenhang mit
oxidierten Fetten in der Vergangenheit als sehr gut geeignet (Luci et al., 2007). Somit ist eine hohe
Übertragbarkeit der gewonnenen Ergebnisse vom Schwein auf den Menschen gewährleistet.
Im zweiten Versuch wurde außerdem ein oxidiertes Behandlungsfett (Rapsöl) verabreicht, in
welchem die Konzentration an polaren Anteilen unter 25 % lag. Dieser Wert wird von der Deutschen
Gesellschaft für Fettwissenschaft e. V. (DGF) als Grenzwert für die Genusstauglichkeit von
Frittierfetten empfohlen (DGF, 2000). Auch aus diesem Grund stellt das verwendete Diätfett des
zweiten Versuches ein in der Humanernährung praxisrelevantes Frittierfett dar.
Um eine potentielle Aktivierung des Nrf2-Signalweges in der Leber zu detektieren, wurden die
nukleären Konzentrationen an Nrf2 sowie die Expressionen Nrf2-abhängiger Gene, welche in die
Phase II des Fremdstoffmetabolismus involviert sind oder antioxidative Funktionen ausüben, in der
Leber gemessen. Phänotypische Auswirkungen einer potentiellen Nrf2-Aktivierung sollten über
Enzymaktivitätsmessungen der SOD und T4-UDP-Glukuronosyltransferase (T4-UGT) sowie
Bestimmung der Gesamt-T4-Konzentration im Plasma ermittelt werden. Um zu überprüfen, ob das
oxidierte Fett den NF-κB als weiteren redox-sensitiven Transkriptionsfaktor aktiviert, sollte
zusätzlich dessen nukleäre Proteinkonzentration bestimmt werden.
Eine detaillierte Beschreibung von Material und Methodik, sowie Ergebnisse und Diskussion sind der
nachfolgend angeführten Publikation zu entnehmen (Studie 2).
Varady J, Gessner DK, Most E, Eder K, Ringseis R. Dietary moderately oxidized oil activates the
Nrf2 signaling pathway in the liver of pigs. Lipids Health Dis. 2012;11:31.
Studie 3:
Im zweiten Teil der Arbeit sollte die Hypothese untersucht werden, dass die Aufnahme eines
oxidierten Fettes über eine Aktivierung des PPARα zur Induktion des FGF21 führt.
Es ist bekannt, dass eine durch Fasten, ketogene Diäten oder synthetische PPARα-Agonisten
verursachte Aktivierung des PPARα den FGF21 in der Leber induziert (Badman et al., 2007; Lundåsen
et al., 2007). Ob auch oxidierte Fette als potente nutritive Aktivatoren des PPARα den FGF21 zu
induzieren vermögen, ist bisher nicht bekannt. Die Ergebnisse dieser dritten Untersuchung sollten
daher einen Beitrag zur Aufklärung des Mechanismus leisten, wonach einige Effekte einer PPARα-
Aktivierung wie sie durch die Aufnahme oxidierter Fette beobachtet werden können, möglicherweise
über den FGF21 vermittelt werden.
Zur Untersuchung der aufgestellten Hypothese wurden die hepatische Genexpression des FGF21
sowie seine Proteinkonzentrationen in Leber und Plasma vom Schwein bestimmt. Um zu überprüfen,

Zielstellung
9
ob durch die Aufnahme des oxidierten Fettes auch eine Aktivierung des PPARα erfolgte, wurden die
hepatischen Genexpressionen klassischer PPARα-Zielgene gemessen.
Eine detaillierte Beschreibung von Material und Methodik, sowie Ergebnisse und Diskussion sind der
nachfolgend angeführten Publikation zu entnehmen (Studie 3).
Varady J, Ringseis R, Eder K. Dietary moderately oxidized oil induces expression of fibroblast
growth factor 21 in the liver of pigs. Lipids Health Dis. 2012;11:34.

3. Originalarbeiten

Eur J Nutr
DOI 10.1007/s00394-011-0181-8
OR IG INAL CONTRIBUTION
Dietary oxidized fat activates the oxidative stress-responsive
pathways NF-κB and Nrf2 in intestinal mucosa of mice Received: 14 December 2010/Accepted: 25 February 2011
© Springer-Verlag 2011
Abstract
Purpose Oxidized fats are known to induce oxidative stress resulting in the upregulation of
antioxidant enzymes as an adaptive response, with the underlying mechanism being unclear. It is
known that the response to oxidative stress is mediated by redox-sensitive transcription factors such
as NF-κB and Nrf2. The aim of the study therefore was to test the hypothesis that ingestion of an
oxidized fat causes the activation of these transcription factors in the small intestinal mucosa.
Methods Female mice were randomly assigned to 2 groups of 12 mice each and administered orally
by gavage either oxidized or fresh fat once per day. Results After 6 days of treatment, mice were
killed, intestinal mucosa was isolated, and nuclear concentration of NF-κB and Nrf2 and expression
of NF-κB- and Nrf2-regulated oxidative stress-responsive genes were determined. Oxidized fat
markedly increased nuclear concentration of NF-κB and Nrf2 and transcript levels of oxidative stress
responsive genes, like aldo-keto reductase 1B8, vanin-1, glutathione peroxidase 1, and superoxide
dismutase-1. In addition, oxidized fat increased the concentrations of PPAR-regulated genes.
Conclusions The activation of oxidative stress-sensitive pathways likely reflects an adaptive
response of the intestinal mucosa to prevent oxidative damage to the intestinal mucosa.
Keywords: Oxidized fat • Mucosa • Nrf2 • NF-κB • Mouse
Juliane Varady • Klaus Eder • Robert Ringseis ()
Institute of Animal Nutrition and Nutrition Physiology, Justus-Liebig-Universität Giessen,
Heinrich-Buff-Ring 26-32, 35392 Giessen, Germany
e-Mail: [email protected]
Published online: 10 March 2011

Introduction
Oxidized lipids as components of heated or fried foods play an important role in human nutrition in
industrialized countries. Animal experiments revealed that ingestion of heated fats containing lipid
peroxidation products provokes a wide array of biological effects. One of the most frequently
reported effects of oxidized fat is induction of oxidative stress and redox imbalances [1-8]. This has
been evidenced by elevated concentrations of lipid peroxidation products, reduced concentrations
of exogenous (e.g. tocopherols, ascorbic acid) and endogenous antioxidants (e.g. glutathione), and
a decreased glutathione/glutathione disulfide ratio in tissues of animals fed oxidized fat [1-8].
Because the intestinal mucosa is directly exposed to oxidized fats during intestinal passage after
ingestion of such fats, it is not surprising that oxidative stress and redox imbalances are especially
induced by oxidized fat in the intestinal mucosa [7, 8].
It is generally accepted that reactive oxygen metabolites generated during oxidative stress
are important mediators in the expression of genes involved in antioxidant defense and
inflammatory processes [9, 10]. This is explained by the activation of redox-sensitive transcription
factors, such as nuclear factor-κB (NF-κB), acting as regulators of genes involved in inflammation
and the adaptation to oxidative stress [11]. Intestinal NF-κB has long been considered to be
specifically associated with gut-associated lymphoid tissue. However, it has now been established
that NF-κB is also present in intestinal epithelial cells, which are the main cellular component of
the intestinal mucosa. Epithelial NF-κB is critical for maintaining the function and integrity of the
gut epithelial barrier by up-regulating genes involved in antimicrobial defense and attenuation of
oxidative stress [12]. Transcription factor nuclear factor-erythroid 2-related factor 2 (Nrf2) is
another important redox-sensitive transcription factor that protects tissues including the intestine
by inducing antioxidant and detoxifying genes in response to oxidative or xenobiotic stress [13].
A recent study clearly demonstrated that administration of a thermally oxidized oil strongly
induces the expression of antioxidant enzymes such as glutathione peroxidase 1 (GPX-1) and Co/Zn-
superoxide dismutase (SOD1) in the small intestine of rats being indicative of an adaptive response
of the intestine to the oxidative stress induced by the ingested lipid peroxidation products [14]. The
molecular mechanisms underlying this adaptive response, however, have not been studied yet. In
light of the role of NF-κB and Nrf2 in mediating the adaptive response to oxidative stress, we
hypothesize that ingestion of an oxidized fats causes activation of these transcription factors in the
small intestinal mucosa. To test this hypothesis, we performed a feeding experiment with mice that
were treated orally with an oxidized fat prepared under deep-frying conditions. As parameters we
considered the nuclear concentrations of the NF-κB subunit p50 and Nrf2 and the transcript levels of
genes which are involved in oxidative stress response and known to be regulated by either NF-κB or
Nrf2 or both of them, such as GPX-1, SOD1, heme-oxygenase 1 (HO-1), aldo-keto reductase 1B8
(AKR1B8), and vanin-1 [15-18]. Because the peroxidation products of oxidized fats, such as hydroxy
and hydroperoxy fatty acids, were shown to be ligands and activators of peroxisome proliferator-
activated receptor (PPAR) α and γ [19, 20], which are a known to interfere with oxidative stress
signalling pathways including NF-κB, we also studied the effect of oxidized fat on the activation of
PPARα and γ.
Materials and methods
Animals and diets
Female mice (129S4/SvImJ) supplied by Charles River with an initial body weight of 21.8 ± 1.9 g
(mean ± SD) were randomly assigned to 2 groups of 12 mice each. They were kept individually in
Macrolon cages in a room controlled for temperature (22 ± 2°C), relative humidity (50–60%), and
light (12-h-light/-dark cycle). All experimental procedures described followed established guidelines
for the care and handling of laboratory animals [21] and were approved by the council of Saxony-
Anhalt. All mice were orally administered 0.25 mL fresh or oxidized sunflower oil by gavage once

per day on 6 consecutive days 2 h after the beginning of the light cycle. All mice were fed a
commercial standard basal diet (Altromin 1324, Altromin, Lage, Germany). According to the
declaration of the manufacturer, this diet contained (per kilogram) 11.9 MJ metabolizable energy,
190 g crude protein, 60 g crude fiber, 40 g crude fat, and 70 g crude ash. To standardize food
intake, the diets were fed daily in restricted amounts of 2.5 g/d, equivalent to an intake of 29.8 kJ
metabolizable energy per day. Water was available ad libitum from nipple drinkers during the
entire experiment.
Preparation and chemical analysis of the oxidized oil
The thermo-oxidized oil was prepared by heating sunflower oil (obtained from a local supermarket)
in a domestic fryer from Bartscher GmbH (GF-8SE, Salzkotten, Germany) at 190°C for 48 h. The
extent of lipid peroxidation was determined by assaying the peroxide value [22], concentration of
thiobarbituric acid substances (TBARS) [23], concentration of conjugated dienes [24], acid value
[22] and the percentage of total polar compounds [25]. The fatty acid composition of the dietary
fats was determined by gas chromatography (GC). For that purpose, fats were methylated with
trimethylsulfonium hydroxide to fatty acid methyl esters (FAME) [26]. FAME were separated by GC
using a Chrompack 9000 system (Fa. Agilent Technologies, Waldbronn, Germany) fitted with an
automatic on-column injector, a polar capillary column (60 m FFAP, 0.25 mm internal diameter,
0.25 µm particle size, Macherey and Nagel, Düren, Germany), and a flame ionization detector [27].
Sample collection
At day 6, mice received the last dose of fresh or oxidized oil and 0.5 g of the diet 2 h after the
beginning of the light cycle and were killed 4 h later by decapitation under light anesthesia with
CO2. Blood was collected into heparinized polyethylene tubes. Plasma was obtained by
centrifugation of the blood (1,100 g, 10 min, 4°C) and stored at -20°C. The small intestine was
rapidly excised, flushed with ice-cold 0.9% NaCl (w/v), and opened length-wide. The mucosa was
scraped off, and snap-frozen for RNA isolation. All tissue samples were stored at -80°C pending
further analysis.
RNA isolation and real-time detection RT-PCR analysis
Total RNA was isolated from mucosa samples using TrizolTM reagent (Invitrogen, Karlsruhe, Germany)
according to the manufacturer’s protocol. Total RNA concentration and purity were estimated from
the optical density at 260 and 280 nm, respectively. Synthesis of cDNA and determination of mRNA
abundance by real-time detection PCR was performed as recently described in detail [28]. The
mRNA abundance of genes was measured by real-time detection PCR using SYBR Green I and the
Rotor Gene 2000 system (Corbett Research). Real-time detection PCR was performed with 1.25 units
Taq DNA polymerase, 500 mmol desoxyribonucleotide triphosphates, and 26.7 pmol of the gene-
specific primers (Operon, Köln, Germany). For determination of mRNA concentration, a threshold
cycle was obtained from each amplification curve using the software RotorGene 5.0 (Corbett
Research). Calculation of the relative mRNA concentration was made using the 2-ΔΔCt-method with
GAPDH as reference gene as previously described [29]. The relative mRNA concentration of the
genes investigated is expressed as fold change in the oxidized fat group compared to the fresh fat
group. Characteristics of gene-specific primers were as follows (forward 5’-3’, reverse 5’-3’; NCBI
GenBank): Adipophilin (CCCTGTCTACCAAGCTCT GC, CGATGCTTCTCTTCCACTCC; NM_007408.3),
AKR1B8 (CAAACCAAGTCAAAGAACTG, CTTG CTGACAATGAAGAGGTC; NM_008012.1), GAPDH
(TGTTCCTACCCCCAATGTGT, GTCATTGAGA GCAATGCCAG; NT_109320.4), GPX-1
(CTCTTTACCTTCCTGCGGAA, GGACAGCAGGGTTTCTAT GT; NM_008160.5), HO-1
(GATTTGTCTGAGGCCTTGAAG, CTTAAAGCCTTCTCTGGACAC; NM_0 10442.1), PPARα
(AGGCAGATGACCTGGAAAGTC, ATGCGTGAACTCCGTAGTGG; NM_01114 4.3), PPARγ
(CCAGTTTCGATCCGTAGAAG, CCATAAAGTCACCAAAGGGC; NM_011146.2), SOD1 (GATG
ACTTGGGCAAAGGTGG, CTGCGCAATCCCAATCACTC; NM_011434.1), Vanin-1 (GAAGT
GGTATCTATGCACCC, CTGTAGGTAGTACTGCCCTT; NM_011704.1).
Immunoblot analysis
Nuclear extracts were prepared from scraped mouse mucosa with a Nuclear Extract Kit (Active
Motif, Rixensart, Belgium) according to the manufacturer’s protocol. Protein concentrations in the
nuclear extracts were determined by the bicinchoninic acid protein assay kit (Interchim, Montluçon,
France) with BSA as standard. Four nuclear extract pools of each group were prepared with three
animals contributing equally to each pool. From each nuclear extract pool, 30 µg protein were
separated on 10% SDS-PAGE for PPARγ and NF-κB/p50 and 12.5% SDS-PAGE for Nrf2 and
electrotransferred to a nitrocellulose membrane (Pall Corporation, Pensacola, FL, USA). Loading of
equal amounts of protein in each line was verified by Ponceau S staining. After incubation the
membranes overnight at 4°C in blocking solution, membranes were incubated with primary
antibodies against Nrf2 (polyclonal anti-Nrf2 antibody; Abcam, Cambridge, UK) and NF-κB/p50
(polyclonal anti-NF-κB/p50 antibody; Santa Cruz, Heidelberg, Germany) and β-actin (monoclonal
anti-β-actin antibody, Abcam, Cambridge, UK) as a housekeeping protein to control for adequate
normalization at room temperature. The membranes were washed, and then incubated with a
horseradish peroxidase conjugated secondary anti-rabbit-IgG antibody (Sigma-Aldrich, Steinheim,
Germany) at room temperature. Afterwards, blots were developed using ECL Plus (GE Healthcare,
München, Germany). The signal intensities of specific bands were detected with a Bio-Imaging
system (Syngene, Cambridge, UK) and quantified using Syngene GeneTools software (nonlinear
dynamics).
Statistical analysis
Treatment effects were analyzed with one-way ANOVA using the Minitab Statistical software Rel.
13.0 (Minitab). For significant F-values, means were compared by Fisher’s multiple range test.
Values in the text are means ± SD. Means were considered significantly different at p < 0.05.
Results
Characterization of the experimental fats
Palmitic (16:0), stearic (18:0), oleic (18:1), and linoleic acid [18:2 (n-6)] were the major fatty acids
in both fats (Table 1). Due to oxidation-dependent loss of PUFA during heat treatment of the fat,
the oxidized fat had a lower proportion of linoleic acid and slightly higher proportions of saturated
fatty acids and oleic acid than the fresh fat (Table 1). The oxidized fat had much higher
concentrations of peroxides (2.9-fold), conjugated dienes (19-fold), TBARS (422-fold), polar
compounds (10.8-fold), and a higher acid value (2.7-fold) than the fresh fat (Table 1).
Table 1 Characteristics of the experimental oils
Fresh oil Oxidized oil
Major fatty acids, g/100 g total fatty acids
14:0 0.62 0.78
16:0 5.95 7.03
18:0 3.83 4.51
18:1 23.5 26.2
18:2 (n-6) 64.0 59.0
18:3 (n-3) 0.13 0.09
22:0 0.91 1.02
Peroxidation products
Peroxide value, mEq O2/kg 2.47 7.16
Conjugated dienes, mmol/kg 6.91 131
TBARS, mmol/kg < 0.1 422
Total polar compounds, % 2.87 31.0
Acid value, g KOH/kg 0.20 0.53
Initial and final body weights
Initial (fresh fat, 21.8 ± 1.98 g; oxidized fat, 21.7 ± 1.94 g; n = 12; p < 0.05) and final (fresh fat,
21.1 ± 1.68 g; oxidized fat, 21.4 ± 1.63 g; n = 12; P < 0.05) body weights did not differ between both
groups of mice.
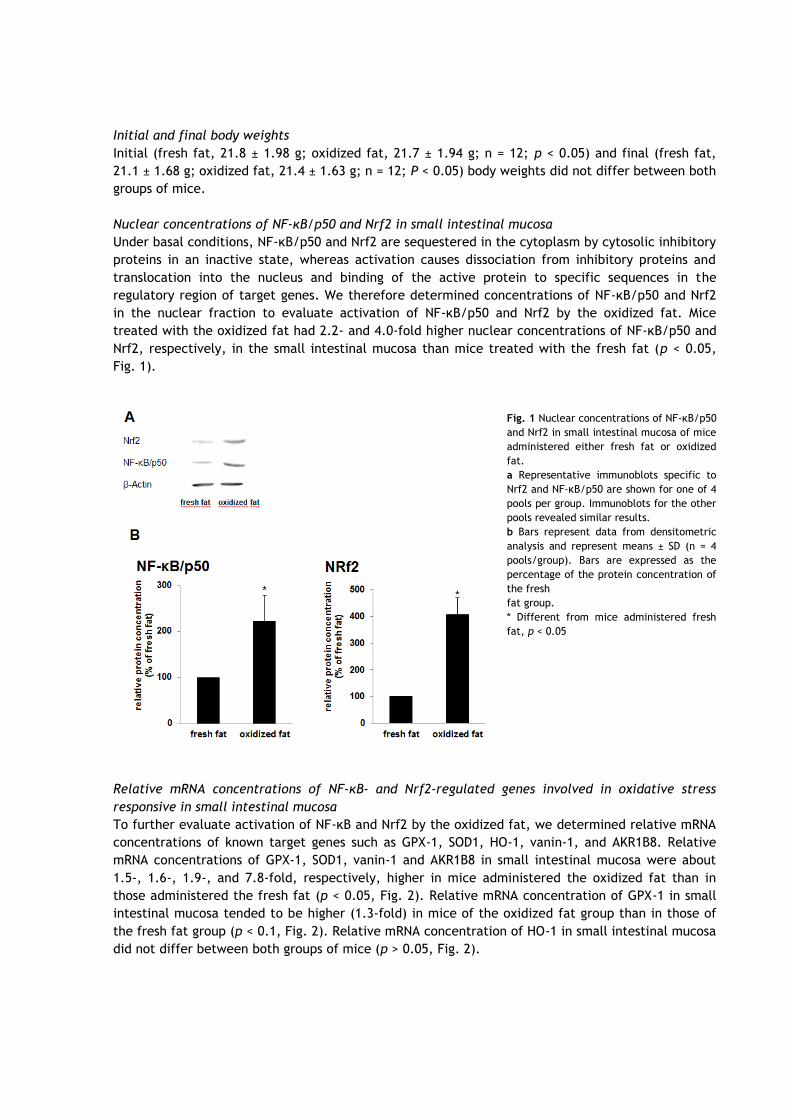
Nuclear concentrations of NF-κB/p50 and Nrf2 in small intestinal mucosa
Under basal conditions, NF-κB/p50 and Nrf2 are sequestered in the cytoplasm by cytosolic inhibitory
proteins in an inactive state, whereas activation causes dissociation from inhibitory proteins and
translocation into the nucleus and binding of the active protein to specific sequences in the
regulatory region of target genes. We therefore determined concentrations of NF-κB/p50 and Nrf2
in the nuclear fraction to evaluate activation of NF-κB/p50 and Nrf2 by the oxidized fat. Mice
treated with the oxidized fat had 2.2- and 4.0-fold higher nuclear concentrations of NF-κB/p50 and
Nrf2, respectively, in the small intestinal mucosa than mice treated with the fresh fat (p < 0.05,
Fig. 1).
Fig. 1 Nuclear concentrations of NF-κB/p50
and Nrf2 in small intestinal mucosa of mice
administered either fresh fat or oxidized
fat.
a Representative immunoblots specific to
Nrf2 and NF-κB/p50 are shown for one of 4
pools per group. Immunoblots for the other
pools revealed similar results.
b Bars represent data from densitometric
analysis and represent means ± SD (n = 4
pools/group). Bars are expressed as the
percentage of the protein concentration of
the fresh
fat group.
* Different from mice administered fresh
fat, p < 0.05
Relative mRNA concentrations of NF-κB- and Nrf2-regulated genes involved in oxidative stress
responsive in small intestinal mucosa
To further evaluate activation of NF-κB and Nrf2 by the oxidized fat, we determined relative mRNA
concentrations of known target genes such as GPX-1, SOD1, HO-1, vanin-1, and AKR1B8. Relative
mRNA concentrations of GPX-1, SOD1, vanin-1 and AKR1B8 in small intestinal mucosa were about
1.5-, 1.6-, 1.9-, and 7.8-fold, respectively, higher in mice administered the oxidized fat than in
those administered the fresh fat (p < 0.05, Fig. 2). Relative mRNA concentration of GPX-1 in small
intestinal mucosa tended to be higher (1.3-fold) in mice of the oxidized fat group than in those of
the fresh fat group (p < 0.1, Fig. 2). Relative mRNA concentration of HO-1 in small intestinal mucosa
did not differ between both groups of mice (p > 0.05, Fig. 2).

Relative mRNA concentrations of PPARα and PPARγ and PPARα/PPARγ-regulated genes in small
intestinal mucosa
To evaluate activation of the PPARα and PPARγ pathways by the oxidized fat, we determined
relative mRNA concentrations of the receptors and selected PPARα/PPARγ-target genes. Mice
administered the oxidized fat had 3.6-, 2.6-, 5.6-, and 1.5-fold higher relative mRNA concentrations
of PPARα, FATP, CYP4A10, and adipophilin in small intestinal mucosa than mice administered the
fresh fat (p < 0.05, Fig. 3). Relative mRNA concentration of PPARγ in small intestinal did not differ
between both groups of mice (p > 0.05, Fig. 3).
Fig. 2 Relative mRNA
concentrations of NF-κB-
and Nrf2-regulated genes
involved in oxidative
stress response in the
intestinal mucosa of mice
administered either fresh
fat or oxidized fat. Bars
represent means ± SD (n =
12/group) and are
expressed as the
percentage of the
relative mRNA
concentration of the
fresh fat group.
*Different from mice
administered fresh fat, p
< 0.05
Fig. 3 Relative mRNA
concentrations of PPARα
and PPARγ and
PPARα/PPARγ-regulated
genes in the intestinal
mucosa of mice
administered either
fresh fat or oxidized fat.
Bars represent means ±
SD (n = 12/group) and
are expressed as the
percentage of the
relative mRNA
concentration of the
fresh fat group.
*Different from mice
administered fresh fat,
p < 0.05

Discussion
The present study tested the hypothesis that ingestion of an oxidized fat causes activation of
oxidative stress-sensitive transcription factors, such as NF-κB and Nrf2, in the small intestinal
mucosa. The oxidized fat used in this study was prepared by heating sunflower oil at a relatively
high temperature over a short period. This reflects a fat that has been prepared under conventional
deep-frying conditions. The relatively high concentrations of lipid peroxidation products
(conjugated dienes, TBARS, peroxides and carbonyls) in the oxidized fat indicate that this fat was
strongly oxidized. Such fats generally contain high concentrations of secondary lipid peroxidation
products such as aldehydes or ketones and rather low concentrations of primary lipid peroxidation
products such as hydroxy and hydroperoxy fatty acids.
The present study shows for the first time that administration of an oxidized fat activates
the transcription factors NF-κB and Nrf2 in the intestinal mucosa of mice. It is well known that
peroxidation products from oxidized fats are efficiently taken up by the intestinal epithelial cells
[30, 31], and that the cellular exposure to oxidized fats leads to the generation of ROS [32].
Regarding that NF-κB and Nrf2 are activated by various stimuli including ROS [13, 33], it is therefore
very likely that the ingestion of lipid peroxidation products and the resulting oxidative stress in the
intestinal mucosa has led to the activation of NF-κB and Nrf2. This finding in mice is in contrast to a
recent observation in pigs, in which oxidized fat did not activate NF-κB in the small intestinal
mucosa [34]. Although it cannot be excluded that the effect in mice is a species-specific
phenomenon, it is more likely that differences in the administration of the oxidized fat are
responsible for the divergent effect between mice and pigs. In the pig study, the oxidized fat was
fed as part of a normal diet, whereas in the mice study, the oxidized fat was administered by
gavage. In the latter case, the intestinal mucosa is probably more directly exposed to the ingested
oxidized fat than in the case that the oxidized fat is virtually diluted by the other feed components.
A further reason for the lack of response in the pig study might be that the amount of fat
administered to the animals when related to their body weights was clearly higher in the mice study
than in the pig study. Our study, moreover, showed that administration of oxidized fat caused an
increase in the transcript levels of several oxidative stress-responsive genes, including SOD1,
AKR1B8, vanin-1, and GPX-1, in the intestinal mucosa. Up-regulation of SOD1 and GPX-1 in the
intestine and other tissues by oxidized lipids has been already observed in previous studies [14, 35],
and up-regulation of these genes has been generally interpreted as an adaptive, defensive response
of tissues to prevent possible harmful effects of ROS [36, 37]. A recent study demonstrated that the
feeding of oxidized fat also results in increased activities of antioxidant enzymes such as catalase
and selendependent GPX in the intestine [14] indicating that transcriptional up-regulation of
antioxidant genes is also translated to the metabolic level. To the best of our knowledge, up-
regulation of AKR1B8 and vanin-1 by oxidized fat has not been described yet. AKRs like AKR1B8
catalyze the reduction of both environmental aldehydes and aldehydes generated endogenously
from lipid peroxidation such as 4-hydroxynonenal or core aldehydes [38]. Due to these properties, it
has been suggested that AKR1B8, like GPX-1 and SOD1, is an important component of the cellular
antioxidant defense mechanisms that protect against injurious lipid peroxidation products [39].
Induction of the epithelial cell-specific vanin-1 is also indicative of the induction of oxidative stress,
because cysteamine provided by the pantetheinase activity of this ectoenzyme acts pro-oxidative in
glutathione metabolism and activates stress pathways. In addition, vanin-1 facilitates the intestinal
epithelial cells to produce inflammatory mediators. This likely explains that vanin-1-deficient mice,
which lack tissue cysteamine, show a remarkably increased resistance to stress and acute intestinal
inflammation [40, 41]. We, therefore, postulate that the induction of AKR1B8 and vanin-1 by
oxidized fats is probably also part of the adaptive response of the intestinal mucosa to cope with
the oxidative stress induced by the lipid peroxidation products.
Induction of the abovementioned oxidative stress-responsive genes in response to ROS is
well-known to be mediated by the binding of activated NF-κB/p50 and Nrf2 to specific DNA-
sequences, called NF-κB response element (NF-κB-RE) and antioxidant response element (ARE),
respectively, which are present in the 5´-flanking regulatory region of those genes. Although we did

not perform gel shift experiments in this study to confirm increased binding to NF-κB-REs and AREs,
we are confident about this because nuclear concentrations of NF-κB/p50 and Nrf2 were markedly
elevated by the oxidized fat and the increased nuclear concentrations of these transcription factors
correlated well with the increased levels of NF-κB-RE- and ARE-mediated gene products. It might be
argued that nuclear protein concentrations of NF-κB/p50 and Nrf2 were elevated due to inadequate
normalization with β-actin which is known to translocate between the cytosol and nucleus [42] and
is therefore present in both cell fractions. However, β-actin is frequently used as a housekeeping
protein for nuclear proteins [43, 44] and considered suitable for normalization of nuclear protein
levels provided that it is constitutively and stably expressed under the specific experimental
conditions. Since there is no evidence from the literature that oxidized fat influences the
intracellular translocation of β-actin, the band intensities for β-actin were similar between both
groups, and equal amounts of protein were separated by SDS–PAGE, we are confident that protein
levels of NF-κB/p50 and Nrf2 were adequately normalized. Thus, we propose that the observed up-
regulation of the abovementioned genes in the intestinal mucosa of mice administered the oxidized
fat is mediated by the activation of oxidative stress-sensitive transcription factors such as NF-κB
and Nrf2. Since the abovementioned genes are also regulated, at least partially, by other oxidative
stress-responsive transcription factors such as activator protein (AP)-1 [45], it is not unlikely that
activation of the abovementioned genes by oxidized fat is mediated in cooperation of NF-κB and
Nrf2 with other transcription factors. This has to be clarified in future studies.
In contrast to the present study, recent studies showed that Nrf2 activation causes
inhibition of NF-κB [46–48]. The negative crosstalk between these two transcription factors has been
explained by different mechanisms, such as competition between Nrf2 and NF-κB for the CREB-
binding protein (CBP) and recruitment of histone deacetylase 3, a co-repressor of ARE, through the
NF-κB subunit p65 and subsequent interaction with CBP or Maf kinases[49]. The fact that we did not
find evidence for a negative cross-talk between Nrf2 and NF-κB may indicate that components of
oxidized fat, unlike phytochemicals such as sulforaphane, stimulate a further yet unidentified
pathway preventing inhibition of the NF-κB pathway by Nrf2.
In contrast to SOD1, GPX-1, AKR1B8 and vanin-1, HO-1, which is involved in heme
catabolism, was not induced in the intestinal mucosa by the oxidized fat, although HO-1 is also
regarded as a sensitive and reliable indicator of cellular oxidative stress [50], and the HO-1 gene
promoter was shown to contain binding sites for both, NF-kB and Nrf2 [51, 52]. Therefore, we
cannot explain the observation that oxidized fat did not induce HO-1 in the intestinal mucosa,
though the nuclear concentrations of NF-κB and Nrf2 were significantly elevated. However, one can
speculate that in the present mice experiment, NF-κB has inhibited the binding of Nrf2 to the HO-1
promoter by specifically impeding binding of the above-mentioned CBP to Nrf2 at the HO-1
promoter
In the present study we also considered the PPARα- and PPARγ-signaling pathways, because
it was shown that these pathways can negatively interfere with oxidative stress signalling pathways
including NF-κB. Herein, we showed that the transcript levels of PPARα/γ-responsive genes, like
FATP, CYP4A1 and adipophilin, were strongly increased by the oxidized fat in the intestinal mucosa
of the mice. These findings are in good agreement with recent studies showing that oxidized fats
cause activation of PPARα in liver and intestine of rats and pigs [53-55]. From the present findings,
we cannot distinguish whether up-regulation of FATP, CYP4A1, and adipophilin was due to
activation of PPARα, PPARγ or both of them, because the components of oxidized fat supposed to
be responsible for PPARα activation are also ligands and activators of PPARγ and FATP, CYP4A1, and
adipophilin are regulated by different PPAR isotypes [56-59]. In any case, the present findings
clearly indicate that activation of PPAR-signaling by oxidized fat in the intestinal mucosa does not
counteract the activation of oxidative stress-responsive pathways such as NF-κB and Nrf2.
In conclusion, the present study shows that ingestion of an oxidized fat causes a significant
up-regulation of several oxidative stress-responsive genes including SOD1, GPX-1, AKR1B8, and
vanin-1 in the intestinal mucosa which is likely mediated by the activation of oxidative stress-
sensitive transcription factors such as NF-κB and Nrf2. We postulate that up-regulation of these
genes which have antioxidant, cytoprotective and detoxifying functions is an adaptive response of

the intestinal mucosa to cope with the luminal diet-derived oxidants thereby preventing ROS-
mediated damage to the intestinal mucosa. In this context, we further postulate that oxidized fat,
similar to the previously reported effects of resveratrol and physical exercise [60, 61], may induce
the generation of low levels of ROS which could comprise a positive ‘‘hormetic redox signal’’ in the
enterocyte thereby enhancing stress resistance. This is in contrast to the detrimental ‘‘cellular
stress signal’’ induced by high levels of ROS generated from high doses of ascorbic acid resulting in
inactivation of Nrf2 [60].
Acknowledgment This study was supported by a grant from the Deutsche Forschungsgemeinschaft (DFG; grant no. RI
1537/1-1).
References
1. Izaki Y, Yoshikawa S, Uchiyama M. Effect of ingestion of thermally oxidized frying oil on peroxidative criteria in rats. Lipids 1984; 19: 324-31.
2. Kok TS, Harris PG, Alexander JC. Heated canola oil and oxidative stress in rats. Nutr Res 1988; 8: 673-84.
3. Liu JF, Huang CJ. Tissue α-tocopherol retention in male rats is compromised by feeding diets containing oxidized frying oil. J Nutr 1995; 125: 3071-80.
4. Liu JF, Huang CJ. Dietary oxidized frying oil enhances tissue α-tocopherol depletion and radioisotope tracer excretion in vitamin E-deficient rats. J Nutr 1996; 126: 2227-35.
5. Eder K, Keller U, Hirche F, Brandsch C. Thermally oxidized dietary fats increase the susceptibility of rat LDL to lipid peroxidation but not their uptake by macrophages. J Nutr 2003; 133: 2830-7.
6. Keller U, Brandsch C, Eder K. Supplementation of vitamins C and E increases the vitamin E status but does not prevent the formation of oxysterols in the liver of guinea pigs fed an oxidised fat. Eur J Nutr 2004; 43: 353-9.
7. Tsunada S, Iwakiri R, Noda T, Fujimoto K, Fuseler J, Rhoads CA, Aw TY. Chronic exposure to subtoxic levels of peroxidized lipids suppresses mucosal cell turnover in rat small intestine and reversal by glutathione. Dig Dis Sci 2003; 48: 210-22.
8. Tsunada S, Iwakiri R, Fujimoto K, Aw TY. Chronic lipid hydroperoxide stress suppresses mucosal proliferation in rat intestine: potentiation of ornithine decarboxylase activity by epidermal growth factor. Dig Dis Sci 2003; 48: 2333-41.
9. Schreck R, Rieber P, Baeuerle PA. Reactive oxygen intermediates as apparently widely used messengers in the activation of the NF-κB transcription factor and HIV-1. EMBO J 1991; 10: 2247-58.
10. Sen CK, Packer L. Antioxidant and redox regulation of gene transcription. FASEB J 1996; 10: 709-20.
11. Barnes PJ, Karin M. Nuclear factor-κB: a pivotal transcription factor in chronic inflammatory diseases. New Engl J Med 1997; 336: 1066-71.
12. Ben-Neriah Y, Schmidt-Supprian M. Epithelial NF-κB maintains host gut microflora homeostasis. Nat Immunol 2007; 8: 479-81.
13. Niture SK, Kaspar JW, Shen J, Jaiswal AK (2010) Nrf2 signaling and cell survival. Toxicol Appl
Pharmacol 244:37–42
14. Olivero David R, Bastida S, Schultz A, Gonza´lez Torres L, González-Muñoz MJ, Sánchez-Muniz
FJ, Benedí J (2010) Fasting status and thermally oxidized sunflower oil ingestion affect the
intestinal antioxidant enzyme activity and gene expression of male Wistar rats. J Agric Food
Chem 58:2498–2504
15. Rojo AI, Salinas M, Martı´n D, Perona R, Cuadrado A (2004) Regulation of Cu/Zn-superoxide
dismutase expression via the phosphatidylinositol 3 kinase/Akt pathway and nuclear factor-
κB. J Neurosci 24:7324–7334
16. Andreadi CK, Howells LM, Atherfold PA, Manson MM (2006) Involvement of Nrf2, p38, B-Raf,
and nuclear factor-κB, but not phosphatidylinositol 3-kinase, in induction of hemeoxygenase-1
by dietary polyphenols. Mol Pharmacol 69:1033–1040
17. Zhou LZ, Johnson AP, Rando TA (2001) NF-κB and AP-1 mediate transcriptional responses to
oxidative stress in skeletal muscle cells. Free Radic Biol Med 31:1405–1416

18. Malone PE, Hernandez MR (2007) 4-Hydroxynonenal, a product of oxidative stress, leads to an
antioxidant response in optic nerve head astrocytes. Exp Eye Res 84:444–454
19. Bull AW, Steffensen KR, Leers J, Rafter JJ (2003) Activation of PPARγ in colon tumor cell lines
by oxidized metabolites of linoleic acid, endogenous ligands for PPARγ. Carcinogenesis
24:1717–1722
20. Delerive P, Furman C, Teissier E, Fruchart J, Duriez P, Staels B (2000) Oxidized phospholipids
activate PPARα in a phospholipase A2-dependent manner. FEBS Lett 471:34–38
21. National Research Council (1985) Guide for the care and use of laboratory animals.
Publication no. 85–23 (rev.). National Institutes of Health, Washington
22. Deutsche Gesellschaft für Fettwissenschaften (1994) Einheitsmethoden zur Untersuchung von
Fetten, Fettprodukten, Tensiden und verwandten Stoffen. Stuttgart (Germany),
Wissenschaftliche Verlagsgesellschaft
23. Sidwell CG, Salwin H, Benca M, Mitchell JH Jr (1954) The use of thiobarbituric acid as a
measure of fat oxidation. J Am Oil Chem Soc 31:603–617
24. Recknagel RO, Glende EA Jr (1984) Spectrophotometric detection of lipid conjugated dienes.
Methods Enzymol 105:331–337
25. International Union of Pure and Applied Chemistry (IUPAC) (2000) Determination of polar
compounds, polymerized and oxidized triacylglycerols, and diacylglycerols in oils and fats.
Pure Appl Chem 72:1563–1575
26. Butte W (1983) Rapid method for the determination of fatty acid profiles from fats and oils
using trimethylsulfonium hydroxide for transesterification. J Chromatogr 261:142–145
27. Brandsch C, Ringseis R, Eder K (2002) High dietary iron concentrations enhance the formation
of cholesterol oxidation products in the liver of adult rats fed salmon oil with minimal effects
on antioxidant status. J Nutr 132:2263–2269
28. Ringseis R, Pösel S, Hirche F, Eder K (2007) Treatment with pharmacological peroxisome
proliferator-activated receptor a agonist clofibrate causes upregulation of organic cation
transporter 2 in liver and small intestine of rats. Pharmacol Res 56:175–183
29. Ringseis R, Muschick A, Eder K (2007) Dietary oxidized fat prevents ethanol-induced
triacylglycerol accumulation and increases expression of PPARα target genes in rat liver. J
Nutr 137:77–83
30. Penumetcha M, Khan N, Parthasarathy S (2000) Dietary oxidized fatty acids: an atherogenic
risk? J Lipid Res 41:1473–1480
31. Staprans I, Rapp JH, Pan XM, Feingold KR (1993) The effect of oxidized lipids in the diet on
serum lipoprotein peroxides in control and diabetic rats. J Clin Investig 92:638–643
32. Liu JF, Lee YW (1998) Vitamin C supplementation restores the impaired vitamin E status of
guinea pigs fed oxidized frying oil. J Nutr 128:116–122
33. Schoonbroodt S, Piette J (2000) Oxidative stress interference with the nuclear factor-κb
activation pathways. Biochem Pharmacol 60:1075–1083
34. Ringseis R, Piwek N, Eder K (2007) Oxidized fat induces oxidative stress but has no effect on
NF-kappaB-mediated proinflammatory gene transcription in porcine intestinal epithelial cells.
Inflamm Res 56:118–125
35. Ringseis R, Eder K (2004) Increases expression and activity of antioxidative enzymes and
reduces the concentration of glutathione in the liver of rats. Int J Vitam Nutr Res 74:86–92
36. Levonen AL, Inkala M, Heikura T et al (2007) Nrf2 gene transfer induces antioxidant enzymes
and suppresses smooth muscle cell growth in vitro and reduces oxidative stress in rabbit aorta
in vivo. Arterioscl Thromb Vasc Biol 27:741–747
37. Takada Y, Mukhopadhyay A, Kundu GC, Mahabeleshwar GH, Singh S, Aggarwal BB (2003)
Hydrogen peroxide activates NF-κB through tyrosine phosphorylation of IkBα and serine
phosphorylation of p65. J Biol Chem 278:24233–24241
38. Srivastava S, Harter TM, Chandra A, Bhatnagar A, Srivastava SK, Petrash JM (1998) Kinetic
studies of FR-1, a growth factorinducible aldo-keto reductase. Biochemistry 37:12909–12917

39. Spite M, Baba SP, Ahmed Y, Barski OA, Nijhawan K, Petrash JM, Bhatnagar A, Srivastava S
(2007) Substrate specificity and catalytic efficiency of aldo-keto reductases with phospholipid
aldehydes. Biochem J 405:95–105
40. Berruyer C, Pouyet L, Millet V et al (2006) Vanin-1 licenses inflammatory mediator production
by gut epithelial cells and controls colitis by antagonizing peroxisome proliferator-activated
receptor c activity. J Exp Med 203:2817–2827
41. Martin F, Penet MF, Malergue F et al (2004) Vanin-1(-/-) mice show decreased NSAID- and
Schistosoma-induced intestinal inflammation associated with higher glutathione stores. J Clin
Investig 113:591–597
42. Xu YZ, Thuraisingam T, de Lima Morais DA, Rola-Pleszczynski M, Radzioch D (2010) Nuclear
translocation of ß-actin is involved in transcriptional regulation during macrophage
differentiation of HL-60 cells. Mol Biol Cell 21:811–820
43. Long I, Suppian R, Ismail Z (2011) Increases in mRNA and DREAM protein expression in the rat
spinal cord after formalin induced pain. Neurochem Res 36:533–539
44. Sahin K, Tuzcu M, Gencoglu H, Dogukan A, Timurkan M, Sahin N, Aslan A, Kucuk O (2010)
Epigallocatechin-3-gallate activates Nrf2/HO-1 signaling pathway in cisplatin-induced
nephrotoxicity in rats. Life Sci 87:240–245
45. Harada H, Sugimoto R, Watanabe A et al (2008) Differential roles for Nrf2 and AP-1 in
upregulation of HO-1 expression by arsenite in murine embryonic fibroblasts. Free Radic Res
42:297–304
46. Lin W, Wu RT, Wu T, Khor TO, Wang H, Kong AN (2008) Sulforaphane suppressed LPS-induced
inflammation in mouse peritoneal macrophages through Nrf2 dependent pathway. Biochem
Pharmacol 76:967–973
47. Liu YC, Hsieh CW, Wenig YC, Chuang SH, Hsieh CY, Wung BS (2008) Sulforaphane inhibition of
monocyte adhesion via the suppression of ICAM-1 and NF-kappaB is dependent upon
glutathione depletion in endothelial cells. Vasc Pharmacol 48:54–61
48. Wagner AE, Ernst I, Iori R, Desel C, Rimbach G (2010) Sulforaphane but not ascorbigen,
indole-3-carbinole and ascorbic acid activates the transcription factor Nrf2 and induces
phase-2 and antioxidant enzymes in human keratinocytes in culture. Exp Dermatol 19:137–144
49. Liu GH, Qu J, Shen X (2008) NF-κB/p65 antagonizes Nrf2-ARE pathway by depriving CBP from
Nrf2 and facilitating recruitment of HDAC3 to MafK. Biochim Biophys Acta 1783:713–727
50. Shibahara S (1988) Regulation of heme oxygenase gene expression. Semin Hematol 25:370–376
51. Alam J, Stewart D, Touchard C, Boinapally S, Choi AM, Cook JL (1999) Nrf2, a Cap’n’Collar
transcription factor, regulates induction of the heme oxygenase-1 gene. J Biol Chem
274:26071–26078
52. Lavrovsky Y, Schwartzman ML, Levere RD, Kappas A, Abraham NG (1994) Identification of
binding sites for transcription factors NF-kappa B and AP-2 in the promoter region of the
human heme oxygenase 1 gene. PNAS 91:5987–5991
53. Chao PM, Chao CY, Lin FJ, Huang C (2001) Oxidized frying oil up-regulates hepatic acyl-CoA
oxidase and cytochrome P450 4 A1 genes in rats and activates PPARα. J Nutr 131:3166–3174
54. Sülzle A, Hirche F, Eder K (2004) Thermally oxidized dietary fat upregulates the expression of
target genes of PPARα in rat liver. J Nutr 134:1375–1383
55. Luci S, König B, Giemsa B, Huber S, Hause G, Kluge H, Stangl GI, Eder K (2007) Feeding of a
deep-fried fat causes PPARα activation in the liver of pigs as a non-proliferating species. Br J
Nutr 97:872–882
56. Zhou XR, Sun CH, Liu JR, Zhao D (2008) Dietary conjugated linoleic acid increases PPARγ gene
expression in adipose tissue of obese rat, and improves insulin resistance. Growth Horm IGF
Res 18:361–368
57. Martin G, Schoonjans K, Lefebvre AM, Staels B, Auwerx J (1997) Coordinate regulation of the
expression of the fatty acid transport protein and acyl-CoA synthetase genes by PPARα and
PPARγ activators. J Biol Chem 272:28210–28217

58. Bildirici I, Roh CR, Schaiff WT, Lewkowski BM, Nelson DM, Sadovsky Y (2003) The lipid droplet-
associated protein adipophilin is expressed in human trophoblasts and is regulated by
peroxisomal proliferator-activated receptor-γ/retinoid X receptor.
J Clin Endocrinol Metab 88:6056–6062
59. Barclay TB, Peters JM, Sewer MB, Ferrari L, Gonzalez FJ, Morgan ET (1999) Modulation of
cytochrome P-450 gene expression in endotoxemic mice is tissue specific and peroxisome
proliferator-activated receptor-α dependent. J Pharmacol Exp Therap 290:1250–1257
60. Wagner AE, Boesch-Saadatmandi C, Breckwoldt D, Schrader C, Schmelzer C, Döring F, Hashida
K, Hori O, Matsugo S, Rimbach G (2011) Ascorbic acid partly antagonizes resveratrol mediated
heme oxygenase-1 but not paraoxonase-1 induction in cultured hepatocytes—role of the
redox-regulated transcription factor Nrf2. BMC Complement Altern Med 11:1
61. Ristow M, Zarse K, Oberbach A, Klöting N, Birringer M, Kiehntopf M, Stumvoll M, Kahn CR,
Blüher M (2009) Antioxidants prevent health-promoting effects of physical exercise in
humans. Proc Natl Acad Sci USA 106:8665–8670

RESEARCH Open Access
Dietary moderately oxidized oil activates the Nrf2signaling pathway in the liver of pigsJuliane Varady, Denise K Gessner, Erika Most, Klaus Eder and Robert Ringseis*
Abstract
Background: Previous studies have shown that administration of oxidized oils increases gene expression andactivities of various enzymes involved in xenobiotic metabolism and stress response in the liver of rats and guineapigs. As these genes are controlled by nuclear factor erythroid-derived 2-like 2 (Nrf2), we investigated thehypothesis that feeding of oxidized fats causes an activation of that transcription factor in the liver which in turnactivates the expression of antioxidant, cytoprotective and detoxifying genes.
Methods: Twenty four crossbred pigs were allocated to two groups of 12 pigs each and fed nutritionally adequatediets with either fresh rapeseed oil (fresh fat group) or oxidized rapeseed oil prepared by heating at a temperatureof 175°C for 72 h (oxidized fat group).
Results: After 29 days of feeding, pigs of the oxidized fat group had a markedly increased nuclear concentration ofthe transcription factor Nrf2 and a higher activity of cellular superoxide dismutase and T4-UDPglucuronosyltransferase in liver than the fresh fat group (P < 0.05). In addition, transcript levels of antioxidant andphase II genes in liver, like superoxide dismutase 1, heme oxygenase 1, glutathione peroxidase 1, thioredoxinreductase 1, microsomal glutathione-S-transferase 1, UDP glucuronosyltransferase 1A1 and NAD(P)H:quinoneoxidoreductase 1 in the liver were higher in the oxidized fat group than in the fresh fat group (P < 0.05).Moreover, pigs of the oxidized fat group had an increased hepatic nuclear concentration of the transcription factorNF-�B which is also an important transcription factor mediating cellular stress response.
Conclusion: The present study shows for the first time that administration of an oxidized fat activates the Nrf2 inthe liver of pigs which likely reflects an adaptive mechanism to prevent cellular oxidative damage. Activation of theNF-�B pathway might also contribute to this effect of oxidized fat.
Keywords: Antioxidant enzymes, Liver, Nrf2, Oxidized fat, Phase II enzymes, Pig
BackgroundIn recent years, the contribution of oxidized fats to totalenergy intake has markedly increased in industrializedcountries due to the rising consumption of deep-friedproducts [1]. In fast food restaurants, foodstuffs are typi-cally fried in fats in fryers at temperatures close to 180°C.During the frying process, several chemical reactionsoccur within the frying oil resulting in the formation of amixture of chemically distinct lipid peroxidation products[2]. Large quantities of the frying oil are absorbed intothe fried food during deep-frying and therefore ingestedduring their consumption.
Feeding experiments with animals revealed that inges-tion of oxidized fats provokes a wide array of biologicaleffects [3-5]. One of the most striking effects of oxidizedfat is the induction of oxidative stress which is due tolipid hydroperoxides absorbed from the ingested oxidizedfats and reactive oxygen species (ROS) generated frommicrosomal cytochrome P450 enzymes which areinduced by oxidized fat [6-8]. Oxidative stress in animalsfed oxidized fats is evident by elevated concentrations oflipid peroxidation products, reduced concentrations ofexogenous and endogenous antioxidants, and a decreasedratio of reduced and oxidized glutathione in plasma andtissues [8-13].Previous studies have shown that administration of
oxidized oils increases gene expression and activities of* Correspondence: [email protected] of Animal Nutrition and Nutrition Physiology, Justus-Liebig-University Giessen, Giessen, Germany
Varady et al. Lipids in Health and Disease 2012, 11:31http://www.lipidworld.com/content/11/1/31
© 2012 Varady et al; licensee BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative CommonsAttribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction inany medium, provided the original work is properly cited.

various enzymes involved in xenobiotic metabolism andstress response in the liver of rats and guinea pigs[14-18]. Hepatic xenobiotic metabolism and stressresponse is mainly controlled by nuclear factor ery-throid-derived 2-like 2 (Nrf2), a master transcriptionfactor shown to regulate more than 200 genes, includingthose involved in phase II detoxification and antioxidantdefense [19]. Nrf2 pathway is regarded as the mostimportant pathway in the cell to protect cells againstoxidative stress [20,21]. Thus, the regulation of Nrf2activity represents a critical step in initiating a cellularantioxidant response to ROS. It has been shown thatoxidation products of n-3 fatty acids are able to activateNrf2 in a human liver cell line, and thus to induce theexpression of Nrf2 target genes involved in cellulardefense [22]. More recently, we found that the ingestionof a dietary oxidized fat activates Nrf2 pathway in theintestinal mucosa of mice [23]. Based on these findings, itis likely that the induction of xenobiotic metabolism inthe liver of animals fed an oxidized fat is due to an activa-tion of Nrf2, which however has not yet been investi-gated. Therefore, the present study was performedto investigate the hypothesis that administration of anoxidized oil leads to an activation of Nrf2 pathway in theliver. For this end, we performed an experiment withpigs, an animal model which is closer to human physiol-ogy with respect to xenobiotic metabolism than rodentswhich are commonly used for the investigation of thebiological effects of oxidized fats [24]. In order to reflectthe practical situation of deep frying of foods in humannutrition, we used rapeseed oil–an oil commonly usedfor deep frying of foods–as source of fat which washeated at a temperature of 175°C for 72 h. To detect apotential activation of Nrf2 in the liver, we determinednuclear concentrations of Nrf2 and transcript levels ofvarious Nrf2-regulated genes involved in phase II meta-bolism or antioxidant defense in the liver of pigs. Inorder to investigate whether changes in mRNA concen-trations of Nrf2 target genes are reflected by alteredactivities, we determined activities of two Nrf2 targetgenes, superoxide dismutase (SOD) and thyroxine UDP-glucuronosyltransferase (T4-UGT) in the liver. As aninfluence on the activity of T4-UGT affects the degrada-tion of thyroxine, we also determined the concentrationof that thyroid hormone in plasma of the pigs.
Materials and methodsAnimals and dietsFor the experiment, 24 (12 male, 12 female) six weekold crossbred pigs [(German Landrace × Duroc) × Pie-train] were used. The animals were kept in a pigpencontrolled for temperature (23 ± 2°C), relative humidity(50-60%), and light from 06.00 to 19.00. After one weekof adaptation, the pigs were weighed and randomly
allocated to two groups, each consisting of 6 male and 6female pigs, with similar average body weights. All pigswere housed in pairs in flat-deck pens and received anutritionally adequate diet (Table 1). The diets con-tained 16.4 MJ metabolizable energy and 224 g crudeprotein per kg. The two experimental diets varied in thetype of fat. The diet of the treatment group (“oxidizedfat group”), contained rapeseed oil (obtained from alocal supermarket) which was heated in a domestic fryer(GF-8SE, Bartscher, Salzkotten, Germany) at a tempera-ture of 175°C for 72 h, without addition of any food-stuffs. The diet of the control group (“fresh fat group”)contained a mixture of fresh rapeseed oil and freshpalm oil (91.6:8.4, w/w). This fat mixture was used inorder to equalize the fatty acid composition of the twooils (since the heating process caused a partial loss ofpolyunsaturated fatty acids (PUFA) in the rapeseed oil).Because the frying process caused a loss of tocopherolsin the rapeseed oil, the native concentrations of toco-pherols of all experimental fats were analysed. Based onthe native concentrations of the fats, the vitamin E con-centration of the oxidized fat was adjusted to that of thefresh fat by supplementation with all-rac-a-tocopherylacetate (the biopotency of all-rac-a-tocopheryl acetate isconsidered to be 67% of that of a-tocopherol). The vita-min E concentration in the fresh fat diet was 97.6 mga-tocopherol per kg diet, and that of the oxidized fatdiet was adjusted accordingly.
Table 1 Composition of the experimental diets
Fresh fatdiet(g/kg)
Oxidized fatdiet(g/kg)
Composition
Wheat 181.9 181.9
Barley 100 100
Soy bean meal (44% crude protein) 350 350
Wheat bran 146 146
Fresh rapeseed oil 137.4 -
Palm oil 12.6 -
Oxidized rapeseed oil - 150
Choline chloride (50%) 1.5 1.5
Calcium hydrogen phosphatedihydrate
9 9
Mineral and vitamin premix* 40 40
L-Lysine (50.7%) 16.5 16.5
DL-methionine 3.55 3.55
L-threonine 3.36 3.36
L-tryptophan 0.5 0.5
*The mineral and vitamin premix provided the following per kg diet: 3.6 glysine; 1 g methionine; 1.4 g threonine; 6.8 g calcium; 1.6 g phosphorus;16,000 IE vitamin A; 2,000 IE vitamin D3; 100 mg a-tocopherol acetate; 2 gsodium; 0.8 g magnesium; 120 mg iron; 160 mg copper; 10 mg zinc; 100 mgmanganese; 2 mg iodine; 0.4 mg selenium; 3 mg vitamin B1; 8 mg vitamin B2;8 mg vitamin B6; 60 μg vitamin B12; 40 mg nicotinic acid; 20 mg pantothenicacid; 2 mg folic acid; 160 μg biotin
Varady et al. Lipids in Health and Disease 2012, 11:31http://www.lipidworld.com/content/11/1/31
Page 2 of 9

In order to avoid potential differences in food intake,due to adverse sensoric properties of the oxidized fat, acontrolled feeding system was applied in which pigs wereoffered the same amount of diet during the 29 days feed-ing period. Water was available ad libitum from nippledrinkers during the entire experiment. All experimentalprocedures described followed established guidelines forthe care and handling of laboratory animals and wereapproved by the local Animal Care and Use Committee(Regierungspräsidium Giessen; permission no: GI 19/3No. 49/2010).
Sample collectionAfter completion of the feeding period the animals wereanaesthesised and exsanguinated 2.5 h after their lastmeal. Blood was collected into heparinized polyethylenetubes and plasma was subsequently obtained by centri-fugation of the blood (1100 × g, 10 min, 4°C). Tissuesamples from liver were dissected and stored at -80°Cuntil analysis.
Lipid analysisThe fatty acid composition of the dietary fats was deter-mined by gas chromatography after methylation of fattyacids by trimethylsulfonium hydroxide [25]. The extentof lipid peroxidation of the experimental fats beforeinclusion into the diets was determined by assaying theperoxide value [26], thiobarbituric acid substances(TBARS) [26] and the percentage of total polar com-pounds [27].
RNA isolation and quantitative RT-PCR (qPCR)For the determination of mRNA expression levels totalRNA was isolated from liver tissue using Trizol™ reagent(Invitrogen, Karlsruhe, Germany) according to the manu-facturer’s protocol. Determination of total RNA concen-tration and purity, cDNA synthesis and qPCR analysiswere performed as described recently in detail [28].Gene-specific primer pairs obtained from Eurofins MWGOperon were designed using Primer3 and BLAST.Features of primer pairs are listed in Table 2. For deter-mination of relative expression levels relative quantitieswere calculated using GeNorm normalization factor.In order to calculate the normalization factor, all Ct-values were transformed into relative quantification databy using the 2-ΔCt equation, and the highest relativequantities for each gene were set to 1. From these valuesthe normalization factor was calculated as the geometricmean of expression data of the three most stable out offive tested potential reference genes (Table 2). Referencegene stability across samples was determined by perform-ing GeNorm analysis [29]. After normalization of geneexpression data using the calculated GeNorm normaliza-tion factor, means and SD were calculated from
normalized expression data for samples of the sametreatment group. The mean of the FF group was set to 1and mean and SD of the OF group were scaled propor-tionally. Data on qPCR performance for each gene mea-sured in liver are also shown in Table 2.
Immunoblot analysisNuclear extracts from liver were prepared from six ani-mals per group with a Nuclear Extract Kit (ActiveMotif, Rixensart, Belgium) according to the manufac-turer’s protocol. Protein concentrations in the nuclearextracts were determined by the bicinchoninic acid(BCA) protein assay kit (Interchim, Montluçon, France)with BSA as standard. From each sample of nuclearextract, 30 μg protein were separated on 12.5% SDS-PAGE and electro-transferred to a nitrocellulose mem-brane (Pall, Pensacola, FL, USA). Loading of equalamounts of protein in each line was verified by PonceauS (Carl Roth, Karlsruhe, Germany) staining. After incu-bation the membranes overnight at 4°C in blockingsolution, membranes were incubated with primary anti-bodies against Nrf2 (polyclonal anti-Nrf2 antibody;Abcam, Cambridge, UK), NF-�B/p50 (polyclonal anti-NF-�B/p50 antibody; Santa Cruz, Heidelberg, Germany)and Histone H1 (polyclonal anti-Histone H1 antibody;Active Motif, La Hulpe, Belgium) as a nuclear referenceprotein to control for adequate normalization at roomtemperature. The membranes were washed, and thenincubated with a horseradish peroxidise-conjugated sec-ondary polyclonal anti-rabbit-IgG antibody (Sigma-Aldrich, Steinheim, Germany) for Nrf2, NF-�B/p50 andHistone H1, respectively, at room temperature. After-wards, blots were developed using ECL Advance (GEHealthcare, München, Germany). The signal intensitiesof specific bands were detected with a Bio-Imaging sys-tem (Syngene, Cambridge, UK) and quantified usingSyngene GeneTools software (nonlinear dynamics).
Tocopherol concentrationsConcentrations of tocopherols in experimental diets,liver and plasma were determined by high performance-liquid chromatography (HPLC) with fluorescence detec-tion [30,31]. Tocopherol esters in the samples weresaponified with sodium hydroxide. Tocopherols wereextracted with n-hexane, separated isocratically on aC-18-reversed phase column (Purospher 100 RP-18;Merck-Hitachi, Darmstadt, Germany), using methanolas mobile phase and detected by fluorescence (excitationwavelength: 295 nm, emission wavelength: 325 nm).
SOD activityActivity of hepatic SOD was determined according tothe method of Marklund and Marklund [32]. SOD activ-ity in the homogenates was related to the protein
Varady et al. Lipids in Health and Disease 2012, 11:31http://www.lipidworld.com/content/11/1/31
Page 3 of 9
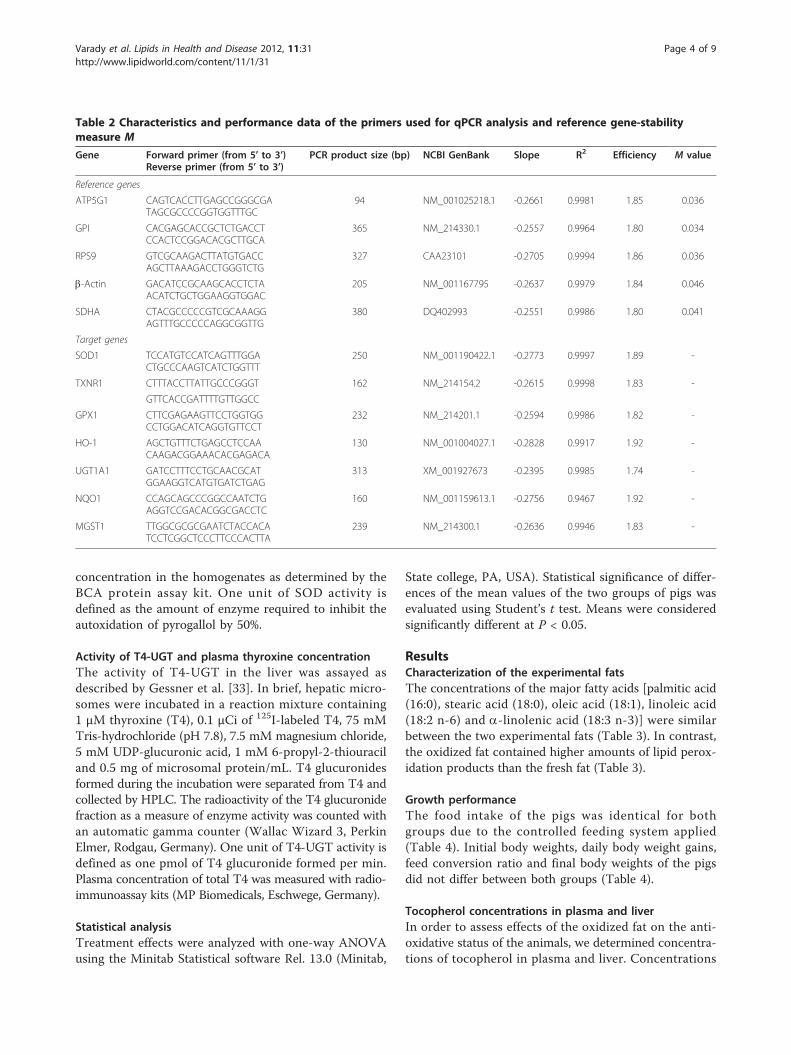
concentration in the homogenates as determined by theBCA protein assay kit. One unit of SOD activity isdefined as the amount of enzyme required to inhibit theautoxidation of pyrogallol by 50%.
Activity of T4-UGT and plasma thyroxine concentrationThe activity of T4-UGT in the liver was assayed asdescribed by Gessner et al. [33]. In brief, hepatic micro-somes were incubated in a reaction mixture containing1 μM thyroxine (T4), 0.1 μCi of 125I-labeled T4, 75 mMTris-hydrochloride (pH 7.8), 7.5 mM magnesium chloride,5 mM UDP-glucuronic acid, 1 mM 6-propyl-2-thiouraciland 0.5 mg of microsomal protein/mL. T4 glucuronidesformed during the incubation were separated from T4 andcollected by HPLC. The radioactivity of the T4 glucuronidefraction as a measure of enzyme activity was counted withan automatic gamma counter (Wallac Wizard 3, PerkinElmer, Rodgau, Germany). One unit of T4-UGT activity isdefined as one pmol of T4 glucuronide formed per min.Plasma concentration of total T4 was measured with radio-immunoassay kits (MP Biomedicals, Eschwege, Germany).
Statistical analysisTreatment effects were analyzed with one-way ANOVAusing the Minitab Statistical software Rel. 13.0 (Minitab,
State college, PA, USA). Statistical significance of differ-ences of the mean values of the two groups of pigs wasevaluated using Student’s t test. Means were consideredsignificantly different at P < 0.05.
ResultsCharacterization of the experimental fatsThe concentrations of the major fatty acids [palmitic acid(16:0), stearic acid (18:0), oleic acid (18:1), linoleic acid(18:2 n-6) and a-linolenic acid (18:3 n-3)] were similarbetween the two experimental fats (Table 3). In contrast,the oxidized fat contained higher amounts of lipid perox-idation products than the fresh fat (Table 3).
Growth performanceThe food intake of the pigs was identical for bothgroups due to the controlled feeding system applied(Table 4). Initial body weights, daily body weight gains,feed conversion ratio and final body weights of the pigsdid not differ between both groups (Table 4).
Tocopherol concentrations in plasma and liverIn order to assess effects of the oxidized fat on the anti-oxidative status of the animals, we determined concentra-tions of tocopherol in plasma and liver. Concentrations
Table 2 Characteristics and performance data of the primers used for qPCR analysis and reference gene-stabilitymeasure M
Gene Forward primer (from 5’ to 3’)Reverse primer (from 5’ to 3’)
PCR product size (bp) NCBI GenBank Slope R2 Efficiency M value
Reference genes
ATP5G1 CAGTCACCTTGAGCCGGGCGATAGCGCCCCGGTGGTTTGC
94 NM_001025218.1 -0.2661 0.9981 1.85 0.036
GPI CACGAGCACCGCTCTGACCTCCACTCCGGACACGCTTGCA
365 NM_214330.1 -0.2557 0.9964 1.80 0.034
RPS9 GTCGCAAGACTTATGTGACCAGCTTAAAGACCTGGGTCTG
327 CAA23101 -0.2705 0.9994 1.86 0.036
b-Actin GACATCCGCAAGCACCTCTAACATCTGCTGGAAGGTGGAC
205 NM_001167795 -0.2637 0.9979 1.84 0.046
SDHA CTACGCCCCCGTCGCAAAGGAGTTTGCCCCCAGGCGGTTG
380 DQ402993 -0.2551 0.9986 1.80 0.041
Target genes
SOD1 TCCATGTCCATCAGTTTGGACTGCCCAAGTCATCTGGTTT
250 NM_001190422.1 -0.2773 0.9997 1.89 -
TXNR1 CTTTACCTTATTGCCCGGGT 162 NM_214154.2 -0.2615 0.9998 1.83 -
GTTCACCGATTTTGTTGGCC
GPX1 CTTCGAGAAGTTCCTGGTGGCCTGGACATCAGGTGTTCCT
232 NM_214201.1 -0.2594 0.9986 1.82 -
HO-1 AGCTGTTTCTGAGCCTCCAACAAGACGGAAACACGAGACA
130 NM_001004027.1 -0.2828 0.9917 1.92 -
UGT1A1 GATCCTTTCCTGCAACGCATGGAAGGTCATGTGATCTGAG
313 XM_001927673 -0.2395 0.9985 1.74 -
NQO1 CCAGCAGCCCGGCCAATCTGAGGTCCGACACGGCGACCTC
160 NM_001159613.1 -0.2756 0.9467 1.92 -
MGST1 TTGGCGCGCGAATCTACCACATCCTCGGCTCCCTTCCCACTTA
239 NM_214300.1 -0.2636 0.9946 1.83 -
Varady et al. Lipids in Health and Disease 2012, 11:31http://www.lipidworld.com/content/11/1/31
Page 4 of 9
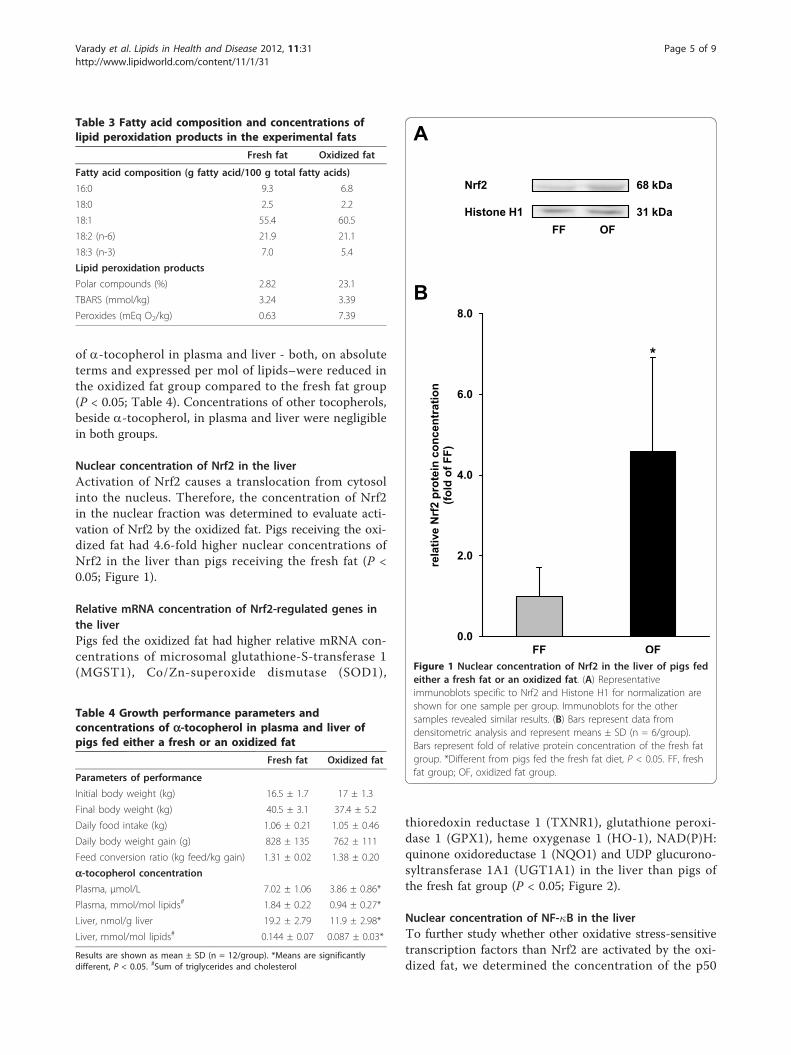
of a-tocopherol in plasma and liver - both, on absoluteterms and expressed per mol of lipids–were reduced inthe oxidized fat group compared to the fresh fat group(P < 0.05; Table 4). Concentrations of other tocopherols,beside a-tocopherol, in plasma and liver were negligiblein both groups.
Nuclear concentration of Nrf2 in the liverActivation of Nrf2 causes a translocation from cytosolinto the nucleus. Therefore, the concentration of Nrf2in the nuclear fraction was determined to evaluate acti-vation of Nrf2 by the oxidized fat. Pigs receiving the oxi-dized fat had 4.6-fold higher nuclear concentrations ofNrf2 in the liver than pigs receiving the fresh fat (P <0.05; Figure 1).
Relative mRNA concentration of Nrf2-regulated genes inthe liverPigs fed the oxidized fat had higher relative mRNA con-centrations of microsomal glutathione-S-transferase 1(MGST1), Co/Zn-superoxide dismutase (SOD1),
thioredoxin reductase 1 (TXNR1), glutathione peroxi-dase 1 (GPX1), heme oxygenase 1 (HO-1), NAD(P)H:quinone oxidoreductase 1 (NQO1) and UDP glucurono-syltransferase 1A1 (UGT1A1) in the liver than pigs ofthe fresh fat group (P < 0.05; Figure 2).
Nuclear concentration of NF-�B in the liverTo further study whether other oxidative stress-sensitivetranscription factors than Nrf2 are activated by the oxi-dized fat, we determined the concentration of the p50
Table 3 Fatty acid composition and concentrations oflipid peroxidation products in the experimental fats
Fresh fat Oxidized fat
Fatty acid composition (g fatty acid/100 g total fatty acids)
16:0 9.3 6.8
18:0 2.5 2.2
18:1 55.4 60.5
18:2 (n-6) 21.9 21.1
18:3 (n-3) 7.0 5.4
Lipid peroxidation products
Polar compounds (%) 2.82 23.1
TBARS (mmol/kg) 3.24 3.39
Peroxides (mEq O2/kg) 0.63 7.39
Table 4 Growth performance parameters andconcentrations of a-tocopherol in plasma and liver ofpigs fed either a fresh or an oxidized fat
Fresh fat Oxidized fat
Parameters of performance
Initial body weight (kg) 16.5 ± 1.7 17 ± 1.3
Final body weight (kg) 40.5 ± 3.1 37.4 ± 5.2
Daily food intake (kg) 1.06 ± 0.21 1.05 ± 0.46
Daily body weight gain (g) 828 ± 135 762 ± 111
Feed conversion ratio (kg feed/kg gain) 1.31 ± 0.02 1.38 ± 0.20
a-tocopherol concentrationPlasma, μmol/L 7.02 ± 1.06 3.86 ± 0.86*
Plasma, mmol/mol lipids# 1.84 ± 0.22 0.94 ± 0.27*
Liver, nmol/g liver 19.2 ± 2.79 11.9 ± 2.98*
Liver, mmol/mol lipids# 0.144 ± 0.07 0.087 ± 0.03*
Results are shown as mean ± SD (n = 12/group). *Means are significantlydifferent, P < 0.05. #Sum of triglycerides and cholesterol
*
0.0
2.0
4.0
6.0
8.0
FF OF
rela
tive
Nrf
2 pr
otei
n co
ncen
trat
ion
(fold
of F
F)
A
B
Nrf2
Histone H1FF OF
68 kDa
31 kDa
Figure 1 Nuclear concentration of Nrf2 in the liver of pigs fedeither a fresh fat or an oxidized fat. (A) Representativeimmunoblots specific to Nrf2 and Histone H1 for normalization areshown for one sample per group. Immunoblots for the othersamples revealed similar results. (B) Bars represent data fromdensitometric analysis and represent means ± SD (n = 6/group).Bars represent fold of relative protein concentration of the fresh fatgroup. *Different from pigs fed the fresh fat diet, P < 0.05. FF, freshfat group; OF, oxidized fat group.
Varady et al. Lipids in Health and Disease 2012, 11:31http://www.lipidworld.com/content/11/1/31
Page 5 of 9
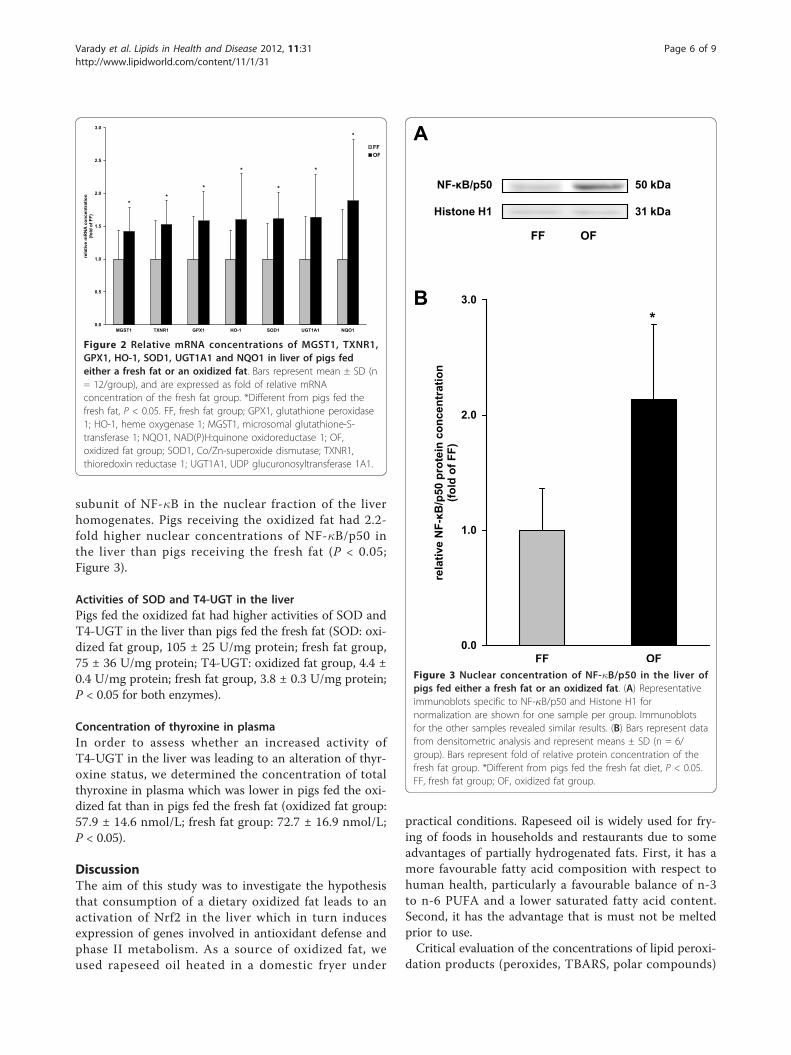
subunit of NF-�B in the nuclear fraction of the liverhomogenates. Pigs receiving the oxidized fat had 2.2-fold higher nuclear concentrations of NF-�B/p50 inthe liver than pigs receiving the fresh fat (P < 0.05;Figure 3).
Activities of SOD and T4-UGT in the liverPigs fed the oxidized fat had higher activities of SOD andT4-UGT in the liver than pigs fed the fresh fat (SOD: oxi-dized fat group, 105 ± 25 U/mg protein; fresh fat group,75 ± 36 U/mg protein; T4-UGT: oxidized fat group, 4.4 ±0.4 U/mg protein; fresh fat group, 3.8 ± 0.3 U/mg protein;P < 0.05 for both enzymes).
Concentration of thyroxine in plasmaIn order to assess whether an increased activity ofT4-UGT in the liver was leading to an alteration of thyr-oxine status, we determined the concentration of totalthyroxine in plasma which was lower in pigs fed the oxi-dized fat than in pigs fed the fresh fat (oxidized fat group:57.9 ± 14.6 nmol/L; fresh fat group: 72.7 ± 16.9 nmol/L;P < 0.05).
DiscussionThe aim of this study was to investigate the hypothesisthat consumption of a dietary oxidized fat leads to anactivation of Nrf2 in the liver which in turn inducesexpression of genes involved in antioxidant defense andphase II metabolism. As a source of oxidized fat, weused rapeseed oil heated in a domestic fryer under
practical conditions. Rapeseed oil is widely used for fry-ing of foods in households and restaurants due to someadvantages of partially hydrogenated fats. First, it has amore favourable fatty acid composition with respect tohuman health, particularly a favourable balance of n-3to n-6 PUFA and a lower saturated fatty acid content.Second, it has the advantage that is must not be meltedprior to use.Critical evaluation of the concentrations of lipid peroxi-
dation products (peroxides, TBARS, polar compounds)
**
*
*
*
*
*
0.0
0.5
1.0
1.5
2.0
2.5
3.0
MGST1 TXNR1 GPX1 HO-1 SOD1 UGT1A1 NQO1
rela
tive
mR
NA
con
cent
ratio
n(fo
ld o
f FF)
FFOF
Figure 2 Relative mRNA concentrations of MGST1, TXNR1,GPX1, HO-1, SOD1, UGT1A1 and NQO1 in liver of pigs fedeither a fresh fat or an oxidized fat. Bars represent mean ± SD (n= 12/group), and are expressed as fold of relative mRNAconcentration of the fresh fat group. *Different from pigs fed thefresh fat, P < 0.05. FF, fresh fat group; GPX1, glutathione peroxidase1; HO-1, heme oxygenase 1; MGST1, microsomal glutathione-S-transferase 1; NQO1, NAD(P)H:quinone oxidoreductase 1; OF,oxidized fat group; SOD1, Co/Zn-superoxide dismutase; TXNR1,thioredoxin reductase 1; UGT1A1, UDP glucuronosyltransferase 1A1.
*
0.0
1.0
2.0
3.0
FF OF
rela
tive
NF-
/p50
pro
tein
con
cent
ratio
n(fo
ld o
f FF)
A
B
NF- B/p50
Histone H1
FF OF
50 kDa
31 kDa
Figure 3 Nuclear concentration of NF-�B/p50 in the liver ofpigs fed either a fresh fat or an oxidized fat. (A) Representativeimmunoblots specific to NF-�B/p50 and Histone H1 fornormalization are shown for one sample per group. Immunoblotsfor the other samples revealed similar results. (B) Bars represent datafrom densitometric analysis and represent means ± SD (n = 6/group). Bars represent fold of relative protein concentration of thefresh fat group. *Different from pigs fed the fresh fat diet, P < 0.05.FF, fresh fat group; OF, oxidized fat group.
Varady et al. Lipids in Health and Disease 2012, 11:31http://www.lipidworld.com/content/11/1/31
Page 6 of 9

indicates that oxidation of PUFA in the oil during heatingoccurred. Nevertheless, concentrations of these lipid per-oxidation products were lower than in most other studiesdealing with the effects of oxidized fats [34-36]. Althoughthe parameters determined to assess the degree of oxida-tion such as TBARS are not very specific, the oil used inthis study can be regarded as moderately oxidized. Onthe base of analyses of a large number of used frying fats,it has been found that heated fats are acceptable untilthey reach a level of 25% polar compounds [37]. Thus,the oxidized fat used in this study, containing 23% ofpolar compounds, would be just acceptable for frying offoods.Heating an oil leads to a loss of its PUFA and its
native antioxidants such as tocopherols [2,38]. In orderto avoid secondary effects on metabolism due to differ-ences in the intake of PUFA and antioxidants betweenthe two groups, we equalized the control fat and theoxidized fat for their fatty acid compositions and theirvitamin E concentrations. The vitamin E concentrationsof the diets, being around 90 mg a-tocopherol equiva-lents per kg diet, were in excess of the vitamin Erequirement which might be around 40 mg a-toco-pherol equivalents per kg for the specific diets used inthis study (according to calculations of the vitamin Erequirement as a function of amount and type of PUFAas suggested by Muggli [39]).In most studies dealing with the effects of thermoxi-
dized fats, feeding the diet containing the oxidized fatlowered body weight gains in the experimental animalsdue to a diminished feed intake, a reduced digestibility ofnutrients and general toxic effects [34,35,40-42]. In thepresent study, feeding the diet containing the oxidized fatdid not influence the growth of the pigs. This might bedue to the facts that feed intake was equalized betweenthe two groups of pigs by a controlled feeding systemand that the heated oil was only moderately oxidized.The finding that the body weight development was notaffected by the oxidized fat is advantageous from a meth-odological viewpoint because the effects of the oxidizedfat were not confounded by secondary effects of reducedgrowth. Recent studies have shown that consumption ofoxidized fats leads to a reduction of tocopherol concen-trations in animal tissues due to a reduced digestibilityand an enhanced turnover of vitamin E [8,11,12,43]. Thefinding of reduced a-tocopherol concentrations inplasma and liver of pigs fed the oxidized fat in the pre-sent study indicates that even moderately oxidized fatscompromise tissue vitamin E status in animals.According to the hypothesis of this study, we observed
for the first time that administration of an oxidized fatleads to an activation of the transcription factor Nrf2 inthe liver. Activation of Nrf2 was evident by an increasedconcentration of nuclear Nrf2 and increased transcript
levels of several Nrf2 target genes involved in the anti-oxidant defense system (SOD1, GPX1, TXNR1, HO-1)and phase II metabolism (NQO1, MGST1, UGT1A1).Under normal conditions, Nrf2 is located in the cytosolof cells where it is constantly degraded via the proteaso-mal system. In response to various stimuli like oxidativestress, Keap1–a cytosolic inhibitory protein–dissociatesfrom Nrf2 and Nrf2 translocates into the nucleus, whereit binds to specific DNA-sequences called antioxidantresponse element (ARE) in the regulatory region of tar-get genes [44]. Regarding that Nrf2 is activated by ROS[45,46], the occurrence of oxidative stress commonlyobserved in animals fed oxidized fats might be one pos-sible explanation for activation of Nrf2 in pigs fed theoxidized fat. This possibility is strengthened by theobservation that the up-regulation of enzymes involvedin xenobiotic metabolism and stress response due tofeeding of oxidized fats was attenuated by concomitantsupplementation of vitamin C or vitamin E in guineapigs and rats, resp [15,16]. Recently, it has been foundthat n-3 fatty acid oxidation products deriving fromeicosapentaenoic or docosahexaenoic acid are able todirectly activate Nrf2 by initiating dissociation of Keap1[22]. Therefore, the possibility that Nrf2 activation wasdirectly induced by specific oxidation products presentin the oxidized oil cannot be excluded.Some of the genes determined in this study, such as
HO-1, GPX1 and SOD1, also contain binding sites forother oxidative stress-sensitive transcription factors, suchas NF-�B, in their regulatory region [47-49]. Therefore, itis likely that the up-regulation of those genes was at leastin part mediated by activation of NF-�B observed in thepigs fed the oxidized fat, as indicated by an increasednuclear concentration of p50. Thus, it is very likely that acoordinated activation of multiple oxidative stress-sensi-tive signaling pathways is responsible for the observedup-regulation of antioxidant, cytoprotective and detoxify-ing genes in the liver of pigs fed the oxidized fat diet, par-ticularly because ROS, whose formation is induced by theadministration of oxidized fats, are common stimuli ofthese pathways.A further interesting observation of this study is that
hepatic activity of T4-UGT, an enzyme which belongs tothe phase II system, in the liver was increased in pigs fedthe oxidized fat. T4-UGT catalyses the glucuronidationof thyroxine, and thus is the key enzyme of its elimina-tion in the liver [50]. The finding of an increased activityof that enzyme concurs with a reduced concentration oftotal thyroxine in plasma of pigs fed the oxidized fat sug-gesting that hepatic elimination of that hormone wasenhanced in those pigs. Thus, the present study indicatesthat oxidized fats could affect thyroid hormone status viaan induction of the thyroid hormone elimination processin the liver such as observed for several other compounds
Varady et al. Lipids in Health and Disease 2012, 11:31http://www.lipidworld.com/content/11/1/31
Page 7 of 9

which are acting as inducers of the hepatic xenobioticsystem [51-53]. It is well known that hepatic phase I andphase II enzymes play an important role in eliminationand detoxification of many drugs [54,55]. Thus, it is likelythat ingestion of oxidized fats enhances the eliminationof medical drugs, due to an induction of the hepaticxenobiotic system.In conclusion, the present study shows for the first
time that the ingestion of a moderately oxidized oilcauses an activation of Nrf2 in the liver of pigs. Thisprobably provides an explanation for the concomitantup-regulation of hepatic genes involved in the antioxi-dant defense system and phase II metabolism as observedherein in pigs and in recent studies in rats and guineapigs [14-19]. Induction of Nrf2 in the liver of pigs fed anoxidized fat can be interpreted as an adaptive response ofthe liver to cope with oxidative stress induced by admin-istration of oxidized fats, thereby, preventing ROS-mediated damage. Activation of Nrf2 is generallyregarded as a beneficial effect as it causes an up-regula-tion of a wide spectrum of antioxidant, cytoprotectiveand detoxifying genes and thus protects the cell againstROS and toxic compounds. Indeed, the coordinated up-regulation of these genes by Nrf2 activators is consideredas a potential therapeutic strategy to protect againstinsults such as inflammation and oxidative stress inducedin various chronic diseases [56]. Nevertheless, the resultsof the present study must not be interpreted in the waythat oxidized fats can be regarded as health-promotingcomponents of the diet, as components of oxidized fatsmight have several adverse effects in human subjects.The results of this study rather suggest that oxidized fatsare a mixture of chemically distinct substances, some ofwhich exhibit a significant biological activity.
AbbreviationsFF: Fresh fat; GPX1: Glutathione peroxidase 1; HO-1: Heme oxygenase 1;Keap1: Kelch-like ECH-associated protein 1; MGST1: Microsomal glutathione-S-transferase 1; NQO1: NAD(P)H: Quinone oxidoreductase 1; OF: Oxidized fat;PUFA: Polyunsaturated fatty acid; SOD1: Co/Zn-superoxide dismutase; TBARS:Thiobarbituric acid substances; T4-UGT: Thyroxine UDP-glucuronosyltransferase; TXNR1: Thioredoxin reductase 1; UGT1A1: UDPGlucuronosyltransferase 1A1.
AcknowledgementsThis study was supported by a grant from the DeutscheForschungsgemeinschaft (DFG; grant no. RI 1537/1-1).
Authors’ contributionsJV carried out the experiments and participated in the interpretation of thedata and drafted the manuscript. DKG and EM participated in analysis. KEand RR conceived of the study and its design, coordinated work,participated in the interpretation of the results, and helped to draft themanuscript. All authors read and approved the final manuscript.
Competing interestsThe authors declare that they have no competing interests.
Received: 11 January 2012 Accepted: 24 February 2012Published: 24 February 2012
References1. Guthrie JF, Lin BH, Frazao E: Role of food prepared away from home in
the American diet, 1977-78 versus 1994-96: changes and consequences.J Nutr Educ Behav 2002, 34:140-150.
2. Choe E, Min DB: Chemistry of deep-fat frying oils. J Food Sci 2007,72:77-86.
3. Cohn JS: Oxidized fat in the diet, postprandial lipaemia andcardiovascular disease. Curr Opin Lipidol 2002, 13:19-24.
4. Staprans I, Pan XM, Rapp JH, Feingold KR: The role of dietary oxidizedcholesterol and oxidized fatty acids in the development ofatherosclerosis. Mol Nutr Food Res 2005, 49:1075-1082.
5. Ringseis R, Eder K: Regulation of genes involved in lipid metabolism bydietary oxidized fat. Mol Nutr Food Res 2011, 1:109-121.
6. Staprans I, Rapp JH, Pan XM, Kim KY, Feingold KR: Oxidized lipids in thediet are a source of oxidized lipid in chylomicrons of human serum.Arterioscler Thromb 1994, 14:1900-1905.
7. Hayam I, Cogan U, Mokady S: Dietary oxidized oil and the activity ofantioxidant enzymes and lipoprotein oxidation in rats. Nutr Res 1995,15:1037-1044.
8. Keller U, Brandsch C, Eder K: Supplementation of vitamins C and Eincreases the vitamin E status but does not prevent the formation ofoxysterols in the liver of guinea pigs fed an oxidised fat. Eur J Nutr 2004,43:353-359.
9. Izaki Y, Yoshikawa S, Uchiyama M: Effect of ingestion of thermallyoxidized frying oil on peroxidative criteria in rats. Lipids 1984, 19:324-331.
10. Kok TS, Harris PG, Alexander JC: Heated canola oil and oxidative stress inrats. Nutr Res 1988, 8:673-684.
11. Liu JF, Huang CJ: Tissue alpha-tocopherol retention in male rats iscompromised by feeding diets containing oxidized frying oil. J Nutr1995, 125:3071-3080.
12. Liu JF, Huang CJ: Dietary oxidized frying oil enhances tissue alpha-tocopherol depletion and radioisotope tracer excretion in vitamin E-deficient rats. J Nutr 1996, 126:2227-2235.
13. Eder K, Keller U, Hirche F, Brandsch C: Thermally oxidized dietary fatsincrease the susceptibility of rat LDL to lipid peroxidation but not theiruptake by macrophages. J Nutr 2003, 133:2830-2837.
14. Huang CJ, Cheung NS, Lu VR: Effects of deteriorated frying oil and dietaryprotein levels on liver microsomal enzymes in rats. J Am Oil Chem Soc1988, 65:1796-1803.
15. Liu JF, Chan FC: Forms of cytochrome P450 in the liver microsome ofoxidized frying oil-fed guinea pigs. J Nutr Sci Vitaminol 2000, 46:240-245.
16. Sülzle A, Hirche F, Eder K: Thermally oxidized dietary fat upregulates theexpression of target genes of PPARα in rat liver. J Nutr 2004,134:1375-1383.
17. Chen YY, Chen CM, Chao PY, Chang TJ, Liu JF: Effects of frying oil andHouttuynia cordata thunb on xenobiotic-metabolizing enzyme systemof rodents. World J Gastroenterol 2005, 11:389-392.
18. Liu JF, Lee YW, Chang FC: Effect of oxidized frying oil and vitamin Clevels on the hepatic xenobiotic-metabolizing enzyme system of guineapigs. J Nutr Sci Vitaminol 2000, 46:137-140.
19. Kwak MK, Wakabayashi N, Itoh K, Motohashi H, Yamamoto M, Kensler TW:Modulation of gene expression by cancer chemopreventivedithiolethiones through the Keap1-Nrf2 pathway. Identification of novelgene clusters for cell survival. J Biol Chem 2003, 278:8135-8145.
20. Jaiswal AK: Nrf2 signaling in coordinated activation of antioxidant geneexpression. Free Radic Biol Med 2004, 36:1199-1207.
21. Copple IM, Goldring CE, Kitteringham NR, Park BK: The Nrf2-Keap1 defencepathway: role in protection against drug-induced toxicity. Toxicol 2008,246:24-33.
22. Gao L, Wang J, Sekhar KR, Yin H, Yared NF, Schneider SN, Sasi S, Dalton TP,Anderson ME, Chan JY, Morrow JD, Freeman ML: Novel n-3 fatty acidoxidation products activate Nrf2 by destabilizing the associationbetween Keap1 and Cullin3. J Biol Chem 2007, 282:2529-2537.
23. Varady J, Eder K, Ringseis R: Dietary oxidized fat activates the oxidativestress-responsive transcription factors NF-κB and Nrf2 in intestinalmucosa of mice. Eur J Nutr 2011, 50:601-609.
Varady et al. Lipids in Health and Disease 2012, 11:31http://www.lipidworld.com/content/11/1/31
Page 8 of 9

24. Puccinelli E, Gervasi PG, Longo V: Xenobiotic metabolizing cytochromeP450 in pig, a promising animal model. Curr Drug Metab 2011,12:507-525.
25. Butte W: Rapid method for the determination of fatty acid profiles fromfats and oils using trimethylsulfonium hydroxide for transesterification. JChromatogr 1983, 261:142-145.
26. Deutsche Gesellschaft für Fettwissenschaft: Einheitsmethoden zurUntersuchung von Fetten, Fettprodukten, Tensiden und verwandten StoffenStuttgart: Wissenschaftliche Verlagsgesellschaft; 1994.
27. International Union of Pure and Applied Chemistry (IUPAC): Determinationof polar compounds, polymerized and oxidized triacylglycerols, anddiacylglycerols in oils and fats. Pure Appl Chem 2000, 72:1563-1575.
28. Keller J, Ringseis R, Koc A, Lukas I, Kluge H, Eder K: Supplementation withL-carnitine downregulates genes of the ubiquitin proteasome system inthe skeletal muscle and liver of piglets. Animal 2011, 6:70-78.
29. Vandesompele J, De Preter K, Pattyn F, Poppe B, Van Roy N, De Paepe A,Speleman F: Accurate normalization of real-time quantitative RT-PCRdata by geometric averaging of multiple internal control genes. GenomeBiol 2002, 3:RESEARCH0034.
30. Catignani GL, Bieri JG: Simultaneous determination of retinol and alpha-tocopherol in serum or plasma by liquid chromatography. Clin Chem1983, 29:708-712.
31. Cort WMS, Vicente TS, Waysek EH, Williams BD: Vitamin E content offeedstuffs determined by high-performance liquid chromatographicfluorescence. J Agric Food Chem 1983, 31:1330-1333.
32. Marklund S, Marklund G: Involvement of the superoxide anion radical inthe autoxidation of pyrogallol and a convenient assay for superoxidedismutase. Eur J Biochem 1974, 47:469-474.
33. Gessner DK, Ringseis R, Möller C, Eder K: Increased plasma thyroidhormone concentrations in LDL receptor deficient mice may beexplained by inhibition of aryl hydrocarbon receptor-dependentexpression of hepatic UDP-glucuronosyltransferases. Biochim Biophys Acta.
34. Yoshida H, Kajimoto G: Effect of dietary vitamin E on the toxicity ofautoxidized oil to rats. Ann Nutr Metab 1989, 33:153-161.
35. Corcos Benedetti P, Di Felice M, Gentili V, Tagliamonte B, Tomassi G:Influence of dietary thermally oxidized soybean oil on the oxidativestatus of rats of different ages. Ann Nutr Metab 1990, 34:221-231.
36. Ammouche A, Rouaki F, Bitam A, Bellal MM: Effect of ingestion ofthermally oxidized sunflower oil on the fatty acid composition andantioxidant enzymes of rat liver and brain in development. Ann NutrMetab 2002, 46:268-275.
37. Frankel EN: Lipid oxidation Dundee, UK: The Oily Press; 1998.38. Eder K, Kirchgessner M: The effect of dietary vitamin E supply and a
moderately oxidized oil on activities of hepatic lipogenic enzymes inrats. Lipids 1998, 33:277-283.
39. Muggli R: Physiological requirements of vitamin E as a function of theamount and type of polyunsaturated fatty acid. World Rev Nutr Diet 1994,75:166-168.
40. Blanc P, Revol A, Pacheco H: Chronical ingestion of oxidized oil in the rat:effect on lipid composition and cytidilyl transferase activity in varioustissues. Nutr Res 1992, 12:833-844.
41. Hochgraf E, Mokady S, Cogan U: Dietary oxidized linoleic acid modifieslipid composition of rat liver microsomes and increases their fluidity. JNutr 1997, 127:681-686.
42. Engberg RM, Lauridsen C, Jensen SK, Jakobsen K: Inclusion of oxidizedvegetable oil in broiler diets. Its influence on nutrient balance and onthe antioxidative status of broilers. Poult Sci 1996, 75:1003-1011.
43. Eder K, Stangl GI: Plasma thyroxine and cholesterol concentrations ofminiature pigs are influenced by thermally oxidized dietary lipids. J Nutr2000, 130:116-121.
44. Dhakshinamoorthy S, Long DJ, Jaiswal AK: Antioxidant regulation of genesencoding enzymes that detoxify xenobiotics and carcinogens. Curr TopCell Regul 2000, 36:201-216.
45. Kaspar JW, Niture SK, Jaiswal AK: Nrf2:INrf2 (Keap1) signaling in oxidativestress. Free Radic Biol Med 2009, 47:1304-1309.
46. Niture SK, Kaspar JW, Shen J, Jaiswal AK: Nrf2 signaling and cell survival.Toxicol Appl Pharmacol 2010, 244:37-42.
47. Lavrovsky Y, Schwartzman ML, Levere RD, Kappas A, Abraham NG:Identification of binding sites for transcription factors NF-kappa B and
AP-2 in the promoter region of the human heme oxygenase 1 gene.Proc Natl Acad Sci USA 1994, 91:5987-5991.
48. Zhou LZ, Johnson AP, Rando TA: NF-κB and AP-1 mediate transcriptionalresponses to oxidative stress in skeletal muscle cells. Free Radic Biol Med2001, 31:1405-1416.
49. Maehara K, Hasegawa T, Isobe KI: A NF-κB p65 subunit is indispensablefor activating manganese superoxide: dismutase gene transcriptionmediated by tumor necrosis factor-α. J Cell Biochem 2000, 77:474-486.
50. Beetstra JB, van Engelen JG, Karels P, van der Hoek HJ, de Jong M,Docter R, Krenning EP, Hennemann G, Brouwer A, Visser TJ: Thyroxine and3,3’,5-triiodothyronine are glucuronidated in rat liver by different uridinediphosphate-glucuronyltransferases. Endocrinol 1991, 128:741-746.
51. Luci S, Kluge H, Hirche F, Eder K: Clofibrate increases hepatictriiodothyronine (T3)- and thyroxine (T4)-glucuronosyltransferaseactivities and lowers plasma T3 and T4 concentrations in pigs. DrugMetab Dispos 2006, 34:1887-1892.
52. Vansell NR, Klaassen CD: Effect of microsomal enzyme inducers on thebiliary excretion of triiodothyronine (T(3)) and its metabolites. Toxicol Sci2002, 65:184-191.
53. Hood A, Allen ML, Liu Y, Liu J, Klaassen CD: Induction of T(4) UDP-GTactivity, serum thyroid stimulating hormone, and thyroid follicular cellproliferation in mice treated with microsomal enzyme inducers. ToxicolAppl Pharmacol 2003, 188:6-13.
54. Xu C, Li CY, Kong AN: Induction of phase I, II and III drug metabolism/transport by xenobiotics. Arch Pharm Res 2005, 28:249-268.
55. Wang H, LeCluyse EL: Role of orphan nuclear receptors in the regulationof drug-metabolising enzymes. Clin Pharmacokinet 2003, 42:1331-1357.
56. de Haan JB: Nrf2 activators as attractive therapeutics for diabeticnephropathy. Diabetes 2011, 60:2683-2684.
doi:10.1186/1476-511X-11-31Cite this article as: Varady et al.: Dietary moderately oxidized oilactivates the Nrf2 signaling pathway in the liver of pigs. Lipids in Healthand Disease 2012 11:31.
Submit your next manuscript to BioMed Centraland take full advantage of:
• Convenient online submission
• Thorough peer review
• No space constraints or color figure charges
• Immediate publication on acceptance
• Inclusion in PubMed, CAS, Scopus and Google Scholar
• Research which is freely available for redistribution
Submit your manuscript at www.biomedcentral.com/submit
Varady et al. Lipids in Health and Disease 2012, 11:31http://www.lipidworld.com/content/11/1/31
Page 9 of 9

SHORT REPORT Open Access
Dietary moderately oxidized oil inducesexpression of fibroblast growth factor21 in the liver of pigsJuliane Varady, Robert Ringseis* and Klaus Eder
Abstract
Background: Fibroblast growth factor 21 (FGF21), whose expression is induced by peroxisome proliferator-activated receptor a (PPARa), has been recently identified as a novel metabolic regulator which plays a crucial rolein glucose homeostasis, lipid metabolism, insulin sensitivity and obesity. Previous studies have shown thatadministration of oxidized fats leads to an activation of PPARa in the liver. Therefore, the present studyinvestigated the hypothesis that feeding of oxidized fats causes an induction of FGF21 in the liver.
Methods: Twenty four crossbred pigs were allocated to two groups of 12 pigs each and fed nutritionally adequatediets with either fresh rapeseed oil or oxidized rapeseed oil prepared by heating at a temperature of 175°C for 72h.
Results: In pigs fed the oxidized fat mRNA abundance and protein concentrations of FGF21 in liver weresignificantly increased (P < 0.05), and the protein concentrations of FGF21 in plasma tended to be increased (P <0.1) in comparison to control pigs. Moreover, pigs fed the oxidized fat had increased transcript levels of the PPARatarget genes acyl-CoA oxidase, carnitine palmitoyltransferase-1 and novel organic cation transporter 2 in the liver (P< 0.05), indicative of PPARa activation.
Conclusion: The present study shows for the first time that administration of an oxidized fat induces theexpression of FGF21 in the liver, probably mediated by activation of PPARa. Induction of FGF21 could be involvedin several effects observed in animals administered an oxidized fat.
Keywords: Fibroblast growth factor 21, Liver, Peroxisome proliferator-activated receptor α, Pig, Oxidized fat
BackgroundFibroblast growth factor 21 (FGF21) has been recentlyidentified as a novel metabolic regulator which plays acrucial role in glucose homeostasis, lipid metabolism,insulin sensitivity and obesity [1-3]. The physiologicimportance of FGF21 is evident from studies showingthat systemic administration of FGF21 reduces serumand liver triacylglycerol (TAG) concentrations, bodyweight and obesity in obese mice [4-6]. Hepatic expres-sion of FGF21 is strongly up-regulated during fastingand is rapidly suppressed by refeeding, indicating a roleof FGF21 in fasting-induced response [1-3]. In the liver,FGF21 stimulates hepatic lipid oxidation, ketogenesis
and gluconeogenesis [1]. Recently, it has been foundthat expression of FGF21 is induced by peroxisome pro-liferator-activated receptor a (PPARa), a transcriptionfactor which is activated during fasting by non-esterifiedfatty acids released from adipose tissue or by adminis-tration of synthetic or natural PPARa ligands such asfibrates or various types of fatty acids [1,3,7,8].Oxidized lipids as components of heated or fried foods
play an important role in nutrition in industrializedcountries [9]. The ingestion of oxidized fats is generallyregarded as detrimental for health. However, some stu-dies in rodents have shown that oxidized fats could alsoexert some beneficial effects on metabolism such as areduction of plasma and liver TAG and cholesterol con-centrations, a reduction of obesity and even an inhibi-tion of atherosclerosis [10-16]. Moreover, it has been
* Correspondence: [email protected] of Animal Nutrition and Nutrition Physiology, Justus-Liebig-University Giessen, Giessen, Germany
Varady et al. Lipids in Health and Disease 2012, 11:34http://www.lipidworld.com/content/11/1/34
© 2012 Varady et al; licensee BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative CommonsAttribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction inany medium, provided the original work is properly cited.
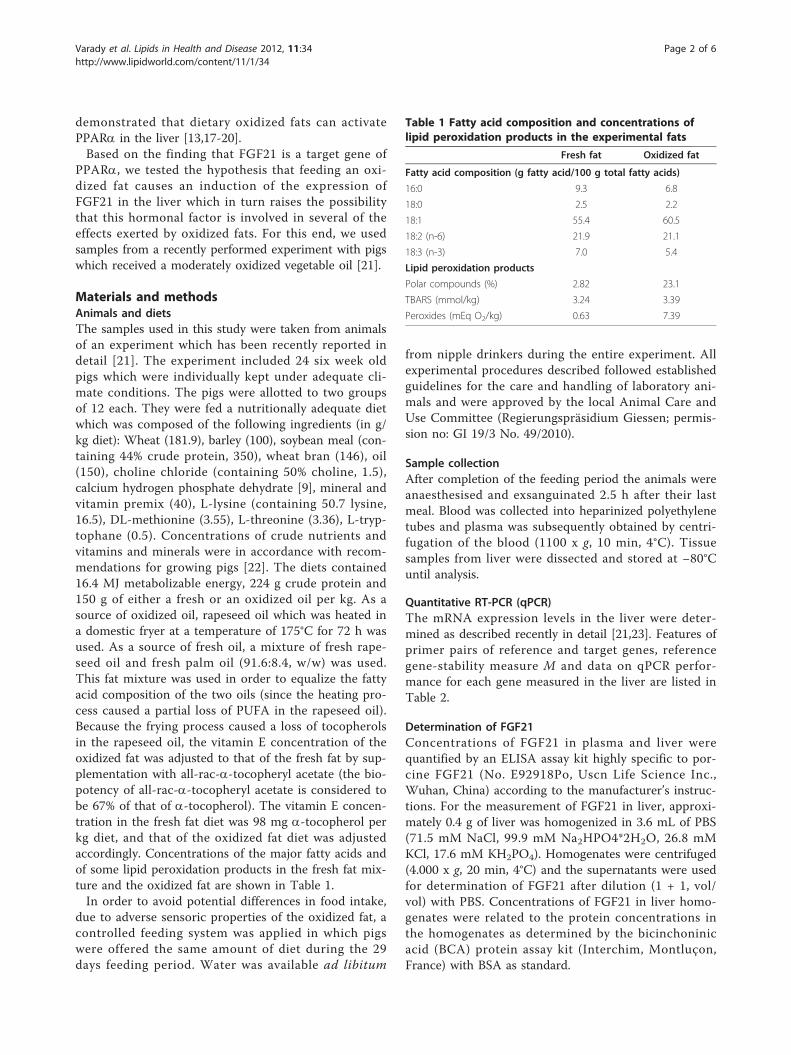
demonstrated that dietary oxidized fats can activatePPARa in the liver [13,17-20].Based on the finding that FGF21 is a target gene of
PPARa, we tested the hypothesis that feeding an oxi-dized fat causes an induction of the expression ofFGF21 in the liver which in turn raises the possibilitythat this hormonal factor is involved in several of theeffects exerted by oxidized fats. For this end, we usedsamples from a recently performed experiment with pigswhich received a moderately oxidized vegetable oil [21].
Materials and methodsAnimals and dietsThe samples used in this study were taken from animalsof an experiment which has been recently reported indetail [21]. The experiment included 24 six week oldpigs which were individually kept under adequate cli-mate conditions. The pigs were allotted to two groupsof 12 each. They were fed a nutritionally adequate dietwhich was composed of the following ingredients (in g/kg diet): Wheat (181.9), barley (100), soybean meal (con-taining 44% crude protein, 350), wheat bran (146), oil(150), choline chloride (containing 50% choline, 1.5),calcium hydrogen phosphate dehydrate [9], mineral andvitamin premix (40), L-lysine (containing 50.7 lysine,16.5), DL-methionine (3.55), L-threonine (3.36), L-tryp-tophane (0.5). Concentrations of crude nutrients andvitamins and minerals were in accordance with recom-mendations for growing pigs [22]. The diets contained16.4 MJ metabolizable energy, 224 g crude protein and150 g of either a fresh or an oxidized oil per kg. As asource of oxidized oil, rapeseed oil which was heated ina domestic fryer at a temperature of 175°C for 72 h wasused. As a source of fresh oil, a mixture of fresh rape-seed oil and fresh palm oil (91.6:8.4, w/w) was used.This fat mixture was used in order to equalize the fattyacid composition of the two oils (since the heating pro-cess caused a partial loss of PUFA in the rapeseed oil).Because the frying process caused a loss of tocopherolsin the rapeseed oil, the vitamin E concentration of theoxidized fat was adjusted to that of the fresh fat by sup-plementation with all-rac-a-tocopheryl acetate (the bio-potency of all-rac-a-tocopheryl acetate is considered tobe 67% of that of a-tocopherol). The vitamin E concen-tration in the fresh fat diet was 98 mg a-tocopherol perkg diet, and that of the oxidized fat diet was adjustedaccordingly. Concentrations of the major fatty acids andof some lipid peroxidation products in the fresh fat mix-ture and the oxidized fat are shown in Table 1.In order to avoid potential differences in food intake,
due to adverse sensoric properties of the oxidized fat, acontrolled feeding system was applied in which pigswere offered the same amount of diet during the 29days feeding period. Water was available ad libitum
from nipple drinkers during the entire experiment. Allexperimental procedures described followed establishedguidelines for the care and handling of laboratory ani-mals and were approved by the local Animal Care andUse Committee (Regierungspräsidium Giessen; permis-sion no: GI 19/3 No. 49/2010).
Sample collectionAfter completion of the feeding period the animals wereanaesthesised and exsanguinated 2.5 h after their lastmeal. Blood was collected into heparinized polyethylenetubes and plasma was subsequently obtained by centri-fugation of the blood (1100 x g, 10 min, 4°C). Tissuesamples from liver were dissected and stored at −80°Cuntil analysis.
Quantitative RT-PCR (qPCR)The mRNA expression levels in the liver were deter-mined as described recently in detail [21,23]. Features ofprimer pairs of reference and target genes, referencegene-stability measure M and data on qPCR perfor-mance for each gene measured in the liver are listed inTable 2.
Determination of FGF21Concentrations of FGF21 in plasma and liver werequantified by an ELISA assay kit highly specific to por-cine FGF21 (No. E92918Po, Uscn Life Science Inc.,Wuhan, China) according to the manufacturer’s instruc-tions. For the measurement of FGF21 in liver, approxi-mately 0.4 g of liver was homogenized in 3.6 mL of PBS(71.5 mM NaCl, 99.9 mM Na2HPO4*2H2O, 26.8 mMKCl, 17.6 mM KH2PO4). Homogenates were centrifuged(4.000 x g, 20 min, 4°C) and the supernatants were usedfor determination of FGF21 after dilution (1 + 1, vol/vol) with PBS. Concentrations of FGF21 in liver homo-genates were related to the protein concentrations inthe homogenates as determined by the bicinchoninicacid (BCA) protein assay kit (Interchim, Montluçon,France) with BSA as standard.
Table 1 Fatty acid composition and concentrations oflipid peroxidation products in the experimental fats
Fresh fat Oxidized fat
Fatty acid composition (g fatty acid/100 g total fatty acids)
16:0 9.3 6.8
18:0 2.5 2.2
18:1 55.4 60.5
18:2 (n-6) 21.9 21.1
18:3 (n-3) 7.0 5.4
Lipid peroxidation products
Polar compounds (%) 2.82 23.1
TBARS (mmol/kg) 3.24 3.39
Peroxides (mEq O2/kg) 0.63 7.39
Varady et al. Lipids in Health and Disease 2012, 11:34http://www.lipidworld.com/content/11/1/34
Page 2 of 6
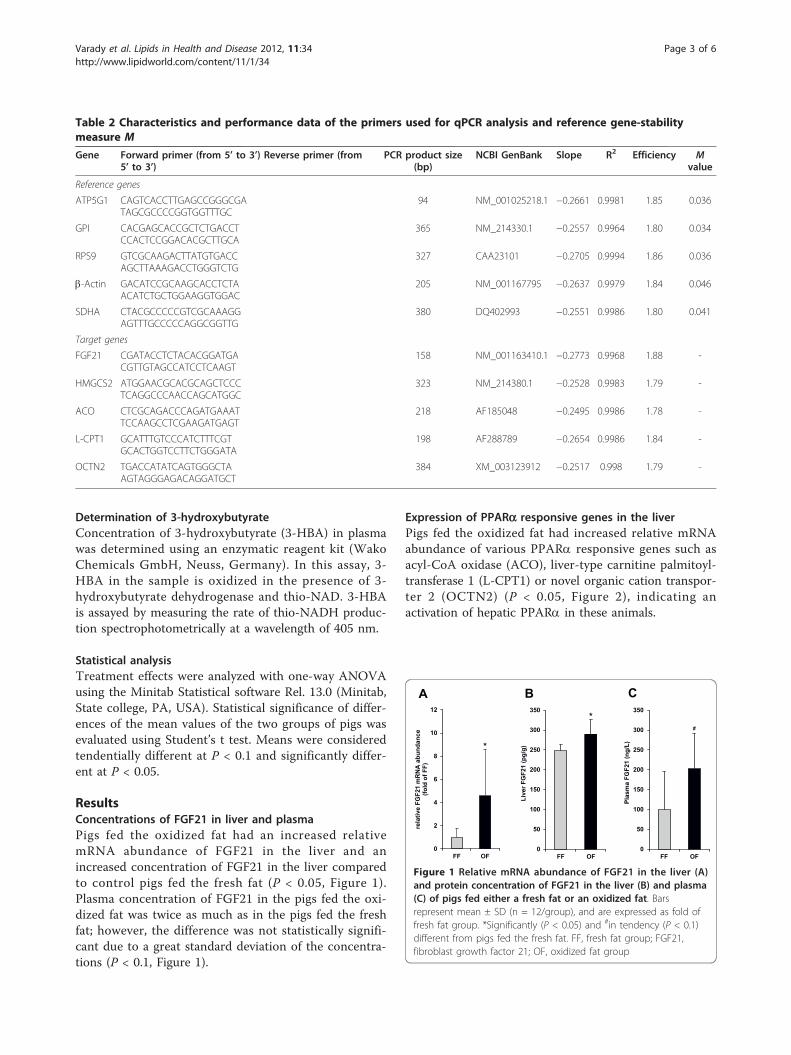
Determination of 3-hydroxybutyrateConcentration of 3-hydroxybutyrate (3-HBA) in plasmawas determined using an enzymatic reagent kit (WakoChemicals GmbH, Neuss, Germany). In this assay, 3-HBA in the sample is oxidized in the presence of 3-hydroxybutyrate dehydrogenase and thio-NAD. 3-HBAis assayed by measuring the rate of thio-NADH produc-tion spectrophotometrically at a wavelength of 405 nm.
Statistical analysisTreatment effects were analyzed with one-way ANOVAusing the Minitab Statistical software Rel. 13.0 (Minitab,State college, PA, USA). Statistical significance of differ-ences of the mean values of the two groups of pigs wasevaluated using Student’s t test. Means were consideredtendentially different at P < 0.1 and significantly differ-ent at P < 0.05.
ResultsConcentrations of FGF21 in liver and plasmaPigs fed the oxidized fat had an increased relativemRNA abundance of FGF21 in the liver and anincreased concentration of FGF21 in the liver comparedto control pigs fed the fresh fat (P < 0.05, Figure 1).Plasma concentration of FGF21 in the pigs fed the oxi-dized fat was twice as much as in the pigs fed the freshfat; however, the difference was not statistically signifi-cant due to a great standard deviation of the concentra-tions (P < 0.1, Figure 1).
Expression of PPARa responsive genes in the liverPigs fed the oxidized fat had increased relative mRNAabundance of various PPARa responsive genes such asacyl-CoA oxidase (ACO), liver-type carnitine palmitoyl-transferase 1 (L-CPT1) or novel organic cation transpor-ter 2 (OCTN2) (P < 0.05, Figure 2), indicating anactivation of hepatic PPARa in these animals.
Table 2 Characteristics and performance data of the primers used for qPCR analysis and reference gene-stabilitymeasure M
Gene Forward primer (from 5’ to 3’) Reverse primer (from5’ to 3’)
PCR product size(bp)
NCBI GenBank Slope R2 Efficiency Mvalue
Reference genes
ATP5G1 CAGTCACCTTGAGCCGGGCGATAGCGCCCCGGTGGTTTGC
94 NM_001025218.1 −0.2661 0.9981 1.85 0.036
GPI CACGAGCACCGCTCTGACCTCCACTCCGGACACGCTTGCA
365 NM_214330.1 −0.2557 0.9964 1.80 0.034
RPS9 GTCGCAAGACTTATGTGACCAGCTTAAAGACCTGGGTCTG
327 CAA23101 −0.2705 0.9994 1.86 0.036
b-Actin GACATCCGCAAGCACCTCTAACATCTGCTGGAAGGTGGAC
205 NM_001167795 −0.2637 0.9979 1.84 0.046
SDHA CTACGCCCCCGTCGCAAAGGAGTTTGCCCCCAGGCGGTTG
380 DQ402993 −0.2551 0.9986 1.80 0.041
Target genes
FGF21 CGATACCTCTACACGGATGACGTTGTAGCCATCCTCAAGT
158 NM_001163410.1 −0.2773 0.9968 1.88 -
HMGCS2 ATGGAACGCACGCAGCTCCCTCAGGCCCAACCAGCATGGC
323 NM_214380.1 −0.2528 0.9983 1.79 -
ACO CTCGCAGACCCAGATGAAATTCCAAGCCTCGAAGATGAGT
218 AF185048 −0.2495 0.9986 1.78 -
L-CPT1 GCATTTGTCCCATCTTTCGTGCACTGGTCCTTCTGGGATA
198 AF288789 −0.2654 0.9986 1.84 -
OCTN2 TGACCATATCAGTGGGCTAAGTAGGGAGACAGGATGCT
384 XM_003123912 −0.2517 0.998 1.79 -
Figure 1 Relative mRNA abundance of FGF21 in the liver (A)and protein concentration of FGF21 in the liver (B) and plasma(C) of pigs fed either a fresh fat or an oxidized fat. Barsrepresent mean ± SD (n = 12/group), and are expressed as fold offresh fat group. *Significantly (P < 0.05) and #in tendency (P < 0.1)different from pigs fed the fresh fat. FF, fresh fat group; FGF21,fibroblast growth factor 21; OF, oxidized fat group
Varady et al. Lipids in Health and Disease 2012, 11:34http://www.lipidworld.com/content/11/1/34
Page 3 of 6
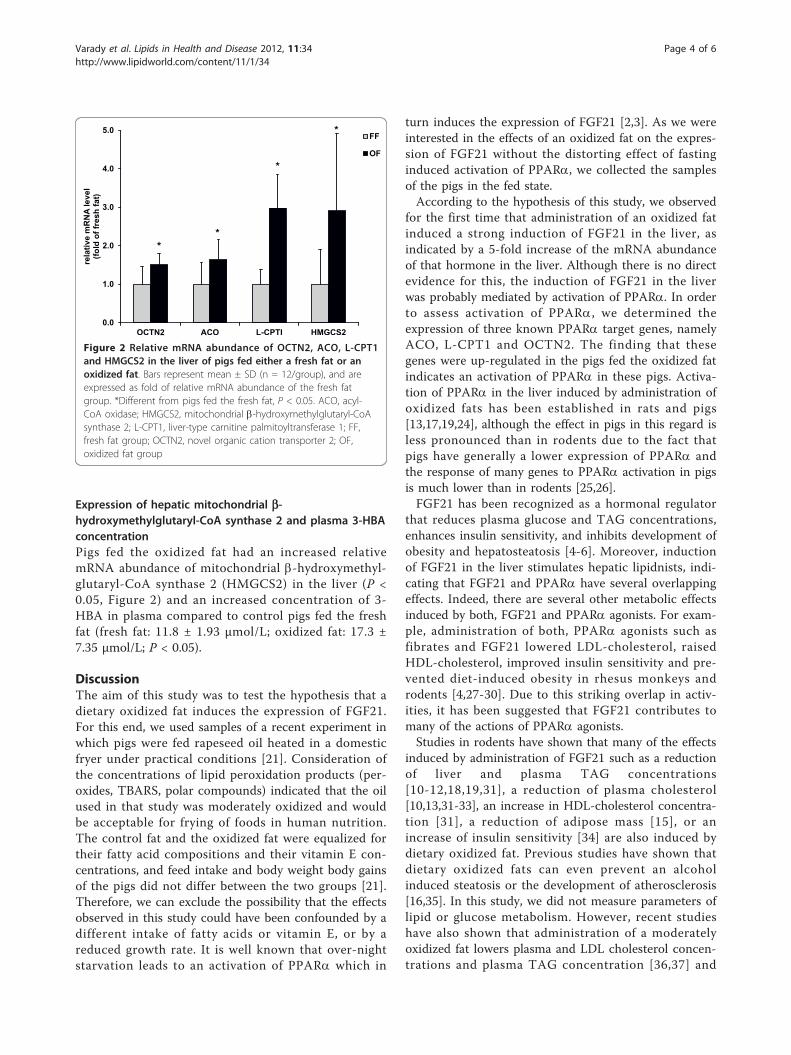
Expression of hepatic mitochondrial b-hydroxymethylglutaryl-CoA synthase 2 and plasma 3-HBAconcentrationPigs fed the oxidized fat had an increased relativemRNA abundance of mitochondrial b-hydroxymethyl-glutaryl-CoA synthase 2 (HMGCS2) in the liver (P <0.05, Figure 2) and an increased concentration of 3-HBA in plasma compared to control pigs fed the freshfat (fresh fat: 11.8 ± 1.93 μmol/L; oxidized fat: 17.3 ±7.35 μmol/L; P < 0.05).
DiscussionThe aim of this study was to test the hypothesis that adietary oxidized fat induces the expression of FGF21.For this end, we used samples of a recent experiment inwhich pigs were fed rapeseed oil heated in a domesticfryer under practical conditions [21]. Consideration ofthe concentrations of lipid peroxidation products (per-oxides, TBARS, polar compounds) indicated that the oilused in that study was moderately oxidized and wouldbe acceptable for frying of foods in human nutrition.The control fat and the oxidized fat were equalized fortheir fatty acid compositions and their vitamin E con-centrations, and feed intake and body weight body gainsof the pigs did not differ between the two groups [21].Therefore, we can exclude the possibility that the effectsobserved in this study could have been confounded by adifferent intake of fatty acids or vitamin E, or by areduced growth rate. It is well known that over-nightstarvation leads to an activation of PPARa which in
turn induces the expression of FGF21 [2,3]. As we wereinterested in the effects of an oxidized fat on the expres-sion of FGF21 without the distorting effect of fastinginduced activation of PPARa, we collected the samplesof the pigs in the fed state.According to the hypothesis of this study, we observed
for the first time that administration of an oxidized fatinduced a strong induction of FGF21 in the liver, asindicated by a 5-fold increase of the mRNA abundanceof that hormone in the liver. Although there is no directevidence for this, the induction of FGF21 in the liverwas probably mediated by activation of PPARa. In orderto assess activation of PPARa, we determined theexpression of three known PPARa target genes, namelyACO, L-CPT1 and OCTN2. The finding that thesegenes were up-regulated in the pigs fed the oxidized fatindicates an activation of PPARa in these pigs. Activa-tion of PPARa in the liver induced by administration ofoxidized fats has been established in rats and pigs[13,17,19,24], although the effect in pigs in this regard isless pronounced than in rodents due to the fact thatpigs have generally a lower expression of PPARa andthe response of many genes to PPARa activation in pigsis much lower than in rodents [25,26].FGF21 has been recognized as a hormonal regulator
that reduces plasma glucose and TAG concentrations,enhances insulin sensitivity, and inhibits development ofobesity and hepatosteatosis [4-6]. Moreover, inductionof FGF21 in the liver stimulates hepatic lipidnists, indi-cating that FGF21 and PPARa have several overlappingeffects. Indeed, there are several other metabolic effectsinduced by both, FGF21 and PPARa agonists. For exam-ple, administration of both, PPARa agonists such asfibrates and FGF21 lowered LDL-cholesterol, raisedHDL-cholesterol, improved insulin sensitivity and pre-vented diet-induced obesity in rhesus monkeys androdents [4,27-30]. Due to this striking overlap in activ-ities, it has been suggested that FGF21 contributes tomany of the actions of PPARa agonists.Studies in rodents have shown that many of the effects
induced by administration of FGF21 such as a reductionof liver and plasma TAG concentrations[10-12,18,19,31], a reduction of plasma cholesterol[10,13,31-33], an increase in HDL-cholesterol concentra-tion [31], a reduction of adipose mass [15], or anincrease of insulin sensitivity [34] are also induced bydietary oxidized fat. Previous studies have shown thatdietary oxidized fats can even prevent an alcoholinduced steatosis or the development of atherosclerosis[16,35]. In this study, we did not measure parameters oflipid or glucose metabolism. However, recent studieshave also shown that administration of a moderatelyoxidized fat lowers plasma and LDL cholesterol concen-trations and plasma TAG concentration [36,37] and
Figure 2 Relative mRNA abundance of OCTN2, ACO, L-CPT1and HMGCS2 in the liver of pigs fed either a fresh fat or anoxidized fat. Bars represent mean ± SD (n = 12/group), and areexpressed as fold of relative mRNA abundance of the fresh fatgroup. *Different from pigs fed the fresh fat, P < 0.05. ACO, acyl-CoA oxidase; HMGCS2, mitochondrial b-hydroxymethylglutaryl-CoAsynthase 2; L-CPT1, liver-type carnitine palmitoyltransferase 1; FF,fresh fat group; OCTN2, novel organic cation transporter 2; OF,oxidized fat group
Varady et al. Lipids in Health and Disease 2012, 11:34http://www.lipidworld.com/content/11/1/34
Page 4 of 6

increases plasma 3-HBA concentration in pigs [24].These studies indicate that administration of an oxidizedfat exerts also effects on lipid metabolism in pigs.Although concentrations of FGF21 in plasma and liverwere less increased than FGF21 mRNA abundance byadministration of the oxidized fat, it is likely that atleast a part of the metabolic effects observed in animalsfed an oxidized fat might be induced by FGF21. Up-reg-ulation of HMGCS2, the key enzyme of ketogenesis, andan increased plasma concentration of 3-HBA observedin the pigs fed the oxidized fat in the present study,might also be - at least in part - induced by FGF21which is regarded as key regulator of ketogenesis [2].Given the similarities between pigs and humans withrespect to expression of PPARa [26] as an importantregulator of FGF21 expression, the possibility exists thata dietary regime rich in fried foods containing oxidizedfats induces hepatic expression of FGF21 in humans.In conclusion, the present study shows for the first
time that the ingestion of a moderately oxidized oilcauses an induction of FGF21 in the liver of pigs, prob-ably induced by activation of PPARa. This findingopens up the possibility that at least some of the effectsobserved in animals administered an oxidized fat areinduced by FGF21.
AbbreviationsACO: Acyl-CoA oxidase; ATP5G1: ATP synthase, H + transporting:mitochondrial Fo complex, subunit C1; 3-HBA: 3-hydroxybutyrate; HMGCS2:Mitochondrial β-hydroxymethylglutaryl-CoA synthase 2; FF: Fresh fat group;FGF21: Fibroblast growth factor 21; GPI: Glucose-6-phosphate isomerase; L-CPT1: Liver isoform of carnitine palmitoyl transferase 1; OCTN2: Novelorganic cation transporter 2; OF: Oxidized fat group; PPARα: Peroxisomeproliferator-activated receptor α; RPS9: Ribosomal protein S9; SDHA:Succinate dehydrogenase complex, subunit A.
AcknowledgementsThis study was supported by a grant from the DeutscheForschungsgemeinschaft (DFG; grant no. RI 1537/1-1).
Authors’ contributionsJV carried out the experiments and participated in the interpretation of thedata and drafted the manuscript. RR participated in the design of the studyand supervised the experiment and analysis. KE conceived of the study andits design, coordinated work, participated in the interpretation of the results,and helped to draft the manuscript. All authors read and approved the finalmanuscript.
Competing interestsThe authors declare that they have no competing interests.
Received: 4 February 2012 Accepted: 6 March 2012Published: 6 March 2012
References1. Badman MK, Pissios P, Kennedy AR, Koukos G, Flier JS, Maratos-Flier E:
Hepatic fibroblast growth factor 21 is regulated by PPARalpha and is akey mediator of hepatic lipid metabolism in ketotic states. Cell Metab2007, 5:426-437.
2. Inagaki T, Dutchak P, Zhao G, Ding X, Gautron L, Parameswara V, Li Y,Goetz R, Mohammadi M, Esser V, Elmquist JK, Gerard RD, Burgess SC,
Hammer RE, Mangelsdorf DJ, Kliewer SA, et al: Endocrine regulation of thefasting response by PPARalpha-mediated induction of fibroblast growthfactor 21. Cell Metab 2007, 5:415-425.
3. Lundåsen T, Hunt MC, Nilsson LM, Sanyal S, Angelin B, Alexson SE,Rudling M, et al: PPARalpha is a key regulator of hepatic FGF21. BiochemBiophys Res Commun 2007, 360:437-440.
4. Kharitonenkov A, Shiyanova TL, Koester A, Ford AM, Micanovic R,Galbreath EJ, Sandusky GE, Hammond LJ, Moyers JS, Owens RA, Gromada J,Brozinick JT, Hawkins ED, Wroblewski VJ, Li DS, Mehrbod F, Jaskunas SR,Shanafelt AB, et al: FGF-21 as a novel metabolic regulator. J Clin Invest2005, 115:1627-1635.
5. Coskun T, Bina HA, Schneider MA, Dunbar JD, Hu CC, Chen Y, Moller DE,Kharitonenkov A, et al: Fibroblast growth factor 21 corrects obesity inmice. Endocrinology 2007, 149:6018-6027.
6. Xu J, Lloyd DJ, Hale C, Stanislaus S, Chen M, Sivits G, Vonderfecht S,Hecht R, Li YS, Lindberg RA, Chen JL, Zhang Z, Ko HJ, Kim JK, Véniant MM,et al: Fibroblast growth factor 21 reverses hepatic steatosis, increasesenergy expenditure, and improves insulin sensitivity in diet-inducedobese mice. Diabetes 2009, 58:250-259.
7. Oishi K, Uchida D, Ishida N, et al: Circadian expression of FGF21 isinduced by PPARalpha activation in the mouse liver. FEBS Lett 2008,582:3639-3642.
8. Yu J, Yu B, Jiang H, Chen D, et al: Conjugated linoleic acid induceshepatic expression of fibroblast growth factor 21 through PPAR-α. Br JNutr 2011, 18:1-5.
9. Guthrie JF, Lin BH, Frazao E, et al: Role of food prepared away from homein the American diet, 1977-78 versus 1994-96: changes andconsequences. J Nutr Educ Behav 2002, 34:140-150.
10. Huang CJ, Cheung NS, Lu VR, et al: Effects of deteriorated frying oil anddietary protein levels on liver microsomal enzymes in rats. J Am OilChem Soc 1988, 65:1796-1803.
11. Eder K, Kirchgessner M, et al: The effect of dietary vitamin E supply and amoderately oxidized oil on activities of hepatic lipogenic enzymes inrats. Lipids 1998, 33:277-283.
12. Eder K, et al: The effects of a dietary oxidized oil on lipid metabolism inrats. Lipids 1999, 34:717-725.
13. Chao PM, Chao CY, Lin FJ, Huang C, et al: Oxidized frying oil up-regulateshepatic acyl-CoA oxidase cytochrome P450 4 A1 genes in rats activatesPPARalpha. J Nutr 2001, 131:3166-3174.
14. Eder K, Suelzle A, Skufca P, Brandsch C, Hirche F, et al: Effects of dietarythermoxidized fats on expression and activities of hepatic lipogenicenzymes in rats. Lipids 2003, 38:31-38.
15. Chao PM, Huang HL, Liao CH, Huang ST, Huang CJ, et al: A high oxidisedfrying oil content diet is less adipogenic, but induces glucoseintolerance in rodents. Br J Nutr 2007, 98:63-71.
16. Kämmerer I, Ringseis R, Eder K, et al: Feeding a thermally oxidised fatinhibits atherosclerotic plaque formation in the aortic root of LDLreceptor-deficient mice. Br J Nutr 2011, 105:190-199.
17. Chao PM, Hsu SC, Lin FJ, Li YJ, Huang CJ, et al: The up-regulation ofhepatic acyl-CoA oxidase and cytochrome P450 4A1 mRNA expressionby dietary oxidized frying oil is comparable between male and femalerats. Lipids 2004, 39:233-238.
18. Chao PM, Yang MF, Tseng YN, Chang KM, Lu KS, Huang CJ, et al:Peroxisome proliferation in liver of rats fed oxidized frying oil. J Nutr SciVitaminol 2005, 51:361-368.
19. Sülzle A, Hirche F, Eder K, et al: Thermally oxidized dietary fat upregulatesthe expression of target genes of PPARα in rat liver. J Nutr 2004,134:1375-1383.
20. Ringseis R, Eder K: Regulation of genes involved in lipid metabolism bydietary oxidized fat. Mol Nutr Food Res 2011, 55:109-121.
21. Varady J, Gessner DK, Most E, Eder K, Ringseis R: Dietary moderatelyoxidized oil activates the Nrf2 signaling pathway in the liver of pigs.Lipids Health Dis (in press) .
22. Gesellschaft für Ernährungsphysiologie: Empfehlungen zur Energie undNährstoffversorgung bei Schweinen Frankfurt, Germany DLG-Verlag; 2006.
23. Keller J, Ringseis R, Koc A, Lukas I, Kluge H, Eder K: Supplementation withL-carnitine downregulates genes of the ubiquitin proteasome system inthe skeletal muscle and liver of piglets. Animal 2011, 6:70-78.
24. Luci S, König B, Giemsa B, Huber S, Hause GI, Kluge H, Stangl , Eder K:Feeding of a deep-fried fat causes PPARalpha activation in the liver ofpigs as a non-proliferating species. Br J Nutr 2007, 97:872-882.
Varady et al. Lipids in Health and Disease 2012, 11:34http://www.lipidworld.com/content/11/1/34
Page 5 of 6

25. Cheon Y, Nara TY, Band MR, Beever JE, Wallig MA, Nakamura MT: Inductionof overlapping genes by fasting and a peroxisome proliferator in pigs:evidence of functional PPARalpha in nonproliferating species. Am JPhysiol Regul Integr Comp Physiol 2005, 288:1525-1535.
26. Luci S, Giemsa B, Kluge H, Eder K: Clofibrate causes an upregulation ofPPARα target genes but does not alter expression of SREBP targetgenes in liver and adipose tissue of pigs. Am J Physiol Regul Integr CompPhysiol 2007, 293:70-77.
27. Kharitonenkov A, Wroblewski VJ, Koester A, Chen YF, Clutinger CK, Tigno XT,Hansen BC, Shanafelt AB, Etgen GJ: The metabolic state of diabeticmonkeys is regulated by fibroblast growth factor-21. Endocrinology 2007,148:774-781.
28. Winegar DA, Brown PJ, Wilkison WO, Lewis MC, Ott RJ, Tong WQ,Brown HR, Lehmann JM, Kliewer SA, Plunket KD, Way JM, Bodkin NL,Hansen BC: Effects of fenofibrate on lipid parameters in obese rhesusmonkeys. J Lipid Res 2001, 42:1543-1551.
29. Chou CJ, Haluzik M, Gregory C, Dietz KR, Vinson C, Gavrilova O, Reitman ML:WY14,643, a peroxisome proliferator-activated receptor alpha(PPARalpha ) agonist, improves hepatic and muscle steatosis andreverses insulin resistance in lipoatrophic A-ZIP/F-1 mice. J Biol Chem2002, 277:24484-24489.
30. Tsuchida A, Yamauchi T, Takekawa S, Hada Y, Ito Y, Maki T, Kadowaki T:Peroxisome proliferator-activated receptor (PPAR)alpha activationincreases adiponectin receptors and reduces obesity-relatedinflammation in adipose tissue: comparison of activation of PPARα,PPARγ, and their combination. Diabetes 2005, 54:3358-3370.
31. Eder K, Keller U, Hirche F, Brandsch C: Thermally oxidized dietary fatsincrease the susceptibility of rat LDL to lipid peroxidation but not theiruptake by macrophages. J Nutr 2003, 133:2830-2837.
32. Ammouche A, Rouaki F, Bitam A, Bellal MM: Effect of ingestion ofthermally oxidized sunflower oil on the fatty acid composition andantioxidant enzymes of rat liver and brain in development. Ann NutrMetab 2002, 46:268-275.
33. Koch A, König B, Spielmann J, Leitner A, Stangl GI, Eder K: Thermallyoxidized oil increases the expression of insulin-induced genes andinhibits activation of sterol regulatory element-binding protein-2 in ratliver. J Nutr 2007, 137:2018-2023.
34. Liao CH, Shaw HM, Chao PM: Impairment of glucose metabolism in miceinduced by dietary oxidized frying oil is different from that induced byconjugated linoleic acid. Nutrition 2008, 24:744-752.
35. Ringseis R, Muschick A, Eder K: Dietary oxidized fat prevents ethanol-induced triacylglycerol accumulation and increases expression of PPARαtarget genes in rat liver. J Nutr 2007, 137:77-83.
36. Eder K: The effect of an oxidized dietary oil on plasma cholesterol andthyroid hormone concentrations in miniature pigs fed a hyperlipidemicdiet. J Anim Physiol Anim Nutr 1999, 82:271-281.
37. Eder K, Stangl GI: Plasma thyroxine and cholesterol concentrations ofminiature pigs are influenced by thermally oxidized dietary lipids. J Nutr2000, 130:116-121.
doi:10.1186/1476-511X-11-34Cite this article as: Varady et al.: Dietary moderately oxidized oil inducesexpression of fibroblast growth factor 21 in the liver of pigs. Lipids inHealth and Disease 2012 11:34.
Submit your next manuscript to BioMed Centraland take full advantage of:
• Convenient online submission
• Thorough peer review
• No space constraints or color figure charges
• Immediate publication on acceptance
• Inclusion in PubMed, CAS, Scopus and Google Scholar
• Research which is freely available for redistribution
Submit your manuscript at www.biomedcentral.com/submit
Varady et al. Lipids in Health and Disease 2012, 11:34http://www.lipidworld.com/content/11/1/34
Page 6 of 6

Diskussion
38
4. Diskussion
4.1 Einfluss oxidierter Fette auf den Nrf2 In den eigenen Untersuchungen (Studie 1 und 2) konnte erstmals festgestellt werden, dass die
Verabreichung oxidierter Fette eine starke Aktivierung des redox-sensitiven Transkriptionsfaktors
Nrf2, welche über eine Erhöhung der nukleären Proteinkonzentrationen ermittelt wurde, sowohl in
den Darmepithelzellen der Maus als auch in der Leber vom Schwein verursacht. Als Folge der Nrf2-
Aktivierung konnte ein Anstieg der hepatischen und intestinalen Expression Nrf2-abhängiger
Zielgene, welche in die antioxidative Abwehr (wie SOD1, TXNR1) und/oder in die Phase II des
Fremdstoffmetabolismus (wie NAD(P)H-Quinon-Oxidoreduktase-1 (NQO1), UGT1A1) involviert
sind, beobachtet werden.
Sowohl der Darm als auch die Leber sind durch ihre exponierte Lage permanent Fremdstoffen
ausgesetzt. Um daraus resultierende Gewebeschäden und die Verteilung toxischer Substanzen im
Organismus zu verhindern, sind in beiden Organen effektive Mechanismen entwickelt, um auf
zellulären Stress zu reagieren (Rushmore und Kong, 2002).
Der Darm ist als mechanische Barriere zwischen dem Körperinneren und der Außenwelt
kontinuierlich gegenüber verschiedensten Mikroorganismen, Fremd- und Giftstoffen exponiert. Mit
der Nahrung aufgenommene Lipidperoxidationsprodukte aus oxidierten Fetten können von
Darmepithelzellen aufgenommen und über Chylomikronen und VLDL im Körper verteilt werden
(Penumetcha et al., 2000; Staprans et al., 2003; Staprans et al., 1994; Staprans et al., 1993; Suomela et
al., 2005; Suomela et al., 2004). Der Nrf2 übernimmt im Darm eine maßgebliche Rolle zum Erhalt und
Schutz der intestinalen Funktion und Integrität durch die Regulation von antioxidativen und
detoxifizierenden Enzymen und der Bildung proinflammatorischer Zytokine (McMahon et al., 2001).
In verschiedenen Mausmodellen konnten die günstigen und substantiellen Auswirkungen einer Nrf2-
Aktivierung im Darm und der Leber herausgestellt werden (Osburn et al., 2007; Khor et al. 2008;
Okawa et al., 2006). So waren Nrf2-knock out Mäuse aufgrund einer verminderten Expression Nrf2-
regulierter Gene, wie UGT1A1 und NQO1, und einer gleichzeitig erhöhten Expression
inflammatorischer Zytokine im Vergleich zu Wildtyp-Mäusen anfälliger gegenüber experimentell
induzierter Kolitis (Khor et al., 2006).
Die Leber ist das bedeutendste Organ zur Entgiftung und Ausscheidung toxischer Substanzen im
Rahmen des Fremdstoffmetabolismus. Während der drei Phasen werden lipophile Fremdstoffe
endogenen und exogenen Ursprungs in hydrophilere Metaboliten transformiert, um über die Galle
oder den Urin aus dem Organismus eliminiert zu werden. Der Nrf2 ist ein sehr bedeutender Regulator
der Phase II-Enzyme (Itoh et al., 1997). Über die Induktion dieser detoxifizierenden Enzyme, übt er
eine protektive Schutzfunktion aus. Deutlich wird dies in der Nrf2-null Maus, welche aufgrund einer

Diskussion
39
erhöhten Anfälligkeit gegenüber chemisch-induzierten oxidativen und elektrophilen Stress
hepatische Schäden und Verletzungen aufweist (Lu et al., 2008; Xu et al., 2008). Auch die Anfälligkeit
der Leber gegenüber der Entstehung von Krebs wird durch das Fehlen einer Nrf2-vermittelten
Genexpression in Nrf2-defizienten Mäusen verstärkt (Ramos-Gomez et al., 2001).
Wie aus verschiedenen Untersuchungen bekannt ist, führt die Aufnahme oxidierter Fette aufgrund
einer vermehrten exogenen Aufnahme (Lipidperoxidationsprodukte) und endogenen Bildung von
ROS zur Induktion von oxidativem Stress. ROS sind in der Lage den Nrf2 zu aktivieren, da sie mit
spezifischen Cysteinresten am Keap1 interagieren, woraufhin es zur Konformationsänderung am
Inhibitorprotein Keap1 und zur Freisetzung von Nrf2 kommt (Fourquet et al., 2010; Wakabayashi et
al., 2004). Es ist daher sehr wahrscheinlich, dass auch in den eigenen Untersuchungen (Studie 1
und 2) eine Aktivierung des Nrf2-Signalweges durch ROS verursacht wurde. Auch
4-Hydroxynonenal (4-HNE) muss bei einer Aktivierung des Nrf2 durch oxidierte Fette in Betracht
gezogen werden. 4-HNE ist eines der Hauptendprodukte der Lipidperoxidation und in die
stressvermittelte Signaltransduktion involviert (Camandola et al., 2000; Song et al., 2001). In geringen
Konzentrationen kann es jedoch auch zytoprotektiv wirken, in dem es die Aktivierung des Nrf2 und
der Nrf2-vermittelten Genexpression, wie der Thioredoxinreduktase 1, bewirkt (Chen et al., 2005;
Ishii et al., 2004). Möglicherweise wurde der Nrf2 auch direkt durch spezifische
Lipidperoxidationsprodukte, welche im oxidierten Behandlungsfett enthalten waren, aktiviert.
Hinweise dafür gibt es aus einer Untersuchung, in welcher Oxidationsprodukte der ω-3-Fettsäuren
Eicosapentaensäure und Docosahexaensäure mit Thiolgruppen am Keap1 reagierten und so die
Abdissoziierung des Nrf2 von Keap1 veranlassten (Gao et al., 2007).
Innerhalb der Wirbeltiere ist die Familie der CNC-bZip-Proteine, zu denen auch der Nrf2 zählt,
evolutionär hoch konserviert. So ist die DNA- und Protein-Homologie des murinen Nrf2s im
Vergleich zum humanen Nrf2 mit 83,4 % und 82,5 % relativ stark ausgeprägt. Für das Schwein gibt es
diesbezüglich noch keine Daten, jedoch ist die Struktur- und Sequenzhomologie zwischen Mensch
und Schwein generell hoch (Lunney, 2007). Da eine Aktivierung des Nrf2-Signalweges durch
oxidierte Fette sowohl in der Maus und als auch im Schwein nachgewiesen werden konnte, kann von
einer speziesübergreifenden Wirkung der oxidierten Fette gesprochen werden. Insbesondere in der
Leber ist eine Aktivierung der Phase II des Fremdstoffmetabolismus durch oxidierte Fette als kritisch
zu betrachten, da viele Arzneimittel über dieselben Mechanismen metabolisiert werden. So könnte es
zu Veränderungen in der Pharmakokinetik von Arzneimitteln mit unerwünschten Neben- und
Wechselwirkungen kommen. Beispielsweise wäre denkbar, dass aufgrund einer gesteigerten
Metabolisierung von Arzneimitteln durch Phase II-Enzyme die normalen Standarddosen nicht richtig
wirken bzw. ausreichen könnten, um die erwünschte Wirkung zu erzielen.
Eine weitere Beobachtung der eigenen Untersuchung (Studie 2), welche als kritisch zu bewerten gilt,
ist die Beeinflussung des Schilddrüsenhormonstatus. So konnte gezeigt werden, dass die

Diskussion
40
Konzentration des Schilddrüsenhormons Gesamt-T4 im Plasma durch die Aufnahme des oxidierten
Fettes sank. Die Steigerung der UGT1A1-Genexpression und der T4-UGT-Aktivität sind sehr
wahrscheinlich ursächlich für die erniedrigten Konzentrationen an Gesamt-T4 im Plasma.
In der Maus führte die Verabreichung spezifischer Nrf2-Aktivatoren zu einer Erhöhung der
hepatischen UGT1A1-mRNA-Expression, was auf eine Regulation von UGT1A1 durch Nrf2 hinweist
(Buckley et al., 2009). Eine funktionelle ARE-Sequenz konnte im UGT1A1-Promotorbereich von
HepG2-Zellen nachgewiesen werden (Yueh und Tukey, 2007). Im Menschen sind Mitglieder der
UGT1A-Familie, insbesondere UGT1A1 verantwortlich für die Glukuronidierung des
Schilddrüsenhormons T4 in der Leber (Kato et al., 2008; Yamanaka et al., 2007). Beim Menschen ist
ein Polymorphismus im Promotorbereich des UGT1A1-Gens (UGT1A1*28) bekannt, der zu einer
verminderten hepatischen Enzymaktivität und zu erniedrigten Konzentrationen an T4-Glucuroniden
führt (Yoder Graber et al., 2007). Dass UGT1A1 eine T4-UGT-Aktivität besitzt, konnte bisher im
Menschen und der Ratte nachgewiesen werden (Vansell et al., 2002; Yamanaka et al., 2007).
Die Ergebnisse der eigenen Untersuchung (Studie 2) stehen somit aber auch im Widerspruch zu
vorangegangenen Untersuchungen, in welchen die Aufnahme oxidierter Fette zu einer Erhöhung der
Plasmakonzentrationen von freiem T4 und Gesamt-T4 in der Ratte und im Schwein führte (Eder und
Stangl, 2000; Eder et al., 2002; Skufca et al., 2003). Jedoch stehen sie im Einklang mit den Ergebnissen
einer Untersuchung am Schwein, in welcher die Verabreichung von Clofibrat als synthetischen
PPARα-Agonisten zu einem erhöhten Abbau der Schilddrüsenhormone T3 und T4 durch eine
Steigerung der UGT-Aktivität in der Leber führte. Es wäre daher denkbar, dass auch in der eigenen
Untersuchung nicht die Aktivierung des Nrf2 sondern die des PPARα durch oxidierte Fette
ursächlich für den verstärkten Abbau des Hormons T4 ist. Dafür spricht auch, dass im UGT1A1-
Promotor Bindungsstellen für mehrere Transkriptionsfaktoren, wie dem PPARα, gefunden wurden
(Buckley und Klaassen, 2009).
Möglicherweise sind aber auch methodische Unterschiede zwischen den verschiedenen
Untersuchungen ursächlich für die gegensätzlichen Ergebnisse. In den vorangegangen Studien,
welche sich mit dem Einfluss oxidierter Fette auf Schilddrüsenparameter beschäftigten, wurden als
Versuchsfette in aller Regel Sonnenblumenöl und Schweinefett/-schmalz verwendet, welche
wesentlich stärker oxidiert waren, wie an den gemessenen Lipidperoxidationsprodukten zu erkennen
ist. Im Gegensatz dazu wurde in der eigenen Untersuchung (Studie 2) erstmals Rapsöl verwendet,
welches nur sehr moderat erhitzt wurde. Es ist daher denkbar, dass verschiedenartige Fette mit
verschiedenen Oxidationsgraden auch unterschiedliche Wirkungen im Organismus vermitteln. Zur
genauen Beurteilung dieser Effekte und aufgrund der inkonsistenten Datenlage wären weitere
Untersuchungen zur Klärung des Einflusses oxidierter Fette auf den Schilddrüsenhormonstatus
notwendig.

Diskussion
41
Aus der Literatur gibt es Hinweise, dass der Nrf2 mit verschiedenen anderen Signalwegen interagiert,
insbesondere mit dem des NF-κB (Kratschmar et al., 2012; Levy und Forman, 2010; Venugopal und
Jaiswal, 1996). Für diese beiden Transkriptionsfaktoren sind größtenteils antagonisierende Effekte
bekannt (Li et al., 2008; Yang et al., 2012). Die Regulation der redox-sensitiven
Transkriptionsfaktoren ist sehr wahrscheinlich abhängig von der Höhe der ROS-Exposition, den
intrazellulären antioxidativen Kapazitäten und den Veränderungen im zellulären Redoxpotential. So
scheint in der frühen Phase des oxidativen Stresses bzw. bei mildem oxidativen Stress, welcher nur
mit einer geringen Veränderung des zellulären Redoxpotentials einhergeht, die Aktivierung des Nrf2-
Signalwegs zur Wiederherstellung der zellulären Homöostase die vorherrschende Rolle zu spielen.
Zahlreiche potente Aktivatoren des Nrf2, sogenannte chemopräventive Substanzen, wie das
Isothiocyanat Sulforaphan, welche geringe Veränderungen im Redoxstatus der Zelle bewirken, üben
eine gleichzeitige Hemmung auf den NF-κB und die Expression seiner Zielgene aus (Heiss et al., 2001;
Karuri et al., 2006). Wahrscheinlich geschieht dies über eine Hemmung der Phosphorylierung des
NF-κB-Inhibitors IκB (Xu et al., 2005). Zahlreiche Studien weisen auch auf eine zentrale Rolle des
Nrf2 zum Schutz vor inflammatorischen Prozessen und Schädigungen hin. Eine Aktivierung des Nrf2
ist essentiell, um eine koordinierte und kontrollierte Aktivierung der NF-κB-vermittelten Induktion
proinflammatorischer Zytokine zu gewährleisten. So kann in Nrf2-knock out und Nrf2-defizienten
Mäusen im Vergleich zu Wildtyp-Mäusen eine überschießende Aktivierung des NF-κB mit erhöhten
Konzentrationen an proinflammatorischen Markern, wie Interleukin (IL)-6, Tumor necrosis factor
(TNF)-α, inducible nitric oxide synthase (iNOS) und Cytochrome c oxidase (COX-2) beobachtet werden
(Khor et al., 2006; Osburn et al., 2007).
Starker oxidativer Stress führt hingegen nicht zur Aktivierung des Nrf2-Signalweges (Garbin et al.,
2009). Vielmehr steht hier die Aktivierung des NF-κB-Signalweges im Vordergrund. In diesem
Zusammenhang konnte nachgewiesen werden, dass der NF-κB/p65 ist in der Lage ist, den Nrf2/ARE-
Signalweg zu supprimieren, indem er die Bindung von Nrf2 an seinen Coaktivator CRB verhindert
und weiterhin die Rekrutierung des Corepressors Histondeacetylase 3 an ARE veranlasst (Liu et al.,
2008).
Es gibt in der Literatur jedoch auch Hinweise auf eine simultane Aktivierung beider
Transkriptionsfaktoren. Viele pathologische Stimuli, wie z.B. ROS, sind potente Aktivatoren sowohl
von Nrf2 als auch von NF-κB (Gloire et al., 2006; Itoh et al., 1997). Auch einige Enzyme, wie
Hämoxygenase (HO)-1, Glutathionperoxidase (GPX)1 und SOD1 enthalten sowohl ARE- und als auch
κB-Sequenzen in ihrer Promotorregion, so dass eine Aktivierung dieser Gene durch beide
Transkriptionsfaktoren möglich ist (Banning et al., 2005; Dreger et al., 2009; Inamdar et al., 1996;
Lavrovsky et al., 1994; Rojo et al., 2004).
In moderaten und starken Rauchern konnte eine negative Korrelation für die Proteinexpressionen von
Nrf2 und NF-κB festgestellt werden (Garbin et al., 2009). Im Vergleich zu Nichtrauchern führte in
jungen Rauchern der Konsum von 5-10 Zigaretten täglich zu einer simultanen Aktivierung des Nrf2-

Diskussion
42
und des NF-κB-Signalweges. Bei starkem Zigarettenkonsum mit 25-40 Zigaretten täglich konnte
hingegen nur noch eine Aktivierung des NF-κB festgestellt werden. Die Nrf2-Proteinkonzentrationen
pegelten sich in Höhe der Nichtraucher ein. Der erhöhte oxidative Stress ausgelöst durch starkes
Rauchen verursachte wahrscheinlich die bekannte negative Regulierung des Nrf2 durch den NF-κB.
Diese Beobachtungen an moderaten Rauchern sind in guter Übereinstimmung mit den eigenen
Ergebnissen (Studie 1 und 2), welche ebenfalls eine simultane Aktivierung beider
Transkriptionsfaktoren aufzeigen. Möglicherweise verursachte die Aufnahme der moderat oxidierten
Fette nur milde bis mäßige Veränderungen im zellulären Redoxstatus in der Leber und den
Darmepithelzellen. In Analogie zu den Starkrauchern ist daher denkbar, dass bei Aufnahme eines
stärker oxidierten Fettes eine noch stärkere NF-κB-Aktivierung und eine Hemmung des Nrf2 hätten
nachgewiesen werden können.
Zusammenfassend lässt sich feststellen, dass in den eigenen Untersuchungen (Studie 1 und 2)
erstmals eine Aktivierung des Nrf2-Signalweges durch oxidierte Fette, welche unter
Frittierbedingungen hergestellt wurden, ermittelt werden konnte. Somit scheint der Nrf2 das
mechanistische Bindeglied zwischen dem durch die Aufnahme oxidierter Fette verursachten
oxidativen Stress und der Aktivierung zellulärer Adaptationsreaktionen durch die Induktion
antioxidativer und detoxifizierender Enzyme zu sein.
Eine Aktivierung des Nrf2-Signalweges wird in der Literatur überwiegend als günstig für den
Organismus gewertet, da durch eine Aktivierung einer Vielzahl von zytoprotektiven Enzymen
Schäden durch ROS oder anderen Toxinen an Biomolekülen verhindert bzw. vermindert werden
können (Baird et al., 2011; Kim et al., 2010; Kwak und Kensler, 2010). Im Hinblick auf eine verstärkte
Phase II-Metabolisierung und eine Beeinflussung des Schilddrüsenhormonstatus ist die Aktivierung
des Nrf2 durch oxidiertes Fett in den eigenen Untersuchungen eher als ungünstig für den Organismus
zu bewerten. Die koordinierte simultane Aktivierung des NF-κB- und Nrf2-Signalweges in der Leber
und den Darmepithelzellen stellt vielmehr einen adaptiven und protektiven Mechanismus dar, um
den negativen Wirkungen der oxidierten Fette entgegenzuwirken.
4.2 Einfluss oxidierter Fette auf den NF-κB
In den eigenen Untersuchungen (Studie 1 und 2) konnte erstmals festgestellt werden, dass die
Verabreichung oxidierter Fette eine starke Aktivierung des redox-sensitiven Transkriptionsfaktors
NF-κB, welche über eine Erhöhung der nukleären Proteinkonzentrationen ermittelt wurde, sowohl in
den Darmepithelzellen der Maus als auch in der Leber vom Schwein verursacht. In diesem
Zusammenhang konnte ein Anstieg der Expression antioxidativer und oxidativer Stress-responsiver
Gene, welche in Teilen auch durch den NF-κB reguliert werden, beobachtet werden.

Diskussion
43
Ursächlich für die Aktivierung des redox-sensitiven NF-κB in der Leber und den Darmepithelzellen
ist mit hoher Wahrscheinlichkeit die Aufnahme von Lipidperoxidationsprodukten aus dem
oxidierten Fett und der daraus resultierende oxidative Stress, welcher mit einer verstärkten Bildung
von ROS einhergeht (Gloire et al., 2006). Außerdem ist eine direkte Aktivierung des NF-κB durch
Bestandteile des oxidierten Fettes, wie 13-HPODE, denkbar (Natarajan et al., 2001).
Der NF-κB übernimmt im Organismus wichtige Funktionen in physiologischen und
pathophysiologischen Prozessen. Er ist einer der zentralen Regulatoren der proinflammatorischen
Genexpression. Eine erhöhte nukleäre Lokalisation und verstärkte Aktivität des NF-κB zeigt sich bei
einer Reihe von Krankheiten, wie chronisch-entzündlichen Darmerkrankungen (CED), rheumatoider
Arthritis oder Psoriasis (Ardite et al., 1998; Johansen et al., 2005; Makarov, 2001). Im Rahmen
inflammatorischer Prozesse im Darm kommt dem NF-κB durch die Induktion der Expression
proinflammatorischer Gene, wie Zytokine, Chemokine und Adhäsionsmolekülen eine pathogene
Rolle zu. So kann in Patienten mit CED die Aktivierung des NF-κB und Sekretion
proinflammatorischer Zytokine im Darm nachgewiesen werden (Schreiber et al., 1998). Eine
Inhibierung der NF-κB-Aktivität kann zur Verbesserung von intestinalen Entzündungszuständen,
wie der Colitis, führen (Herfarth et al., 2000).
Gleichzeitig kommen dem NF-κB aber auch protektive Funktionen im Darm zu, indem er als
wichtiger Regulator der intestinalen Immunhomöostase und Integrität der Darmepithelzellen
fungiert (Pasparakis, 2008; Spehlmann und Eckmann, 2009). Eine Inhibierung der NF-κB-Aktivität
speziell in den Darmepithelzellen kann zu Störungen in der Homöostase und zum Auslösen
inflammatorischer Prozesse führen (Nenci et al., 2007). In Mäusen mit einer spezifischen Inhibierung
des NF-κB in Darmepithelzellen durch Ablation von NEMO kommt es aufgrund einer gesteigerten
Apoptoserate von Kolonepithelzellen und einer beeinträchtigten Bildung antimikrobieller Peptide zur
Translokation von Bakterien in die Mukosa und somit zur Entwicklung starker CED (Nenci et al.,
2007). Auch in der Leber scheint die NEMO-abhängige Aktivierung des NF-κBs wichtige
physiologische Funktionen zu vermitteln. Die Ablation von NEMO fördert die Entwicklung
chronischer Hepatitis, welche sich durch eine gesteigerte Apoptoserate von Hepatozyten,
Entzündungen und Steatose äußert und schließlich in die Entstehung von Leberkrebs mündet
(Luedde et al., 2007).
Die eigenen Ergebnisse (Studie 1 und 2) stehen im Widerspruch zu einer vorangegangenen
Untersuchung, welche sich mit dem Einfluss eines oxidierten Fettes auf inflammatorische Prozesse
im Darm vom Schwein beschäftigte (Ringseis et al., 2007b). Obwohl es in dieser Untersuchung
Anzeichen für oxidativen Stress, wie erhöhte Lipidperoxidationsprodukte und erniedrigte
Konzentrationen an Antioxidantien gab, konnte keine Aktivierung des NF-κB in den
Darmepithelzellen ermittelt werden. Möglicherweise war der durch die Aufnahme des oxidierten

Diskussion
44
Fettes ausgelöste oxidative Stress zu schwach bzw. reichten die Kapazitäten des antioxidativen
Abwehrsystems aus, um die NF-κB-vermittelte Signaltransduktion nicht einzuleiten.
Für die unterschiedlichen Ergebnisse bezüglich einer NF-κB-Aktivierung in den Darmepithelzellen
könnte die Art der Verabreichung des Versuchsfettes ursächlich sein. Während die Schweine das
oxidierte Fett im Futter gemischt aufnahmen, wurde den Mäusen der eigenen Untersuchung
(Studie 1) das Versuchsfett per Schlundsonde verabreicht, was zu einer direkten Exposition des
Darms gegenüber dem oxidierten Fett ohne störende Futterkomponenten führte. Bezogen auf das
Körpergewicht hatte die Maus im Vergleich zum Schwein auch eine wesentlich höhere Menge der
Versuchsfette aufgenommen.
PPARα und PPARγ sind in die Kontrolle und Regulation inflammatorischer Prozesse involviert. Ihre
anti-inflammatorischen Wirkungen können sie u. a. über die Transrepression des NF-κB-Signalweges
vermitteln. Der PPARγ vermittelt seine antagonisierenden Eigenschaften wahrscheinlich über die
Bildung eines Repressorkomplexes in der Promotorregion der NF-κB-Zielgene oder durch direkte
Bindung an die p50- und p65-Untereinheiten des NF-κB, wodurch dessen transkriptionelle Aktivität
verloren geht (Chung et al., 2000; Li et al., 2000). Die Verabreichung synthetischer PPARγ-Liganden,
wie Rosiglitazon, führt in Tiermodellen und Patienten mit chronisch-inflammtorischen Krankheiten,
wie ulcerativer Colitis und rheumatoider Arthritis, zur Reduktion inflammatorischer Prozesse und
Verminderung inflammatorischer Marker, wie Zytokinen und Adhäsionsmolekülen (Kawahito et al.,
2000; Lewis et al., 2001). Auch für den PPARα sind in der Literatur verschiedene molekulare
Mechanismen beschrieben, über welche er den NF-κB-Signalweg antagonisieren kann. So ist PPARα
in der Lage, inaktive Komplexe mit der NF-κB-Untereinheit p65 zu bilden und die mRNA- und
Protein-Expression des Inhibitorproteins IκBα zu induzieren, wodurch die Bindung des NF-κB an die
DNA verringert wird (Delerive et al, 2000; Delerive et al., 1999).
Um zu überprüfen, ob auch in den eigenen Untersuchungen (Studie 1 und 2) eine Aktivierung der
PPARs durch oxidiertes Fett eine Transrepressionsaktivität auf den NF-κB ausübt, wurde zusätzlich
die hepatische und intestinale mRNA-Expression PPAR-responsiver Gene bestimmt. Die
vorherrschende PPAR-Isoform in der Leber ist der PPARα, wohingegen in den Darmepithelzellen
hohe Expressionsraten an PPARγ zu finden sind. Dass oxidierte Fette in der Lage sind, den PPARα
und PPARγ in diesen Geweben zu aktivieren, konnte bereits in verschiedenen Untersuchungen an
Nagern und im Schwein nachgewiesen werden (Ringseis et al., 2007b; Luci et al., 2007; Sülzle et al.,
2004).
Das Ausmaß der ermittelten PPAR-Aktivierung (Studie 1 und 2) war in den Darmepithelzellen der
Maus stärker als in der Leber vom Schwein. Diese Beobachtung kann verschiedene Gründe zur
Ursache haben. Einerseits zählt die Maus im Gegensatz zum Schwein zur proliferierenden Spezies.
Somit sind auch die Effekte einer PPARα-abhängigen Aktivierung generell stärker. Andererseits war
das Versuchsfett der Maus stärker oxidiert als das Versuchsfett vom Schwein. Möglicherweise waren

Diskussion
45
deshalb in dem Fett mehr Lipidperoxidationsprodukte enthalten, welche als Liganden der PPARs
fungierten.
Die Aktivierung des PPAR-Signalweges konnte jedoch weder in der Leber noch in den
Darmepithelzellen die Aktivierung NF-κB-Signalweges hemmen, was die Stärke des durch oxidierte
Fette verursachten oxidativen Stresses in den betreffenden Geweben deutlich macht.
Zusammenfassend kann daher festgestellt werden, dass die Aufnahme unter Frittierbedingungen
hergestellter oxidierter Fette zur Aktivierung des NF-κB-Signalweges im Darm und der Leber führt.
4.3 Einfluss eines oxidierten Fettes auf den FGF21
Die Bedeutung des FGF21 als metabolischer Regulator konnte erstmals in vitro in murinen 3T3-L1- und
primären humanen Adipozyten beschrieben werden, wo die Behandlung mit rekombinantem FGF21
die insulinunabhängige Glukoseaufnahme durch die Erhöhung der Transkript- und Proteinmenge des
Glukosetransporters (GLUT)1 stimulierte (Kharitonenkov et al., 2005). In zahlreichen
Untersuchungen an genetisch modifizierten (ob/ob, db/db) und diätinduzierten Mausmodellen für
Diabetes mellitus Typ II und Fettleibigkeit ließ sich ein positiver Einfluss einer systemischen
Verabreichung von rekombinantem FGF21 auf metabolische Parameter und Zustände, wie die
Glukosetoleranz, die Insulinsensitivität, das Lipidprofil in Plasma und Leber, die hepatische Steatose
sowie auf Körpermasse und -fettanteil, feststellen (Coskun et al., 2008; Xu et al., 2009a; Xu et al.,
2009b; Berglund et al., 2009). Ähnliche Ergebnisse zeigten sich in einer weiterführenden
Untersuchung an diabetischen Rhesusaffen, in welchen die Plasmakonzentrationen an Glukose,
Insulin und Triglyceriden und low density lipoprotein (LDL)-Cholesterin ab- und an high density lipoprotein
(HDL)-Cholesterin zunahmen und die Körpermasse der Tiere moderat sank (Kharitonenkov et al.,
2007).
Vergleichbare positive Wirkungen auf verschiedene metabolische Parameter zeigen sich durch die
Verabreichung synthetischer PPARα-Agonisten, wie den Fibraten. So können sie ebenfalls eine
Verbesserung des Lipidprofils durch Triglycerid- und LDL-Cholesterin-senkende Wirkungen, eine
Erhöhung des HDL-Cholesterins sowie eine Verbesserung der Insulinsensitivität und hepatischen
Steatose in Nagern und Rhesusaffen hervorrufen (Chou et al., 2002; Tsuchida et al., 2005; Winegar et
al., 2001). Diese günstigen Wirkungen der Fibrate macht man sich seit einiger Zeit in der
Humantherapie zur Behandlung von Hypercholesterinämie und Hypertriglyceridämie zu nutze.
Die zentrale Funktion des PPARα in der Regulation des Lipidstoffwechsels und der Adaptation des
Stoffwechsels an das Fasten und Hungern durch die direkte Induktion der Transkription von Genen,
welche in die Aufnahme, den Transport und die Oxidation von Fettsäuren sowie in die
Ketonkörperproduktion involviert sind, ist hinreichend bekannt (Mandard et al., 2004).

Diskussion
46
Dass der hepatische FGF21 unter der Kontrolle des PPARα steht, konnte in verschiedenen
Untersuchungen belegt werden. So führte in Wildtyp-Mäusen eine durch Fasten oder WY-14,643
verursachte Aktivierung des PPARα zur Induktion des FGF21, während in PPARα-defizienten
Mäusen mit einer sehr niedrigen basalen Expression an FGF21 dieser Effekt ausblieb (Lundåsen et al.,
2007). In vitro stimulierte die Behandlung von humanen Primärhepatozyten mit WY-14,643 als
synthetischen PPARα-Agonisten die FGF21-mRNA Expression (Lundasen et al., 2007). Auch im
Menschen selbst ist eine Induktion des FGF21 nach PPARα-Aktivierung beschrieben. In
hypertriglyzeridämischen Patienten führte die Gabe von Fenofibrat neben der triglyceridsenkenden
Wirkung zu einer mäßigen, jedoch signifikanten Erhöhung der Konzentration an FGF21 im Serum
(Gälman et al., 2008). Ebenso konnte eine durch längeranhaltendes Fasten induzierte Erhöhung der
FGF21-Serumspiegel im Menschen nachgewiesen werden (Gälman et al., 2008). In einem weiteren
Fastenversuch mit Wildtyp-Mäusen sank die gesteigerte FGF21-mRNA-Expression durch Gabe von
Futter rasch zu ihren Ausgangswerten zurück (Inagaki et al., 2007; Badman et al., 2007). Putative
PPREs konnten in der Promotorregion des murinen und humanen FGF21-Gens identifiziert werden
(Lundasen et al., 2007).
Transgene Mäuse, welche FGF21 überexprimieren, sind geschützt vor diätinduzierter
Gewichtszunahme (Kharitonenkov et al., 2005). Im Vergleich zu Wildtyp-Mäusen zeigen sie eine
gesteigerte Ketonkörperproduktion und deutlich reduzierte Plasmakonzentrationen an Cholesterin,
Glukose und Insulin im Plasma (Inagaki et al., 2007).
In PPARα knock out- und leberspezifischen FGF21 knock down-Tieren kommt es zur Ausbildung
sehr ähnlicher Symptome. So zeigen Mäuse mit leberspezifischem FGF21-knock down unter einer
ketogenen Diät eine verminderte Expression der Gene der peroxisomalen β-Oxidation (z.B. ACO),
des Fettsäuretransportes ins Mitochondrium (z.B. L-Carnitin-Palmitoyltransferase (CPT)1) und der
Ketonkörpersynthese (mitochondriale 3-Hydroxy-3-Methylglutaryl-CoA Synthase (HMGCoAS2)),
welche mit der Entwicklung einer Fettleber, Lipämie, Abnahme an Ketonkörpern und Erhöhung von
Triglyceriden und Cholesterin im Serum einhergeht (Badman et al., 2007). Fastende PPARα knock
out-Mäuse leiden ebenfalls unter einer beeinträchtigten β-Oxidation und Ketogenese sowie
vermehrter hepatischer Lipidakkumulation, wodurch es zur Ausbildung von Hypoketonämie,
Hypoglykämie und hepatischer Steatose kommt (Hashimoto et al., 2000; Kersten et al., 1999; Leone et
al., 1999). Diese Symptome lassen sich durch eine systemische Verabreichung von rekombinantem
FGF21 vermindern. Dabei konnte in Wildtyp- und PPARα knock out-Mäusen eine Erhöhung der
Konzentrationen an β-Hydroxybutyrat und eine entsprechende Abnahme der
Triglyceridkonzentrationen im Serum beobachtet werden (Badman et al., 2007; Inagaki et al., 2007).
Diese zusammengefassten Literaturergebnisse deuten auf eine wesentliche Rolle des FGF21 als
PPARα-abhängigen Regulator der Ketogenese und Fettsäureoxidation hin.
Dass oxidierte Fette charakteristische Bestandteile enthalten, welche potente Liganden des
hepatischen PPARα darstellen, konnte in zahlreichen Untersuchungen an der Ratte und am Schwein

Diskussion
47
nachgewiesen werden (Luci et al., 2007; Sülzle et al., 2004). So konnte durch die Verabreichung
oxidierter Fette u. a. eine Erhöhung der Transkription klassischer PPARα-Zielgene, welche in den
Lipidstoffwechsel (ACO, L-CPT1, Stearoyl-CoA Desaturase (SCD)) und die Ketogenese
(HMGCoAS2) involviert sind, eine Senkung der Lipide (Triglyceride und Cholesterin) in Leber und
Plasma und eine Erhöhung des Ketonkörpers β-Hydroxybutyrat beobachtet werden (Chao et al.,
2001; Luci et al., 2007; Sülzle et al., 2004). Auch der FGF21 ist im Rahmen der Adaptation des
Stoffwechsels während des Fastens in die Regulation der hepatischen Glukoneogenese, Ketogenese
und Fettsäureoxidation und der Lipolyse im Fettgewebe involviert (Badman et al., 2007; Inagaki et al.,
2007; Potthoff et al., 2009). Die Ergebnisse dieser Untersuchungen zeigen, dass der PPARα in die
Regulation des hepatischen FGF21 involviert ist. Eine Aktivierung des PPARα durch Fasten, ketogene
Diät oder synthetische Agonisten führt zur Induktion des FGF21. Weiterhin ist auffällig, dass sowohl
FGF21 als auch oxidierte Fette verschiedene Wirkungen im Lipid- und Ketonkörperstoffwechsel
vermitteln und eine Vielzahl der hervorgerufenen metabolischen Effekte, welche über eine
Verabreichung von FGF21 verursacht werden, ebenso durch oxidierte Fette hervorgerufen werden.
Dazu zählen u. a. die Reduktion der Triglyzeridkonzentrationen im Serum und der Leber, die
Reduktion von Plasmacholesterin, Erhöhung der Konzentration an β-Hydroxybutyrat im Plasma und
die Verbesserung einer hepatischen Steatose. Sowohl der PPARα als auch der FGF21 sind in die
hepatische Stimulation der Fettsäureoxidation, Ketogenese und Glukoneogenese involviert.
In den eigenen Untersuchungen (Studie 3) konnte erstmals gezeigt werden, dass die Fütterung eines
moderat oxidierten Fettes den FGF21 in der Leber vom Schwein induziert. Die Erhöhung der FGF21-
Transkription und der FGF21-Proteinmenge zeigt eine Steigerung der Synthese dieses Proteins in der
Leber durch das oxidierte Fett. Da die Konzentration an FGF21 im Serum mit der hepatischen FGF21-
Genexpression positiv korreliert (Badman et al., 2007) und in den eigenen Untersuchungen eine
tendentielle Erhöhung der Proteinkonzentration an FGF21 im Plasma ermittelt werden konnte, ist
davon auszugehen, dass auch die hepatische Sekretion von FGF21 in die Zirkulation durch das
oxidierte Fett gesteigert wurde. Die Aktivierung des PPARα durch oxidiertes Fett, welche über eine
Erhöhung der Transkription typischer PPARα-Zielgene festgestellt wurde (ACO, L-CPT1, novel
organic cation transporter (OCTN)2), führte vermutlich zur Induktion des FGF21. Die beobachtete
ketogene Stoffwechselreaktion auf das oxidierte Fett, an der Erhöhung der Transkription des
Schlüsselenzyms der Ketogenese (HMGCoAS2) und der tendentiellen Erhöhung der Konzentration
des Ketonkörpers β-Hydroxybutyrat im Plasma zu erkennen, wird vermutlich auch, zumindest
teilweise über den FGF21 vermittelt.
Zusammenfassend kann daher festgestellt werden, dass einige der metabolischen Wirkungen, welche
durch die Verabreichung oxidierter Fette beobachtet werden können, anscheinend auch über eine
Induktion des FGF21 vermittelt werden.

Diskussion
48
Die vergleichsweise moderaten Effektstärken der Expression PPARα-relevanter Zielgene der eigenen
Untersuchung im Vergleich zu Nagern (Chao et al., 2001; Koch et al., 2007; Luci et al., 2007; Sülzle et
al., 2004) sind vermutlich sowohl in der generell niedrigeren Expression des PPARα im Schwein als
auch in der Verwendung eines nur moderat erhitzten Behandlungsfettes, wie an den ermittelten
Fettkennzahlen ersichtlich ist, begründet. Die Plasmakonzentrationen an FGF21 der Kontrollgruppe
der eigenen Untersuchung sind jedoch in sehr gutem Einklang mit Werten, wie sie für einen gesunden
Erwachsenen beschrieben werden konnten (Li et al., 2010).
Da der Mensch ebenso wie das Schwein zur nicht-proliferierenden Spezies gehört (S. 7-8) und der
FGF21 unter der Kontrolle des hepatischen PPARα liegt, ist mit hoher Wahrscheinlichkeit davon
auszugehen, dass die Aufnahme einer Kost reich an oxidierten Fetten auch im Menschen zur
Induktion des FGF21 führt.

Diskussion
49
4.4 Schlussfolgerungen
Der Einfluss oxidierter Fette auf den Organismus stellt trotz einer Vielzahl an bereits durchgeführten
Untersuchungen nach wie vor einen hochaktuellen Forschungsschwerpunkt im Bereich der
Humanernährung dar. Durch den anhaltend hohen und stetig steigenden Konsum an Lebensmitteln,
die oxidierte Fette enthalten und ihrem regulatorischen Einfluss auf den Metabolismus, besteht
gerade zu den Wirkmechanismen oxidierter Fette ein besonderes Interesse an der weiterführenden
Aufklärung.
Die Ergebnisse der vorliegenden Arbeit zeigen erstmals, dass oxidierte Fette ihre Wirkungen über
eine gleichzeitige, koordinierte Aktivierung der redox-sensitiven Transkriptionsfaktoren Nrf2 und
NF-κB vermitteln. In diesem Zusammenhang konnten auch eine gesteigerte Genexpressionen und
erhöhte Aktivitäten von Enzymen, welche antioxidative, detoxifizierende oder proinflammatorische
Funktionen übernehmen, ermittelt werden. Dabei konnte nachgewiesen werden, dass in beiden
untersuchten Spezies in den jeweiligen Organen die gleichen Mechanismen mit teilweise ähnlichen
Effektstärken unabhängig von der Art des oxidierten Fettes aktiviert werden. Dies hebt die
substantielle Bedeutung dieser grundlegenden Mechanismen in der zellulären Stressantwort hervor.
Somit stellen diese beiden Transkriptionsfaktoren wahrscheinlich die bisher unbekannten
funktionellen Bindeglieder zwischen dem durch die Aufnahme oxidierter Fette verursachten
oxidativen Stress und der Aktivierung zellulärer Stressreaktionen durch die Induktion antioxidativer,
detoxifizierender und proinflammatorischer Enzyme dar.
Um eine hohe Übertragbarkeit der erzielten Ergebnisse auf den Menschen zu gewährleisten, wurden
die beiden in dieser Arbeit ausgewählten Fette, welche in der menschlichen Ernährung auch zum
Frittieren von Lebensmitteln Einsatz finden, unter möglichst praxisnahen Bedingungen hergestellt.
Die im Schweinversuch (Studie 2 und 3) täglich verfütterte Menge an oxidiertem Fett, die mit 15 %
in der Diät rund 35-Energie % leistete, ist ebenfalls eine durchaus relevante Größe in der
Humanernährung. Extrapoliert man diese Werte auf den Menschen mit einem durchschnittlichen
Energiebedarf von 2000 kcal/d, so kann dieser über eine einzige Mahlzeit frittierter Lebensmittel mit
exemplarisch 200 g Geflügelnuggets (durchschnittlich 28 % Fett) und 200 g Pommes
(durchschnittlich 10 % Fett) die entsprechende Menge an oxidierten Fetten verzehren. Vor allem die
Ernährungsweise der westlichen Bevölkerung zeichnet sich aufgrund des hohen Fast Food-Konsums
durch einen hohen Anteil an oxidierten Fetten aus. Daher ist es für einen Menschen durchaus
möglich, täglich diese Mengen über die Nahrung aufzunehmen. Die im Schweineversuch
beobachteten Effekte oxidierter Fette wären demnach sehr wahrscheinlich auch im Menschen
nachweisbar.
Generell stellen die verwendeten Spezies Maus und Schwein in der wissenschaftlichen Praxis
etablierte Modelltiere dar und eignen sich besonders, um grundlegende Mechanismen zu erforschen.

Diskussion
50
Zudem zeichnen sie sich durch hohe Sequenzhomologien im Vergleich zum Menschen aus, so dass
von einer hohen Übertragbarkeit der erzielten Ergebnisse oxidierter Fette auf den Menschen
ausgegangen werden kann (Waterston et al., 2002; Swindle et al., 2012). Gleichwohl steht das
Schwein dem Menschen aufgrund genetischer, anatomischer und physiologischer Merkmale näher
und stellt damit das besser geeignetere Modelltier dar, um Rückschlüsse auf den Menschen ziehen zu
können. So bestehen auch hinsichtlich der Expression des PPARα Unterschiede zwischen der Maus
als Vertreter der proliferierenden Spezies und dem Schwein als Vertreter der nicht-proliferierenden
Spezies, so dass hier die Übertragbarkeit der Ergebnisse von der Maus auf den Menschen begrenzt ist
(siehe S. 7).
Die Aktivierung des Nrf2 und seiner ARE-abhängigen Zielgene im Organismus wird in der
wissenschaftlichen Literatur überwiegend als günstig bewertet. In diesem Zusammenhang wird
häufig die Hormesistheorie angeführt, in welcher der Nrf2 eine Schlüsselrolle spielt. Diese Theorie
besagt, dass die Toxizität eines Stoffes von dessen Konzentration abhängig ist, wobei geringe
Konzentrationen eine positive Wirkung haben können, da sie zur Adaptation an den milden oder
transienten Stress eine zelluläre Stressantwort im Organismus auslösen. Diese Stoffe, auch Stressoren
genannt, zu denen z.B. Flavonoide und Isothiocyanate zählen, aktivieren den Nrf2 und die ARE-
abhängige Genexpression, wodurch z. B. antioxidative und Phase II-Enzyme aktiviert werden und die
zytoprotektive Kapazität der Zelle ansteigt. Dabei spielt häufig die gleichzeitige Inhibierung des
NF-κB eine Rolle, wobei die zugrunde liegenden Signalkaskaden bisher nur unzureichend verstanden
sind. Übersteigt die Konzentration der Stressoren jedoch ein gewisses Maß, reicht die Kapazität der
zellulären Schutzmechanismen nicht mehr aus und es kann zu negativen Auswirkungen kommen.
Da jede Nahrungsaufnahme per se einen gewissen oxidativen Reiz auslöst, herrscht im Darm eine
basale Nrf2-Aktivierung vor, welche essentiell für die Aufrechterhaltung der Darmintegrität und zum
Schutz des Organismus vor schädlichen Fremdstoffen und oxidativen Stress ist. Die in Studie 1
beobachtete Nrf2-Aktivierung im Darm durch oxidierte Fette könnte daher noch als normale
physiologische Stoffwechselreaktion gewertet werden. Da allerdings auch eine Aktivierung des
NF-κB und die Induktion proinflammatorischer Enzyme festgestellt werden konnte, kann daraus
geschlossen werden, dass der durch das oxidierte Fett ausgelöste oxidative Stress ein gewisses Maß
überstieg und es zur Einleitung inflammatorischer Prozesse kam. Die Aktivierung der redox-
sensitiven Signalwege in der Leber in Studie 2 deutet darauf hin, dass die antioxidative Kapazität im
Darm nicht ausreichend war und Stressoren in den Organismus gelangen konnten. Die in der Leber
ermittelte Aktivierung antioxidativer Enzyme und der Phase II-Enzyme des Fremdstoffmetabolismus
zeigt, dass der Organismus bestrebt ist, die in den Stoffwechsel gelangten oxidierten Fette und deren
Oxidationsprodukte zu detoxifizieren bzw. das durch vermehrte ROS-Bildung gestörte
Redoxpotential der Zelle über die Induktion der entsprechenden Enzyme wieder herzustellen. Das
Ausmaß der Enzymaktivierung war dabei so stark, dass Auswirkungen auf den

Diskussion
51
Schilddrüsenhormonstatus ermittelt werden konnten. Ein deutliches Zeichen für verstärkten
oxidativen Stress war auch die Abnahme der Vitamin E-Konzentrationen in der Leber und im Plasma.
Da in den eigenen Studien mit oxidierten Fetten neben der Nrf2-Aktivierung auch eine Aktivierung
des NF-κB festgestellt werden konnte, ist davon auszugehen, dass oxidierte Fette ein höheres Maß an
oxidativem Stress als die anderen o. g. Nrf2-Induktoren auslösen. Die koordinierte Aktivierung der
beiden Transkriptionsfaktoren im Darm und der Leber diente wahrscheinlich der Adaptation an den
gesteigerten oxidativen Stress und der Protektion der Organe, um ROS-vermittelte oxidative
Schädigungen durch die Aufnahme der oxidierten Fette zu verhindern.
Erstmals konnte im Rahmen der vorliegenden Arbeit auch gezeigt werden, dass die Aufnahme eines
moderat oxidierten Fettes zur Induktion des hepatischen FGF21 führt, welche mit großer
Wahrscheinlichkeit über eine Aktivierung des PPARα vermittelt wurde. Somit besitzen oxidierte
Fette ebenso wie freie Fettsäuren als natürliche und Fibrate als synthetische Agonisten des PPARα
das Potential, den FGF21 zu induzieren. Einige der in Tierstudien beobachteten metabolischen
Wirkungen oxidierter Fette, wie eine gesteigerte Ketonkörperproduktion oder verminderte
Plasmalipidkonzentrationen, könnten daher auch über diesen neuen Mechanismus vermittelt werden.
Aufgrund der in Tierstudien beobachteten günstigen metabolischen Wirkungen nach der
Verabreichung von rekombinantem FGF21, wie der Reduktion der Blutlipide oder des
Körpergewichtes, stellt sich die Frage nach dessen therapeutischen Potential auch in der Therapie
von Humanerkrankungen, wie der Fettleibigkeit oder dem Diabetes mellitus Typ 2. Aus Tier- und
Humanstudien ist bekannt, dass in Zuständen der Insulinresistenz die FGF21-Serumwerte erhöht
sind, was auf einen Kompensationsmechanismus oder eine Resistenz ähnlich des Insulins hinweist
(Chavez et al., 2009; Fisher et al., 2010). Im Tierversuch wurde FGF21 zur Behandlung
hyperglykämischer und hyperlipidämischer Zustände schon erfolgreich eingesetzt, jedoch zeigten nur
pharmakologisch hohe Dosen Wirkung. Im Menschen stehen derartige Untersuchungen derzeit noch
aus. Allerdings wird hier schon der Einsatz des FGF21 in der Diagnostik als frühzeitiger Biomarker
zur Detektion metabolischer Störungen diskutiert. Um sich die positiven metabolischen Wirkungen
einer durch oxidierte Fette verursachten FGF21-Aktivierung nutzbar machen zu können, müssten
zuvor in weiteren Untersuchungen die dafür verantwortlichen, biologisch aktiven Bestandteile der
oxidierten Fettgemische identifiziert und Effektivdosen ermittelt werden, um anschließend einen
Einsatz solcher Isolate abzuleiten. Eine Verabreichung oxidierte Fettgemische per se würde aufgrund
ihrer weiteren, teils auch negativen Wirkungen nicht zweckmäßig sein.
Zusammenfassend konnte anhand der in der vorliegenden Arbeit durchgeführten Untersuchungen
gezeigt werden, dass Bestandteile oxidierter Fette die Fähigkeit besitzen über die Einleitung
zellulärer Signalwege regulatorische Aktivitäten im Stoffwechsel zu vollziehen. Durch die
Aktivierung der redox-sensitiven Transkriptionsfaktoren Nrf2 und NF-κB sowie des
hormonähnlichen, metabolischen Regulators FGF21 konnten neue molekulare Mechanismen

Diskussion
52
aufgezeigt werden, über welche oxidierte Fette ihre biologischen Wirkungen im Organismus
vermitteln.

Zusammenfassung
53
5. Zusammenfassung
Das Ziel der vorliegenden Arbeit war die Aufklärung molekularer Mechanismen, über die oxidierte
Fette ihre Wirkungen im Organismus vermitteln.
Oxidierte Fette sind chemisch veränderte Fette, wie sie in erhitzten und unsachgemäß gelagerten
Lebensmitteln zu finden sind. Insbesondere während des Frittierprozesses laufen verschiedenste
chemische Reaktionen ab, wobei große Mengen an Oxidationsprodukten gebildet und vom
Frittiergut absorbiert werden. Eine bedeutende Quelle oxidierter Fette in der menschlichen
Ernährung stellen daher frittierte Lebensmittel dar. Aus Tier- und Humanstudien ist bekannt, dass
die Aufnahme oxidierter Fette im Organismus verschiedenste biologische Auswirkungen haben kann.
Eine der bedeutendsten ist die Induktion von oxidativem Stress durch die Generierung von reaktiven
Sauerstoffspezies, welcher zur Aktivierung der antioxidativen Abwehr, detoxifizierender
Mechanismen und auch proinflammatorischer Prozesse führen kann. In diesem Zusammenhang
konnte eine verstärkte Expression redox-sensitiver Gene, welche u. a. für antioxidative Enzyme,
proinflammatorische Zytokine und Enzyme des Fremdstoffmetabolismus kodieren, sowie gesteigerte
Enzymaktivitäten beobachtet werden. Die Mechanismen, die diesen Reaktionen zugrunde liegen,
sind bisher nicht vollständig geklärt.
Zwei sehr bedeutende redox-sensitive Transkriptionsfaktoren, welche durch oxidativen Stress
aktiviert werden können und in die Regulation der zellulären Stressantwort involviert sind, sind der
Nrf2 und NF-κB. Aus diesem Grunde wurde im ersten Teil dieser Arbeit dem Einfluss oxidierter Fette
auf die Aktivierung dieser beiden Transkriptionsfaktoren nachgegangen.
Um möglichst praxisnahe Versuchsbedingungen zu schaffen und damit eine hohe Übertragbarkeit
der erzielten Ergebnissen auf den Menschen zu gewährleisten, wurden in dieser Arbeit zwei
Versuchsfette (Sonnenblumenöl und Rapsöl) verwendet, die auch in der menschlichen Ernährung
Einsatz zum Frittieren von Lebensmitteln finden. Zudem erfolgte die Erhitzung der Behandlungsfette
unter Frittierbedingungen.
Studie 1: Das Ziel der ersten Untersuchung am Modelltier Maus war daher die Überprüfung der
Hypothese, dass die Verabreichung eines oxidierten Fettes (Sonnenblumenöl) zu einer Aktivierung
der redox-sensitiven Transkriptionsfaktoren NF-κB und Nrf2 im Darm führt. Weiterhin sollte der
Einfluss des oxidierten Fettes auf die Aktivierung der Transkriptionsfaktoren PPARα und PPARγ und
der PPARα/γ-responsiven Genexpression sowie einer möglichen negativen Interferenz mit dem
NF-κB ermittelt werden.
Die Ergebnisse zeigten erstmals eine Aktivierung des Nrf2 und NF-κB/p50 durch die Verabreichung
eines oxidierten Fettes in der Dünndarmmukosa der Maus. In diesem Zusammenhang konnte eine
gesteigerte zelluläre Stressantwort anhand erhöhter mRNA-Konzentrationen oxidativer Stress-

Zusammenfassung
54
responsiver Gene beobachtet werden. Weiterhin verursachte das oxidierte Fett einen starken Anstieg
der mRNA-Konzentrationen an PPARα sowie PPARα/γ-responsiver Gene. Kein Einfluss konnte auf
der mRNA-Konzentration des PPARγ festgestellt werden.
Studie 2: Basierend auf den Ergebnissen der ersten Untersuchung wurde im zweiten Versuch
überprüft, ob sich diese durch Aufnahme eines oxidierten Fettes ausgelösten Effekte auch im
Schwein, das hinsichtlich der anatomischer und stoffwechselphysiologischer Merkmale ein
geeigneteres Modelltier für den Menschen als die Maus darstellt, nachweisen lassen. Dazu wurde der
Hypothese nachgegangen, dass die Aufnahme eines moderat oxidierten Fettes (Rapsöl) auch in der
Leber des Schweins zu einer Aktivierung der redox-sensitiven Transkriptionsfaktoren Nrf2 und
NF-κB führt. Auch in dieser Untersuchung konnte durch oxidiertes Fett eine Aktivierung des Nrf2
und NF-κB/p50 nachgewiesen werden. Als Folge der Aktivierung wurden erhöhte Enzymaktivitäten
und erhöhte mRNA-Konzentrationen von Genen, welche in die antioxidative Abwehr oder in die
Phase II des Fremdstoffmetabolismus involviert sind, beobachtet. Über die Induktion des
Abbauprozesses von Schilddrüsenhormonen wirkte das oxidierte Fett auch auf den Gesamtthyroxin-
Status. Dass oxidiertes Fett den Antioxidantien-Status ungünstig beeinflusste, konnte anhand
verringerter Vitamin E-Konzentrationen in Leber und Plasma verdeutlicht werden.
Insgesamt konnte in diesen beiden Untersuchungen (Studie 1 und 2) erstmals eine Aktivierung der
redox-sensitiven Transkriptionsfaktoren Nrf2 und NF-κB durch oxidierte Fette im Darm und der
Leber festgestellt werden. Somit stellen diese beiden Transkriptionsfaktoren wahrscheinlich die
bisher unbekannten funktionellen Bindeglieder zwischen dem durch die Aufnahme oxidierter Fette
verursachten oxidativen Stress und der Aktivierung zellulärer Stressreaktionen durch die Induktion
antioxidativer, detoxifizierender und proinflammatorischer Enzyme dar.
Die koordinierte Aktivierung dieser Signalwege durch die Aufnahme oxidierter Fette diente offenbar
der Adaptation der Organe an den hervorgerufenen oxidativen Stress und ihrer Protektion vor
oxidativen Schädigungen.
Studie 3: Im zweiten Teil dieser Arbeit wurde der Hypothese nachgegangen, dass die Verabreichung
eines moderat oxidierten Fettes (Rapsöl) den endokrinen Faktor FGF21 in der Leber vom Schwein
aktiviert. Der FGF21 wurde kürzlich als neuartiger hormonähnlicher Regulator im Stoffwechsel
identifiziert. Seine physiologische Bedeutung liegt in der Kontrolle der Lipid- und Glukose-
homöostase, indem er die Glukoneogenese, die Ketogenese und die Fettsäureoxidation induziert. Die
hepatische Regulation des FGF21 unterliegt der direkten Kontrolle des PPARα. So konnte in
bisherigen Studien gezeigt werden, dass eine durch Fasten, ketogene Diäten und synthetische
PPARα-Agonisten verursachte Aktivierung des PPARα den FGF21 in der Leber induzieren kann.
Inwiefern auch oxidierte Fette als potente nutritive Aktivatoren des PPARα den FGF21 zu induzieren
vermögen, wurde bisher nicht untersucht.

Zusammenfassung
55
Erstmals konnte in dieser Arbeit gezeigt werden, dass die Aufnahme eines oxidierten Fettes nicht nur
zu einem Anstieg der mRNA-Konzentration PPARα-relevanter Zielgene, sondern auch in einer
gesteigerten Expression des FGF21 in der Leber resultierte. Sehr wahrscheinlich führte die
Aktivierung des PPARα, welche über eine erhöhte Expression typischer PPARα-Zielgene festgestellt
wurde, zur Induktion des FGF21. Über diesen neu beschriebenen Mechanismus werden
möglicherweise einige der aus Tierstudien bekannten metabolischen Wirkungen oxidierter Fette, wie
eine gesteigerte Ketonkörperproduktion oder verminderte Plasmalipidkonzentrationen, vermittelt.
Zusammenfassend konnte anhand der in der vorliegenden Arbeit durchgeführten Untersuchungen
gezeigt werden, dass Bestandteile oxidierter Fette die Fähigkeit besitzen über die Einleitung
zellulärer Signalwege regulatorische Aktivitäten im Stoffwechsel zu vollziehen. Durch die
Aktivierung der redox-sensitiven Transkriptionsfaktoren Nrf2 und NF-κB sowie des
hormonähnlichen, metabolischen Regulators FGF21 konnten neue molekulare Mechanismen
aufgezeigt werden, über welche oxidierte Fette ihre biologischen Wirkungen im Organismus
vermitteln.

Summary
56
6. Summary
The aim of the present work was to clarify molecular mechanisms by which oxidized fats mediate
their effects in organism.
Oxidized fats are chemically modified fats that can be found in heated and incorrectly stored food
items. Especially, during the process of deep-frying several chemical reactions occur that generate
large amount of oxidation products which are absorbed by the deep-fried food. Therefore, deep-fried
food is an important source of oxidized fats in human nutrition. Studies on animals and humans
revealed that ingestion of oxidized fats provokes a wide array of biological effects in organism. One of
the most striking effects mediated by oxidized fats is the induction of oxidative stress by the
generation of reactive oxygen species leading to the activation of antioxidative defense, detoxifying
mechanisms, and proinflammatory processes. In this context, an increased expression of redox-
sensitive genes encoding for antioxidative enzymes, proinflammatory enzymes, and enzymes of
phase II detoxification, and increased enzyme activities could be observed. However, till now the
underlying mechanisms of these observations are not completely clarified.
Nrf2 and NF-κB, both being very important redox-sensitive transcription factors, can be activated by
oxidative stress and are involved in the regulation of cellular stress response. For this reason, the first
part of this work aimed to investigate the influence of oxidized fats on the activation of these two
transcription factors. In order to ensure practically relevant conditions of the experiments and, hence,
a high transferability of the study results with regard to human, two experimental fats, which are also
used for deep-frying of food in human nutrition, were used in this work. Furthermore, the treatment
fats were heated under deep-frying conditions.
Study 1: The aim of the first study was to test the hypothesis, that ingestion of an oxidized fat
(sunflower oil) causes an activation of the redox-sensitive transcription factors NF-κB and Nrf2 in
intestine of mice. Furthermore, the influence of the oxidized fat on the activation of the transcription
factors PPARα and PPARγ and the PPARα/γ-responsive gene expression and a possible negative
interference with NF-κB should be investigated.
The results showed for the first time the activation of Nrf2 und NF-κB/p50 by administration of an
oxidized fat in small intestinal mucosa of mice. In this context, an enhanced cellular stress response
was determined by an increased mRNA concentration of oxidative stress-responsive genes.
Furthermore, the oxidized fat led to a strong increase in the mRNA concentration of PPARα and
PPARα/γ-responsive genes whereas mRNA concentration of PPARγ was unaffected.
Study 2: Based on the findings of the first study, the second experiment aimed to examine whether
the effects caused by administration of an oxidized fat, can also be demonstrated in the pig which is in
regard to anatomic and metabolic physiology characteristics a more suitable model for human than
the mouse model.

Summary
57
Therefore, the hypothesis was tested, that ingestion of a moderately oxidized fat (rapeseed oil) also
activates the redox-sensitive transcription factors Nrf2 und NF-κB in the liver of pigs. Also an
activation of Nrf2 and NF-κB by oxidized fat could be demonstrated in this study. As a consequence
of activation, increased enzyme activities and increased mRNA concentration of genes involved in
antioxidative defense or phase II metabolism were observed. The oxidized fat also influenced the total
thyroxin status by induction of thyroid hormone elimination processes. That oxidized fat
compromised status of antioxidants become obvious in decreased concentrations of vitamin E in liver
and plasma.
For the first time, in the present studies (study 1 and 2) an activation of the redox-sensitive
transcription factors Nrf2 and NF-κB by oxidized fats in the intestine and the liver was determined.
Thus, these two transcription factors probably are the unknown functional link between oxidative
stress caused by ingestion of oxidized fats and activation of cellular stress response by induction of
antioxidative, detoxifying and proinflammatory enzymes. The coordinated activation of these
signaling pathways by intake of oxidized fats likely conduced to the adaptation of organs to the
generated oxidative stress and their protection of oxidative damage.
Study 3: The second part of this work aimed to investigate the hypothesis that administration of a
moderately oxidized fat (rapeseed oil) activates the endocrine factor FGF21 in the liver of pigs. FGF21
has been recently identified as a novel hormone-like regulator in metabolism. Its physiological
importance lies in the control of lipid and glucose homeostasis by the induction of gluconeogenesis,
ketogenesis, and fatty acid oxidation. Hepatic induction of FGF21 is under direct control of PPARα.
Previous studies have shown that an activation of PPARα caused by fasting, ketogenic diets, and
synthetic PPARα-agonists can induce FGF21 in the liver. Whether oxidized fats as potent nutritive
activators of PPARα are also able to induce FGF21, has not been investigated yet.
The present study showed for the first time that administration of an oxidized fat not only led to an
increase in mRNA concentration of PPARα-relevant target genes but also results in an increased
expression of FGF21 in the liver. Activation of PPARα determined by increased expression of typical
PPARα target genes induced FGF21 very likely. Possibly, some of the metabolic effects of oxidized fats
known from animal studies, like increased production of ketone bodies or decreased concentration of
lipids in plasma, are mediated by this new described mechanism.
To summarize, the experiments performed in the present work revealed that components of oxidized
fats have the ability to exert regulatory activities in metabolism by induction of cellular signaling
pathways. New molecular mechanisms by which oxidized fats mediate their biological effects in
organism could be demonstrated by the activation of the redox-sensitive transcription factors Nrf2
and NF-kB and the hormone-like, metabolic regulator FGF21.

Literaturverzeichnis
58
7. Literaturverzeichnis
Ardite E, Panés J, Miranda M, Salas A, Elizalde JI, Sans M, Arce Y, Bordas JM, Fernández-Checa JC,
Piqué JM. 1998. Effects of steroid treatment on activation of nuclear factor kappaB in patients
with inflammatory bowel disease. Br J Pharmacol. 124:431-3.
Badman MK, Pissios P, Kennedy AR, Koukos G, Flier JS, Maratos-Flier E. 2007. Hepatic fibroblast
growth factor 21 is regulated by PPARalpha and is a key mediator of hepatic lipid metabolism
in ketotic states. Cell Metab. 5:426-37.
Baird L, Dinkova-Kostova AT. 2011. The cytoprotective role of the Keap1-Nrf2 pathway. Arch Toxicol.
85:241-72.
Banning A, Deubel S, Kluth D, Zhou Z, Brigelius-Flohé R. 2005. The GI-GPx gene is a target for Nrf2.
Mol Cell Biol. 25:4914-23.
Barnes PJ. 1997. Nuclear factor-kappa B. Int J Biochem Cell Biol. 29:867-70.
Barth CA, Pfeuffer M, Scholtissek J. 1990. Animal models for the study of lipid metabolism, with
particular reference to the Göttingen minipig. Adv Anim Physiol Anim Nutr S. 20:39-49.
Bendixen E, Danielsen M, Larsen K, Bendixen C. 2010. Advances in porcine genomics and proteomics-
-a toolbox for developing the pig as a model organism for molecular biomedical research. Brief
Funct Genomics. 9(3):208-19.
Bergamini CM, Gambetti S, Dondi A, Cervellati C. 2004. Oxygen, reactive oxygen species and tissue
damage. Curr Pharm Des. 10:1611-26.
Berglund ED, Li CY, Bina HA, Lynes SE, Michael MD, Shanafelt AB, Kharitonenkov A, Wasserman
DH. 2009. Fibroblast growth factor 21 controls glycemia via regulation of hepatic glucose flux
and insulin sensitivity. Endocrinology. 150:4084-93.
Bode G, Clausing P, Gervais F, Loegsted J, Luft J, Nogues V, Sims J; Steering Group of the RETHINK
Project. 2010. The utility of the minipig as an animal model in regulatory toxicology. J
Pharmacol Toxicol Methods. 62:196-220.
Braissant O, Foufelle F, Scotto C, Dauça M, Wahli W. 1996. Differential expression of peroxisome
proliferator-activated receptors (PPARs): tissue distribution of PPAR-alpha, -beta, and -
gamma in the adult rat. Endocrinology. 137:354-66.
Brandsch C, Nass N, Eder K. 2004. A thermally oxidized dietary oil does not lower the activities of
lipogenic enzymes in mammary glands of lactating rats but reduces the milk triglyceride
concentration. J Nutr. 134:631-6.
Buckley DB, Klaassen CD. 2009. Induction of mouse UDP-glucuronosyltransferase mRNA expression
in liver and intestine by activators of aryl-hydrocarbon receptor, constitutive androstane
receptor, pregnane X receptor, peroxisome proliferator-activated receptor alpha, and nuclear
factor erythroid 2-related factor 2. Drug Metab Dispos. 37:847-56.

Literaturverzeichnis
59
Camandola S, Poli G, Mattson MP. 2000. The lipid peroxidation product 4-hydroxy-2,3-nonenal
increases AP-1-binding activity through caspase activation in neurons. J Neurochem. 74:159-
68.
Chao PM, Chao CY, Lin FJ, Huang C. 2001. Oxidized frying oil up-regulates hepatic acyl-CoA oxidase
and cytochrome P450 4 A1 genes in rats and activates PPARalpha. J Nutr. 131:3166-74.
Chao PM, Huang HL, Liao CH, Huang ST, Huang CJ. 2007. A high oxidised frying oil content diet is
less adipogenic, but induces glucose intolerance in rodents. Br J Nutr. 98:63-71.
Chavez AO, Molina-Carrion M, Abdul-Ghani MA, Folli F, Defronzo RA, Tripathy D. 2009.
Circulating fibroblast growth factor-21 is elevated in impaired glucose tolerance and type 2
diabetes and correlates with muscle and hepatic insulin resistance. Diabetes Care. 32(8):1542-
6.
Chen YY, Chen CM, Chao PY, Chang TJ, Liu JF. 2005. Effects of frying oil and Houttuynia cordata
thunb on xenobiotic-metabolizing enzyme system of rodents. World J Gastroenterol. 11:389-
92.
Cheon Y, Nara TY, Band MR, Beever JE, Wallig MA, Nakamura MT. 2005. Induction of overlapping
genes by fasting and a peroxisome proliferator in pigs: evidence of functional PPARalpha in
nonproliferating species. Am J Physiol Regul Integr Comp Physiol. 288:1525-35.
Choe E, Min DB. 2007. Chemistry of deep-fat frying oils. J Food Sci. 72:77-86.
Chou CJ, Haluzik M, Gregory C, Dietz KR, Vinson C, Gavrilova O, Reitman ML. 2002. WY14,643, a
peroxisome proliferator-activated receptor alpha (PPARalpha ) agonist, improves hepatic and
muscle steatosis and reverses insulin resistance in lipoatrophic A-ZIP/F-1 mice. J Biol Chem.
277:24484-9.
Chung SW, Kang BY, Kim SH, Pak YK, Cho D, Trinchieri G, Kim TS. 2000. Oxidized low density
lipoprotein inhibits interleukin-12 production in lipopolysaccharide-activated mouse
macrophages via direct interactions between peroxisome proliferator-activated receptor-
gamma and nuclear factor-kappa B. J Biol Chem. 275:32681-7.
Cohn JS. 2002. Oxidized fat in the diet, postprandial lipaemia and cardiovascular disease. Curr Opin
Lipidol. 13:19-24.
Coskun T, Bina HA, Schneider MA, Dunbar JD, Hu CC, Chen Y, Moller DE, Kharitonenkov A. 2008.
Fibroblast growth factor 21 corrects obesity in mice. Endocrinology. 149:6018-27.
Delerive P, Fruchart JC, Staels B. 2001. Peroxisome proliferator-activated receptors in inflammation
control. J Endocrinol. 169:453-9.
Delerive P, Furman C, Teissier E, Fruchart J, Duriez P, Staels B. 2000. Oxidized phospholipids activate
PPARalpha in a phospholipase A2-dependent manner. FEBS Lett. 471:34-8.
Delerive P, De Bosscher K, Besnard S, Vanden Berghe W, Peters JM, Gonzalez FJ, Fruchart JC, Tedgui
A, Haegeman G, Staels B. 1999. Peroxisome proliferator-activated receptor alpha negatively
regulates the vascular inflammatory gene response by negative cross-talk with transcription
factors NF-kappaB and AP-1. J Biol Chem. 274:32048-54.

Literaturverzeichnis
60
Deutsche Gesellschaft für Fettwissenschaft e. V. 2000. 3rd International Symposium on Deep Fat
Frying.
Dreger H, Westphal K, Weller A, Baumann G, Stangl V, Meiners S, Stangl K. 2009. Nrf2-dependent
upregulation of antioxidative enzymes: a novel pathway for proteasome inhibitor-mediated
cardioprotection. Cardiovasc Res. 83:354-61.
Eder K, Kirchgessner M. 1998. The effect of dietary vitamin E supply and a moderately oxidized oil on
activities of hepatic lipogenic enzymes in rats. Lipids. 33:277-83.
Eder K, Stangl GI. 2000. Plasma thyroxine and cholesterol concentrations of miniature pigs are
influenced by thermally oxidized dietary lipids. J Nutr. 130:116-21.
Eder K, Skufca P, Brandsch C. 2002. Thermally oxidized dietary fats increase plasma thyroxine
concentrations in rats irrespective of the vitamin E and selenium supply. J Nutr. 132:1275-81.
Eder K, Keller U, Hirche F, Brandsch C. 2003a. Thermally oxidized dietary fats increase the
susceptibility of rat LDL to lipid peroxidation but not their uptake by macrophages. J Nutr.
133:2830-7.
Eder K, Suelzle A, Skufca P, Brandsch C, Hirche F. 2003b. Effects of dietary thermoxidized fats on
expression and activities of hepatic lipogenic enzymes in rats. Lipids. 38:31-8.
Fisher FM, Chui PC, Antonellis PJ, Bina HA, Kharitonenkov A, Flier JS, Maratos-Flier E. 2010.
Obesity is a fibroblast growth factor 21 (FGF21)-resistant state. Diabetes. 59(11):2781-9.
Fourquet S, Guerois R, Biard D, Toledano MB. 2010. Activation of NRF2 by nitrosative agents and
H2O2 involves KEAP1 disulfide formation. J Biol Chem. 285:8463-71.
Gälman C, Lundåsen T, Kharitonenkov A, Bina HA, Eriksson M, Hafström I, Dahlin M, Amark P,
Angelin B, Rudling M. 2008. The circulating metabolic regulator FGF21 is induced by
prolonged fasting and PPARalpha activation in man. Cell Metab. 8:169-74.
Gao L, Wang J, Sekhar KR, Yin H, Yared NF, Schneider SN, Sasi S, Dalton TP, Anderson ME, Chan JY,
Morrow JD, Freeman ML. 2006. Novel n-3 fatty acid oxidation products activate Nrf2 by
destabilizing the association between Keap1 and Cullin3. J Biol Chem. 282:2529-37.
Garbin U, Fratta Pasini A, Stranieri C, Cominacini M, Pasini A, Manfro S, Lugoboni F, Mozzini C,
Guidi G, Faccini G, Cominacini L. 2009. Cigarette smoking blocks the protective expression
of Nrf2/ARE pathway in peripheral mononuclear cells of young heavy smokers favouring
inflammation. PLoS One. 4:8225.
Gloire G, Dejardin E, Piette J. 2006. Extending the nuclear roles of IkappaB kinase subunits. Biochem
Pharmacol. 72:1081-9.
Gulick T, Cresci S, Caira T, Moore DD, Kelly DP. 1994. The peroxisome proliferator-activated
receptor regulates mitochondrial fatty acid oxidative enzyme gene expression. Proc Natl Acad
Sci U S A. 91:11012-6.
Halliwell B. 2007. Biochemistry of oxidative stress. Biochem Soc Trans. 35:1147-50.
Hashimoto T, Cook WS, Qi C, Yeldandi AV, Reddy JK, Rao MS. 2000. Defect in peroxisome
proliferator-activated receptor alpha-inducible fatty acid oxidation determines the severity of
hepatic steatosis in response to fasting. J Biol Chem. 275:28918-28.

Literaturverzeichnis
61
Hayden MS, Ghosh S. 2004. Signaling to NF-kappaB. Genes Dev. 18:2195-224.
Heiss E, Herhaus C, Klimo K, Bartsch H, Gerhäuser C. 2001. Nuclear factor kappa B is a molecular
target for sulforaphane-mediated anti-inflammatory mechanisms. J Biol Chem. 276:32008-15.
Herfarth H, Brand K, Rath HC, Rogler G, Schölmerich J, Falk W. 2000. Nuclear factor-kappa B
activity and intestinal inflammation in dextran sulphate sodium (DSS)-induced colitis in mice
is suppressed by gliotoxin. Clin Exp Immunol. 120:59-65.
Huang CJ, Cheung NS, Lu VR. 1998. Effects of deteriorated frying oil and dietary protein levels on
liver microsomal enzymes in rats. JAOCS. 11:1796-1803.
Inagaki T, Dutchak P, Zhao G, Ding X, Gautron L, Parameswara V, Li Y, Goetz R, Mohammadi M,
Esser V, Elmquist JK, Gerard RD, Burgess SC, Hammer RE, Mangelsdorf DJ, Kliewer SA.
2007. Endocrine regulation of the fasting response by PPARalpha-mediated induction of
fibroblast growth factor 21. Cell Metab. 5:415-25.
Inamdar NM, Ahn YI, Alam J. 1996. The heme-responsive element of the mouse heme oxygenase-1
gene is an extended AP-1 binding site that resembles the recognition sequences for MAF and
NF-E2 transcription factors. Biochem Biophys Res Commun. 221:570-6.
Ishii T, Itoh K, Ruiz E, Leake DS, Unoki H, Yamamoto M, Mann GE. 2004. Role of Nrf2 in the
regulation of CD36 and stress protein expression in murine macrophages: activation by
oxidatively modified LDL and 4-hydroxynonenal. Circ Res. 94:609-16.
Itoh K, Wakabayashi N, Katoh Y, Ishii T, Igarashi K, Engel JD, Yamamoto M. 1999. Keap1 represses
nuclear activation of antioxidant responsive elements by Nrf2 through binding to the amino-
terminal Neh2 domain. Genes Dev. 13:76-86.
Itoh K, Chiba T, Takahashi S, Ishii T, Igarashi K, Katoh Y, Oyake T, Hayashi N, Satoh K, Hatayama I,
Yamamoto M, Nabeshima Y. 1997. An Nrf2/small Maf heterodimer mediates the induction of
phase II detoxifying enzyme genes through antioxidant response elements. Biochem Biophys
Res Commun. 236:313-22.
Izumiya Y, Bina HA, Ouchi N, Akasaki Y, Kharitonenkov A, Walsh K. 2008. FGF21 is an Akt-
regulated myokine. FEBS Lett. 582:3805-10.
Jaiswal AK. 2004. Nrf2 signaling in coordinated activation of antioxidant gene expression. Free Radic
Biol Med. 36:1199-207.
Johansen C, Flindt E, Kragballe K, Henningsen J, Westergaard M, Kristiansen K, Iversen L. 2005.
Inverse regulation of the nuclear factor-kappaB binding to the p53 and interleukin-8 kappaB
response elements in lesional psoriatic skin. J Invest Dermatol. 124:1284-92.
Kämmerer I, Ringseis R, Eder K. 2011. Feeding a thermally oxidised fat inhibits atherosclerotic plaque
formation in the aortic root of LDL receptor-deficient mice. Br J Nutr. 105:190-9.
Karuri AR, Huang Y, Bodreddigari S, Sutter CH, Roebuck BD, Kensler TW, Sutter TR. 2006. 3H-1,2-
dithiole-3-thione targets nuclear factor kappaB to block expression of inducible nitric-oxide
synthase, prevents hypotension, and improves survival in endotoxemic rats. J Pharmacol Exp
Ther. 317:61-7.

Literaturverzeichnis
62
Kato Y, Ikushiro S, Emi Y, Tamaki S, Suzuki H, Sakaki T, Yamada S, Degawa M. 2008. Hepatic UDP-
glucuronosyltransferases responsible for glucuronidation of thyroxine in humans. Drug Metab
Dispos. 36:51-5.
Kawahito Y, Kondo M, Tsubouchi Y, Hashiramoto A, Bishop-Bailey D, Inoue K, Kohno M, Yamada R,
Hla T, Sano H. 2000. 15-deoxy-delta(12,14)-PGJ(2) induces synoviocyte apoptosis and
suppresses adjuvant-induced arthritis in rats. J Clin Invest. 106:189-97.
Keller U, Brandsch C, Eder K. 2004. The effect of dietary oxidized fats on the antioxidant status of
erythrocytes and their susceptibility to haemolysis in rats and guinea pigs. J Anim Physiol
Anim Nutr (Berl). 88(:59-72.
Kersten S, Seydoux J, Peters JM, Gonzalez FJ, Desvergne B, Wahli W. 1999. Peroxisome proliferator-
activated receptor alpha mediates the adaptive response to fasting. J Clin Invest. 103:1489-98.
Kharitonenkov A, Shiyanova TL, Koester A, Ford AM, Micanovic R, Galbreath EJ, Sandusky GE,
Hammond LJ, Moyers JS, Owens RA, Gromada J, Brozinick JT, Hawkins ED, Wroblewski VJ,
Li DS, Mehrbod F, Jaskunas SR, Shanafelt AB. 2005. FGF-21 as a novel metabolic regulator. J
Clin Invest. 115:1627-35.
Kharitonenkov A, Wroblewski VJ, Koester A, Chen YF, Clutinger CK, Tigno XT, Hansen BC,
Shanafelt AB, Etgen GJ. 2007. The metabolic state of diabetic monkeys is regulated by
fibroblast growth factor-21. Endocrinology. 148:774-81.
Kharitonenkov A, Dunbar JD, Bina HA, Bright S, Moyers JS, Zhang C, Ding L, Micanovic R, Mehrbod
SF, Knierman MD, Hale JE, Coskun T, Shanafelt AB. 2008. FGF-21/FGF-21 receptor
interaction and activation is determined by betaKlotho. J Cell Physiol. 215:1-7.
Kharitonenkov A. 2009. FGFs and metabolism. Curr Opin Pharmacol. 9:805-10.
Khor TO, Huang MT, Prawan A, Liu Y, Hao X, Yu S, Cheung WK, Chan JY, Reddy BS, Yang CS, Kong
AN. 2008. Increased susceptibility of Nrf2 knockout mice to colitis-associated colorectal
cancer. Cancer Prev Res (Phila). 1:187-91.
Khor TO, Huang MT, Kwon KH, Chan JY, Reddy BS, Kong AN. 2006. Nrf2-deficient mice have an
increased susceptibility to dextran sulfate sodium-induced colitis. Cancer Res. 66:11580-4.
Kim J, Cha YN, Surh YJ. 2010. A protective role of nuclear factor-erythroid 2-related factor-2 (Nrf2) in
inflammatory disorders. Mutat Res. 690:12-23.
Koch A, König B, Luci S, Stangl GI, Eder K. 2007. Dietary oxidised fat up regulates the expression of
organic cation transporters in liver and small intestine and alters carnitine concentrations in
liver, muscle and plasma of rats. Br J Nutr. 98:882-9.
König B, Eder K. 2006. Differential action of 13-HPODE on PPARalpha downstream genes in rat Fao
and human HepG2 hepatoma cell lines. J Nutr Biochem. 17:410-8.
Kratschmar DV, Calabrese D, Walsh J, Lister A, Birk J, Appenzeller-Herzog C, Moulin P, Goldring
CE, Odermatt A. 2012. Suppression of the Nrf2-dependent antioxidant response by
glucocorticoids and 11β-HSD1-mediated glucocorticoid activation in hepatic cells. PLoS One.
7:e36774.

Literaturverzeichnis
63
Kwak MK, Kensler TW. 2010. Targeting NRF2 signaling for cancer chemoprevention. Toxicol Appl
Pharmacol. 244:66-76.
Lavrovsky Y, Schwartzman ML, Levere RD, Kappas A, Abraham NG. 1994. Identification of binding
sites for transcription factors NF-kappa B and AP-2 in the promoter region of the human
heme oxygenase 1 gene. Proc Natl Acad Sci U S A. 91:5987-91.
Lee JM, Johnson JA. 2004. An important role of Nrf2-ARE pathway in the cellular defense mechanism.
J Biochem Mol Biol. 37:139-43.
Lee JM, Li J, Johnson DA, Stein TD, Kraft AD, Calkins MJ, Jakel RJ, Johnson JA. 2005. Nrf2, a multi-
organ protector? FASEB J. 19:1061-6.
Leonardini A, Laviola L, Perrini S, Natalicchio A, Giorgino F. 2009. Cross-Talk between PPARgamma
and Insulin Signaling and Modulation of Insulin Sensitivity. PPAR Res. 2009;2009:818945.
Leone TC, Weinheimer CJ, Kelly DP. A critical role for the peroxisome proliferator-activated receptor
alpha (PPARalpha) in the cellular fasting response: the PPARalpha-null mouse as a model of
fatty acid oxidation disorders. Proc Natl Acad Sci U S A. 1999 Jun 22;96(13):7473-8.
Levy S, Forman HJ. 2010. C-Myc is a Nrf2-interacting protein that negatively regulates phase II genes
through their electrophile responsive elements. IUBMB Life. 62:237-46.
Lewis JD, Lichtenstein GR, Stein RB, Deren JJ, Judge TA, Fogt F, Furth EE, Demissie EJ, Hurd LB, Su
CG, Keilbaugh SA, Lazar MA, Wu GD. 2001. An open-label trial of the PPAR-gamma ligand
rosiglitazone for active ulcerative colitis. Am J Gastroenterol. 96:3323-8.
Li H, Fang Q, Gao F, Fan J, Zhou J, Wang X, Zhang H, Pan X, Bao Y, Xiang K, Xu A, Jia W. 2010.
Fibroblast growth factor 21 levels are increased in nonalcoholic fatty liver disease patients and
are correlated with hepatic triglyceride. J Hepatol. 53:934-40.
Li JJ, Cao Y, Young MR, Colburn NH. 2000. Induced expression of dominant-negative c-jun
downregulates NFkappaB and AP-1 target genes and suppresses tumor phenotype in human
keratinocytes. Mol Carcinog. 29:159-69.
Li W, Khor TO, Xu C, Shen G, Jeong WS, Yu S, Kong AN. 2008. Activation of Nrf2-antioxidant
signaling attenuates NFkappaB-inflammatory response and elicits apoptosis. Biochem
Pharmacol. 76:1485-9.
Li X, Stark GR. 2002. NFkappaB-dependent signaling pathways. Exp Hematol. 30:285-96.
Liu GH, Qu J, Shen X. 2008. NF-kappaB/p65 antagonizes Nrf2-ARE pathway by depriving CBP from
Nrf2 and facilitating recruitment of HDAC3 to MafK. Biochim Biophys Acta. 1783:713-27.
Liu JF, Huang CJ. 1996. Dietary oxidized frying oil enhances tissue alpha-tocopherol depletion and
radioisotope tracer excretion in vitamin E-deficient rats. J Nutr. 126:2227-35.
Liu JF, Huang CJ. 1995. Tissue alpha-tocopherol retention in male rats is compromised by feeding
diets containing oxidized frying oil. J Nutr. 125:3071-80.
Lu Y, Gong P, Cederbaum AI. 2008. Pyrazole induced oxidative liver injury independent of
CYP2E1/2A5 induction due to Nrf2 deficiency. Toxicology. 252:9-16.

Literaturverzeichnis
64
Luci S, König B, Giemsa B, Huber S, Hause G, Kluge H, Stangl GI, Eder K. 2007. Feeding of a deep-
fried fat causes PPARalpha activation in the liver of pigs as a non-proliferating species. Br J
Nutr. 97:872-82.
Luedde T, Beraza N, Kotsikoris V, van Loo G, Nenci A, De Vos R, Roskams T, Trautwein C,
Pasparakis M. 2007. Deletion of NEMO/IKKgamma in liver parenchymal cells causes
steatohepatitis and hepatocellular carcinoma. Cancer Cell. 11:119-32.
Lundåsen T, Hunt MC, Nilsson LM, Sanyal S, Angelin B, Alexson SE, Rudling M. 2007. PPARalpha is
a key regulator of hepatic FGF21. Biochem Biophys Res Commun. 360:437-40.
Lunney JK. 2007. Advances in swine biomedical model genomics. Int J Biol Sci. 3(3):179-84.
Makarov SS. 2001. NF-kappa B in rheumatoid arthritis: a pivotal regulator of inflammation,
hyperplasia, and tissue destruction. Arthritis Res. 3:200-6.
Mandard S, Müller M, Kersten S. 2004. Peroxisome proliferator-activated receptor alpha target genes.
Cell Mol Life Sci. 61:393-416.
Martin JC, Joffre F, Siess MH, Vernevaut MF, Collenot P, Genty M, Sébédio JL. 2000. Cyclic fatty acid
monomers from heated oil modify the activities of lipid synthesizing and oxidizing enzymes
in rat liver. J Nutr. 130:1524-30.
McMahon M, Itoh K, Yamamoto M, Chanas SA, Henderson CJ, McLellan LI, Wolf CR, Cavin C,
Hayes JD. 2001. The Cap'n'Collar basic leucine zipper transcription factor Nrf2 (NF-E2 p45-
related factor 2) controls both constitutive and inducible expression of intestinal
detoxification and glutathione biosynthetic enzymes. Cancer Res. 61:3299-307.
McMahon M, Itoh K, Yamamoto M, Hayes JD. 2003. Keap1-dependent proteasomal degradation of
transcription factor Nrf2 contributes to the negative regulation of antioxidant response
element-driven gene expression. J Biol Chem. 278:21592-600.
Michiels C, Minet E, Mottet D, Raes M. 2002. Regulation of gene expression by oxygen: NF-kappaB
and HIF-1, two extremes. Free Radic Biol Med. 33:1231-42.
Michiels C, Raes M, Toussaint O, Remacle J. 1994. Importance of Se-glutathione peroxidase, catalase,
and Cu/Zn-SOD for cell survival against oxidative stress. Free Radic Biol Med. 17:235-48.
Moi P, Chan K, Asunis I, Cao A, Kan YW. 1994. Isolation of NF-E2-related factor 2 (Nrf2), a NF-E2-
like basic leucine zipper transcriptional activator that binds to the tandem NF-E2/AP1 repeat
of the beta-globin locus control region. Proc Natl Acad Sci U S A. 91:9926-30.
Moyers JS, Shiyanova TL, Mehrbod F, Dunbar JD, Noblitt TW, Otto KA, Reifel-Miller A,
Kharitonenkov A. 2007. Molecular determinants of FGF-21 activity-synergy and cross-talk
with PPARgamma signaling. J Cell Physiol. 210:1-6.
Muise ES, Azzolina B, Kuo DW, El-Sherbeini M, Tan Y, Yuan X, Mu J, Thompson JR, Berger JP, Wong
KK. 2008. Adipose fibroblast growth factor 21 is up-regulated by peroxisome proliferator-
activated receptor gamma and altered metabolic states. Mol Pharmacol. 74:403-12.
Nagy L, Tontonoz P, Alvarez JG, Chen H, Evans RM. 1998. Oxidized LDL regulates macrophage gene
expression through ligand activation of PPARgamma. Cell. 93:229-40.

Literaturverzeichnis
65
Natarajan R, Reddy MA, Malik KU, Fatima S, Khan BV. 2001. Signaling mechanisms of nuclear factor-
kappab-mediated activation of inflammatory genes by 13-hydroperoxyoctadecadienoic acid in
cultured vascular smooth muscle cells. Arterioscler Thromb Vasc Biol. 21:1408-13.
Nenci A, Becker C, Wullaert A, Gareus R, van Loo G, Danese S, Huth M, Nikolaev A, Neufert C,
Madison B, Gumucio D, Neurath MF, Pasparakis M. 2007. Epithelial NEMO links innate
immunity to chronic intestinal inflammation. Nature. 446:557-61.
Nishimura T, Nakatake Y, Konishi M, Itoh N. 2000. Identification of a novel FGF, FGF-21,
preferentially expressed in the liver. Biochim Biophys Acta. 1492:203-6.
Ogawa Y, Kurosu H, Yamamoto M, Nandi A, Rosenblatt KP, Goetz R, Eliseenkova AV, Mohammadi
M, Kuro-o M. 2007. BetaKlotho is required for metabolic activity of fibroblast growth factor
21. Proc Natl Acad Sci U S A. 104:7432-7.
Okawa H, Motohashi H, Kobayashi A, Aburatani H, Kensler TW, Yamamoto M. 2006. Hepatocyte-
specific deletion of the keap1 gene activates Nrf2 and confers potent resistance against acute
drug toxicity. Biochem Biophys Res Commun. 339:79-88.
Okayasu T, Tomizawa A, Suzuki K, Manaka K, Hattori Y. 2008. PPARalpha activators upregulate
eNOS activity and inhibit cytokine-induced NF-kappaB activation through AMP-activated
protein kinase activation. Life Sci. 82:884-91.
Olivero David R, Bastida S, Schultz A, González Torres L, González-Muñoz MJ, Sánchez-Muniz FJ,
Benedí J. 2010. Fasting status and thermally oxidized sunflower oil ingestion affect the
intestinal antioxidant enzyme activity and gene expression of male Wistar rats. J Agric Food
Chem. 58:2498-504.
Osburn WO, Karim B, Dolan PM, Liu G, Yamamoto M, Huso DL, Kensler TW. 2007. Increased
colonic inflammatory injury and formation of aberrant crypt foci in Nrf2-deficient mice upon
dextran sulfate treatment. Int J Cancer. 121:1883-91.
Pasparakis M. 2008. IKK/NF-kappaB signaling in intestinal epithelial cells controls immune
homeostasis in the gut. Mucosal Immunol. 1:S54-7.
Penumetcha M, Khan N, Parthasarathy S. 2000. Dietary oxidized fatty acids: an atherogenic risk? J
Lipid Res. 41:1473-80.
Petrick JS, Klaassen CD. 2007. Importance of hepatic induction of constitutive androstane receptor
and other transcription factors that regulate xenobiotic metabolism and transport. Drug
Metab Dispos. 35:1806-15.
Picard F, Auwerx J. 2002. PPAR(gamma) and glucose homeostasis. Annu Rev Nutr. 22:167-97.
Potthoff MJ, Inagaki T, Satapati S, Ding X, He T, Goetz R, Mohammadi M, Finck BN, Mangelsdorf DJ,
Kliewer SA, Burgess SC. 2009. FGF21 induces PGC-1alpha and regulates carbohydrate and
fatty acid metabolism during the adaptive starvation response. Proc Natl Acad Sci U S A.
106:10853-8.
Ramos-Gomez M, Kwak MK, Dolan PM, Itoh K, Yamamoto M, Talalay P, Kensler TW. 2001.
Sensitivity to carcinogenesis is increased and chemoprotective efficacy of enzyme inducers is
lost in nrf2 transcription factor-deficient mice. Proc Natl Acad Sci U S A. 98:3410-5.

Literaturverzeichnis
66
Ringseis R, Eder K. Regulation of genes involved in lipid metabolism by dietary oxidized fat. 2011. Mol
Nutr Food Res. 55:109-21.
Ringseis R, Muschick A, Eder K. 2007a. Dietary oxidized fat prevents ethanol-induced triacylglycerol
accumulation and increases expression of PPARalpha target genes in rat liver. J Nutr. 137:77-
83.
Ringseis R, Piwek N, Eder K. 2007b. Oxidized fat induces oxidative stress but has no effect on NF-
kappaB-mediated proinflammatory gene transcription in porcine intestinal epithelial cells.
Inflamm Res. 56:118-25.
Roberts RA, James NH, Hasmall SC, Holden PR, Lambe K, Macdonald N, West D, Woodyatt NJ,
Whitcome D. 2000. Apoptosis and proliferation in nongenotoxic carcinogenesis: species
differences and role of PPARalpha. Toxicol Lett. 112-113:49-57.
Rojo AI, Salinas M, Martín D, Perona R, Cuadrado A. 2004. Regulation of Cu/Zn-superoxide
dismutase expression via the phosphatidylinositol 3 kinase/Akt pathway and nuclear factor-
kappaB. J Neurosci. 24:7324-34.
Rosen ED, Spiegelman BM. PPARgamma : a nuclear regulator of metabolism, differentiation, and cell
growth. 2001. J Biol Chem. 276:37731-4.
Rosen ED, Sarraf P, Troy AE, Bradwin G, Moore K, Milstone DS, Spiegelman BM, Mortensen RM.
1999. PPAR gamma is required for the differentiation of adipose tissue in vivo and in vitro.
Mol Cell. 4:611-7.
Rushmore TH, Kong AN. 2002. Pharmacogenomics, regulation and signaling pathways of phase I and
II drug metabolizing enzymes. Curr Drug Metab. 3:481-90.
Rushmore TH, Morton MR, Pickett CB. 1991. The antioxidant responsive element. Activation by
oxidative stress and identification of the DNA consensus sequence required for functional
activity. J Biol Chem. 266:11632-9.
Schild RL, Schaiff WT, Carlson MG, Cronbach EJ, Nelson DM, Sadovsky Y. 2002. The activity of
PPAR gamma in primary human trophoblasts is enhanced by oxidized lipids. J Clin
Endocrinol Metab. 87:1105-10.
Schreiber S, Nikolaus S, Hampe J. 1998. Activation of nuclear factor kappa B inflammatory bowel
disease. Gut. 42:477-84.
Skufca P, Brandsch C, Hirche F, Eder K. 2003. Effects of a dietary thermally oxidized fat on thyroid
morphology and mRNA concentrations of thyroidal iodide transporter and thyroid
peroxidase in rats. Ann Nutr Metab. 47:207-13.
Song BJ, Soh Y, Bae M, Pie J, Wan J, Jeong K. 2001. Apoptosis of PC12 cells by 4-hydroxy-2-nonenal is
mediated through selective activation of the c-Jun N-terminal protein kinase pathway. Chem
Biol Interact. 130-132:943-54.
Spehlmann ME, Eckmann L. 2009. Nuclear factor-kappa B in intestinal protection and destruction.
Curr Opin Gastroenterol. 25:92-9.
Srinivasan KN, Pugalendi KV. 2000. Effect of excessive intake of thermally oxidized sesame oil on
lipids, lipid peroxidation and antioxidants' status in rats. Indian J Exp Biol. 38:777-80.

Literaturverzeichnis
67
Staprans I, Pan XM, Rapp JH, Feingold KR. 2005. The role of dietary oxidized cholesterol and
oxidized fatty acids in the development of atherosclerosis. Mol Nutr Food Res. 49:1075-82.
Staprans I, Pan XM, Rapp JH, Feingold KR. 2003. Oxidized cholesterol in the diet is a source of
oxidized lipoproteins in human serum. J Lipid Res. 44:705-15.
Staprans I, Rapp JH, Pan XM, Kim KY, Feingold KR. 1994. Oxidized lipids in the diet are a source of
oxidized lipid in chylomicrons of human serum. Arterioscler Thromb. 14:1900-5.
Staprans I, Rapp JH, Pan XM, Feingold KR. 1993. The effect of oxidized lipids in the diet on serum
lipoprotein peroxides in control and diabetic rats. J Clin Invest. 92:638-43.
Sülzle A, Hirche F, Eder K. 2004. Thermally oxidized dietary fat upregulates the expression of target
genes of PPAR alpha in rat liver. J Nutr. 134:1375-83.
Sun Y, Oberley LW. 1996. Redox regulation of transcriptional activators. Free Radic Biol Med. 21:335-
48.
Suomela JP, Ahotupa M, Kallio H. 2005. Triacylglycerol oxidation in pig lipoproteins after a diet rich
in oxidized sunflower seed oil. Lipids. 40:437-44.
Suomela JP, Ahotupa M, Sjövall O, Kurvinen JP, Kallio H. 2004. Diet and lipoprotein oxidation:
analysis of oxidized triacylglycerols in pig lipoproteins. Lipids. 39:639-47.
Surh YJ, Kundu JK, Na HK, Lee JS. 2005. Redox-sensitive transcription factors as prime targets for
chemoprevention with anti-inflammatory and antioxidative phytochemicals. J Nutr. 135:2993-
3001.
Swindle MM, Makin A, Herron AJ, Clubb FJ Jr, Frazier KS. 2012. Swine as models in biomedical
research and toxicology testing. Vet Pathol. 49:344-56.
Tontonoz P, Hu E, Spiegelman BM. 1994. Stimulation of adipogenesis in fibroblasts by PPAR gamma
2, a lipid-activated transcription factor. Cell. 79:1147-56.
Tsuchida A, Yamauchi T, Takekawa S, Hada Y, Ito Y, Maki T, Kadowaki T. 2005. Peroxisome
proliferator-activated receptor (PPAR)alpha activation increases adiponectin receptors and
reduces obesity-related inflammation in adipose tissue: comparison of activation of
PPARalpha, PPARgamma, and their combination. Diabetes. 54:3358-70.
Tugwood JD, Holden PR, James NH, Prince RA, Roberts RA. 1998. A peroxisome proliferator-
activated receptor-alpha (PPARalpha) cDNA cloned from guinea-pig liver encodes a protein
with similar properties to the mouse PPARalpha: implications for species differences in
responses to peroxisome proliferators. Arch Toxicol. 72:169-77.
Turrens JF. 2003. Mitochondrial formation of reactive oxygen species. J Physiol. 552:335-44.
Vansell NR, Klaassen CD. 2002. Increase in rat liver UDP-glucuronosyltransferase mRNA by
microsomal enzyme inducers that enhance thyroid hormone glucuronidation. Drug Metab
Dispos. 30:240-6.
Venugopal R, Jaiswal AK. 1996. Nrf1 and Nrf2 positively and c-Fos and Fra1 negatively regulate the
human antioxidant response element-mediated expression of NAD(P)H:quinone
oxidoreductase1 gene. Proc Natl Acad Sci U S A. 93:14960-5.

Literaturverzeichnis
68
Vo N, Goodman RH. 2001. CREB-binding protein and p300 in transcriptional regulation. J Biol Chem.
276:13505-8.
Wakabayashi N, Dinkova-Kostova AT, Holtzclaw WD, Kang MI, Kobayashi A, Yamamoto M, Kensler
TW, Talalay P. 2004. Protection against electrophile and oxidant stress by induction of the
phase 2 response: fate of cysteines of the Keap1 sensor modified by inducers. Proc Natl Acad
Sci U S A. 101:2040-5.
Wang N, Verna L, Chen NG, Chen J, Li H, Forman BM, Stemerman MB. 2002. Constitutive activation
of peroxisome proliferator-activated receptor-gamma suppresses pro-inflammatory adhesion
molecules in human vascular endothelial cells. J Biol Chem. 277:34176-81.
Waterston RH, Lindblad-Toh K, Birney E, Rogers J, Abril JF, et al. 2002. Initial sequencing and
comparative analysis of the mouse genome. Nature. 420(6915):520-62.
Wente W, Efanov AM, Brenner M, Kharitonenkov A, Köster A, Sandusky GE, Sewing S, Treinies I,
Zitzer H, Gromada J. 2006. Fibroblast growth factor-21 improves pancreatic beta-cell
function and survival by activation of extracellular signal-regulated kinase 1/2 and Akt
signaling pathways. Diabetes. 55:2470-8.
Winegar DA, Brown PJ, Wilkison WO, Lewis MC, Ott RJ, Tong WQ, Brown HR, Lehmann JM,
Kliewer SA, Plunket KD, Way JM, Bodkin NL, Hansen BC. 2001. Effects of fenofibrate on lipid
parameters in obese rhesus monkeys. J Lipid Res. 42:1543-51.
Xu C, Shen G, Chen C, Gélinas C, Kong AN. 2005. Suppression of NF-kappaB and NF-kappaB-
regulated gene expression by sulforaphane and PEITC through IkappaBalpha, IKK pathway
in human prostate cancer PC-3 cells. Oncogene. 24:4486-95.
Xu J, Lloyd DJ, Hale C, Stanislaus S, Chen M, Sivits G, Vonderfecht S, Hecht R, Li YS, Lindberg RA,
Chen JL, Jung DY, Zhang Z, Ko HJ, Kim JK, Véniant MM. 2009a. Fibroblast growth factor 21
reverses hepatic steatosis, increases energy expenditure, and improves insulin sensitivity in
diet-induced obese mice. Diabetes. 58:250-9.
Xu J, Stanislaus S, Chinookoswong N, Lau YY, Hager T, Patel J, Ge H, Weiszmann J, Lu SC, Graham
M, Busby J, Hecht R, Li YS, Li Y, Lindberg R, Véniant MM. 2009b. Acute glucose-lowering
and insulin-sensitizing action of FGF21 in insulin-resistant mouse models--association with
liver and adipose tissue effects. Am J Physiol Endocrinol Metab. 297:1105-14.
Xu W, Hellerbrand C, Köhler UA, Bugnon P, Kan YW, Werner S, Beyer TA. 2008. The Nrf2
transcription factor protects from toxin-induced liver injury and fibrosis. Lab Invest. 88:1068-
78.
Yamanaka H, Nakajima M, Katoh M, Yokoi T. 2007. Glucuronidation of thyroxine in human liver,
jejunum, and kidney microsomes. Drug Metab Dispos. 35:1642-8.
Yang YC, Lii CK, Wei YL, Li CC, Lu CY, Liu KL, Chen HW. 2012. Docosahexaenoic acid inhibition of
inflammation is partially via cross-talk between Nrf2/heme oxygenase 1 and IKK/NF-κB
pathways. J Nutr Biochem. [Epub ahead of print]
Yoder Graber AL, Ramírez J, Innocenti F, Ratain MJ. 2007. UGT1A1*28 genotype affects the in-vitro
glucuronidation of thyroxine in human livers. Pharmacogenet Genomics. 17:619-27.

Literaturverzeichnis
69
Yueh MF, Tukey RH. 2007. Nrf2-Keap1 signaling pathway regulates human UGT1A1 expression in
vitro and in transgenic UGT1 mice. J Biol Chem. 282:8749-58.
Zhu M, Fahl WE. 2001. Functional characterization of transcription regulators that interact with the
electrophile response element. Biochem Biophys Res Commun. 289:212-9.

Erklärung
A
Erklärung „Ich erkläre: Ich habe die vorgelegte Dissertation „Einfluss oxidierter Fette auf zelluläre Signalwege
im Modelltier“ selbständig und ohne unerlaubte fremde Hilfe und nur mit den Hilfen angefertigt, die
ich in der Dissertation angegeben habe. Alle Textstellen, die wörtlich oder sinngemäß aus
veröffentlichten Schriften entnommen sind, und alle Angaben, die auf mündlichen Auskünften
beruhen, sind als solche kenntlich gemacht. Bei den von mir durchgeführten und in der Dissertation
erwähnten Untersuchungen habe ich die Grundsätze guter wissenschaftlicher Praxis, wie sie in der
„Satzung der Justus-Liebig-Universität Gießen zur Sicherung guter wissenschaftlicher Praxis“
niedergelegt sind, eingehalten.“
Halle (Saale), 26.04.2013 _________________________________________
Juliane Varady

Danksagung
B
Danksagung
An dieser Stelle bedanke ich mich bei allen, die mich bei der Durchführung und Vollendung meiner
Promotionsarbeit unterstützt haben.
Ich danke:
an erster Stelle Herrn Prof. Dr. K. Eder für die Aufnahme in seine Forschungsgruppe, die
Überlassung des Promotionsthemas und die wissenschaftliche Betreuung der Arbeit;
Herrn Dr. habil. R. Ringseis für seine stete Bereitschaft zur Diskussion und Hilfestellung, sowie die
vielen Hinweise und Ratschläge bei der Bearbeitung des Promotionsthemas;
Herrn Prof. Dr. U. Wenzel für die Übernahme des Zweitgutachtens;
der Deutschen Forschungsgemeinschaft für die Projektförderung;
allen Kollegen/-innen und Mitarbeiter/-innen am Institut für Tierernährung und
Ernährungsphysiologie der Justus-Liebig-Universität Gießen sowie des Institutes für Agrar- und
Ernährungswissenschaften der Martin Luther-Universität Halle-Wittenberg für ihre Unterstützung
und Hilfe;
meiner Familie, meinen Freunden und meinem Freund für ihre Unterstützung, ihr Verständnis und
ihr Interesse an meiner Arbeit.





























