Embed Size (px)
Citation preview

Research ArticleElectroacupuncture Regulates HippocampalSynaptic Plasticity via miR-134-Mediated LIMK1Function in Rats with Ischemic Stroke
Weilin Liu1 Jie Wu2 Jia Huang1 Peiyuan Zhuo2 Yunjiao Lin2 Lulu Wang2
Ruhui Lin2 Lidian Chen1 and Jing Tao1
1College of Rehabilitation Medicine Fujian University of Traditional Chinese Medicine Fuzhou Fujian 350122 China2Fujian Key Laboratory of Rehabilitation Technology Fuzhou Fujian 350122 China
Correspondence should be addressed to Lidian Chen cldfjtcmeducn and Jing Tao taojing01163com
Received 26 August 2016 Accepted 3 November 2016 Published 2 January 2017
Academic Editor Cun-Zhi Liu
Copyright copy 2017 Weilin Liu et al This is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
MircoRNAs (miRs) have been implicated in learning and memory by regulating LIM domain kinase (LIMK1) to induce synaptic-dendritic plasticity The study aimed to investigate whether miRNAsLIMK1 signaling was involved in electroacupuncture- (EA-) mediated synaptic-dendritic plasticity in a rat model of middle cerebral artery occlusion induced cognitive deficit (MICD)Compared to untreatment or non-acupoint-EA treatment EA at DU20 and DU24 acupoints could shorten escape latency andincrease the frequency of crossing platform in Morris water maze test T2-weighted imaging showed that the MICD rat brainlesions were located in cortex hippocampus corpus striatum and thalamus regions and injured volumes were reduced after EAFurthermore we found that the density of dendritic spine and the number of synapses in the hippocampal CA1 pyramidal cellswere obviously reduced at Day 14 after MICD However synaptic-dendritic loss could be rescued after EA Moreover the synaptic-dendritic plasticitywas associatedwith increases of the total LIMK1 andphospho-LIMK1 levels in hippocampalCA1 regionwhereinEA decreased the expression of miR-134 negatively regulating LIMK1 to enhance synaptic-dendritic plasticityTherefore miR-134-mediated LIMK1 was involved in EA-induced hippocampal synaptic plasticity which served as a contributor to improving learningand memory during the recovery stage of ischemic stroke
1 Introduction
Ischemic stroke results in a high mortality rate and increaseddisability rate all over the world [1] Approximately 64 ofstroke patients are often followed with cognitive impairmentand 33 of them turn into dementia existing for severalmonths during decubation [2] Cognitive deficits arise frequ-ently after ischemic stroke which cause difficulties with anal-ysis concentration organization interpretation and otherabates in cognitive functions that bring about the low qualityof life [3 4] The dysfunction of learning and memory is thecardinal symptom of cognitive impairment after stroke and isthe main culprit of persistent sequelae [5] A recent studydemonstrated that the incidence rate of poststroke mildcognitive impairment was diagnosed in 244 of individuals
after 3 years and each year the mean growth rate is approxi-mately 8 [6] In addition to conventional cognitive trainingelectroacupuncture (EA) is a stretch therapeutic method ofacupuncture which is traditional acupuncture incorporationwith modern electrotherapy The clinical efficacy of EA onpoststroke cognitive impairment has been widely demon-strated [7 8] However the functional mechanism of EA isfar from been fully elucidated
The hippocampus is a pivotal structure of the brain thearea plays an important role in the formation of acquisitionconsolidation and recognition of declarative and spatialmemory [9 10] The loss of hippocampal synapses and neu-rons in poststroke induces cognitive deficits including spatialreference learning and memory impairment [11 12] In theformation of spatial referencememory is closely related to the
HindawiNeural PlasticityVolume 2017 Article ID 9545646 11 pageshttpsdoiorg10115520179545646
2 Neural Plasticity
plasticity of dendritic spines and the morphological changessuch as expansion and contraction [13] Dendritic spines altertheir shape to make the information spreading more easilyand influence the synaptic efficacy (ie long-term potenti-ation and long-term depression) [14 15] which have beenwidely considered as a cellular mechanism for learning andmemory [16] LIM domain kinase (LIMK1) is enriched inboth axonal and dendritic growth cones of hippocampalpyramidal neurons in rats [17] LIMK1 encodes a serinethre-onine protein kinase that regulates the actin cytoskeleton byphosphorylating and inactivating the actin depolymerizationfactor (ADF)cofilin [18] In addition LIMK1 is also knownas having an important role in synapse and dendritic spinefunction It has been reported that the knockout mice lackingLIMK1 are severely impaired in dendritic spine morphologyand hippocampal long-term potentiation [19 20] Evidenceshowed that LIMK1 regulated long-term memory (LTM)and long lasting synaptic plasticity through interacting withand activating cyclic AMP response element-binding protein(CREB) [21]
In addition a potential role for microRNAs (miRNAs ormiRs) in synaptic function has been particularly intriguinggiven the evidence that a brain-specific miRNA contributesto synaptic development maturation andor plasticity [22]miRNAs are endogenous noncoding RNAs that mediate theposttranscriptional regulation of gene expression mainly bybinding to the 31015840-untranslated region of messenger RNAs(mRNAs) [23] A number of miRNAs have been isolatedfrom nervous system and a recent study has demonstrateda crucial role for dynamically regulating synaptic plasticity[24 25] Moreover miRNAs have been implicated in hippo-campus-dependent function which have a significant poten-tial in learning and memory formation regulating LIMK1expression to induce synaptic-dendritic plasticity [22] Den-dritic mRNAs encode diversified functionalities in hippo-campal pyramidal neurons and play an important role insynaptic plasticity as well as learning and memory [26]
Therefore miRNA-LIMK1 can be considered as a targetfor cognitive deficit Our previous study has shown that EAat Baihui (DU20) and Shenting (DU24) acupoints couldimprove cognitive impairment through Rho GTPases toenhance dendritic plasticity in rats with ischemic stroke [27]Interestingly it has been suggested that the activation of theRho GTPases signaling is essential for LIMK1 activation byphosphorylation on threonine 508 which is widely known asmaster regulator of actin dynamics [28 29] Thus the studyaimed to elucidate whether EA at the DU20 and DU24acupoints could improve the cognitive deficits in rats withischemic stroke via miRNA-LIMK1-mediated synaptic plas-ticity to enhance spatial reference learning and memory
2 Materials and Methods
21 Animal Ethics Statement 96 male Sprague-Dawley rats(weight 250 plusmn 20 g) were obtained from Shanghai SLAC Lab-oratoryAnimalCo Ltd (Shanghai China SCXK2013-0005)and housed under pathogen-free conditions with a 12 hlightdark cycleThe study was approved by the Committee ofFujian University of Traditional Chinese Medicine (protocol
FUTCM-2015008) and was performed in accordance withthe national guidelines for the care and use of laboratoryanimals For euthanasia 3 sodium pentobarbital (40mgkgbody weight ip) was used Middle cerebral artery occlusion(MCAO) surgery was carried out under general anesthesia(15 isoflurane in 685 N2O and 30 O2) All efforts weremade to minimize suffering
22 Experimental Procedures TheMCAO-induced cognitivedeficit (MICD)modelwas established as previously described[5 30] Rats were randomly assigned to four groups accordingto the random number table (119899 = 24 rats each group) as fol-lows (1) Sham group (2)MICD group (3)MICD+EA groupand (4) MICD+non-EA group The left middle cerebralartery was occluded by a 4-0 nylon monofilament (023mmin diameter Jialing-bio China) Focal cerebral ischemiawas monitored using transcranial temporal laser Doppler(BIOPAC Systems Goleta CA USA) and an 80 decrease inblood flow after the occlusion was recorded After 90min ofocclusion reperfusion was achieved by pulling out the fila-ment to restore blood flow Sham-operated rats of the Shamgroup underwent the same procedure but arterial occlusionwas not performed
After 24 hours of MCAO surgery rats of the Sham groupand the MICD group received no treatment Rats of theMICD+EAgroupwere givenEA treatment for 30min per dayfor 14 consecutive days The EA needles (diameter 03mmneedle purchased from Hualun acupuncture of Suzhou CoLtd Suzhou China) were inserted at a depth of 2-3mm intothe Baihui (DU20 located in the median of frontalis) andShenting (DU24 located in the median of the parietal bone)acupoints [31] Stimulation was then generated using the EAapparatus [ModelG6805 ShanghaiHuayi (Group)CompanyLtd Shanghai China] and the stimulation parameters wereset as follows dilatational waves of 1sim20Hz (adjusted to themuscle twitch threshold) peak voltage of 6V and 02mAintensity [31] Rats of the MICD+non-EA group were givenEA treatment at the bilateral nonacupoints (located in thecostal region and 10mmdistal to the iliac crest) for 30min perday for 14 consecutive days [32]The EA needles and stimula-tion parameters of the bilateral nonacupoints were consistentwith the EA treatment Manipulators were experienced andblinded to the ratrsquos group
23 Behavioral Assessment Behavioral testingwas conductedby researchers who were blinded to the ratrsquos group At 10 daysafter EA all the rats were subjected to the Morris water mazetest (Shanghai Xinruan Information Technology Co LtdShanghai China) to evaluate spatial reference learning andmemory (Figure 1)TheMorris water maze consisted of a cir-cular pool (diameter 150 cm height 60 cm) filled with water(depth of 30 cm and temperature of 25 plusmn 2∘C) A circularescape platform (diameter 12 cm height 29 cm) was sub-merged 2 cm below the water surface in the middle of thethird quadrant of the pool and the reference objects aroundthe pool were placedMorris watermaze tasksmainly includeorientation navigation and space exploration trials Duringthe first set of trials each rat was placed in the water at eachof the four equidistant locations to the platform When the
Neural Plasticity 3
0
Sham or MCAO surgery Sacrificed
Morris water test
Electroacupuncture
Baseline T2WI T2WI
24 h 14 d10 d7 d1 d
Figure 1 Experimental design used in the study
rats arrived at the platform within the 90 sec time restrictionand remained on it for 3 sec they were considered to havefound the platform and were scored by the time takenlengthof the route When the rats were unable to find the platformwithin 90 sec they were placed on the platform for 10 sec andthe time scorewas 90 secThe time taken and the length of theroute bywhich each rat found the safe platformwere recordedby the computerThe average of the time taken and the lengthof the route for the four quadrants as a result of each rat wereassessed every day The duration of the first set of trials wasperformed on Days 10 to 13 after EA The second part of theexperiment was performed on Day 14 to examine the timein which rats found the location of the platform within the90 sec time restriction which tested their ability to rememberthe position of the platform After all trials the rats weredried thoroughly with a hair drier and returned to their cages(Guangzhou RiboBio Co Ltd Guangzhou China) Morriswater maze test was repeated three times
24 Measurement of Brain Lesions Animal MRI scans wereperformed on a 70 T MRI scanner (7020USR BioSpecBruker Biospin Gmbls Germany) using a 38mmbirdcage ratbrain quadrature resonator for radiofrequency transmissionand reception Animals were anesthetized with isofluraneO2(with 3 induction for 5min and 12ndash15 maintenance inorder to let the rats in the depth of anesthesia state) andkept warm with circuit After anesthesia the rat was putin prone position on a custom-made holder to minimizehead motion set the location of head position and performreal-time monitor of the breathing rate and maintained in40 breathsmin Ratrsquos temperature is maintained at 33 plusmn 2∘Cin the process of scanning holder to minimize head motionwhile respiration was maintained
T2-weighted imaging (T2WI) in three planes with afast spin echo (FSE) pulse sequence was first acquired tocontrol rat head positioning T2WI scan was acquired usinga Rapid Acquisition with Relaxation Enhancement (RARE)pulse sequence with the following parameters field of view =32mmtimes 32mmmatrix size = 256times 256 repetition time (TR)= 4200ms echo time (TE) = 35ms slice thickness = 10mmand slice gap = 0mm
Image J analysis and processing system was applied forT2W images the percentage of the brain lesions = brainlesions volumewhole brain volume times100
25 Golgi Staining Golgi staining of brain was performedusing an FD Rapid GolgiStain Kit following the manufac-turerrsquos instructions (FD Neurotechnologies Inc ColumbiaMD USA) The removed rat brains were placed incubated
in a mixture of A and B solutions from the kit and storedin the dark at room temperature following which they weretransferred into solution C from the kit stored at 4∘C for 7days Finally brains were then frozen and coronal sections(150 120583m) were made using a cryostat (Leica CM3050S LeicaMicrosystems K K Tokyo Japan) The sections incubatedin a mixture of D and E solutions from the kit and thendehydrated in alcohol (50 70 90 and 100 for 5min each)cleared in xylene and were cover-slipped The images finallywere viewed under a microscope (Leica DM6000 B LeicaMicrosystems Wetzlar Germany) The slides were reviewedby two or three pathologists blind to the study
26 Transmission Electron Microscopy Four rats in eachgroup were anesthetized and the left ventricle was perfusedwith 200mL of saline followed by 400mL 4 paraformalde-hyde (pH 74) The tissue was taken from the left ischemichippocampus cut into 1mm3 size cubes and fixed in 1paraformaldehyde with 1 lanthanum nitrate tracer for 24 hfollowed by fixation with 3 glutaraldehyde for 24 h Sampleswere fixed in 1 osmium tetroxide for 2 h and dehydrated ingraded ethanol 1 lanthanum nitrate tracer (LNT) solutionand embedded in araldite Ultrathin hippocampal CA1 sliceswere obtained (90 nm) then they were stained with uranylacetate and lead citrate and observed under TEM (H-7650Hitachi Ltd Tokyo Japan) The photos were obtained onhp digital CCD camera (SIS4 million voxel) Images wereacquired digitally from a randomly selected pool of 10 to 15fields under each condition
27 Western Blotting Isolated left hippocampus tissue waslysed in 100 120583L radioimmunoprecipitation assay (RIPA) buf-fer (Beyotime Haimen Jiangsu China) plus protease inhibi-tors Total protein (50120583g) was loaded into 10 SDS-PAGEgels electrophoresed and then transblotted onto polyvinyli-dene difluoride membranes (Immobilon-P Millipore Bil-lerica MA) in a Tris-glycine transfer buffer After beingblocked in 5 milk in PBST for 1 h at room temperaturewith shaking membranes were incubated with antibodiesovernight at 4∘C against the following anti-LIMK1 antibody(dilution 1 1000 ab81046 Abcam) and anti-LIM kinase1 antibody (phospho-Thr508) (dilution 1 1000 ab131341Abcam) and 120573-actin (dilution 1 1000 ab189073 Abcam)The following day membranes were incubated in 5 milk(in TBST) with an anti-goat or anti-mouse IgG antibody(dilution 1 5000 PerkinElmer Life Sciences Waltham MA)for 1 h at room temperature with shaking Membranes werewashed a minimum of four times (10min per wash) in PBST
4 Neural Plasticity
between each antibody treatment Detected bands were visu-alized using enhanced chemiluminescence and images werecaptured using a Bio-Image system (Bio-Rad LaboratoriesInc Hercules USA) Western blotting was repeated threetimes
28 Real Time Quantitative RT-PCR The expression of miR-NAs was determined by real time quantitative reverse tran-scription polymerase chain reaction (RT-PCR) Total RNAwas extracted from hippocampal region of the ipsilaterallesion tissue using the TRIzol Reagent (Life Technologies(AB amp Invitrogen) Carlsbad USA) Then extracted totalRNA was reverse transcribed to generate cDNA accord-ing to manufacturer instructions of Revert AidTM First-Strand cDNA Synthesis Kit (Thermo Fisher Scientific Bei-jing China) The reverse transcription reaction was ampli-fied using a Bio-Rad CFX96 Detection System (Bio-RadHercules CA USA) with the Plexor One-Step qRT-PCR System (Promega Madison WI USA) For miRNAamplification the following primers were used rno-miR-134 (RmiRQP0168 GeneCopoeia Guangzhou China) Thefold change in relative mRNA and miRNA expression wasdetermined using the 2minusΔΔCt method as described previously[33] and U6 snRNA (RQP047936 GeneCopoeia) as mRNAand miRNA internal control respectively
29 Statistical Analysis Statistical analysis was performedwith the SPSS package for Windows statistical analysissoftware (Version 180 SPSS Inc Chicago IL USA) Thedata from all groups were determined by one-way analysisof variance (ANOVA) and Studentrsquos 119905-testsThe homogeneityof variance was analyzed using the least significant differ-ence method and missing variance using the Games-Howellmethod Intergroup comparisons of the brain lesions volumeat different time points were performed with paired-samples119905-tests All data are presented as mean plusmn standard error andthe significance was regarded as at least 119901 lt 005 All finalresults were analyzed in a blinded manner
3 Results
31 EA Improved Cognitive Impairment in MICD Rats Toassess the effect of EA on spatial reference learning andmemory impairments in MICD rats the Morris water maze(MWM) test was performed All groups of rats learned to findthe platform and the latency time to reach the platform wasreduced in the four training days However learning abilitywas significantly reduced in MICD rats compared with theSham group (119901 lt 001 Figure 2(a)) the MICD rats treatedwith EA significantly took less time to find the platform com-pared with the MICD group and the MICD+non-EA group(119901 lt 001 or 119901 lt 005 Figure 2(a)) There was no significantdifference about path length among the four groups (119901 gt005 Figure 2(b)) As illustrated in Figures 2(c) and 2(d)tracing images from the MWM test showed that in the spaceexploration test where the platform was removed the MICDrats passed through the original position of the platformfewer times than the Sham group (119901 lt 001 Figure 2(d))
whereas in the MICD+EA group the times where the ratscrossed the position of the platformwere significantly increa-sed compared with the MICD group and the MICD+non-EA group (119901 lt 001 119901 lt 005 Figure 2(d)) As illustratedin Figures 2(e) and 2(f) the percentage of time spent in thetarget quadrant was used for statistical analysis during theprobe trial The date showed that the rats spent more time inthe target quadrant compared to other quadrants (119901 lt 001Figure 2(e)) The MICD group spent less time in the targetquadrant compared with the other groups (119901 lt 001 Fig-ure 2(e))TheMICD+EA group spent more time in the targetquadrant compared with the MICD group and MICD+non-EA group (119901 lt 001 119901 lt 005 Figure 2(f)) Therefore theseresults suggest that acquisition or retention of spatial refer-ence learning and memory was ameliorated in MICD rats byEA treatment
32 EA Attenuated Left Cortex Hippocampus Corpus Stria-tum and Thalamus Lesions in MICD Rats The brain lesionswere determined by T2-weighted imaging (T2WI) (Figures3(a) and 3(c)) before and after EA treatment The volumes ofbrain lesions that included left cortex hippocampus corpusstriatum and thalamus regions in the all groups had nosignificant difference before EA treatment (119901 gt 005 Figures3(a) and 3(b)) There was mild spontaneous recovery for 14days in the brain lesions of the MICD group The volumesof left cortex hippocampus corpus striatum and thalamuslesions showed comprising approximately 23 of the wholebrain in theMICDgroup whereas the brain lesions sizes weresignificantly reduced to 15 in the MICD+EA group (119901 lt001 Figures 3(c) and 3(d)) The difference of brain lesionsbetween the MICD+EA and the MICD+non-EA groupshowed significant changes at Day 14 after EA (119901 lt 005Figures 3(c) and 3(d))
33 EA Increased the Density of Dendritic Spines in the Hip-pocampus ofMICDRats To investigate the function of EAonsynaptic plasticity dendritic spine density in the hippocampalneurons was analyzed by the primary basilar dendrites ofGolgi-stained pyramidal neurons at Days 14 after EA Golgi-Cox staining clearly filled the dendritic shafts and the spinesof neurons frompyramidal neurons (Figure 4(a))Hereby therepresentative hippocampus showed that the density of den-dritic spines was reduced in different degree by macroscopicexamination in the MICD group the MICD+EA group andMICD+non-EA group and the loss of dendritic spines inhippocampal CA1 was obvious in the MICD group Thus asillustrated in Figures 4(b) and 4(c) the density of selecteddendritic spines that derived from hippocampal CA1 wassignificantly decreased in the MICD group compared withthe Sham group (119901 lt 001) however the density of dendriticspines of the hippocampal CA1 in the MICD+EA group wasmore than that of the MICD group and the MICD+non-EAgroup (119901 lt 001) In brief EA treatment triggered large-scale remodeling of dendrites in the hippocampal area CA1
34 EA Enhanced the Number of Hippocampus CA1 SynapsesinMICD Rats Furthermore we observed the effect of EA on
Neural Plasticity 5
Esca
pe la
tenc
y (s
)
Day 10 Day 11 Day 12 Day 13Days after MICD
ShamMICD
MICD+EAMICD+non-EA
lowastlowast
lowastlowast
lowastlowastlowastlowast
and
and
andand
0
10
20
30
40
50
60
70
80
90
100
(a)
Day 10 Day 11 Day 12 Day 13Training time
ShamMICD
MICD+EAMICD+non-EA
0
200
400
600
800
1000
1200
Dist
ance
trav
elle
d (c
m)
(b)
Sham MICD
MICD+EA MICD+non-EA
III
III IV
III
III IV
III
III IV
III
III IV
(c)
Sham MICD MICD+EA MICD+non-EA
lowastlowast
and
Prob
e tim
e
0
1
2
3
4
5
6
7
(d)
ShamMICD
MICD+EAMICD+non-EA
lowastlowast and
ti
me s
pent
in ea
ch q
uadr
ant
0
20
40
60
Opp
osite
Left
adja
cent
Targ
etRi
ght a
djac
ent
Opp
osite
Left
adja
cent
Targ
etRi
ght a
djac
ent
Opp
osite
Left
adja
cent
Targ
etRi
ght a
djac
ent
Opp
osite
Left
adja
cent
Targ
etRi
ght a
djac
ent
(e)
Sham MICD MICD+EA MICD+non-EA
lowastlowast
and
ti
me s
pent
in ta
rget
qua
dran
t
0
5
10
15
20
25
30
35
40
(f)
Figure 2 Evaluation of spatial reference learning and memory by Morris water maze test at Days 10ndash14 after EA (a and b) Mean escapelatency time and path length during the orientation navigation test on Days 10ndash13 after EA treatment (c and d) Times the rats crossed overprevious platform location on Day 14 after EA treatment during the spatial memory test in different groups (e)The percentage of times spentin each quadrant in all probe trials is shown on Day 14 after EA treatment (f) The times of passing the hidden platform position on Day 14after EA treatment (each group lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD group andand119901 lt 001 and119901 lt 005versus the MICD+EA group) EA electroacupuncture MICD middle cerebral artery occlusion induced cognitive deficits
6 Neural Plasticity
MICD+non-EA
MICD+EA
MICD
Sham
(a)
MICD+non-EA
MICD+EAMICD
Lesio
ns v
olum
es (
)
0
10
20
30
40
(b)
MICD+non-EA
MICD+EA
MICD
Sham
(c)
MICD+non-EA
MICD+EAMICD
Lesio
ns v
olum
es (
)
0
10
20
30
and
(d)
Figure 3 T2WI signal changes before and after EA (a and c)Thebrain lesionsweremeasured byT2WI (slices from 11 to 13) in the Shamgroupthe MICD group the MICD+EA group and the MICD+non-EA group before (a) and after (c) EA (b and d) Lesion volume is representedas a percentage of the total brain volume and data are presented as the mean plusmn standard deviation from 12 individual rats in each group(lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD group andand119901 lt 001 and119901 lt 005 versus the MICD+EA group)T2WI T2-weighted magnetic resonance imaging MICD middle cerebral artery occlusion induced cognitive deficits
ultrastructural morphology of hippocampal CA1 pyramidalneurons Obviously the density of synapses in the MICDgroup was decreased compared with the Sham group (119901 lt001 Figures 5(a) and 5(b)) whereas there was amplifying inthe number of synapses in the MICD+EA group comparedwith the MICD group and the MICD+non-EA group (119901 lt005 or 119901 lt 001 Figures 5(a) and 5(b))
Taken together these results suggest that EA couldimprove synaptic-dendritic plasticity in vivo
35 EA Promoted LIMK1 Expression and Phosphorylation ofHippocampus CA1 in MICD Rats To explore the underly-ing molecular mechanism of EA-induced synaptic-dendriticplasticity the levels of total LIMK1 and phospho-LIMK1 (P-LIMK1 Thr508) in hippocampal CA1 were investigated Asshown in Figures 6(a) and 6(b) the expression of total LIMK1of hippocampal CA1was significantly decreased in theMICDgroup compared with the Sham group (119901 lt 001) Howeverthe total LIMK1 level in the MICD+EA group was morethan that of the MICD group and the MICD+non-EA group(119901 lt 001 Figures 6(a) and 6(b)) Moreover the changes ofphospho-LIMK1 (Thr508) in groups were similar to the totalLIMK1 expression The level of P-LIMK1 was significantly
increased in the MICD+EA group compared with the MICDgroup and the MICD+non-EA group (119901 lt 001 Figures 6(a)and 6(c))
36 EA Regulated miR-134 Expression in Hippocampal CA1 inMICD Rats To identify the role of synaptic-dendrite-relatedmiR-134 in hippocampus CA1 the levels were detected Asshown in Figure 7 the expression ofmiR-134was significantlyincreased in theMICD group comparedwith the Sham group(119901 lt 001) However repeated EA treatment significantlydecreased the expression of miR-134 compared with theMICD group and the MICD+non-EA group (119901 lt 005)
4 Discussion
Ischemic stroke leads to a high incidence of long-termcognitive impairment which is strongly associated with lossof hippocampal neurons and synapses [34] While review-ing ancient Chinese documents regarding acupuncture andcognitive impairment-related terms we discovered that theDU20 andDU24 acupoints were themost frequently selectedacupoints for cognitive impairment-related rehabilitation inChina A number of studies have shown that EA can improve
Neural Plasticity 7
MICD+non-EAMICD+EA
MICDSham
(a)
MICD+non-EA
MICD+EA
MICD
Sham
(b)
MICD+non-EA
MICD+EAMICDSham0
5
10
15
20
lowastlowastandand
Spin
e den
sity
(10
um)
(c)
Figure 4 The structures and densities of dendritic spine in the hippocampus of rats (a) Representative images of dendritic spine in thehippocampus of each group (Golgi staining times50) (b) Representative images of dendritic spine density and morphology from the pyramidalcell layer of hippocampal CA1 area in each group (Golgi staining times1000) (c)The density of dendritic spine was analyzed in hippocampal CA1pyramidal cells in each group (119899 = 6group lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD group andand119901 lt 001and119901 lt 005 versus the MICD+EA group) All experiments were repeated three times
learning and memory ability and stimulate consciousness[30 31 35]These results indicated that EA could be a comple-mentary therapy for cognitive impairment after stroke Thepresent study found that EA at theDU20 andDU24 acupointsshortened time to find the platform and increased the timesof crossing the position of the platform compared to untreat-ment or nonacupoint EA (stimulation control) in Morriswater maze test suggesting that EA could improve spatialreference learning and memory ability in MCAO-inducedcognitive deficit (MICD) rats Moreover it is worth mention-ing that the path length in the Morris water maze in the fourgroups showed no significant difference indicating that thetime spending ofMICD rats to find the objectwas not affectedby themotor functionHowever themechanisms of cognitivetreatment involved are far from being fully elucidated
Firstly the cognitive deficit-related brain regions weredetermined by a small animal MRI in rats with ischemicstroke T2-weighted imaging showed that MICD caused thelesions of left cortex hippocampus corpus striatum andthalamus regions before EA It had beennoticed that the brain
lesion exhibited amild spontaneous recovery performance inMICD rats However the lesion regions comprising approxi-mately 23of thewhole brain volumewere reduced to 15byEA treatment indicating that EA could attenuate cortex andhippocampus corpus striatum and thalamus region lesionsin theMICD rats Some of these regions such as the hippoca-mpus and cortex are essential for regulating learning andmemory behaviors including spatial exploration [36] Studieshave confirmed that the hippocampus regions play a veryimportant role in learning and memory through their spe-cific structure and location which connect with other brainregions together [37] Furthermore in order to identify thehippocampus function in cognitive deficit the hippocampalmorphology staining was observed
Using Golgi staining and electron microscopy we demo-nstrated that the density of dendritic spine and the number ofsynapses in hippocampal CA1 pyramidal cells were obviouslyreduced at Day 14 after MICD However EA can rescue theloss of dendritic spine and synapses in hippocampal CA1region Moreover EA promoted synaptic-dendritic spine
8 Neural Plasticity
MICD+non-EAMICD+EA
MICDSham
(a)
MICD+non-EA
MICD+EAMICDSham0
2
4
6
8
10
The n
umbe
r of s
ynap
ses (10cm
2)
lowastlowast andand
(b)
Figure 5The number of synapses in hippocampal CA1 pyramidal cells (a) Representative electronmicrographic images of synapses (arrows)in the CA1 region of each group (magnification times50000) (b) Histogram shows the significant difference in the mean plusmn SEM of the numberof synapses in each group (119899 = 5group lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD group andand119901 lt 001and119901 lt 005 versus the MICD+EA group)
MICD+non-EAMICD+EAMICD
120573-Actin
P-LIMK1
T-LIMK1
Sham
(a)
MICD+non-EA
MICD+EAMICDSham00
02
04
06
08
10
Tota
l LIM
K1120573
-act
in
lowastlowastandand
(b)
MICD+non-EA
MICD+EAMICDSham00
02
04
06
08
10
lowastlowastandand
P-LI
MK1
T-L
IMK1
(c)
Figure 6 EA amplified the LIMK1 level in the hippocampus CA1 pyramidal cells (a) The expression of total LIMK1 and phosphorylatedLIMK1was evaluated usingwestern blotting (b and c)Histogram shows significant difference of the levels of P-LIMK1 and phosphor-LIMLK1in each group Data are means plusmn SEM (119899 = 6group lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD groupandand119901 lt 001 and119901 lt 005 versus the MICD+EA group)
Neural Plasticity 9
0
2
4
6
8
10
12
miR
-134
relat
ive e
xpre
ssio
n to
U6 lowastlowast
and
MICD+non-EA
MICD+EAMICDSham
Figure 7 Changes of miR-134 expression in the hippocampusCA1 cells The changes of miR-134 expression level were measuredusing quantitative RT-PCR U6 was used as internal control forquantification of miRNAs Data are mean plusmn SEM (119899 = 6grouplowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus theMICD group andand119901 lt 001 and119901 lt 005 versus the MICD+EA group)
regeneration Dendritic spine is the tiny protrusions withmulti-dense body and ion channels on the surface of varioustypes of neurons which serve as cellular substrates of brainconnectivity and the major sites of information processingin the brain [38 39] Evidence is increasing that synaptic-dendritic plasticity enables the encoding of memory [40]Indeed themodification on dendritic spine number size andshape is of importance for the plasticity of synapses accom-panied by hippocampus-dependent learning and memory[41 42] Studies have well described that dendritic spineremodeling in ischemia surrounding areas may contribute tocognitive functional recovery after stroke [43 44]
Increasing evidence showed that LIMK1 protein is acti-vated at dendritic spine membranes [45] LIMK1 positivecells were located in the CA1 region of the hippocampuswhich is important in the regulation of the spinemorphologyand synaptic function in vivo A prominent form of longlasting synaptic plasticity is thought to be critical to memoryformation [26] The present study found that the expressionof total LIMK1 and phospho-LIMK1 in hippocampal CA1wassignificantly decreased in the MICD rats however EA couldenhance the total LIMK1 and phospho-LIMK1 levels to pro-mote synaptic-dendritic plasticity in hippocampal CA1 area
In addition we have identified a dendritically localizedmiRNA that regulated the expression of the synaptic LIMK1protein thereby controlling dendritic pine sizemiR-134 is thefirst discovered dendritic microRNA enriched in the neu-ronal dendrites of rat hippocampal neurons and negativelycontrols the size of dendritic spines [46] This effect ismediated by miR-134 inhibition of the translation of LIMK1mRNA [47] The present study showed that the expression ofmiR-134 was obviously increased in theMICD rats Howeverrepeated EA treatment could relieve the upregulation of miR-134 expression in hippocampal CA1 area
In conclusion the study indicated that EA at the DU20and DU24 acupoints could ameliorate cognitive impairmentof MICD rats through the regulation of synaptic-dendritic
plasticity of the hippocampal CA1 area and the expressionand phosphorylation of synaptic LIMK1 protein were acti-vated to control dendritic spine number size and shapeFurthermore the mechanism of LIMK1 increase induced byEA treatmentmay be associated withmiR-134 which is local-ized in hippocampal CA1 by negatively regulating LIMK1 toenhance synaptic-dendritic plasticity in the recovery stage ofischemic stroke
Competing Interests
The authors declare no financial or commercial conflict ofinterests
Authorsrsquo Contributions
Weilin Liu Jie Wu and Jia Huang contributed equally to thiswork
Acknowledgments
The present study was supported by the Natural ScienceFoundation of China (no 81403450) and the Natural ScienceFoundation of Fujian Province (no 2016J01382)
References
[1] V L Roger ldquoHeart disease and stroke statisticsmdash2011 update areport from the American Heart Associationrdquo Circulation vol131 no 4 pp 29ndash322 2014
[2] T E Nichols and A P Holmes ldquoNonparametric permutationtests for functional neuroimaging a primer with examplesrdquoHuman Brain Mapping vol 15 no 1 pp 1ndash25 2002
[3] T K Tatemichi D W Desmond Y Stern M Paik M Sanoand E Bagiella ldquoCognitive impairment after stroke frequencypatterns and relationship to functional abilitiesrdquo Journal ofNeurology Neurosurgery amp Psychiatry vol 57 no 2 pp 202ndash207 1994
[4] F Liu Z-M Li Y-J Jiang and L-D Chen ldquoA meta-analysis ofacupuncture use in the treatment of cognitive impairment afterstrokerdquo Journal of Alternative amp Complementary Medicine vol20 no 7 pp 535ndash544 2014
[5] W Li R Huang R A Shetty et al ldquoTransient focal cerebralischemia induces long-termcognitive function deficit in an exp-erimental ischemic stroke modelrdquo Neurobiology of Disease vol59 pp 18ndash25 2013
[6] P S Sachdev X Chen H Brodaty CThompson A Altendorfand W Wen ldquoThe determinants and longitudinal course ofpost-stroke mild cognitive impairmentrdquo Journal of the Inter-national Neuropsychological Society vol 15 no 6 pp 915ndash9232009
[7] A-J Liu J-H Li H-Q Li et al ldquoElectroacupuncture foracute ischemic stroke ameta-analysis of randomized controlledtrialsrdquoThe American Journal of Chinese Medicine vol 43 no 8pp 1541ndash1566 2015
[8] X Ding C-Y Li Q-S Wang et al ldquoPatterns in default-modenetwork connectivity for determining outcomes in cognitivefunction in acute stroke patientsrdquo Neuroscience vol 277 no 10pp 637ndash646 2014
10 Neural Plasticity
[9] H Alipanahzadeh M Soleimani S Soleimani Asl B Pourhey-dar A Nikkhah and M Mehdizadeh ldquoTransforming growthfactor-alpha improves memory impairment and neurogenesisfollowing ischemia reperfusionrdquo Cell Journal vol 16 no 3 pp315ndash324 2014
[10] H Eichenbaum ldquoHippocampus cognitive processes and neuralrepresentations that underlie declarative memoryrdquoNeuron vol44 no 1 pp 109ndash120 2004
[11] T Kadar S Dachir B Shukitt-Hale and A Levy ldquoSub-regionalhippocampal vulnerability in various animal models leading tocognitive dysfunctionrdquo Journal of Neural Transmission vol 105no 8-9 pp 987ndash1004 1998
[12] J B Pereira C Valls-Pedret E Ros et al ldquoRegional vulnera-bility of hippocampal subfields to aging measured by structuraland diffusion MRIrdquo Hippocampus vol 24 no 4 pp 403ndash4142014
[13] S Knafo G Ariav E Barkai and F Libersat ldquoOlfactory learn-ing-induced increase in spine density along the apical dendritesof CA1 hippocampal neuronsrdquo Hippocampus vol 14 no 7 pp819ndash825 2004
[14] S Schacher and J-Y Hu ldquoThe less things change the more theyare different contributions of long-term synaptic plasticity andhomeostasis to memoryrdquo Learning and Memory vol 21 no 3pp 128ndash134 2014
[15] K Park H Heo M E Han et al ldquoLearning-induced synapticpotentiation in implanted neural precursor cell-derived neu-ronsrdquo Scientific Reports vol 5 article 17796 2015
[16] S Knafo F Libersat and E Barkai ldquoDynamics of learning-induced spine redistribution along dendrites of pyramidalneurons in ratsrdquo European Journal of Neuroscience vol 21 no4 pp 927ndash935 2005
[17] V C Foletta N Moussi P D Sarmiere J R Bamburg and OBernard ldquoLIM kinase 1 a key regulator of actin dynamics iswidely expressed in embryonic and adult tissuesrdquo ExperimentalCell Research vol 294 no 2 pp 392ndash405 2004
[18] M Endo K Ohashi Y Sasaki et al ldquoControl of growth conemotility andmorphology by LIMkinase and slingshot via phos-phorylation and dephosphorylation of cofilinrdquo Journal of Neu-roscience vol 23 no 7 pp 2527ndash2537 2003
[19] C Jarrold A D Baddeley and C Phillips ldquoLong-termmemoryfor verbal and visual information in Down syndrome andWilliams syndrome performance on the doors and people testrdquoCortex vol 43 no 2 pp 233ndash247 2007
[20] Y Meng Y Zhang V Tregoubov et al ldquoAbnormal spinemorphology and enhanced LTP in LIMK-1 knockout micerdquoNeuron vol 35 no 1 pp 121ndash133 2002
[21] Z Todorovski S Asrar J Liu et al ldquoLIMK1 regulates long-termmemory and synaptic plasticity via the transcriptional factorCREBrdquo Molecular and Cellular Biology vol 35 no 8 pp 1316ndash1328 2015
[22] G M Schratt F Tuebing E A Nigh et al ldquoA brain-specificmicroRNA regulates dendritic spine developmentrdquo Nature vol439 no 7074 pp 283ndash289 2006
[23] F Capitano J Camon V Ferretti et al ldquomicroRNAs modulatespatial memory in the hippocampus and in the ventral striatumin a region-specific mannerrdquo Molecular Neurobiology vol 53no 7 pp 4618ndash4630 2015
[24] J GaoW-YWang Y-WMao et al ldquoA novel pathway regulatesmemory and plasticity via SIRT1 andmiR-134rdquoNature vol 466no 7310 pp 1105ndash1109 2010
[25] J Remenyi M W M van den Bosch O Palygin et al ldquomiR-132212 knockout mice reveal roles for these miRNAs in regu-lating cortical synaptic transmission and plasticityrdquo PLoS ONEvol 8 no 4 Article ID e62509 p 65 2013
[26] L An J LiuWW Li et al ldquoDistribution of LIM domain kinase1 in the olfactory bulb cerebral cortex hippocampus and cere-bellum of the AppPS+minus micerdquo Genetics amp Molecular Researchvol 14 no 4 pp 17244ndash17251 2015
[27] R Lin Y Wu J Tao et al ldquoElectroacupuncture improves cog-nitive function through Rho GTPases and enhances dendriticspine plasticity in rats with cerebral ischemia-reperfusionrdquoMolecular Medicine Reports vol 13 no 3 pp 2655ndash2660 2016
[28] P Castaneda M Munoz G Garcıa-Rojo et al ldquoAssociation ofN-cadherin levels and downstream effectors of Rho GTPaseswith dendritic spine loss induced by chronic stress in rathippocampal neuronsrdquo Journal of Neuroscience Research vol93 no 10 pp 1476ndash1491 2015
[29] A Romarowski M A Battistone F A La Spina et al ldquoPKA-dependent phosphorylation of LIMK1 and Cofilin is essentialformouse sperm acrosomal exocytosisrdquoDevelopmental Biologyvol 405 no 2 pp 237ndash249 2015
[30] R Lin K Yu X Li et al ldquoElectroacupuncture ameliorates post-stroke learning and memory through minimizing ultrastruc-tural brain damage and inhibiting the expression of MMP-2andMMP-9 in cerebral ischemia-reperfusion injured ratsrdquoMol-ecular Medicine Reports vol 14 no 1 pp 225ndash233 2016
[31] X Feng S Yang J Liu et al ldquoElectroacupuncture amelioratescognitive impairment through inhibition of NF-120581B-mediatedneuronal cell apoptosis in cerebral ischemia-reperfusion inj-ured ratsrdquo Molecular Medicine Reports vol 7 no 5 pp 1516ndash1522 2013
[32] Z K Wang N I Guang-Xia K Liu et al ldquoResearch on thechanges of IL-1 receptor and TNF-120572 receptor in rats with cere-bral ischemia reperfusion and the chronergy of acupunctureinterventionrdquo Zhongguo Zhen Jiu vol 32 no 11 pp 1012ndash10182012
[33] X S LiuM Chopp R L Zhang and Z G Zhang ldquoMicroRNAsin cerebral ischemia-induced neurogenesisrdquo Journal of Neu-ropathology amp Experimental Neurology vol 72 no 8 pp 718ndash722 2013
[34] X He T Yan R Chen and D Ran ldquoAcute effects of electro-acupuncture (EA) on hippocampal long term potentiation(LTP) of perforant path-dentate gyrus granule cells synapserelated to memoryrdquo Acupuncture and Electro-TherapeuticsResearch vol 37 no 2-3 pp 89ndash101 2012
[35] W Dong W Guo X Zheng et al ldquoElectroacupunctureimproves cognitive deficits associated with AMPK activation inSAMP8 micerdquo Metabolic Brain Disease vol 30 no 3 pp 777ndash784 2015
[36] B Milner and D Klein ldquoLoss of recent memory after bilateralhippocampal lesions memory and memoriesmdashlooking backand looking forwardrdquo Journal of Neurology Neurosurgery ampPsychiatry vol 87 no 3 article 230 2016
[37] A Veronica Witte L Kerti D S Margulies and A FloelldquoEffects of resveratrol on memory performance hippocampalfunctional connectivity and glucose metabolism in healthyolder adultsrdquo Journal of Neuroscience vol 34 no 23 pp 7862ndash7870 2014
[38] T Bonhoeffer and R Yuste ldquoSpine motility phenomenologymechanisms and functionrdquoNeuron vol 35 no 6 pp 1019ndash10272002
Neural Plasticity 11
[39] H Qiao M Li C Xu H Chen S An and X Ma ldquoDendriticspines in depression what we learned from animal modelsrdquoNeural Plasticity vol 2016 Article ID 8056370 26 pages 2016
[40] S C McQuown R M Barrett D P Matheos et al ldquoHDAC3 isa critical negative regulator of long-term memory formationrdquoJournal of Neuroscience vol 31 no 2 pp 764ndash774 2011
[41] K-O Lai and N Y Ip ldquoStructural plasticity of dendritic spinesthe underlying mechanisms and its dysregulation in brain dis-ordersrdquo Biochimica et Biophysica ActamdashMolecular Basis of Dis-ease vol 1832 no 12 pp 2257ndash2263 2013
[42] C H Bailey E R Kandel and K M Harris ldquoStructural com-ponents of synaptic plasticity and memory consolidationrdquo ColdSpring Harbor Perspectives in Biology vol 7 no 7 Article IDa021758 2015
[43] C E Brown P Li J D Boyd K R Delaney and T HMurphy ldquoExtensive turnover of dendritic spines and vascularremodeling in cortical tissues recovering from strokerdquo Journalof Neuroscience vol 27 no 15 pp 4101ndash4109 2007
[44] R Mostany T G Chowdhury D G Johnston S A PortonovoS T Carmichael and C Portera-Cailliau ldquoLocal hemodynam-ics dictate long-term dendritic plasticity in peri-infarct cortexrdquoJournal of Neuroscience vol 30 no 42 pp 14116ndash14126 2010
[45] J George C Soares A Montersino J-C Beique and G MThomas ldquoPalmitoylation of LIM kinase-1 ensures spine-specificactin polymerization and morphological plasticityrdquo eLife vol2015 no 4 Article ID e06327 2015
[46] S Bicker M Lackinger K Weiszlig and G Schratt ldquoMicroRNA-132 -134 and -138 a microRNA troika rules in neuronaldendritesrdquo Cellular and molecular life sciences CMLS vol 71no 20 pp 3987ndash4005 2014
[47] H-C Tai and E M Schuman ldquoMicroRNA microRNAs reachout into dendritesrdquoCurrent Biology vol 16 no 4 pp R121ndashR1232006
Submit your manuscripts athttpswwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

2 Neural Plasticity
plasticity of dendritic spines and the morphological changessuch as expansion and contraction [13] Dendritic spines altertheir shape to make the information spreading more easilyand influence the synaptic efficacy (ie long-term potenti-ation and long-term depression) [14 15] which have beenwidely considered as a cellular mechanism for learning andmemory [16] LIM domain kinase (LIMK1) is enriched inboth axonal and dendritic growth cones of hippocampalpyramidal neurons in rats [17] LIMK1 encodes a serinethre-onine protein kinase that regulates the actin cytoskeleton byphosphorylating and inactivating the actin depolymerizationfactor (ADF)cofilin [18] In addition LIMK1 is also knownas having an important role in synapse and dendritic spinefunction It has been reported that the knockout mice lackingLIMK1 are severely impaired in dendritic spine morphologyand hippocampal long-term potentiation [19 20] Evidenceshowed that LIMK1 regulated long-term memory (LTM)and long lasting synaptic plasticity through interacting withand activating cyclic AMP response element-binding protein(CREB) [21]
In addition a potential role for microRNAs (miRNAs ormiRs) in synaptic function has been particularly intriguinggiven the evidence that a brain-specific miRNA contributesto synaptic development maturation andor plasticity [22]miRNAs are endogenous noncoding RNAs that mediate theposttranscriptional regulation of gene expression mainly bybinding to the 31015840-untranslated region of messenger RNAs(mRNAs) [23] A number of miRNAs have been isolatedfrom nervous system and a recent study has demonstrateda crucial role for dynamically regulating synaptic plasticity[24 25] Moreover miRNAs have been implicated in hippo-campus-dependent function which have a significant poten-tial in learning and memory formation regulating LIMK1expression to induce synaptic-dendritic plasticity [22] Den-dritic mRNAs encode diversified functionalities in hippo-campal pyramidal neurons and play an important role insynaptic plasticity as well as learning and memory [26]
Therefore miRNA-LIMK1 can be considered as a targetfor cognitive deficit Our previous study has shown that EAat Baihui (DU20) and Shenting (DU24) acupoints couldimprove cognitive impairment through Rho GTPases toenhance dendritic plasticity in rats with ischemic stroke [27]Interestingly it has been suggested that the activation of theRho GTPases signaling is essential for LIMK1 activation byphosphorylation on threonine 508 which is widely known asmaster regulator of actin dynamics [28 29] Thus the studyaimed to elucidate whether EA at the DU20 and DU24acupoints could improve the cognitive deficits in rats withischemic stroke via miRNA-LIMK1-mediated synaptic plas-ticity to enhance spatial reference learning and memory
2 Materials and Methods
21 Animal Ethics Statement 96 male Sprague-Dawley rats(weight 250 plusmn 20 g) were obtained from Shanghai SLAC Lab-oratoryAnimalCo Ltd (Shanghai China SCXK2013-0005)and housed under pathogen-free conditions with a 12 hlightdark cycleThe study was approved by the Committee ofFujian University of Traditional Chinese Medicine (protocol
FUTCM-2015008) and was performed in accordance withthe national guidelines for the care and use of laboratoryanimals For euthanasia 3 sodium pentobarbital (40mgkgbody weight ip) was used Middle cerebral artery occlusion(MCAO) surgery was carried out under general anesthesia(15 isoflurane in 685 N2O and 30 O2) All efforts weremade to minimize suffering
22 Experimental Procedures TheMCAO-induced cognitivedeficit (MICD)modelwas established as previously described[5 30] Rats were randomly assigned to four groups accordingto the random number table (119899 = 24 rats each group) as fol-lows (1) Sham group (2)MICD group (3)MICD+EA groupand (4) MICD+non-EA group The left middle cerebralartery was occluded by a 4-0 nylon monofilament (023mmin diameter Jialing-bio China) Focal cerebral ischemiawas monitored using transcranial temporal laser Doppler(BIOPAC Systems Goleta CA USA) and an 80 decrease inblood flow after the occlusion was recorded After 90min ofocclusion reperfusion was achieved by pulling out the fila-ment to restore blood flow Sham-operated rats of the Shamgroup underwent the same procedure but arterial occlusionwas not performed
After 24 hours of MCAO surgery rats of the Sham groupand the MICD group received no treatment Rats of theMICD+EAgroupwere givenEA treatment for 30min per dayfor 14 consecutive days The EA needles (diameter 03mmneedle purchased from Hualun acupuncture of Suzhou CoLtd Suzhou China) were inserted at a depth of 2-3mm intothe Baihui (DU20 located in the median of frontalis) andShenting (DU24 located in the median of the parietal bone)acupoints [31] Stimulation was then generated using the EAapparatus [ModelG6805 ShanghaiHuayi (Group)CompanyLtd Shanghai China] and the stimulation parameters wereset as follows dilatational waves of 1sim20Hz (adjusted to themuscle twitch threshold) peak voltage of 6V and 02mAintensity [31] Rats of the MICD+non-EA group were givenEA treatment at the bilateral nonacupoints (located in thecostal region and 10mmdistal to the iliac crest) for 30min perday for 14 consecutive days [32]The EA needles and stimula-tion parameters of the bilateral nonacupoints were consistentwith the EA treatment Manipulators were experienced andblinded to the ratrsquos group
23 Behavioral Assessment Behavioral testingwas conductedby researchers who were blinded to the ratrsquos group At 10 daysafter EA all the rats were subjected to the Morris water mazetest (Shanghai Xinruan Information Technology Co LtdShanghai China) to evaluate spatial reference learning andmemory (Figure 1)TheMorris water maze consisted of a cir-cular pool (diameter 150 cm height 60 cm) filled with water(depth of 30 cm and temperature of 25 plusmn 2∘C) A circularescape platform (diameter 12 cm height 29 cm) was sub-merged 2 cm below the water surface in the middle of thethird quadrant of the pool and the reference objects aroundthe pool were placedMorris watermaze tasksmainly includeorientation navigation and space exploration trials Duringthe first set of trials each rat was placed in the water at eachof the four equidistant locations to the platform When the
Neural Plasticity 3
0
Sham or MCAO surgery Sacrificed
Morris water test
Electroacupuncture
Baseline T2WI T2WI
24 h 14 d10 d7 d1 d
Figure 1 Experimental design used in the study
rats arrived at the platform within the 90 sec time restrictionand remained on it for 3 sec they were considered to havefound the platform and were scored by the time takenlengthof the route When the rats were unable to find the platformwithin 90 sec they were placed on the platform for 10 sec andthe time scorewas 90 secThe time taken and the length of theroute bywhich each rat found the safe platformwere recordedby the computerThe average of the time taken and the lengthof the route for the four quadrants as a result of each rat wereassessed every day The duration of the first set of trials wasperformed on Days 10 to 13 after EA The second part of theexperiment was performed on Day 14 to examine the timein which rats found the location of the platform within the90 sec time restriction which tested their ability to rememberthe position of the platform After all trials the rats weredried thoroughly with a hair drier and returned to their cages(Guangzhou RiboBio Co Ltd Guangzhou China) Morriswater maze test was repeated three times
24 Measurement of Brain Lesions Animal MRI scans wereperformed on a 70 T MRI scanner (7020USR BioSpecBruker Biospin Gmbls Germany) using a 38mmbirdcage ratbrain quadrature resonator for radiofrequency transmissionand reception Animals were anesthetized with isofluraneO2(with 3 induction for 5min and 12ndash15 maintenance inorder to let the rats in the depth of anesthesia state) andkept warm with circuit After anesthesia the rat was putin prone position on a custom-made holder to minimizehead motion set the location of head position and performreal-time monitor of the breathing rate and maintained in40 breathsmin Ratrsquos temperature is maintained at 33 plusmn 2∘Cin the process of scanning holder to minimize head motionwhile respiration was maintained
T2-weighted imaging (T2WI) in three planes with afast spin echo (FSE) pulse sequence was first acquired tocontrol rat head positioning T2WI scan was acquired usinga Rapid Acquisition with Relaxation Enhancement (RARE)pulse sequence with the following parameters field of view =32mmtimes 32mmmatrix size = 256times 256 repetition time (TR)= 4200ms echo time (TE) = 35ms slice thickness = 10mmand slice gap = 0mm
Image J analysis and processing system was applied forT2W images the percentage of the brain lesions = brainlesions volumewhole brain volume times100
25 Golgi Staining Golgi staining of brain was performedusing an FD Rapid GolgiStain Kit following the manufac-turerrsquos instructions (FD Neurotechnologies Inc ColumbiaMD USA) The removed rat brains were placed incubated
in a mixture of A and B solutions from the kit and storedin the dark at room temperature following which they weretransferred into solution C from the kit stored at 4∘C for 7days Finally brains were then frozen and coronal sections(150 120583m) were made using a cryostat (Leica CM3050S LeicaMicrosystems K K Tokyo Japan) The sections incubatedin a mixture of D and E solutions from the kit and thendehydrated in alcohol (50 70 90 and 100 for 5min each)cleared in xylene and were cover-slipped The images finallywere viewed under a microscope (Leica DM6000 B LeicaMicrosystems Wetzlar Germany) The slides were reviewedby two or three pathologists blind to the study
26 Transmission Electron Microscopy Four rats in eachgroup were anesthetized and the left ventricle was perfusedwith 200mL of saline followed by 400mL 4 paraformalde-hyde (pH 74) The tissue was taken from the left ischemichippocampus cut into 1mm3 size cubes and fixed in 1paraformaldehyde with 1 lanthanum nitrate tracer for 24 hfollowed by fixation with 3 glutaraldehyde for 24 h Sampleswere fixed in 1 osmium tetroxide for 2 h and dehydrated ingraded ethanol 1 lanthanum nitrate tracer (LNT) solutionand embedded in araldite Ultrathin hippocampal CA1 sliceswere obtained (90 nm) then they were stained with uranylacetate and lead citrate and observed under TEM (H-7650Hitachi Ltd Tokyo Japan) The photos were obtained onhp digital CCD camera (SIS4 million voxel) Images wereacquired digitally from a randomly selected pool of 10 to 15fields under each condition
27 Western Blotting Isolated left hippocampus tissue waslysed in 100 120583L radioimmunoprecipitation assay (RIPA) buf-fer (Beyotime Haimen Jiangsu China) plus protease inhibi-tors Total protein (50120583g) was loaded into 10 SDS-PAGEgels electrophoresed and then transblotted onto polyvinyli-dene difluoride membranes (Immobilon-P Millipore Bil-lerica MA) in a Tris-glycine transfer buffer After beingblocked in 5 milk in PBST for 1 h at room temperaturewith shaking membranes were incubated with antibodiesovernight at 4∘C against the following anti-LIMK1 antibody(dilution 1 1000 ab81046 Abcam) and anti-LIM kinase1 antibody (phospho-Thr508) (dilution 1 1000 ab131341Abcam) and 120573-actin (dilution 1 1000 ab189073 Abcam)The following day membranes were incubated in 5 milk(in TBST) with an anti-goat or anti-mouse IgG antibody(dilution 1 5000 PerkinElmer Life Sciences Waltham MA)for 1 h at room temperature with shaking Membranes werewashed a minimum of four times (10min per wash) in PBST
4 Neural Plasticity
between each antibody treatment Detected bands were visu-alized using enhanced chemiluminescence and images werecaptured using a Bio-Image system (Bio-Rad LaboratoriesInc Hercules USA) Western blotting was repeated threetimes
28 Real Time Quantitative RT-PCR The expression of miR-NAs was determined by real time quantitative reverse tran-scription polymerase chain reaction (RT-PCR) Total RNAwas extracted from hippocampal region of the ipsilaterallesion tissue using the TRIzol Reagent (Life Technologies(AB amp Invitrogen) Carlsbad USA) Then extracted totalRNA was reverse transcribed to generate cDNA accord-ing to manufacturer instructions of Revert AidTM First-Strand cDNA Synthesis Kit (Thermo Fisher Scientific Bei-jing China) The reverse transcription reaction was ampli-fied using a Bio-Rad CFX96 Detection System (Bio-RadHercules CA USA) with the Plexor One-Step qRT-PCR System (Promega Madison WI USA) For miRNAamplification the following primers were used rno-miR-134 (RmiRQP0168 GeneCopoeia Guangzhou China) Thefold change in relative mRNA and miRNA expression wasdetermined using the 2minusΔΔCt method as described previously[33] and U6 snRNA (RQP047936 GeneCopoeia) as mRNAand miRNA internal control respectively
29 Statistical Analysis Statistical analysis was performedwith the SPSS package for Windows statistical analysissoftware (Version 180 SPSS Inc Chicago IL USA) Thedata from all groups were determined by one-way analysisof variance (ANOVA) and Studentrsquos 119905-testsThe homogeneityof variance was analyzed using the least significant differ-ence method and missing variance using the Games-Howellmethod Intergroup comparisons of the brain lesions volumeat different time points were performed with paired-samples119905-tests All data are presented as mean plusmn standard error andthe significance was regarded as at least 119901 lt 005 All finalresults were analyzed in a blinded manner
3 Results
31 EA Improved Cognitive Impairment in MICD Rats Toassess the effect of EA on spatial reference learning andmemory impairments in MICD rats the Morris water maze(MWM) test was performed All groups of rats learned to findthe platform and the latency time to reach the platform wasreduced in the four training days However learning abilitywas significantly reduced in MICD rats compared with theSham group (119901 lt 001 Figure 2(a)) the MICD rats treatedwith EA significantly took less time to find the platform com-pared with the MICD group and the MICD+non-EA group(119901 lt 001 or 119901 lt 005 Figure 2(a)) There was no significantdifference about path length among the four groups (119901 gt005 Figure 2(b)) As illustrated in Figures 2(c) and 2(d)tracing images from the MWM test showed that in the spaceexploration test where the platform was removed the MICDrats passed through the original position of the platformfewer times than the Sham group (119901 lt 001 Figure 2(d))
whereas in the MICD+EA group the times where the ratscrossed the position of the platformwere significantly increa-sed compared with the MICD group and the MICD+non-EA group (119901 lt 001 119901 lt 005 Figure 2(d)) As illustratedin Figures 2(e) and 2(f) the percentage of time spent in thetarget quadrant was used for statistical analysis during theprobe trial The date showed that the rats spent more time inthe target quadrant compared to other quadrants (119901 lt 001Figure 2(e)) The MICD group spent less time in the targetquadrant compared with the other groups (119901 lt 001 Fig-ure 2(e))TheMICD+EA group spent more time in the targetquadrant compared with the MICD group and MICD+non-EA group (119901 lt 001 119901 lt 005 Figure 2(f)) Therefore theseresults suggest that acquisition or retention of spatial refer-ence learning and memory was ameliorated in MICD rats byEA treatment
32 EA Attenuated Left Cortex Hippocampus Corpus Stria-tum and Thalamus Lesions in MICD Rats The brain lesionswere determined by T2-weighted imaging (T2WI) (Figures3(a) and 3(c)) before and after EA treatment The volumes ofbrain lesions that included left cortex hippocampus corpusstriatum and thalamus regions in the all groups had nosignificant difference before EA treatment (119901 gt 005 Figures3(a) and 3(b)) There was mild spontaneous recovery for 14days in the brain lesions of the MICD group The volumesof left cortex hippocampus corpus striatum and thalamuslesions showed comprising approximately 23 of the wholebrain in theMICDgroup whereas the brain lesions sizes weresignificantly reduced to 15 in the MICD+EA group (119901 lt001 Figures 3(c) and 3(d)) The difference of brain lesionsbetween the MICD+EA and the MICD+non-EA groupshowed significant changes at Day 14 after EA (119901 lt 005Figures 3(c) and 3(d))
33 EA Increased the Density of Dendritic Spines in the Hip-pocampus ofMICDRats To investigate the function of EAonsynaptic plasticity dendritic spine density in the hippocampalneurons was analyzed by the primary basilar dendrites ofGolgi-stained pyramidal neurons at Days 14 after EA Golgi-Cox staining clearly filled the dendritic shafts and the spinesof neurons frompyramidal neurons (Figure 4(a))Hereby therepresentative hippocampus showed that the density of den-dritic spines was reduced in different degree by macroscopicexamination in the MICD group the MICD+EA group andMICD+non-EA group and the loss of dendritic spines inhippocampal CA1 was obvious in the MICD group Thus asillustrated in Figures 4(b) and 4(c) the density of selecteddendritic spines that derived from hippocampal CA1 wassignificantly decreased in the MICD group compared withthe Sham group (119901 lt 001) however the density of dendriticspines of the hippocampal CA1 in the MICD+EA group wasmore than that of the MICD group and the MICD+non-EAgroup (119901 lt 001) In brief EA treatment triggered large-scale remodeling of dendrites in the hippocampal area CA1
34 EA Enhanced the Number of Hippocampus CA1 SynapsesinMICD Rats Furthermore we observed the effect of EA on
Neural Plasticity 5
Esca
pe la
tenc
y (s
)
Day 10 Day 11 Day 12 Day 13Days after MICD
ShamMICD
MICD+EAMICD+non-EA
lowastlowast
lowastlowast
lowastlowastlowastlowast
and
and
andand
0
10
20
30
40
50
60
70
80
90
100
(a)
Day 10 Day 11 Day 12 Day 13Training time
ShamMICD
MICD+EAMICD+non-EA
0
200
400
600
800
1000
1200
Dist
ance
trav
elle
d (c
m)
(b)
Sham MICD
MICD+EA MICD+non-EA
III
III IV
III
III IV
III
III IV
III
III IV
(c)
Sham MICD MICD+EA MICD+non-EA
lowastlowast
and
Prob
e tim
e
0
1
2
3
4
5
6
7
(d)
ShamMICD
MICD+EAMICD+non-EA
lowastlowast and
ti
me s
pent
in ea
ch q
uadr
ant
0
20
40
60
Opp
osite
Left
adja
cent
Targ
etRi
ght a
djac
ent
Opp
osite
Left
adja
cent
Targ
etRi
ght a
djac
ent
Opp
osite
Left
adja
cent
Targ
etRi
ght a
djac
ent
Opp
osite
Left
adja
cent
Targ
etRi
ght a
djac
ent
(e)
Sham MICD MICD+EA MICD+non-EA
lowastlowast
and
ti
me s
pent
in ta
rget
qua
dran
t
0
5
10
15
20
25
30
35
40
(f)
Figure 2 Evaluation of spatial reference learning and memory by Morris water maze test at Days 10ndash14 after EA (a and b) Mean escapelatency time and path length during the orientation navigation test on Days 10ndash13 after EA treatment (c and d) Times the rats crossed overprevious platform location on Day 14 after EA treatment during the spatial memory test in different groups (e)The percentage of times spentin each quadrant in all probe trials is shown on Day 14 after EA treatment (f) The times of passing the hidden platform position on Day 14after EA treatment (each group lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD group andand119901 lt 001 and119901 lt 005versus the MICD+EA group) EA electroacupuncture MICD middle cerebral artery occlusion induced cognitive deficits
6 Neural Plasticity
MICD+non-EA
MICD+EA
MICD
Sham
(a)
MICD+non-EA
MICD+EAMICD
Lesio
ns v
olum
es (
)
0
10
20
30
40
(b)
MICD+non-EA
MICD+EA
MICD
Sham
(c)
MICD+non-EA
MICD+EAMICD
Lesio
ns v
olum
es (
)
0
10
20
30
and
(d)
Figure 3 T2WI signal changes before and after EA (a and c)Thebrain lesionsweremeasured byT2WI (slices from 11 to 13) in the Shamgroupthe MICD group the MICD+EA group and the MICD+non-EA group before (a) and after (c) EA (b and d) Lesion volume is representedas a percentage of the total brain volume and data are presented as the mean plusmn standard deviation from 12 individual rats in each group(lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD group andand119901 lt 001 and119901 lt 005 versus the MICD+EA group)T2WI T2-weighted magnetic resonance imaging MICD middle cerebral artery occlusion induced cognitive deficits
ultrastructural morphology of hippocampal CA1 pyramidalneurons Obviously the density of synapses in the MICDgroup was decreased compared with the Sham group (119901 lt001 Figures 5(a) and 5(b)) whereas there was amplifying inthe number of synapses in the MICD+EA group comparedwith the MICD group and the MICD+non-EA group (119901 lt005 or 119901 lt 001 Figures 5(a) and 5(b))
Taken together these results suggest that EA couldimprove synaptic-dendritic plasticity in vivo
35 EA Promoted LIMK1 Expression and Phosphorylation ofHippocampus CA1 in MICD Rats To explore the underly-ing molecular mechanism of EA-induced synaptic-dendriticplasticity the levels of total LIMK1 and phospho-LIMK1 (P-LIMK1 Thr508) in hippocampal CA1 were investigated Asshown in Figures 6(a) and 6(b) the expression of total LIMK1of hippocampal CA1was significantly decreased in theMICDgroup compared with the Sham group (119901 lt 001) Howeverthe total LIMK1 level in the MICD+EA group was morethan that of the MICD group and the MICD+non-EA group(119901 lt 001 Figures 6(a) and 6(b)) Moreover the changes ofphospho-LIMK1 (Thr508) in groups were similar to the totalLIMK1 expression The level of P-LIMK1 was significantly
increased in the MICD+EA group compared with the MICDgroup and the MICD+non-EA group (119901 lt 001 Figures 6(a)and 6(c))
36 EA Regulated miR-134 Expression in Hippocampal CA1 inMICD Rats To identify the role of synaptic-dendrite-relatedmiR-134 in hippocampus CA1 the levels were detected Asshown in Figure 7 the expression ofmiR-134was significantlyincreased in theMICD group comparedwith the Sham group(119901 lt 001) However repeated EA treatment significantlydecreased the expression of miR-134 compared with theMICD group and the MICD+non-EA group (119901 lt 005)
4 Discussion
Ischemic stroke leads to a high incidence of long-termcognitive impairment which is strongly associated with lossof hippocampal neurons and synapses [34] While review-ing ancient Chinese documents regarding acupuncture andcognitive impairment-related terms we discovered that theDU20 andDU24 acupoints were themost frequently selectedacupoints for cognitive impairment-related rehabilitation inChina A number of studies have shown that EA can improve
Neural Plasticity 7
MICD+non-EAMICD+EA
MICDSham
(a)
MICD+non-EA
MICD+EA
MICD
Sham
(b)
MICD+non-EA
MICD+EAMICDSham0
5
10
15
20
lowastlowastandand
Spin
e den
sity
(10
um)
(c)
Figure 4 The structures and densities of dendritic spine in the hippocampus of rats (a) Representative images of dendritic spine in thehippocampus of each group (Golgi staining times50) (b) Representative images of dendritic spine density and morphology from the pyramidalcell layer of hippocampal CA1 area in each group (Golgi staining times1000) (c)The density of dendritic spine was analyzed in hippocampal CA1pyramidal cells in each group (119899 = 6group lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD group andand119901 lt 001and119901 lt 005 versus the MICD+EA group) All experiments were repeated three times
learning and memory ability and stimulate consciousness[30 31 35]These results indicated that EA could be a comple-mentary therapy for cognitive impairment after stroke Thepresent study found that EA at theDU20 andDU24 acupointsshortened time to find the platform and increased the timesof crossing the position of the platform compared to untreat-ment or nonacupoint EA (stimulation control) in Morriswater maze test suggesting that EA could improve spatialreference learning and memory ability in MCAO-inducedcognitive deficit (MICD) rats Moreover it is worth mention-ing that the path length in the Morris water maze in the fourgroups showed no significant difference indicating that thetime spending ofMICD rats to find the objectwas not affectedby themotor functionHowever themechanisms of cognitivetreatment involved are far from being fully elucidated
Firstly the cognitive deficit-related brain regions weredetermined by a small animal MRI in rats with ischemicstroke T2-weighted imaging showed that MICD caused thelesions of left cortex hippocampus corpus striatum andthalamus regions before EA It had beennoticed that the brain
lesion exhibited amild spontaneous recovery performance inMICD rats However the lesion regions comprising approxi-mately 23of thewhole brain volumewere reduced to 15byEA treatment indicating that EA could attenuate cortex andhippocampus corpus striatum and thalamus region lesionsin theMICD rats Some of these regions such as the hippoca-mpus and cortex are essential for regulating learning andmemory behaviors including spatial exploration [36] Studieshave confirmed that the hippocampus regions play a veryimportant role in learning and memory through their spe-cific structure and location which connect with other brainregions together [37] Furthermore in order to identify thehippocampus function in cognitive deficit the hippocampalmorphology staining was observed
Using Golgi staining and electron microscopy we demo-nstrated that the density of dendritic spine and the number ofsynapses in hippocampal CA1 pyramidal cells were obviouslyreduced at Day 14 after MICD However EA can rescue theloss of dendritic spine and synapses in hippocampal CA1region Moreover EA promoted synaptic-dendritic spine
8 Neural Plasticity
MICD+non-EAMICD+EA
MICDSham
(a)
MICD+non-EA
MICD+EAMICDSham0
2
4
6
8
10
The n
umbe
r of s
ynap
ses (10cm
2)
lowastlowast andand
(b)
Figure 5The number of synapses in hippocampal CA1 pyramidal cells (a) Representative electronmicrographic images of synapses (arrows)in the CA1 region of each group (magnification times50000) (b) Histogram shows the significant difference in the mean plusmn SEM of the numberof synapses in each group (119899 = 5group lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD group andand119901 lt 001and119901 lt 005 versus the MICD+EA group)
MICD+non-EAMICD+EAMICD
120573-Actin
P-LIMK1
T-LIMK1
Sham
(a)
MICD+non-EA
MICD+EAMICDSham00
02
04
06
08
10
Tota
l LIM
K1120573
-act
in
lowastlowastandand
(b)
MICD+non-EA
MICD+EAMICDSham00
02
04
06
08
10
lowastlowastandand
P-LI
MK1
T-L
IMK1
(c)
Figure 6 EA amplified the LIMK1 level in the hippocampus CA1 pyramidal cells (a) The expression of total LIMK1 and phosphorylatedLIMK1was evaluated usingwestern blotting (b and c)Histogram shows significant difference of the levels of P-LIMK1 and phosphor-LIMLK1in each group Data are means plusmn SEM (119899 = 6group lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD groupandand119901 lt 001 and119901 lt 005 versus the MICD+EA group)
Neural Plasticity 9
0
2
4
6
8
10
12
miR
-134
relat
ive e
xpre
ssio
n to
U6 lowastlowast
and
MICD+non-EA
MICD+EAMICDSham
Figure 7 Changes of miR-134 expression in the hippocampusCA1 cells The changes of miR-134 expression level were measuredusing quantitative RT-PCR U6 was used as internal control forquantification of miRNAs Data are mean plusmn SEM (119899 = 6grouplowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus theMICD group andand119901 lt 001 and119901 lt 005 versus the MICD+EA group)
regeneration Dendritic spine is the tiny protrusions withmulti-dense body and ion channels on the surface of varioustypes of neurons which serve as cellular substrates of brainconnectivity and the major sites of information processingin the brain [38 39] Evidence is increasing that synaptic-dendritic plasticity enables the encoding of memory [40]Indeed themodification on dendritic spine number size andshape is of importance for the plasticity of synapses accom-panied by hippocampus-dependent learning and memory[41 42] Studies have well described that dendritic spineremodeling in ischemia surrounding areas may contribute tocognitive functional recovery after stroke [43 44]
Increasing evidence showed that LIMK1 protein is acti-vated at dendritic spine membranes [45] LIMK1 positivecells were located in the CA1 region of the hippocampuswhich is important in the regulation of the spinemorphologyand synaptic function in vivo A prominent form of longlasting synaptic plasticity is thought to be critical to memoryformation [26] The present study found that the expressionof total LIMK1 and phospho-LIMK1 in hippocampal CA1wassignificantly decreased in the MICD rats however EA couldenhance the total LIMK1 and phospho-LIMK1 levels to pro-mote synaptic-dendritic plasticity in hippocampal CA1 area
In addition we have identified a dendritically localizedmiRNA that regulated the expression of the synaptic LIMK1protein thereby controlling dendritic pine sizemiR-134 is thefirst discovered dendritic microRNA enriched in the neu-ronal dendrites of rat hippocampal neurons and negativelycontrols the size of dendritic spines [46] This effect ismediated by miR-134 inhibition of the translation of LIMK1mRNA [47] The present study showed that the expression ofmiR-134 was obviously increased in theMICD rats Howeverrepeated EA treatment could relieve the upregulation of miR-134 expression in hippocampal CA1 area
In conclusion the study indicated that EA at the DU20and DU24 acupoints could ameliorate cognitive impairmentof MICD rats through the regulation of synaptic-dendritic
plasticity of the hippocampal CA1 area and the expressionand phosphorylation of synaptic LIMK1 protein were acti-vated to control dendritic spine number size and shapeFurthermore the mechanism of LIMK1 increase induced byEA treatmentmay be associated withmiR-134 which is local-ized in hippocampal CA1 by negatively regulating LIMK1 toenhance synaptic-dendritic plasticity in the recovery stage ofischemic stroke
Competing Interests
The authors declare no financial or commercial conflict ofinterests
Authorsrsquo Contributions
Weilin Liu Jie Wu and Jia Huang contributed equally to thiswork
Acknowledgments
The present study was supported by the Natural ScienceFoundation of China (no 81403450) and the Natural ScienceFoundation of Fujian Province (no 2016J01382)
References
[1] V L Roger ldquoHeart disease and stroke statisticsmdash2011 update areport from the American Heart Associationrdquo Circulation vol131 no 4 pp 29ndash322 2014
[2] T E Nichols and A P Holmes ldquoNonparametric permutationtests for functional neuroimaging a primer with examplesrdquoHuman Brain Mapping vol 15 no 1 pp 1ndash25 2002
[3] T K Tatemichi D W Desmond Y Stern M Paik M Sanoand E Bagiella ldquoCognitive impairment after stroke frequencypatterns and relationship to functional abilitiesrdquo Journal ofNeurology Neurosurgery amp Psychiatry vol 57 no 2 pp 202ndash207 1994
[4] F Liu Z-M Li Y-J Jiang and L-D Chen ldquoA meta-analysis ofacupuncture use in the treatment of cognitive impairment afterstrokerdquo Journal of Alternative amp Complementary Medicine vol20 no 7 pp 535ndash544 2014
[5] W Li R Huang R A Shetty et al ldquoTransient focal cerebralischemia induces long-termcognitive function deficit in an exp-erimental ischemic stroke modelrdquo Neurobiology of Disease vol59 pp 18ndash25 2013
[6] P S Sachdev X Chen H Brodaty CThompson A Altendorfand W Wen ldquoThe determinants and longitudinal course ofpost-stroke mild cognitive impairmentrdquo Journal of the Inter-national Neuropsychological Society vol 15 no 6 pp 915ndash9232009
[7] A-J Liu J-H Li H-Q Li et al ldquoElectroacupuncture foracute ischemic stroke ameta-analysis of randomized controlledtrialsrdquoThe American Journal of Chinese Medicine vol 43 no 8pp 1541ndash1566 2015
[8] X Ding C-Y Li Q-S Wang et al ldquoPatterns in default-modenetwork connectivity for determining outcomes in cognitivefunction in acute stroke patientsrdquo Neuroscience vol 277 no 10pp 637ndash646 2014
10 Neural Plasticity
[9] H Alipanahzadeh M Soleimani S Soleimani Asl B Pourhey-dar A Nikkhah and M Mehdizadeh ldquoTransforming growthfactor-alpha improves memory impairment and neurogenesisfollowing ischemia reperfusionrdquo Cell Journal vol 16 no 3 pp315ndash324 2014
[10] H Eichenbaum ldquoHippocampus cognitive processes and neuralrepresentations that underlie declarative memoryrdquoNeuron vol44 no 1 pp 109ndash120 2004
[11] T Kadar S Dachir B Shukitt-Hale and A Levy ldquoSub-regionalhippocampal vulnerability in various animal models leading tocognitive dysfunctionrdquo Journal of Neural Transmission vol 105no 8-9 pp 987ndash1004 1998
[12] J B Pereira C Valls-Pedret E Ros et al ldquoRegional vulnera-bility of hippocampal subfields to aging measured by structuraland diffusion MRIrdquo Hippocampus vol 24 no 4 pp 403ndash4142014
[13] S Knafo G Ariav E Barkai and F Libersat ldquoOlfactory learn-ing-induced increase in spine density along the apical dendritesof CA1 hippocampal neuronsrdquo Hippocampus vol 14 no 7 pp819ndash825 2004
[14] S Schacher and J-Y Hu ldquoThe less things change the more theyare different contributions of long-term synaptic plasticity andhomeostasis to memoryrdquo Learning and Memory vol 21 no 3pp 128ndash134 2014
[15] K Park H Heo M E Han et al ldquoLearning-induced synapticpotentiation in implanted neural precursor cell-derived neu-ronsrdquo Scientific Reports vol 5 article 17796 2015
[16] S Knafo F Libersat and E Barkai ldquoDynamics of learning-induced spine redistribution along dendrites of pyramidalneurons in ratsrdquo European Journal of Neuroscience vol 21 no4 pp 927ndash935 2005
[17] V C Foletta N Moussi P D Sarmiere J R Bamburg and OBernard ldquoLIM kinase 1 a key regulator of actin dynamics iswidely expressed in embryonic and adult tissuesrdquo ExperimentalCell Research vol 294 no 2 pp 392ndash405 2004
[18] M Endo K Ohashi Y Sasaki et al ldquoControl of growth conemotility andmorphology by LIMkinase and slingshot via phos-phorylation and dephosphorylation of cofilinrdquo Journal of Neu-roscience vol 23 no 7 pp 2527ndash2537 2003
[19] C Jarrold A D Baddeley and C Phillips ldquoLong-termmemoryfor verbal and visual information in Down syndrome andWilliams syndrome performance on the doors and people testrdquoCortex vol 43 no 2 pp 233ndash247 2007
[20] Y Meng Y Zhang V Tregoubov et al ldquoAbnormal spinemorphology and enhanced LTP in LIMK-1 knockout micerdquoNeuron vol 35 no 1 pp 121ndash133 2002
[21] Z Todorovski S Asrar J Liu et al ldquoLIMK1 regulates long-termmemory and synaptic plasticity via the transcriptional factorCREBrdquo Molecular and Cellular Biology vol 35 no 8 pp 1316ndash1328 2015
[22] G M Schratt F Tuebing E A Nigh et al ldquoA brain-specificmicroRNA regulates dendritic spine developmentrdquo Nature vol439 no 7074 pp 283ndash289 2006
[23] F Capitano J Camon V Ferretti et al ldquomicroRNAs modulatespatial memory in the hippocampus and in the ventral striatumin a region-specific mannerrdquo Molecular Neurobiology vol 53no 7 pp 4618ndash4630 2015
[24] J GaoW-YWang Y-WMao et al ldquoA novel pathway regulatesmemory and plasticity via SIRT1 andmiR-134rdquoNature vol 466no 7310 pp 1105ndash1109 2010
[25] J Remenyi M W M van den Bosch O Palygin et al ldquomiR-132212 knockout mice reveal roles for these miRNAs in regu-lating cortical synaptic transmission and plasticityrdquo PLoS ONEvol 8 no 4 Article ID e62509 p 65 2013
[26] L An J LiuWW Li et al ldquoDistribution of LIM domain kinase1 in the olfactory bulb cerebral cortex hippocampus and cere-bellum of the AppPS+minus micerdquo Genetics amp Molecular Researchvol 14 no 4 pp 17244ndash17251 2015
[27] R Lin Y Wu J Tao et al ldquoElectroacupuncture improves cog-nitive function through Rho GTPases and enhances dendriticspine plasticity in rats with cerebral ischemia-reperfusionrdquoMolecular Medicine Reports vol 13 no 3 pp 2655ndash2660 2016
[28] P Castaneda M Munoz G Garcıa-Rojo et al ldquoAssociation ofN-cadherin levels and downstream effectors of Rho GTPaseswith dendritic spine loss induced by chronic stress in rathippocampal neuronsrdquo Journal of Neuroscience Research vol93 no 10 pp 1476ndash1491 2015
[29] A Romarowski M A Battistone F A La Spina et al ldquoPKA-dependent phosphorylation of LIMK1 and Cofilin is essentialformouse sperm acrosomal exocytosisrdquoDevelopmental Biologyvol 405 no 2 pp 237ndash249 2015
[30] R Lin K Yu X Li et al ldquoElectroacupuncture ameliorates post-stroke learning and memory through minimizing ultrastruc-tural brain damage and inhibiting the expression of MMP-2andMMP-9 in cerebral ischemia-reperfusion injured ratsrdquoMol-ecular Medicine Reports vol 14 no 1 pp 225ndash233 2016
[31] X Feng S Yang J Liu et al ldquoElectroacupuncture amelioratescognitive impairment through inhibition of NF-120581B-mediatedneuronal cell apoptosis in cerebral ischemia-reperfusion inj-ured ratsrdquo Molecular Medicine Reports vol 7 no 5 pp 1516ndash1522 2013
[32] Z K Wang N I Guang-Xia K Liu et al ldquoResearch on thechanges of IL-1 receptor and TNF-120572 receptor in rats with cere-bral ischemia reperfusion and the chronergy of acupunctureinterventionrdquo Zhongguo Zhen Jiu vol 32 no 11 pp 1012ndash10182012
[33] X S LiuM Chopp R L Zhang and Z G Zhang ldquoMicroRNAsin cerebral ischemia-induced neurogenesisrdquo Journal of Neu-ropathology amp Experimental Neurology vol 72 no 8 pp 718ndash722 2013
[34] X He T Yan R Chen and D Ran ldquoAcute effects of electro-acupuncture (EA) on hippocampal long term potentiation(LTP) of perforant path-dentate gyrus granule cells synapserelated to memoryrdquo Acupuncture and Electro-TherapeuticsResearch vol 37 no 2-3 pp 89ndash101 2012
[35] W Dong W Guo X Zheng et al ldquoElectroacupunctureimproves cognitive deficits associated with AMPK activation inSAMP8 micerdquo Metabolic Brain Disease vol 30 no 3 pp 777ndash784 2015
[36] B Milner and D Klein ldquoLoss of recent memory after bilateralhippocampal lesions memory and memoriesmdashlooking backand looking forwardrdquo Journal of Neurology Neurosurgery ampPsychiatry vol 87 no 3 article 230 2016
[37] A Veronica Witte L Kerti D S Margulies and A FloelldquoEffects of resveratrol on memory performance hippocampalfunctional connectivity and glucose metabolism in healthyolder adultsrdquo Journal of Neuroscience vol 34 no 23 pp 7862ndash7870 2014
[38] T Bonhoeffer and R Yuste ldquoSpine motility phenomenologymechanisms and functionrdquoNeuron vol 35 no 6 pp 1019ndash10272002
Neural Plasticity 11
[39] H Qiao M Li C Xu H Chen S An and X Ma ldquoDendriticspines in depression what we learned from animal modelsrdquoNeural Plasticity vol 2016 Article ID 8056370 26 pages 2016
[40] S C McQuown R M Barrett D P Matheos et al ldquoHDAC3 isa critical negative regulator of long-term memory formationrdquoJournal of Neuroscience vol 31 no 2 pp 764ndash774 2011
[41] K-O Lai and N Y Ip ldquoStructural plasticity of dendritic spinesthe underlying mechanisms and its dysregulation in brain dis-ordersrdquo Biochimica et Biophysica ActamdashMolecular Basis of Dis-ease vol 1832 no 12 pp 2257ndash2263 2013
[42] C H Bailey E R Kandel and K M Harris ldquoStructural com-ponents of synaptic plasticity and memory consolidationrdquo ColdSpring Harbor Perspectives in Biology vol 7 no 7 Article IDa021758 2015
[43] C E Brown P Li J D Boyd K R Delaney and T HMurphy ldquoExtensive turnover of dendritic spines and vascularremodeling in cortical tissues recovering from strokerdquo Journalof Neuroscience vol 27 no 15 pp 4101ndash4109 2007
[44] R Mostany T G Chowdhury D G Johnston S A PortonovoS T Carmichael and C Portera-Cailliau ldquoLocal hemodynam-ics dictate long-term dendritic plasticity in peri-infarct cortexrdquoJournal of Neuroscience vol 30 no 42 pp 14116ndash14126 2010
[45] J George C Soares A Montersino J-C Beique and G MThomas ldquoPalmitoylation of LIM kinase-1 ensures spine-specificactin polymerization and morphological plasticityrdquo eLife vol2015 no 4 Article ID e06327 2015
[46] S Bicker M Lackinger K Weiszlig and G Schratt ldquoMicroRNA-132 -134 and -138 a microRNA troika rules in neuronaldendritesrdquo Cellular and molecular life sciences CMLS vol 71no 20 pp 3987ndash4005 2014
[47] H-C Tai and E M Schuman ldquoMicroRNA microRNAs reachout into dendritesrdquoCurrent Biology vol 16 no 4 pp R121ndashR1232006
Submit your manuscripts athttpswwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

Neural Plasticity 3
0
Sham or MCAO surgery Sacrificed
Morris water test
Electroacupuncture
Baseline T2WI T2WI
24 h 14 d10 d7 d1 d
Figure 1 Experimental design used in the study
rats arrived at the platform within the 90 sec time restrictionand remained on it for 3 sec they were considered to havefound the platform and were scored by the time takenlengthof the route When the rats were unable to find the platformwithin 90 sec they were placed on the platform for 10 sec andthe time scorewas 90 secThe time taken and the length of theroute bywhich each rat found the safe platformwere recordedby the computerThe average of the time taken and the lengthof the route for the four quadrants as a result of each rat wereassessed every day The duration of the first set of trials wasperformed on Days 10 to 13 after EA The second part of theexperiment was performed on Day 14 to examine the timein which rats found the location of the platform within the90 sec time restriction which tested their ability to rememberthe position of the platform After all trials the rats weredried thoroughly with a hair drier and returned to their cages(Guangzhou RiboBio Co Ltd Guangzhou China) Morriswater maze test was repeated three times
24 Measurement of Brain Lesions Animal MRI scans wereperformed on a 70 T MRI scanner (7020USR BioSpecBruker Biospin Gmbls Germany) using a 38mmbirdcage ratbrain quadrature resonator for radiofrequency transmissionand reception Animals were anesthetized with isofluraneO2(with 3 induction for 5min and 12ndash15 maintenance inorder to let the rats in the depth of anesthesia state) andkept warm with circuit After anesthesia the rat was putin prone position on a custom-made holder to minimizehead motion set the location of head position and performreal-time monitor of the breathing rate and maintained in40 breathsmin Ratrsquos temperature is maintained at 33 plusmn 2∘Cin the process of scanning holder to minimize head motionwhile respiration was maintained
T2-weighted imaging (T2WI) in three planes with afast spin echo (FSE) pulse sequence was first acquired tocontrol rat head positioning T2WI scan was acquired usinga Rapid Acquisition with Relaxation Enhancement (RARE)pulse sequence with the following parameters field of view =32mmtimes 32mmmatrix size = 256times 256 repetition time (TR)= 4200ms echo time (TE) = 35ms slice thickness = 10mmand slice gap = 0mm
Image J analysis and processing system was applied forT2W images the percentage of the brain lesions = brainlesions volumewhole brain volume times100
25 Golgi Staining Golgi staining of brain was performedusing an FD Rapid GolgiStain Kit following the manufac-turerrsquos instructions (FD Neurotechnologies Inc ColumbiaMD USA) The removed rat brains were placed incubated
in a mixture of A and B solutions from the kit and storedin the dark at room temperature following which they weretransferred into solution C from the kit stored at 4∘C for 7days Finally brains were then frozen and coronal sections(150 120583m) were made using a cryostat (Leica CM3050S LeicaMicrosystems K K Tokyo Japan) The sections incubatedin a mixture of D and E solutions from the kit and thendehydrated in alcohol (50 70 90 and 100 for 5min each)cleared in xylene and were cover-slipped The images finallywere viewed under a microscope (Leica DM6000 B LeicaMicrosystems Wetzlar Germany) The slides were reviewedby two or three pathologists blind to the study
26 Transmission Electron Microscopy Four rats in eachgroup were anesthetized and the left ventricle was perfusedwith 200mL of saline followed by 400mL 4 paraformalde-hyde (pH 74) The tissue was taken from the left ischemichippocampus cut into 1mm3 size cubes and fixed in 1paraformaldehyde with 1 lanthanum nitrate tracer for 24 hfollowed by fixation with 3 glutaraldehyde for 24 h Sampleswere fixed in 1 osmium tetroxide for 2 h and dehydrated ingraded ethanol 1 lanthanum nitrate tracer (LNT) solutionand embedded in araldite Ultrathin hippocampal CA1 sliceswere obtained (90 nm) then they were stained with uranylacetate and lead citrate and observed under TEM (H-7650Hitachi Ltd Tokyo Japan) The photos were obtained onhp digital CCD camera (SIS4 million voxel) Images wereacquired digitally from a randomly selected pool of 10 to 15fields under each condition
27 Western Blotting Isolated left hippocampus tissue waslysed in 100 120583L radioimmunoprecipitation assay (RIPA) buf-fer (Beyotime Haimen Jiangsu China) plus protease inhibi-tors Total protein (50120583g) was loaded into 10 SDS-PAGEgels electrophoresed and then transblotted onto polyvinyli-dene difluoride membranes (Immobilon-P Millipore Bil-lerica MA) in a Tris-glycine transfer buffer After beingblocked in 5 milk in PBST for 1 h at room temperaturewith shaking membranes were incubated with antibodiesovernight at 4∘C against the following anti-LIMK1 antibody(dilution 1 1000 ab81046 Abcam) and anti-LIM kinase1 antibody (phospho-Thr508) (dilution 1 1000 ab131341Abcam) and 120573-actin (dilution 1 1000 ab189073 Abcam)The following day membranes were incubated in 5 milk(in TBST) with an anti-goat or anti-mouse IgG antibody(dilution 1 5000 PerkinElmer Life Sciences Waltham MA)for 1 h at room temperature with shaking Membranes werewashed a minimum of four times (10min per wash) in PBST
4 Neural Plasticity
between each antibody treatment Detected bands were visu-alized using enhanced chemiluminescence and images werecaptured using a Bio-Image system (Bio-Rad LaboratoriesInc Hercules USA) Western blotting was repeated threetimes
28 Real Time Quantitative RT-PCR The expression of miR-NAs was determined by real time quantitative reverse tran-scription polymerase chain reaction (RT-PCR) Total RNAwas extracted from hippocampal region of the ipsilaterallesion tissue using the TRIzol Reagent (Life Technologies(AB amp Invitrogen) Carlsbad USA) Then extracted totalRNA was reverse transcribed to generate cDNA accord-ing to manufacturer instructions of Revert AidTM First-Strand cDNA Synthesis Kit (Thermo Fisher Scientific Bei-jing China) The reverse transcription reaction was ampli-fied using a Bio-Rad CFX96 Detection System (Bio-RadHercules CA USA) with the Plexor One-Step qRT-PCR System (Promega Madison WI USA) For miRNAamplification the following primers were used rno-miR-134 (RmiRQP0168 GeneCopoeia Guangzhou China) Thefold change in relative mRNA and miRNA expression wasdetermined using the 2minusΔΔCt method as described previously[33] and U6 snRNA (RQP047936 GeneCopoeia) as mRNAand miRNA internal control respectively
29 Statistical Analysis Statistical analysis was performedwith the SPSS package for Windows statistical analysissoftware (Version 180 SPSS Inc Chicago IL USA) Thedata from all groups were determined by one-way analysisof variance (ANOVA) and Studentrsquos 119905-testsThe homogeneityof variance was analyzed using the least significant differ-ence method and missing variance using the Games-Howellmethod Intergroup comparisons of the brain lesions volumeat different time points were performed with paired-samples119905-tests All data are presented as mean plusmn standard error andthe significance was regarded as at least 119901 lt 005 All finalresults were analyzed in a blinded manner
3 Results
31 EA Improved Cognitive Impairment in MICD Rats Toassess the effect of EA on spatial reference learning andmemory impairments in MICD rats the Morris water maze(MWM) test was performed All groups of rats learned to findthe platform and the latency time to reach the platform wasreduced in the four training days However learning abilitywas significantly reduced in MICD rats compared with theSham group (119901 lt 001 Figure 2(a)) the MICD rats treatedwith EA significantly took less time to find the platform com-pared with the MICD group and the MICD+non-EA group(119901 lt 001 or 119901 lt 005 Figure 2(a)) There was no significantdifference about path length among the four groups (119901 gt005 Figure 2(b)) As illustrated in Figures 2(c) and 2(d)tracing images from the MWM test showed that in the spaceexploration test where the platform was removed the MICDrats passed through the original position of the platformfewer times than the Sham group (119901 lt 001 Figure 2(d))
whereas in the MICD+EA group the times where the ratscrossed the position of the platformwere significantly increa-sed compared with the MICD group and the MICD+non-EA group (119901 lt 001 119901 lt 005 Figure 2(d)) As illustratedin Figures 2(e) and 2(f) the percentage of time spent in thetarget quadrant was used for statistical analysis during theprobe trial The date showed that the rats spent more time inthe target quadrant compared to other quadrants (119901 lt 001Figure 2(e)) The MICD group spent less time in the targetquadrant compared with the other groups (119901 lt 001 Fig-ure 2(e))TheMICD+EA group spent more time in the targetquadrant compared with the MICD group and MICD+non-EA group (119901 lt 001 119901 lt 005 Figure 2(f)) Therefore theseresults suggest that acquisition or retention of spatial refer-ence learning and memory was ameliorated in MICD rats byEA treatment
32 EA Attenuated Left Cortex Hippocampus Corpus Stria-tum and Thalamus Lesions in MICD Rats The brain lesionswere determined by T2-weighted imaging (T2WI) (Figures3(a) and 3(c)) before and after EA treatment The volumes ofbrain lesions that included left cortex hippocampus corpusstriatum and thalamus regions in the all groups had nosignificant difference before EA treatment (119901 gt 005 Figures3(a) and 3(b)) There was mild spontaneous recovery for 14days in the brain lesions of the MICD group The volumesof left cortex hippocampus corpus striatum and thalamuslesions showed comprising approximately 23 of the wholebrain in theMICDgroup whereas the brain lesions sizes weresignificantly reduced to 15 in the MICD+EA group (119901 lt001 Figures 3(c) and 3(d)) The difference of brain lesionsbetween the MICD+EA and the MICD+non-EA groupshowed significant changes at Day 14 after EA (119901 lt 005Figures 3(c) and 3(d))
33 EA Increased the Density of Dendritic Spines in the Hip-pocampus ofMICDRats To investigate the function of EAonsynaptic plasticity dendritic spine density in the hippocampalneurons was analyzed by the primary basilar dendrites ofGolgi-stained pyramidal neurons at Days 14 after EA Golgi-Cox staining clearly filled the dendritic shafts and the spinesof neurons frompyramidal neurons (Figure 4(a))Hereby therepresentative hippocampus showed that the density of den-dritic spines was reduced in different degree by macroscopicexamination in the MICD group the MICD+EA group andMICD+non-EA group and the loss of dendritic spines inhippocampal CA1 was obvious in the MICD group Thus asillustrated in Figures 4(b) and 4(c) the density of selecteddendritic spines that derived from hippocampal CA1 wassignificantly decreased in the MICD group compared withthe Sham group (119901 lt 001) however the density of dendriticspines of the hippocampal CA1 in the MICD+EA group wasmore than that of the MICD group and the MICD+non-EAgroup (119901 lt 001) In brief EA treatment triggered large-scale remodeling of dendrites in the hippocampal area CA1
34 EA Enhanced the Number of Hippocampus CA1 SynapsesinMICD Rats Furthermore we observed the effect of EA on
Neural Plasticity 5
Esca
pe la
tenc
y (s
)
Day 10 Day 11 Day 12 Day 13Days after MICD
ShamMICD
MICD+EAMICD+non-EA
lowastlowast
lowastlowast
lowastlowastlowastlowast
and
and
andand
0
10
20
30
40
50
60
70
80
90
100
(a)
Day 10 Day 11 Day 12 Day 13Training time
ShamMICD
MICD+EAMICD+non-EA
0
200
400
600
800
1000
1200
Dist
ance
trav
elle
d (c
m)
(b)
Sham MICD
MICD+EA MICD+non-EA
III
III IV
III
III IV
III
III IV
III
III IV
(c)
Sham MICD MICD+EA MICD+non-EA
lowastlowast
and
Prob
e tim
e
0
1
2
3
4
5
6
7
(d)
ShamMICD
MICD+EAMICD+non-EA
lowastlowast and
ti
me s
pent
in ea
ch q
uadr
ant
0
20
40
60
Opp
osite
Left
adja
cent
Targ
etRi
ght a
djac
ent
Opp
osite
Left
adja
cent
Targ
etRi
ght a
djac
ent
Opp
osite
Left
adja
cent
Targ
etRi
ght a
djac
ent
Opp
osite
Left
adja
cent
Targ
etRi
ght a
djac
ent
(e)
Sham MICD MICD+EA MICD+non-EA
lowastlowast
and
ti
me s
pent
in ta
rget
qua
dran
t
0
5
10
15
20
25
30
35
40
(f)
Figure 2 Evaluation of spatial reference learning and memory by Morris water maze test at Days 10ndash14 after EA (a and b) Mean escapelatency time and path length during the orientation navigation test on Days 10ndash13 after EA treatment (c and d) Times the rats crossed overprevious platform location on Day 14 after EA treatment during the spatial memory test in different groups (e)The percentage of times spentin each quadrant in all probe trials is shown on Day 14 after EA treatment (f) The times of passing the hidden platform position on Day 14after EA treatment (each group lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD group andand119901 lt 001 and119901 lt 005versus the MICD+EA group) EA electroacupuncture MICD middle cerebral artery occlusion induced cognitive deficits
6 Neural Plasticity
MICD+non-EA
MICD+EA
MICD
Sham
(a)
MICD+non-EA
MICD+EAMICD
Lesio
ns v
olum
es (
)
0
10
20
30
40
(b)
MICD+non-EA
MICD+EA
MICD
Sham
(c)
MICD+non-EA
MICD+EAMICD
Lesio
ns v
olum
es (
)
0
10
20
30
and
(d)
Figure 3 T2WI signal changes before and after EA (a and c)Thebrain lesionsweremeasured byT2WI (slices from 11 to 13) in the Shamgroupthe MICD group the MICD+EA group and the MICD+non-EA group before (a) and after (c) EA (b and d) Lesion volume is representedas a percentage of the total brain volume and data are presented as the mean plusmn standard deviation from 12 individual rats in each group(lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD group andand119901 lt 001 and119901 lt 005 versus the MICD+EA group)T2WI T2-weighted magnetic resonance imaging MICD middle cerebral artery occlusion induced cognitive deficits
ultrastructural morphology of hippocampal CA1 pyramidalneurons Obviously the density of synapses in the MICDgroup was decreased compared with the Sham group (119901 lt001 Figures 5(a) and 5(b)) whereas there was amplifying inthe number of synapses in the MICD+EA group comparedwith the MICD group and the MICD+non-EA group (119901 lt005 or 119901 lt 001 Figures 5(a) and 5(b))
Taken together these results suggest that EA couldimprove synaptic-dendritic plasticity in vivo
35 EA Promoted LIMK1 Expression and Phosphorylation ofHippocampus CA1 in MICD Rats To explore the underly-ing molecular mechanism of EA-induced synaptic-dendriticplasticity the levels of total LIMK1 and phospho-LIMK1 (P-LIMK1 Thr508) in hippocampal CA1 were investigated Asshown in Figures 6(a) and 6(b) the expression of total LIMK1of hippocampal CA1was significantly decreased in theMICDgroup compared with the Sham group (119901 lt 001) Howeverthe total LIMK1 level in the MICD+EA group was morethan that of the MICD group and the MICD+non-EA group(119901 lt 001 Figures 6(a) and 6(b)) Moreover the changes ofphospho-LIMK1 (Thr508) in groups were similar to the totalLIMK1 expression The level of P-LIMK1 was significantly
increased in the MICD+EA group compared with the MICDgroup and the MICD+non-EA group (119901 lt 001 Figures 6(a)and 6(c))
36 EA Regulated miR-134 Expression in Hippocampal CA1 inMICD Rats To identify the role of synaptic-dendrite-relatedmiR-134 in hippocampus CA1 the levels were detected Asshown in Figure 7 the expression ofmiR-134was significantlyincreased in theMICD group comparedwith the Sham group(119901 lt 001) However repeated EA treatment significantlydecreased the expression of miR-134 compared with theMICD group and the MICD+non-EA group (119901 lt 005)
4 Discussion
Ischemic stroke leads to a high incidence of long-termcognitive impairment which is strongly associated with lossof hippocampal neurons and synapses [34] While review-ing ancient Chinese documents regarding acupuncture andcognitive impairment-related terms we discovered that theDU20 andDU24 acupoints were themost frequently selectedacupoints for cognitive impairment-related rehabilitation inChina A number of studies have shown that EA can improve
Neural Plasticity 7
MICD+non-EAMICD+EA
MICDSham
(a)
MICD+non-EA
MICD+EA
MICD
Sham
(b)
MICD+non-EA
MICD+EAMICDSham0
5
10
15
20
lowastlowastandand
Spin
e den
sity
(10
um)
(c)
Figure 4 The structures and densities of dendritic spine in the hippocampus of rats (a) Representative images of dendritic spine in thehippocampus of each group (Golgi staining times50) (b) Representative images of dendritic spine density and morphology from the pyramidalcell layer of hippocampal CA1 area in each group (Golgi staining times1000) (c)The density of dendritic spine was analyzed in hippocampal CA1pyramidal cells in each group (119899 = 6group lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD group andand119901 lt 001and119901 lt 005 versus the MICD+EA group) All experiments were repeated three times
learning and memory ability and stimulate consciousness[30 31 35]These results indicated that EA could be a comple-mentary therapy for cognitive impairment after stroke Thepresent study found that EA at theDU20 andDU24 acupointsshortened time to find the platform and increased the timesof crossing the position of the platform compared to untreat-ment or nonacupoint EA (stimulation control) in Morriswater maze test suggesting that EA could improve spatialreference learning and memory ability in MCAO-inducedcognitive deficit (MICD) rats Moreover it is worth mention-ing that the path length in the Morris water maze in the fourgroups showed no significant difference indicating that thetime spending ofMICD rats to find the objectwas not affectedby themotor functionHowever themechanisms of cognitivetreatment involved are far from being fully elucidated
Firstly the cognitive deficit-related brain regions weredetermined by a small animal MRI in rats with ischemicstroke T2-weighted imaging showed that MICD caused thelesions of left cortex hippocampus corpus striatum andthalamus regions before EA It had beennoticed that the brain
lesion exhibited amild spontaneous recovery performance inMICD rats However the lesion regions comprising approxi-mately 23of thewhole brain volumewere reduced to 15byEA treatment indicating that EA could attenuate cortex andhippocampus corpus striatum and thalamus region lesionsin theMICD rats Some of these regions such as the hippoca-mpus and cortex are essential for regulating learning andmemory behaviors including spatial exploration [36] Studieshave confirmed that the hippocampus regions play a veryimportant role in learning and memory through their spe-cific structure and location which connect with other brainregions together [37] Furthermore in order to identify thehippocampus function in cognitive deficit the hippocampalmorphology staining was observed
Using Golgi staining and electron microscopy we demo-nstrated that the density of dendritic spine and the number ofsynapses in hippocampal CA1 pyramidal cells were obviouslyreduced at Day 14 after MICD However EA can rescue theloss of dendritic spine and synapses in hippocampal CA1region Moreover EA promoted synaptic-dendritic spine
8 Neural Plasticity
MICD+non-EAMICD+EA
MICDSham
(a)
MICD+non-EA
MICD+EAMICDSham0
2
4
6
8
10
The n
umbe
r of s
ynap
ses (10cm
2)
lowastlowast andand
(b)
Figure 5The number of synapses in hippocampal CA1 pyramidal cells (a) Representative electronmicrographic images of synapses (arrows)in the CA1 region of each group (magnification times50000) (b) Histogram shows the significant difference in the mean plusmn SEM of the numberof synapses in each group (119899 = 5group lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD group andand119901 lt 001and119901 lt 005 versus the MICD+EA group)
MICD+non-EAMICD+EAMICD
120573-Actin
P-LIMK1
T-LIMK1
Sham
(a)
MICD+non-EA
MICD+EAMICDSham00
02
04
06
08
10
Tota
l LIM
K1120573
-act
in
lowastlowastandand
(b)
MICD+non-EA
MICD+EAMICDSham00
02
04
06
08
10
lowastlowastandand
P-LI
MK1
T-L
IMK1
(c)
Figure 6 EA amplified the LIMK1 level in the hippocampus CA1 pyramidal cells (a) The expression of total LIMK1 and phosphorylatedLIMK1was evaluated usingwestern blotting (b and c)Histogram shows significant difference of the levels of P-LIMK1 and phosphor-LIMLK1in each group Data are means plusmn SEM (119899 = 6group lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD groupandand119901 lt 001 and119901 lt 005 versus the MICD+EA group)
Neural Plasticity 9
0
2
4
6
8
10
12
miR
-134
relat
ive e
xpre
ssio
n to
U6 lowastlowast
and
MICD+non-EA
MICD+EAMICDSham
Figure 7 Changes of miR-134 expression in the hippocampusCA1 cells The changes of miR-134 expression level were measuredusing quantitative RT-PCR U6 was used as internal control forquantification of miRNAs Data are mean plusmn SEM (119899 = 6grouplowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus theMICD group andand119901 lt 001 and119901 lt 005 versus the MICD+EA group)
regeneration Dendritic spine is the tiny protrusions withmulti-dense body and ion channels on the surface of varioustypes of neurons which serve as cellular substrates of brainconnectivity and the major sites of information processingin the brain [38 39] Evidence is increasing that synaptic-dendritic plasticity enables the encoding of memory [40]Indeed themodification on dendritic spine number size andshape is of importance for the plasticity of synapses accom-panied by hippocampus-dependent learning and memory[41 42] Studies have well described that dendritic spineremodeling in ischemia surrounding areas may contribute tocognitive functional recovery after stroke [43 44]
Increasing evidence showed that LIMK1 protein is acti-vated at dendritic spine membranes [45] LIMK1 positivecells were located in the CA1 region of the hippocampuswhich is important in the regulation of the spinemorphologyand synaptic function in vivo A prominent form of longlasting synaptic plasticity is thought to be critical to memoryformation [26] The present study found that the expressionof total LIMK1 and phospho-LIMK1 in hippocampal CA1wassignificantly decreased in the MICD rats however EA couldenhance the total LIMK1 and phospho-LIMK1 levels to pro-mote synaptic-dendritic plasticity in hippocampal CA1 area
In addition we have identified a dendritically localizedmiRNA that regulated the expression of the synaptic LIMK1protein thereby controlling dendritic pine sizemiR-134 is thefirst discovered dendritic microRNA enriched in the neu-ronal dendrites of rat hippocampal neurons and negativelycontrols the size of dendritic spines [46] This effect ismediated by miR-134 inhibition of the translation of LIMK1mRNA [47] The present study showed that the expression ofmiR-134 was obviously increased in theMICD rats Howeverrepeated EA treatment could relieve the upregulation of miR-134 expression in hippocampal CA1 area
In conclusion the study indicated that EA at the DU20and DU24 acupoints could ameliorate cognitive impairmentof MICD rats through the regulation of synaptic-dendritic
plasticity of the hippocampal CA1 area and the expressionand phosphorylation of synaptic LIMK1 protein were acti-vated to control dendritic spine number size and shapeFurthermore the mechanism of LIMK1 increase induced byEA treatmentmay be associated withmiR-134 which is local-ized in hippocampal CA1 by negatively regulating LIMK1 toenhance synaptic-dendritic plasticity in the recovery stage ofischemic stroke
Competing Interests
The authors declare no financial or commercial conflict ofinterests
Authorsrsquo Contributions
Weilin Liu Jie Wu and Jia Huang contributed equally to thiswork
Acknowledgments
The present study was supported by the Natural ScienceFoundation of China (no 81403450) and the Natural ScienceFoundation of Fujian Province (no 2016J01382)
References
[1] V L Roger ldquoHeart disease and stroke statisticsmdash2011 update areport from the American Heart Associationrdquo Circulation vol131 no 4 pp 29ndash322 2014
[2] T E Nichols and A P Holmes ldquoNonparametric permutationtests for functional neuroimaging a primer with examplesrdquoHuman Brain Mapping vol 15 no 1 pp 1ndash25 2002
[3] T K Tatemichi D W Desmond Y Stern M Paik M Sanoand E Bagiella ldquoCognitive impairment after stroke frequencypatterns and relationship to functional abilitiesrdquo Journal ofNeurology Neurosurgery amp Psychiatry vol 57 no 2 pp 202ndash207 1994
[4] F Liu Z-M Li Y-J Jiang and L-D Chen ldquoA meta-analysis ofacupuncture use in the treatment of cognitive impairment afterstrokerdquo Journal of Alternative amp Complementary Medicine vol20 no 7 pp 535ndash544 2014
[5] W Li R Huang R A Shetty et al ldquoTransient focal cerebralischemia induces long-termcognitive function deficit in an exp-erimental ischemic stroke modelrdquo Neurobiology of Disease vol59 pp 18ndash25 2013
[6] P S Sachdev X Chen H Brodaty CThompson A Altendorfand W Wen ldquoThe determinants and longitudinal course ofpost-stroke mild cognitive impairmentrdquo Journal of the Inter-national Neuropsychological Society vol 15 no 6 pp 915ndash9232009
[7] A-J Liu J-H Li H-Q Li et al ldquoElectroacupuncture foracute ischemic stroke ameta-analysis of randomized controlledtrialsrdquoThe American Journal of Chinese Medicine vol 43 no 8pp 1541ndash1566 2015
[8] X Ding C-Y Li Q-S Wang et al ldquoPatterns in default-modenetwork connectivity for determining outcomes in cognitivefunction in acute stroke patientsrdquo Neuroscience vol 277 no 10pp 637ndash646 2014
10 Neural Plasticity
[9] H Alipanahzadeh M Soleimani S Soleimani Asl B Pourhey-dar A Nikkhah and M Mehdizadeh ldquoTransforming growthfactor-alpha improves memory impairment and neurogenesisfollowing ischemia reperfusionrdquo Cell Journal vol 16 no 3 pp315ndash324 2014
[10] H Eichenbaum ldquoHippocampus cognitive processes and neuralrepresentations that underlie declarative memoryrdquoNeuron vol44 no 1 pp 109ndash120 2004
[11] T Kadar S Dachir B Shukitt-Hale and A Levy ldquoSub-regionalhippocampal vulnerability in various animal models leading tocognitive dysfunctionrdquo Journal of Neural Transmission vol 105no 8-9 pp 987ndash1004 1998
[12] J B Pereira C Valls-Pedret E Ros et al ldquoRegional vulnera-bility of hippocampal subfields to aging measured by structuraland diffusion MRIrdquo Hippocampus vol 24 no 4 pp 403ndash4142014
[13] S Knafo G Ariav E Barkai and F Libersat ldquoOlfactory learn-ing-induced increase in spine density along the apical dendritesof CA1 hippocampal neuronsrdquo Hippocampus vol 14 no 7 pp819ndash825 2004
[14] S Schacher and J-Y Hu ldquoThe less things change the more theyare different contributions of long-term synaptic plasticity andhomeostasis to memoryrdquo Learning and Memory vol 21 no 3pp 128ndash134 2014
[15] K Park H Heo M E Han et al ldquoLearning-induced synapticpotentiation in implanted neural precursor cell-derived neu-ronsrdquo Scientific Reports vol 5 article 17796 2015
[16] S Knafo F Libersat and E Barkai ldquoDynamics of learning-induced spine redistribution along dendrites of pyramidalneurons in ratsrdquo European Journal of Neuroscience vol 21 no4 pp 927ndash935 2005
[17] V C Foletta N Moussi P D Sarmiere J R Bamburg and OBernard ldquoLIM kinase 1 a key regulator of actin dynamics iswidely expressed in embryonic and adult tissuesrdquo ExperimentalCell Research vol 294 no 2 pp 392ndash405 2004
[18] M Endo K Ohashi Y Sasaki et al ldquoControl of growth conemotility andmorphology by LIMkinase and slingshot via phos-phorylation and dephosphorylation of cofilinrdquo Journal of Neu-roscience vol 23 no 7 pp 2527ndash2537 2003
[19] C Jarrold A D Baddeley and C Phillips ldquoLong-termmemoryfor verbal and visual information in Down syndrome andWilliams syndrome performance on the doors and people testrdquoCortex vol 43 no 2 pp 233ndash247 2007
[20] Y Meng Y Zhang V Tregoubov et al ldquoAbnormal spinemorphology and enhanced LTP in LIMK-1 knockout micerdquoNeuron vol 35 no 1 pp 121ndash133 2002
[21] Z Todorovski S Asrar J Liu et al ldquoLIMK1 regulates long-termmemory and synaptic plasticity via the transcriptional factorCREBrdquo Molecular and Cellular Biology vol 35 no 8 pp 1316ndash1328 2015
[22] G M Schratt F Tuebing E A Nigh et al ldquoA brain-specificmicroRNA regulates dendritic spine developmentrdquo Nature vol439 no 7074 pp 283ndash289 2006
[23] F Capitano J Camon V Ferretti et al ldquomicroRNAs modulatespatial memory in the hippocampus and in the ventral striatumin a region-specific mannerrdquo Molecular Neurobiology vol 53no 7 pp 4618ndash4630 2015
[24] J GaoW-YWang Y-WMao et al ldquoA novel pathway regulatesmemory and plasticity via SIRT1 andmiR-134rdquoNature vol 466no 7310 pp 1105ndash1109 2010
[25] J Remenyi M W M van den Bosch O Palygin et al ldquomiR-132212 knockout mice reveal roles for these miRNAs in regu-lating cortical synaptic transmission and plasticityrdquo PLoS ONEvol 8 no 4 Article ID e62509 p 65 2013
[26] L An J LiuWW Li et al ldquoDistribution of LIM domain kinase1 in the olfactory bulb cerebral cortex hippocampus and cere-bellum of the AppPS+minus micerdquo Genetics amp Molecular Researchvol 14 no 4 pp 17244ndash17251 2015
[27] R Lin Y Wu J Tao et al ldquoElectroacupuncture improves cog-nitive function through Rho GTPases and enhances dendriticspine plasticity in rats with cerebral ischemia-reperfusionrdquoMolecular Medicine Reports vol 13 no 3 pp 2655ndash2660 2016
[28] P Castaneda M Munoz G Garcıa-Rojo et al ldquoAssociation ofN-cadherin levels and downstream effectors of Rho GTPaseswith dendritic spine loss induced by chronic stress in rathippocampal neuronsrdquo Journal of Neuroscience Research vol93 no 10 pp 1476ndash1491 2015
[29] A Romarowski M A Battistone F A La Spina et al ldquoPKA-dependent phosphorylation of LIMK1 and Cofilin is essentialformouse sperm acrosomal exocytosisrdquoDevelopmental Biologyvol 405 no 2 pp 237ndash249 2015
[30] R Lin K Yu X Li et al ldquoElectroacupuncture ameliorates post-stroke learning and memory through minimizing ultrastruc-tural brain damage and inhibiting the expression of MMP-2andMMP-9 in cerebral ischemia-reperfusion injured ratsrdquoMol-ecular Medicine Reports vol 14 no 1 pp 225ndash233 2016
[31] X Feng S Yang J Liu et al ldquoElectroacupuncture amelioratescognitive impairment through inhibition of NF-120581B-mediatedneuronal cell apoptosis in cerebral ischemia-reperfusion inj-ured ratsrdquo Molecular Medicine Reports vol 7 no 5 pp 1516ndash1522 2013
[32] Z K Wang N I Guang-Xia K Liu et al ldquoResearch on thechanges of IL-1 receptor and TNF-120572 receptor in rats with cere-bral ischemia reperfusion and the chronergy of acupunctureinterventionrdquo Zhongguo Zhen Jiu vol 32 no 11 pp 1012ndash10182012
[33] X S LiuM Chopp R L Zhang and Z G Zhang ldquoMicroRNAsin cerebral ischemia-induced neurogenesisrdquo Journal of Neu-ropathology amp Experimental Neurology vol 72 no 8 pp 718ndash722 2013
[34] X He T Yan R Chen and D Ran ldquoAcute effects of electro-acupuncture (EA) on hippocampal long term potentiation(LTP) of perforant path-dentate gyrus granule cells synapserelated to memoryrdquo Acupuncture and Electro-TherapeuticsResearch vol 37 no 2-3 pp 89ndash101 2012
[35] W Dong W Guo X Zheng et al ldquoElectroacupunctureimproves cognitive deficits associated with AMPK activation inSAMP8 micerdquo Metabolic Brain Disease vol 30 no 3 pp 777ndash784 2015
[36] B Milner and D Klein ldquoLoss of recent memory after bilateralhippocampal lesions memory and memoriesmdashlooking backand looking forwardrdquo Journal of Neurology Neurosurgery ampPsychiatry vol 87 no 3 article 230 2016
[37] A Veronica Witte L Kerti D S Margulies and A FloelldquoEffects of resveratrol on memory performance hippocampalfunctional connectivity and glucose metabolism in healthyolder adultsrdquo Journal of Neuroscience vol 34 no 23 pp 7862ndash7870 2014
[38] T Bonhoeffer and R Yuste ldquoSpine motility phenomenologymechanisms and functionrdquoNeuron vol 35 no 6 pp 1019ndash10272002
Neural Plasticity 11
[39] H Qiao M Li C Xu H Chen S An and X Ma ldquoDendriticspines in depression what we learned from animal modelsrdquoNeural Plasticity vol 2016 Article ID 8056370 26 pages 2016
[40] S C McQuown R M Barrett D P Matheos et al ldquoHDAC3 isa critical negative regulator of long-term memory formationrdquoJournal of Neuroscience vol 31 no 2 pp 764ndash774 2011
[41] K-O Lai and N Y Ip ldquoStructural plasticity of dendritic spinesthe underlying mechanisms and its dysregulation in brain dis-ordersrdquo Biochimica et Biophysica ActamdashMolecular Basis of Dis-ease vol 1832 no 12 pp 2257ndash2263 2013
[42] C H Bailey E R Kandel and K M Harris ldquoStructural com-ponents of synaptic plasticity and memory consolidationrdquo ColdSpring Harbor Perspectives in Biology vol 7 no 7 Article IDa021758 2015
[43] C E Brown P Li J D Boyd K R Delaney and T HMurphy ldquoExtensive turnover of dendritic spines and vascularremodeling in cortical tissues recovering from strokerdquo Journalof Neuroscience vol 27 no 15 pp 4101ndash4109 2007
[44] R Mostany T G Chowdhury D G Johnston S A PortonovoS T Carmichael and C Portera-Cailliau ldquoLocal hemodynam-ics dictate long-term dendritic plasticity in peri-infarct cortexrdquoJournal of Neuroscience vol 30 no 42 pp 14116ndash14126 2010
[45] J George C Soares A Montersino J-C Beique and G MThomas ldquoPalmitoylation of LIM kinase-1 ensures spine-specificactin polymerization and morphological plasticityrdquo eLife vol2015 no 4 Article ID e06327 2015
[46] S Bicker M Lackinger K Weiszlig and G Schratt ldquoMicroRNA-132 -134 and -138 a microRNA troika rules in neuronaldendritesrdquo Cellular and molecular life sciences CMLS vol 71no 20 pp 3987ndash4005 2014
[47] H-C Tai and E M Schuman ldquoMicroRNA microRNAs reachout into dendritesrdquoCurrent Biology vol 16 no 4 pp R121ndashR1232006
Submit your manuscripts athttpswwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

4 Neural Plasticity
between each antibody treatment Detected bands were visu-alized using enhanced chemiluminescence and images werecaptured using a Bio-Image system (Bio-Rad LaboratoriesInc Hercules USA) Western blotting was repeated threetimes
28 Real Time Quantitative RT-PCR The expression of miR-NAs was determined by real time quantitative reverse tran-scription polymerase chain reaction (RT-PCR) Total RNAwas extracted from hippocampal region of the ipsilaterallesion tissue using the TRIzol Reagent (Life Technologies(AB amp Invitrogen) Carlsbad USA) Then extracted totalRNA was reverse transcribed to generate cDNA accord-ing to manufacturer instructions of Revert AidTM First-Strand cDNA Synthesis Kit (Thermo Fisher Scientific Bei-jing China) The reverse transcription reaction was ampli-fied using a Bio-Rad CFX96 Detection System (Bio-RadHercules CA USA) with the Plexor One-Step qRT-PCR System (Promega Madison WI USA) For miRNAamplification the following primers were used rno-miR-134 (RmiRQP0168 GeneCopoeia Guangzhou China) Thefold change in relative mRNA and miRNA expression wasdetermined using the 2minusΔΔCt method as described previously[33] and U6 snRNA (RQP047936 GeneCopoeia) as mRNAand miRNA internal control respectively
29 Statistical Analysis Statistical analysis was performedwith the SPSS package for Windows statistical analysissoftware (Version 180 SPSS Inc Chicago IL USA) Thedata from all groups were determined by one-way analysisof variance (ANOVA) and Studentrsquos 119905-testsThe homogeneityof variance was analyzed using the least significant differ-ence method and missing variance using the Games-Howellmethod Intergroup comparisons of the brain lesions volumeat different time points were performed with paired-samples119905-tests All data are presented as mean plusmn standard error andthe significance was regarded as at least 119901 lt 005 All finalresults were analyzed in a blinded manner
3 Results
31 EA Improved Cognitive Impairment in MICD Rats Toassess the effect of EA on spatial reference learning andmemory impairments in MICD rats the Morris water maze(MWM) test was performed All groups of rats learned to findthe platform and the latency time to reach the platform wasreduced in the four training days However learning abilitywas significantly reduced in MICD rats compared with theSham group (119901 lt 001 Figure 2(a)) the MICD rats treatedwith EA significantly took less time to find the platform com-pared with the MICD group and the MICD+non-EA group(119901 lt 001 or 119901 lt 005 Figure 2(a)) There was no significantdifference about path length among the four groups (119901 gt005 Figure 2(b)) As illustrated in Figures 2(c) and 2(d)tracing images from the MWM test showed that in the spaceexploration test where the platform was removed the MICDrats passed through the original position of the platformfewer times than the Sham group (119901 lt 001 Figure 2(d))
whereas in the MICD+EA group the times where the ratscrossed the position of the platformwere significantly increa-sed compared with the MICD group and the MICD+non-EA group (119901 lt 001 119901 lt 005 Figure 2(d)) As illustratedin Figures 2(e) and 2(f) the percentage of time spent in thetarget quadrant was used for statistical analysis during theprobe trial The date showed that the rats spent more time inthe target quadrant compared to other quadrants (119901 lt 001Figure 2(e)) The MICD group spent less time in the targetquadrant compared with the other groups (119901 lt 001 Fig-ure 2(e))TheMICD+EA group spent more time in the targetquadrant compared with the MICD group and MICD+non-EA group (119901 lt 001 119901 lt 005 Figure 2(f)) Therefore theseresults suggest that acquisition or retention of spatial refer-ence learning and memory was ameliorated in MICD rats byEA treatment
32 EA Attenuated Left Cortex Hippocampus Corpus Stria-tum and Thalamus Lesions in MICD Rats The brain lesionswere determined by T2-weighted imaging (T2WI) (Figures3(a) and 3(c)) before and after EA treatment The volumes ofbrain lesions that included left cortex hippocampus corpusstriatum and thalamus regions in the all groups had nosignificant difference before EA treatment (119901 gt 005 Figures3(a) and 3(b)) There was mild spontaneous recovery for 14days in the brain lesions of the MICD group The volumesof left cortex hippocampus corpus striatum and thalamuslesions showed comprising approximately 23 of the wholebrain in theMICDgroup whereas the brain lesions sizes weresignificantly reduced to 15 in the MICD+EA group (119901 lt001 Figures 3(c) and 3(d)) The difference of brain lesionsbetween the MICD+EA and the MICD+non-EA groupshowed significant changes at Day 14 after EA (119901 lt 005Figures 3(c) and 3(d))
33 EA Increased the Density of Dendritic Spines in the Hip-pocampus ofMICDRats To investigate the function of EAonsynaptic plasticity dendritic spine density in the hippocampalneurons was analyzed by the primary basilar dendrites ofGolgi-stained pyramidal neurons at Days 14 after EA Golgi-Cox staining clearly filled the dendritic shafts and the spinesof neurons frompyramidal neurons (Figure 4(a))Hereby therepresentative hippocampus showed that the density of den-dritic spines was reduced in different degree by macroscopicexamination in the MICD group the MICD+EA group andMICD+non-EA group and the loss of dendritic spines inhippocampal CA1 was obvious in the MICD group Thus asillustrated in Figures 4(b) and 4(c) the density of selecteddendritic spines that derived from hippocampal CA1 wassignificantly decreased in the MICD group compared withthe Sham group (119901 lt 001) however the density of dendriticspines of the hippocampal CA1 in the MICD+EA group wasmore than that of the MICD group and the MICD+non-EAgroup (119901 lt 001) In brief EA treatment triggered large-scale remodeling of dendrites in the hippocampal area CA1
34 EA Enhanced the Number of Hippocampus CA1 SynapsesinMICD Rats Furthermore we observed the effect of EA on
Neural Plasticity 5
Esca
pe la
tenc
y (s
)
Day 10 Day 11 Day 12 Day 13Days after MICD
ShamMICD
MICD+EAMICD+non-EA
lowastlowast
lowastlowast
lowastlowastlowastlowast
and
and
andand
0
10
20
30
40
50
60
70
80
90
100
(a)
Day 10 Day 11 Day 12 Day 13Training time
ShamMICD
MICD+EAMICD+non-EA
0
200
400
600
800
1000
1200
Dist
ance
trav
elle
d (c
m)
(b)
Sham MICD
MICD+EA MICD+non-EA
III
III IV
III
III IV
III
III IV
III
III IV
(c)
Sham MICD MICD+EA MICD+non-EA
lowastlowast
and
Prob
e tim
e
0
1
2
3
4
5
6
7
(d)
ShamMICD
MICD+EAMICD+non-EA
lowastlowast and
ti
me s
pent
in ea
ch q
uadr
ant
0
20
40
60
Opp
osite
Left
adja
cent
Targ
etRi
ght a
djac
ent
Opp
osite
Left
adja
cent
Targ
etRi
ght a
djac
ent
Opp
osite
Left
adja
cent
Targ
etRi
ght a
djac
ent
Opp
osite
Left
adja
cent
Targ
etRi
ght a
djac
ent
(e)
Sham MICD MICD+EA MICD+non-EA
lowastlowast
and
ti
me s
pent
in ta
rget
qua
dran
t
0
5
10
15
20
25
30
35
40
(f)
Figure 2 Evaluation of spatial reference learning and memory by Morris water maze test at Days 10ndash14 after EA (a and b) Mean escapelatency time and path length during the orientation navigation test on Days 10ndash13 after EA treatment (c and d) Times the rats crossed overprevious platform location on Day 14 after EA treatment during the spatial memory test in different groups (e)The percentage of times spentin each quadrant in all probe trials is shown on Day 14 after EA treatment (f) The times of passing the hidden platform position on Day 14after EA treatment (each group lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD group andand119901 lt 001 and119901 lt 005versus the MICD+EA group) EA electroacupuncture MICD middle cerebral artery occlusion induced cognitive deficits
6 Neural Plasticity
MICD+non-EA
MICD+EA
MICD
Sham
(a)
MICD+non-EA
MICD+EAMICD
Lesio
ns v
olum
es (
)
0
10
20
30
40
(b)
MICD+non-EA
MICD+EA
MICD
Sham
(c)
MICD+non-EA
MICD+EAMICD
Lesio
ns v
olum
es (
)
0
10
20
30
and
(d)
Figure 3 T2WI signal changes before and after EA (a and c)Thebrain lesionsweremeasured byT2WI (slices from 11 to 13) in the Shamgroupthe MICD group the MICD+EA group and the MICD+non-EA group before (a) and after (c) EA (b and d) Lesion volume is representedas a percentage of the total brain volume and data are presented as the mean plusmn standard deviation from 12 individual rats in each group(lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD group andand119901 lt 001 and119901 lt 005 versus the MICD+EA group)T2WI T2-weighted magnetic resonance imaging MICD middle cerebral artery occlusion induced cognitive deficits
ultrastructural morphology of hippocampal CA1 pyramidalneurons Obviously the density of synapses in the MICDgroup was decreased compared with the Sham group (119901 lt001 Figures 5(a) and 5(b)) whereas there was amplifying inthe number of synapses in the MICD+EA group comparedwith the MICD group and the MICD+non-EA group (119901 lt005 or 119901 lt 001 Figures 5(a) and 5(b))
Taken together these results suggest that EA couldimprove synaptic-dendritic plasticity in vivo
35 EA Promoted LIMK1 Expression and Phosphorylation ofHippocampus CA1 in MICD Rats To explore the underly-ing molecular mechanism of EA-induced synaptic-dendriticplasticity the levels of total LIMK1 and phospho-LIMK1 (P-LIMK1 Thr508) in hippocampal CA1 were investigated Asshown in Figures 6(a) and 6(b) the expression of total LIMK1of hippocampal CA1was significantly decreased in theMICDgroup compared with the Sham group (119901 lt 001) Howeverthe total LIMK1 level in the MICD+EA group was morethan that of the MICD group and the MICD+non-EA group(119901 lt 001 Figures 6(a) and 6(b)) Moreover the changes ofphospho-LIMK1 (Thr508) in groups were similar to the totalLIMK1 expression The level of P-LIMK1 was significantly
increased in the MICD+EA group compared with the MICDgroup and the MICD+non-EA group (119901 lt 001 Figures 6(a)and 6(c))
36 EA Regulated miR-134 Expression in Hippocampal CA1 inMICD Rats To identify the role of synaptic-dendrite-relatedmiR-134 in hippocampus CA1 the levels were detected Asshown in Figure 7 the expression ofmiR-134was significantlyincreased in theMICD group comparedwith the Sham group(119901 lt 001) However repeated EA treatment significantlydecreased the expression of miR-134 compared with theMICD group and the MICD+non-EA group (119901 lt 005)
4 Discussion
Ischemic stroke leads to a high incidence of long-termcognitive impairment which is strongly associated with lossof hippocampal neurons and synapses [34] While review-ing ancient Chinese documents regarding acupuncture andcognitive impairment-related terms we discovered that theDU20 andDU24 acupoints were themost frequently selectedacupoints for cognitive impairment-related rehabilitation inChina A number of studies have shown that EA can improve
Neural Plasticity 7
MICD+non-EAMICD+EA
MICDSham
(a)
MICD+non-EA
MICD+EA
MICD
Sham
(b)
MICD+non-EA
MICD+EAMICDSham0
5
10
15
20
lowastlowastandand
Spin
e den
sity
(10
um)
(c)
Figure 4 The structures and densities of dendritic spine in the hippocampus of rats (a) Representative images of dendritic spine in thehippocampus of each group (Golgi staining times50) (b) Representative images of dendritic spine density and morphology from the pyramidalcell layer of hippocampal CA1 area in each group (Golgi staining times1000) (c)The density of dendritic spine was analyzed in hippocampal CA1pyramidal cells in each group (119899 = 6group lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD group andand119901 lt 001and119901 lt 005 versus the MICD+EA group) All experiments were repeated three times
learning and memory ability and stimulate consciousness[30 31 35]These results indicated that EA could be a comple-mentary therapy for cognitive impairment after stroke Thepresent study found that EA at theDU20 andDU24 acupointsshortened time to find the platform and increased the timesof crossing the position of the platform compared to untreat-ment or nonacupoint EA (stimulation control) in Morriswater maze test suggesting that EA could improve spatialreference learning and memory ability in MCAO-inducedcognitive deficit (MICD) rats Moreover it is worth mention-ing that the path length in the Morris water maze in the fourgroups showed no significant difference indicating that thetime spending ofMICD rats to find the objectwas not affectedby themotor functionHowever themechanisms of cognitivetreatment involved are far from being fully elucidated
Firstly the cognitive deficit-related brain regions weredetermined by a small animal MRI in rats with ischemicstroke T2-weighted imaging showed that MICD caused thelesions of left cortex hippocampus corpus striatum andthalamus regions before EA It had beennoticed that the brain
lesion exhibited amild spontaneous recovery performance inMICD rats However the lesion regions comprising approxi-mately 23of thewhole brain volumewere reduced to 15byEA treatment indicating that EA could attenuate cortex andhippocampus corpus striatum and thalamus region lesionsin theMICD rats Some of these regions such as the hippoca-mpus and cortex are essential for regulating learning andmemory behaviors including spatial exploration [36] Studieshave confirmed that the hippocampus regions play a veryimportant role in learning and memory through their spe-cific structure and location which connect with other brainregions together [37] Furthermore in order to identify thehippocampus function in cognitive deficit the hippocampalmorphology staining was observed
Using Golgi staining and electron microscopy we demo-nstrated that the density of dendritic spine and the number ofsynapses in hippocampal CA1 pyramidal cells were obviouslyreduced at Day 14 after MICD However EA can rescue theloss of dendritic spine and synapses in hippocampal CA1region Moreover EA promoted synaptic-dendritic spine
8 Neural Plasticity
MICD+non-EAMICD+EA
MICDSham
(a)
MICD+non-EA
MICD+EAMICDSham0
2
4
6
8
10
The n
umbe
r of s
ynap
ses (10cm
2)
lowastlowast andand
(b)
Figure 5The number of synapses in hippocampal CA1 pyramidal cells (a) Representative electronmicrographic images of synapses (arrows)in the CA1 region of each group (magnification times50000) (b) Histogram shows the significant difference in the mean plusmn SEM of the numberof synapses in each group (119899 = 5group lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD group andand119901 lt 001and119901 lt 005 versus the MICD+EA group)
MICD+non-EAMICD+EAMICD
120573-Actin
P-LIMK1
T-LIMK1
Sham
(a)
MICD+non-EA
MICD+EAMICDSham00
02
04
06
08
10
Tota
l LIM
K1120573
-act
in
lowastlowastandand
(b)
MICD+non-EA
MICD+EAMICDSham00
02
04
06
08
10
lowastlowastandand
P-LI
MK1
T-L
IMK1
(c)
Figure 6 EA amplified the LIMK1 level in the hippocampus CA1 pyramidal cells (a) The expression of total LIMK1 and phosphorylatedLIMK1was evaluated usingwestern blotting (b and c)Histogram shows significant difference of the levels of P-LIMK1 and phosphor-LIMLK1in each group Data are means plusmn SEM (119899 = 6group lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD groupandand119901 lt 001 and119901 lt 005 versus the MICD+EA group)
Neural Plasticity 9
0
2
4
6
8
10
12
miR
-134
relat
ive e
xpre
ssio
n to
U6 lowastlowast
and
MICD+non-EA
MICD+EAMICDSham
Figure 7 Changes of miR-134 expression in the hippocampusCA1 cells The changes of miR-134 expression level were measuredusing quantitative RT-PCR U6 was used as internal control forquantification of miRNAs Data are mean plusmn SEM (119899 = 6grouplowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus theMICD group andand119901 lt 001 and119901 lt 005 versus the MICD+EA group)
regeneration Dendritic spine is the tiny protrusions withmulti-dense body and ion channels on the surface of varioustypes of neurons which serve as cellular substrates of brainconnectivity and the major sites of information processingin the brain [38 39] Evidence is increasing that synaptic-dendritic plasticity enables the encoding of memory [40]Indeed themodification on dendritic spine number size andshape is of importance for the plasticity of synapses accom-panied by hippocampus-dependent learning and memory[41 42] Studies have well described that dendritic spineremodeling in ischemia surrounding areas may contribute tocognitive functional recovery after stroke [43 44]
Increasing evidence showed that LIMK1 protein is acti-vated at dendritic spine membranes [45] LIMK1 positivecells were located in the CA1 region of the hippocampuswhich is important in the regulation of the spinemorphologyand synaptic function in vivo A prominent form of longlasting synaptic plasticity is thought to be critical to memoryformation [26] The present study found that the expressionof total LIMK1 and phospho-LIMK1 in hippocampal CA1wassignificantly decreased in the MICD rats however EA couldenhance the total LIMK1 and phospho-LIMK1 levels to pro-mote synaptic-dendritic plasticity in hippocampal CA1 area
In addition we have identified a dendritically localizedmiRNA that regulated the expression of the synaptic LIMK1protein thereby controlling dendritic pine sizemiR-134 is thefirst discovered dendritic microRNA enriched in the neu-ronal dendrites of rat hippocampal neurons and negativelycontrols the size of dendritic spines [46] This effect ismediated by miR-134 inhibition of the translation of LIMK1mRNA [47] The present study showed that the expression ofmiR-134 was obviously increased in theMICD rats Howeverrepeated EA treatment could relieve the upregulation of miR-134 expression in hippocampal CA1 area
In conclusion the study indicated that EA at the DU20and DU24 acupoints could ameliorate cognitive impairmentof MICD rats through the regulation of synaptic-dendritic
plasticity of the hippocampal CA1 area and the expressionand phosphorylation of synaptic LIMK1 protein were acti-vated to control dendritic spine number size and shapeFurthermore the mechanism of LIMK1 increase induced byEA treatmentmay be associated withmiR-134 which is local-ized in hippocampal CA1 by negatively regulating LIMK1 toenhance synaptic-dendritic plasticity in the recovery stage ofischemic stroke
Competing Interests
The authors declare no financial or commercial conflict ofinterests
Authorsrsquo Contributions
Weilin Liu Jie Wu and Jia Huang contributed equally to thiswork
Acknowledgments
The present study was supported by the Natural ScienceFoundation of China (no 81403450) and the Natural ScienceFoundation of Fujian Province (no 2016J01382)
References
[1] V L Roger ldquoHeart disease and stroke statisticsmdash2011 update areport from the American Heart Associationrdquo Circulation vol131 no 4 pp 29ndash322 2014
[2] T E Nichols and A P Holmes ldquoNonparametric permutationtests for functional neuroimaging a primer with examplesrdquoHuman Brain Mapping vol 15 no 1 pp 1ndash25 2002
[3] T K Tatemichi D W Desmond Y Stern M Paik M Sanoand E Bagiella ldquoCognitive impairment after stroke frequencypatterns and relationship to functional abilitiesrdquo Journal ofNeurology Neurosurgery amp Psychiatry vol 57 no 2 pp 202ndash207 1994
[4] F Liu Z-M Li Y-J Jiang and L-D Chen ldquoA meta-analysis ofacupuncture use in the treatment of cognitive impairment afterstrokerdquo Journal of Alternative amp Complementary Medicine vol20 no 7 pp 535ndash544 2014
[5] W Li R Huang R A Shetty et al ldquoTransient focal cerebralischemia induces long-termcognitive function deficit in an exp-erimental ischemic stroke modelrdquo Neurobiology of Disease vol59 pp 18ndash25 2013
[6] P S Sachdev X Chen H Brodaty CThompson A Altendorfand W Wen ldquoThe determinants and longitudinal course ofpost-stroke mild cognitive impairmentrdquo Journal of the Inter-national Neuropsychological Society vol 15 no 6 pp 915ndash9232009
[7] A-J Liu J-H Li H-Q Li et al ldquoElectroacupuncture foracute ischemic stroke ameta-analysis of randomized controlledtrialsrdquoThe American Journal of Chinese Medicine vol 43 no 8pp 1541ndash1566 2015
[8] X Ding C-Y Li Q-S Wang et al ldquoPatterns in default-modenetwork connectivity for determining outcomes in cognitivefunction in acute stroke patientsrdquo Neuroscience vol 277 no 10pp 637ndash646 2014
10 Neural Plasticity
[9] H Alipanahzadeh M Soleimani S Soleimani Asl B Pourhey-dar A Nikkhah and M Mehdizadeh ldquoTransforming growthfactor-alpha improves memory impairment and neurogenesisfollowing ischemia reperfusionrdquo Cell Journal vol 16 no 3 pp315ndash324 2014
[10] H Eichenbaum ldquoHippocampus cognitive processes and neuralrepresentations that underlie declarative memoryrdquoNeuron vol44 no 1 pp 109ndash120 2004
[11] T Kadar S Dachir B Shukitt-Hale and A Levy ldquoSub-regionalhippocampal vulnerability in various animal models leading tocognitive dysfunctionrdquo Journal of Neural Transmission vol 105no 8-9 pp 987ndash1004 1998
[12] J B Pereira C Valls-Pedret E Ros et al ldquoRegional vulnera-bility of hippocampal subfields to aging measured by structuraland diffusion MRIrdquo Hippocampus vol 24 no 4 pp 403ndash4142014
[13] S Knafo G Ariav E Barkai and F Libersat ldquoOlfactory learn-ing-induced increase in spine density along the apical dendritesof CA1 hippocampal neuronsrdquo Hippocampus vol 14 no 7 pp819ndash825 2004
[14] S Schacher and J-Y Hu ldquoThe less things change the more theyare different contributions of long-term synaptic plasticity andhomeostasis to memoryrdquo Learning and Memory vol 21 no 3pp 128ndash134 2014
[15] K Park H Heo M E Han et al ldquoLearning-induced synapticpotentiation in implanted neural precursor cell-derived neu-ronsrdquo Scientific Reports vol 5 article 17796 2015
[16] S Knafo F Libersat and E Barkai ldquoDynamics of learning-induced spine redistribution along dendrites of pyramidalneurons in ratsrdquo European Journal of Neuroscience vol 21 no4 pp 927ndash935 2005
[17] V C Foletta N Moussi P D Sarmiere J R Bamburg and OBernard ldquoLIM kinase 1 a key regulator of actin dynamics iswidely expressed in embryonic and adult tissuesrdquo ExperimentalCell Research vol 294 no 2 pp 392ndash405 2004
[18] M Endo K Ohashi Y Sasaki et al ldquoControl of growth conemotility andmorphology by LIMkinase and slingshot via phos-phorylation and dephosphorylation of cofilinrdquo Journal of Neu-roscience vol 23 no 7 pp 2527ndash2537 2003
[19] C Jarrold A D Baddeley and C Phillips ldquoLong-termmemoryfor verbal and visual information in Down syndrome andWilliams syndrome performance on the doors and people testrdquoCortex vol 43 no 2 pp 233ndash247 2007
[20] Y Meng Y Zhang V Tregoubov et al ldquoAbnormal spinemorphology and enhanced LTP in LIMK-1 knockout micerdquoNeuron vol 35 no 1 pp 121ndash133 2002
[21] Z Todorovski S Asrar J Liu et al ldquoLIMK1 regulates long-termmemory and synaptic plasticity via the transcriptional factorCREBrdquo Molecular and Cellular Biology vol 35 no 8 pp 1316ndash1328 2015
[22] G M Schratt F Tuebing E A Nigh et al ldquoA brain-specificmicroRNA regulates dendritic spine developmentrdquo Nature vol439 no 7074 pp 283ndash289 2006
[23] F Capitano J Camon V Ferretti et al ldquomicroRNAs modulatespatial memory in the hippocampus and in the ventral striatumin a region-specific mannerrdquo Molecular Neurobiology vol 53no 7 pp 4618ndash4630 2015
[24] J GaoW-YWang Y-WMao et al ldquoA novel pathway regulatesmemory and plasticity via SIRT1 andmiR-134rdquoNature vol 466no 7310 pp 1105ndash1109 2010
[25] J Remenyi M W M van den Bosch O Palygin et al ldquomiR-132212 knockout mice reveal roles for these miRNAs in regu-lating cortical synaptic transmission and plasticityrdquo PLoS ONEvol 8 no 4 Article ID e62509 p 65 2013
[26] L An J LiuWW Li et al ldquoDistribution of LIM domain kinase1 in the olfactory bulb cerebral cortex hippocampus and cere-bellum of the AppPS+minus micerdquo Genetics amp Molecular Researchvol 14 no 4 pp 17244ndash17251 2015
[27] R Lin Y Wu J Tao et al ldquoElectroacupuncture improves cog-nitive function through Rho GTPases and enhances dendriticspine plasticity in rats with cerebral ischemia-reperfusionrdquoMolecular Medicine Reports vol 13 no 3 pp 2655ndash2660 2016
[28] P Castaneda M Munoz G Garcıa-Rojo et al ldquoAssociation ofN-cadherin levels and downstream effectors of Rho GTPaseswith dendritic spine loss induced by chronic stress in rathippocampal neuronsrdquo Journal of Neuroscience Research vol93 no 10 pp 1476ndash1491 2015
[29] A Romarowski M A Battistone F A La Spina et al ldquoPKA-dependent phosphorylation of LIMK1 and Cofilin is essentialformouse sperm acrosomal exocytosisrdquoDevelopmental Biologyvol 405 no 2 pp 237ndash249 2015
[30] R Lin K Yu X Li et al ldquoElectroacupuncture ameliorates post-stroke learning and memory through minimizing ultrastruc-tural brain damage and inhibiting the expression of MMP-2andMMP-9 in cerebral ischemia-reperfusion injured ratsrdquoMol-ecular Medicine Reports vol 14 no 1 pp 225ndash233 2016
[31] X Feng S Yang J Liu et al ldquoElectroacupuncture amelioratescognitive impairment through inhibition of NF-120581B-mediatedneuronal cell apoptosis in cerebral ischemia-reperfusion inj-ured ratsrdquo Molecular Medicine Reports vol 7 no 5 pp 1516ndash1522 2013
[32] Z K Wang N I Guang-Xia K Liu et al ldquoResearch on thechanges of IL-1 receptor and TNF-120572 receptor in rats with cere-bral ischemia reperfusion and the chronergy of acupunctureinterventionrdquo Zhongguo Zhen Jiu vol 32 no 11 pp 1012ndash10182012
[33] X S LiuM Chopp R L Zhang and Z G Zhang ldquoMicroRNAsin cerebral ischemia-induced neurogenesisrdquo Journal of Neu-ropathology amp Experimental Neurology vol 72 no 8 pp 718ndash722 2013
[34] X He T Yan R Chen and D Ran ldquoAcute effects of electro-acupuncture (EA) on hippocampal long term potentiation(LTP) of perforant path-dentate gyrus granule cells synapserelated to memoryrdquo Acupuncture and Electro-TherapeuticsResearch vol 37 no 2-3 pp 89ndash101 2012
[35] W Dong W Guo X Zheng et al ldquoElectroacupunctureimproves cognitive deficits associated with AMPK activation inSAMP8 micerdquo Metabolic Brain Disease vol 30 no 3 pp 777ndash784 2015
[36] B Milner and D Klein ldquoLoss of recent memory after bilateralhippocampal lesions memory and memoriesmdashlooking backand looking forwardrdquo Journal of Neurology Neurosurgery ampPsychiatry vol 87 no 3 article 230 2016
[37] A Veronica Witte L Kerti D S Margulies and A FloelldquoEffects of resveratrol on memory performance hippocampalfunctional connectivity and glucose metabolism in healthyolder adultsrdquo Journal of Neuroscience vol 34 no 23 pp 7862ndash7870 2014
[38] T Bonhoeffer and R Yuste ldquoSpine motility phenomenologymechanisms and functionrdquoNeuron vol 35 no 6 pp 1019ndash10272002
Neural Plasticity 11
[39] H Qiao M Li C Xu H Chen S An and X Ma ldquoDendriticspines in depression what we learned from animal modelsrdquoNeural Plasticity vol 2016 Article ID 8056370 26 pages 2016
[40] S C McQuown R M Barrett D P Matheos et al ldquoHDAC3 isa critical negative regulator of long-term memory formationrdquoJournal of Neuroscience vol 31 no 2 pp 764ndash774 2011
[41] K-O Lai and N Y Ip ldquoStructural plasticity of dendritic spinesthe underlying mechanisms and its dysregulation in brain dis-ordersrdquo Biochimica et Biophysica ActamdashMolecular Basis of Dis-ease vol 1832 no 12 pp 2257ndash2263 2013
[42] C H Bailey E R Kandel and K M Harris ldquoStructural com-ponents of synaptic plasticity and memory consolidationrdquo ColdSpring Harbor Perspectives in Biology vol 7 no 7 Article IDa021758 2015
[43] C E Brown P Li J D Boyd K R Delaney and T HMurphy ldquoExtensive turnover of dendritic spines and vascularremodeling in cortical tissues recovering from strokerdquo Journalof Neuroscience vol 27 no 15 pp 4101ndash4109 2007
[44] R Mostany T G Chowdhury D G Johnston S A PortonovoS T Carmichael and C Portera-Cailliau ldquoLocal hemodynam-ics dictate long-term dendritic plasticity in peri-infarct cortexrdquoJournal of Neuroscience vol 30 no 42 pp 14116ndash14126 2010
[45] J George C Soares A Montersino J-C Beique and G MThomas ldquoPalmitoylation of LIM kinase-1 ensures spine-specificactin polymerization and morphological plasticityrdquo eLife vol2015 no 4 Article ID e06327 2015
[46] S Bicker M Lackinger K Weiszlig and G Schratt ldquoMicroRNA-132 -134 and -138 a microRNA troika rules in neuronaldendritesrdquo Cellular and molecular life sciences CMLS vol 71no 20 pp 3987ndash4005 2014
[47] H-C Tai and E M Schuman ldquoMicroRNA microRNAs reachout into dendritesrdquoCurrent Biology vol 16 no 4 pp R121ndashR1232006
Submit your manuscripts athttpswwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
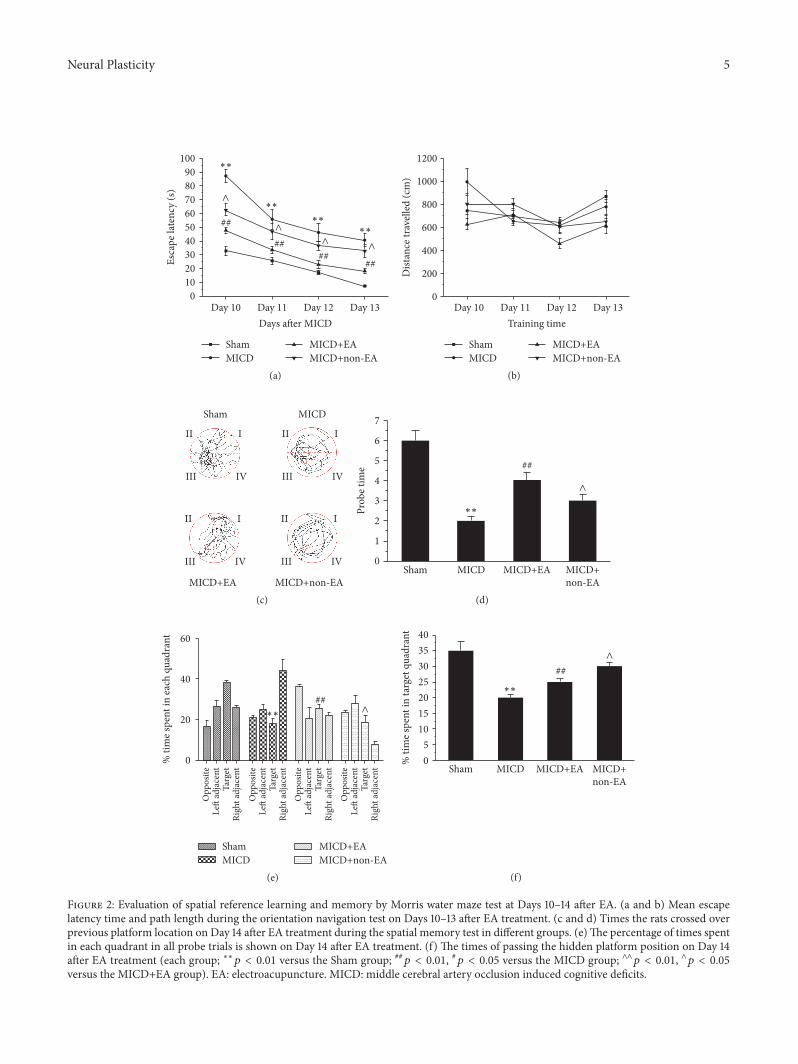
Neural Plasticity 5
Esca
pe la
tenc
y (s
)
Day 10 Day 11 Day 12 Day 13Days after MICD
ShamMICD
MICD+EAMICD+non-EA
lowastlowast
lowastlowast
lowastlowastlowastlowast
and
and
andand
0
10
20
30
40
50
60
70
80
90
100
(a)
Day 10 Day 11 Day 12 Day 13Training time
ShamMICD
MICD+EAMICD+non-EA
0
200
400
600
800
1000
1200
Dist
ance
trav
elle
d (c
m)
(b)
Sham MICD
MICD+EA MICD+non-EA
III
III IV
III
III IV
III
III IV
III
III IV
(c)
Sham MICD MICD+EA MICD+non-EA
lowastlowast
and
Prob
e tim
e
0
1
2
3
4
5
6
7
(d)
ShamMICD
MICD+EAMICD+non-EA
lowastlowast and
ti
me s
pent
in ea
ch q
uadr
ant
0
20
40
60
Opp
osite
Left
adja
cent
Targ
etRi
ght a
djac
ent
Opp
osite
Left
adja
cent
Targ
etRi
ght a
djac
ent
Opp
osite
Left
adja
cent
Targ
etRi
ght a
djac
ent
Opp
osite
Left
adja
cent
Targ
etRi
ght a
djac
ent
(e)
Sham MICD MICD+EA MICD+non-EA
lowastlowast
and
ti
me s
pent
in ta
rget
qua
dran
t
0
5
10
15
20
25
30
35
40
(f)
Figure 2 Evaluation of spatial reference learning and memory by Morris water maze test at Days 10ndash14 after EA (a and b) Mean escapelatency time and path length during the orientation navigation test on Days 10ndash13 after EA treatment (c and d) Times the rats crossed overprevious platform location on Day 14 after EA treatment during the spatial memory test in different groups (e)The percentage of times spentin each quadrant in all probe trials is shown on Day 14 after EA treatment (f) The times of passing the hidden platform position on Day 14after EA treatment (each group lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD group andand119901 lt 001 and119901 lt 005versus the MICD+EA group) EA electroacupuncture MICD middle cerebral artery occlusion induced cognitive deficits
6 Neural Plasticity
MICD+non-EA
MICD+EA
MICD
Sham
(a)
MICD+non-EA
MICD+EAMICD
Lesio
ns v
olum
es (
)
0
10
20
30
40
(b)
MICD+non-EA
MICD+EA
MICD
Sham
(c)
MICD+non-EA
MICD+EAMICD
Lesio
ns v
olum
es (
)
0
10
20
30
and
(d)
Figure 3 T2WI signal changes before and after EA (a and c)Thebrain lesionsweremeasured byT2WI (slices from 11 to 13) in the Shamgroupthe MICD group the MICD+EA group and the MICD+non-EA group before (a) and after (c) EA (b and d) Lesion volume is representedas a percentage of the total brain volume and data are presented as the mean plusmn standard deviation from 12 individual rats in each group(lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD group andand119901 lt 001 and119901 lt 005 versus the MICD+EA group)T2WI T2-weighted magnetic resonance imaging MICD middle cerebral artery occlusion induced cognitive deficits
ultrastructural morphology of hippocampal CA1 pyramidalneurons Obviously the density of synapses in the MICDgroup was decreased compared with the Sham group (119901 lt001 Figures 5(a) and 5(b)) whereas there was amplifying inthe number of synapses in the MICD+EA group comparedwith the MICD group and the MICD+non-EA group (119901 lt005 or 119901 lt 001 Figures 5(a) and 5(b))
Taken together these results suggest that EA couldimprove synaptic-dendritic plasticity in vivo
35 EA Promoted LIMK1 Expression and Phosphorylation ofHippocampus CA1 in MICD Rats To explore the underly-ing molecular mechanism of EA-induced synaptic-dendriticplasticity the levels of total LIMK1 and phospho-LIMK1 (P-LIMK1 Thr508) in hippocampal CA1 were investigated Asshown in Figures 6(a) and 6(b) the expression of total LIMK1of hippocampal CA1was significantly decreased in theMICDgroup compared with the Sham group (119901 lt 001) Howeverthe total LIMK1 level in the MICD+EA group was morethan that of the MICD group and the MICD+non-EA group(119901 lt 001 Figures 6(a) and 6(b)) Moreover the changes ofphospho-LIMK1 (Thr508) in groups were similar to the totalLIMK1 expression The level of P-LIMK1 was significantly
increased in the MICD+EA group compared with the MICDgroup and the MICD+non-EA group (119901 lt 001 Figures 6(a)and 6(c))
36 EA Regulated miR-134 Expression in Hippocampal CA1 inMICD Rats To identify the role of synaptic-dendrite-relatedmiR-134 in hippocampus CA1 the levels were detected Asshown in Figure 7 the expression ofmiR-134was significantlyincreased in theMICD group comparedwith the Sham group(119901 lt 001) However repeated EA treatment significantlydecreased the expression of miR-134 compared with theMICD group and the MICD+non-EA group (119901 lt 005)
4 Discussion
Ischemic stroke leads to a high incidence of long-termcognitive impairment which is strongly associated with lossof hippocampal neurons and synapses [34] While review-ing ancient Chinese documents regarding acupuncture andcognitive impairment-related terms we discovered that theDU20 andDU24 acupoints were themost frequently selectedacupoints for cognitive impairment-related rehabilitation inChina A number of studies have shown that EA can improve
Neural Plasticity 7
MICD+non-EAMICD+EA
MICDSham
(a)
MICD+non-EA
MICD+EA
MICD
Sham
(b)
MICD+non-EA
MICD+EAMICDSham0
5
10
15
20
lowastlowastandand
Spin
e den
sity
(10
um)
(c)
Figure 4 The structures and densities of dendritic spine in the hippocampus of rats (a) Representative images of dendritic spine in thehippocampus of each group (Golgi staining times50) (b) Representative images of dendritic spine density and morphology from the pyramidalcell layer of hippocampal CA1 area in each group (Golgi staining times1000) (c)The density of dendritic spine was analyzed in hippocampal CA1pyramidal cells in each group (119899 = 6group lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD group andand119901 lt 001and119901 lt 005 versus the MICD+EA group) All experiments were repeated three times
learning and memory ability and stimulate consciousness[30 31 35]These results indicated that EA could be a comple-mentary therapy for cognitive impairment after stroke Thepresent study found that EA at theDU20 andDU24 acupointsshortened time to find the platform and increased the timesof crossing the position of the platform compared to untreat-ment or nonacupoint EA (stimulation control) in Morriswater maze test suggesting that EA could improve spatialreference learning and memory ability in MCAO-inducedcognitive deficit (MICD) rats Moreover it is worth mention-ing that the path length in the Morris water maze in the fourgroups showed no significant difference indicating that thetime spending ofMICD rats to find the objectwas not affectedby themotor functionHowever themechanisms of cognitivetreatment involved are far from being fully elucidated
Firstly the cognitive deficit-related brain regions weredetermined by a small animal MRI in rats with ischemicstroke T2-weighted imaging showed that MICD caused thelesions of left cortex hippocampus corpus striatum andthalamus regions before EA It had beennoticed that the brain
lesion exhibited amild spontaneous recovery performance inMICD rats However the lesion regions comprising approxi-mately 23of thewhole brain volumewere reduced to 15byEA treatment indicating that EA could attenuate cortex andhippocampus corpus striatum and thalamus region lesionsin theMICD rats Some of these regions such as the hippoca-mpus and cortex are essential for regulating learning andmemory behaviors including spatial exploration [36] Studieshave confirmed that the hippocampus regions play a veryimportant role in learning and memory through their spe-cific structure and location which connect with other brainregions together [37] Furthermore in order to identify thehippocampus function in cognitive deficit the hippocampalmorphology staining was observed
Using Golgi staining and electron microscopy we demo-nstrated that the density of dendritic spine and the number ofsynapses in hippocampal CA1 pyramidal cells were obviouslyreduced at Day 14 after MICD However EA can rescue theloss of dendritic spine and synapses in hippocampal CA1region Moreover EA promoted synaptic-dendritic spine
8 Neural Plasticity
MICD+non-EAMICD+EA
MICDSham
(a)
MICD+non-EA
MICD+EAMICDSham0
2
4
6
8
10
The n
umbe
r of s
ynap
ses (10cm
2)
lowastlowast andand
(b)
Figure 5The number of synapses in hippocampal CA1 pyramidal cells (a) Representative electronmicrographic images of synapses (arrows)in the CA1 region of each group (magnification times50000) (b) Histogram shows the significant difference in the mean plusmn SEM of the numberof synapses in each group (119899 = 5group lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD group andand119901 lt 001and119901 lt 005 versus the MICD+EA group)
MICD+non-EAMICD+EAMICD
120573-Actin
P-LIMK1
T-LIMK1
Sham
(a)
MICD+non-EA
MICD+EAMICDSham00
02
04
06
08
10
Tota
l LIM
K1120573
-act
in
lowastlowastandand
(b)
MICD+non-EA
MICD+EAMICDSham00
02
04
06
08
10
lowastlowastandand
P-LI
MK1
T-L
IMK1
(c)
Figure 6 EA amplified the LIMK1 level in the hippocampus CA1 pyramidal cells (a) The expression of total LIMK1 and phosphorylatedLIMK1was evaluated usingwestern blotting (b and c)Histogram shows significant difference of the levels of P-LIMK1 and phosphor-LIMLK1in each group Data are means plusmn SEM (119899 = 6group lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD groupandand119901 lt 001 and119901 lt 005 versus the MICD+EA group)
Neural Plasticity 9
0
2
4
6
8
10
12
miR
-134
relat
ive e
xpre
ssio
n to
U6 lowastlowast
and
MICD+non-EA
MICD+EAMICDSham
Figure 7 Changes of miR-134 expression in the hippocampusCA1 cells The changes of miR-134 expression level were measuredusing quantitative RT-PCR U6 was used as internal control forquantification of miRNAs Data are mean plusmn SEM (119899 = 6grouplowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus theMICD group andand119901 lt 001 and119901 lt 005 versus the MICD+EA group)
regeneration Dendritic spine is the tiny protrusions withmulti-dense body and ion channels on the surface of varioustypes of neurons which serve as cellular substrates of brainconnectivity and the major sites of information processingin the brain [38 39] Evidence is increasing that synaptic-dendritic plasticity enables the encoding of memory [40]Indeed themodification on dendritic spine number size andshape is of importance for the plasticity of synapses accom-panied by hippocampus-dependent learning and memory[41 42] Studies have well described that dendritic spineremodeling in ischemia surrounding areas may contribute tocognitive functional recovery after stroke [43 44]
Increasing evidence showed that LIMK1 protein is acti-vated at dendritic spine membranes [45] LIMK1 positivecells were located in the CA1 region of the hippocampuswhich is important in the regulation of the spinemorphologyand synaptic function in vivo A prominent form of longlasting synaptic plasticity is thought to be critical to memoryformation [26] The present study found that the expressionof total LIMK1 and phospho-LIMK1 in hippocampal CA1wassignificantly decreased in the MICD rats however EA couldenhance the total LIMK1 and phospho-LIMK1 levels to pro-mote synaptic-dendritic plasticity in hippocampal CA1 area
In addition we have identified a dendritically localizedmiRNA that regulated the expression of the synaptic LIMK1protein thereby controlling dendritic pine sizemiR-134 is thefirst discovered dendritic microRNA enriched in the neu-ronal dendrites of rat hippocampal neurons and negativelycontrols the size of dendritic spines [46] This effect ismediated by miR-134 inhibition of the translation of LIMK1mRNA [47] The present study showed that the expression ofmiR-134 was obviously increased in theMICD rats Howeverrepeated EA treatment could relieve the upregulation of miR-134 expression in hippocampal CA1 area
In conclusion the study indicated that EA at the DU20and DU24 acupoints could ameliorate cognitive impairmentof MICD rats through the regulation of synaptic-dendritic
plasticity of the hippocampal CA1 area and the expressionand phosphorylation of synaptic LIMK1 protein were acti-vated to control dendritic spine number size and shapeFurthermore the mechanism of LIMK1 increase induced byEA treatmentmay be associated withmiR-134 which is local-ized in hippocampal CA1 by negatively regulating LIMK1 toenhance synaptic-dendritic plasticity in the recovery stage ofischemic stroke
Competing Interests
The authors declare no financial or commercial conflict ofinterests
Authorsrsquo Contributions
Weilin Liu Jie Wu and Jia Huang contributed equally to thiswork
Acknowledgments
The present study was supported by the Natural ScienceFoundation of China (no 81403450) and the Natural ScienceFoundation of Fujian Province (no 2016J01382)
References
[1] V L Roger ldquoHeart disease and stroke statisticsmdash2011 update areport from the American Heart Associationrdquo Circulation vol131 no 4 pp 29ndash322 2014
[2] T E Nichols and A P Holmes ldquoNonparametric permutationtests for functional neuroimaging a primer with examplesrdquoHuman Brain Mapping vol 15 no 1 pp 1ndash25 2002
[3] T K Tatemichi D W Desmond Y Stern M Paik M Sanoand E Bagiella ldquoCognitive impairment after stroke frequencypatterns and relationship to functional abilitiesrdquo Journal ofNeurology Neurosurgery amp Psychiatry vol 57 no 2 pp 202ndash207 1994
[4] F Liu Z-M Li Y-J Jiang and L-D Chen ldquoA meta-analysis ofacupuncture use in the treatment of cognitive impairment afterstrokerdquo Journal of Alternative amp Complementary Medicine vol20 no 7 pp 535ndash544 2014
[5] W Li R Huang R A Shetty et al ldquoTransient focal cerebralischemia induces long-termcognitive function deficit in an exp-erimental ischemic stroke modelrdquo Neurobiology of Disease vol59 pp 18ndash25 2013
[6] P S Sachdev X Chen H Brodaty CThompson A Altendorfand W Wen ldquoThe determinants and longitudinal course ofpost-stroke mild cognitive impairmentrdquo Journal of the Inter-national Neuropsychological Society vol 15 no 6 pp 915ndash9232009
[7] A-J Liu J-H Li H-Q Li et al ldquoElectroacupuncture foracute ischemic stroke ameta-analysis of randomized controlledtrialsrdquoThe American Journal of Chinese Medicine vol 43 no 8pp 1541ndash1566 2015
[8] X Ding C-Y Li Q-S Wang et al ldquoPatterns in default-modenetwork connectivity for determining outcomes in cognitivefunction in acute stroke patientsrdquo Neuroscience vol 277 no 10pp 637ndash646 2014
10 Neural Plasticity
[9] H Alipanahzadeh M Soleimani S Soleimani Asl B Pourhey-dar A Nikkhah and M Mehdizadeh ldquoTransforming growthfactor-alpha improves memory impairment and neurogenesisfollowing ischemia reperfusionrdquo Cell Journal vol 16 no 3 pp315ndash324 2014
[10] H Eichenbaum ldquoHippocampus cognitive processes and neuralrepresentations that underlie declarative memoryrdquoNeuron vol44 no 1 pp 109ndash120 2004
[11] T Kadar S Dachir B Shukitt-Hale and A Levy ldquoSub-regionalhippocampal vulnerability in various animal models leading tocognitive dysfunctionrdquo Journal of Neural Transmission vol 105no 8-9 pp 987ndash1004 1998
[12] J B Pereira C Valls-Pedret E Ros et al ldquoRegional vulnera-bility of hippocampal subfields to aging measured by structuraland diffusion MRIrdquo Hippocampus vol 24 no 4 pp 403ndash4142014
[13] S Knafo G Ariav E Barkai and F Libersat ldquoOlfactory learn-ing-induced increase in spine density along the apical dendritesof CA1 hippocampal neuronsrdquo Hippocampus vol 14 no 7 pp819ndash825 2004
[14] S Schacher and J-Y Hu ldquoThe less things change the more theyare different contributions of long-term synaptic plasticity andhomeostasis to memoryrdquo Learning and Memory vol 21 no 3pp 128ndash134 2014
[15] K Park H Heo M E Han et al ldquoLearning-induced synapticpotentiation in implanted neural precursor cell-derived neu-ronsrdquo Scientific Reports vol 5 article 17796 2015
[16] S Knafo F Libersat and E Barkai ldquoDynamics of learning-induced spine redistribution along dendrites of pyramidalneurons in ratsrdquo European Journal of Neuroscience vol 21 no4 pp 927ndash935 2005
[17] V C Foletta N Moussi P D Sarmiere J R Bamburg and OBernard ldquoLIM kinase 1 a key regulator of actin dynamics iswidely expressed in embryonic and adult tissuesrdquo ExperimentalCell Research vol 294 no 2 pp 392ndash405 2004
[18] M Endo K Ohashi Y Sasaki et al ldquoControl of growth conemotility andmorphology by LIMkinase and slingshot via phos-phorylation and dephosphorylation of cofilinrdquo Journal of Neu-roscience vol 23 no 7 pp 2527ndash2537 2003
[19] C Jarrold A D Baddeley and C Phillips ldquoLong-termmemoryfor verbal and visual information in Down syndrome andWilliams syndrome performance on the doors and people testrdquoCortex vol 43 no 2 pp 233ndash247 2007
[20] Y Meng Y Zhang V Tregoubov et al ldquoAbnormal spinemorphology and enhanced LTP in LIMK-1 knockout micerdquoNeuron vol 35 no 1 pp 121ndash133 2002
[21] Z Todorovski S Asrar J Liu et al ldquoLIMK1 regulates long-termmemory and synaptic plasticity via the transcriptional factorCREBrdquo Molecular and Cellular Biology vol 35 no 8 pp 1316ndash1328 2015
[22] G M Schratt F Tuebing E A Nigh et al ldquoA brain-specificmicroRNA regulates dendritic spine developmentrdquo Nature vol439 no 7074 pp 283ndash289 2006
[23] F Capitano J Camon V Ferretti et al ldquomicroRNAs modulatespatial memory in the hippocampus and in the ventral striatumin a region-specific mannerrdquo Molecular Neurobiology vol 53no 7 pp 4618ndash4630 2015
[24] J GaoW-YWang Y-WMao et al ldquoA novel pathway regulatesmemory and plasticity via SIRT1 andmiR-134rdquoNature vol 466no 7310 pp 1105ndash1109 2010
[25] J Remenyi M W M van den Bosch O Palygin et al ldquomiR-132212 knockout mice reveal roles for these miRNAs in regu-lating cortical synaptic transmission and plasticityrdquo PLoS ONEvol 8 no 4 Article ID e62509 p 65 2013
[26] L An J LiuWW Li et al ldquoDistribution of LIM domain kinase1 in the olfactory bulb cerebral cortex hippocampus and cere-bellum of the AppPS+minus micerdquo Genetics amp Molecular Researchvol 14 no 4 pp 17244ndash17251 2015
[27] R Lin Y Wu J Tao et al ldquoElectroacupuncture improves cog-nitive function through Rho GTPases and enhances dendriticspine plasticity in rats with cerebral ischemia-reperfusionrdquoMolecular Medicine Reports vol 13 no 3 pp 2655ndash2660 2016
[28] P Castaneda M Munoz G Garcıa-Rojo et al ldquoAssociation ofN-cadherin levels and downstream effectors of Rho GTPaseswith dendritic spine loss induced by chronic stress in rathippocampal neuronsrdquo Journal of Neuroscience Research vol93 no 10 pp 1476ndash1491 2015
[29] A Romarowski M A Battistone F A La Spina et al ldquoPKA-dependent phosphorylation of LIMK1 and Cofilin is essentialformouse sperm acrosomal exocytosisrdquoDevelopmental Biologyvol 405 no 2 pp 237ndash249 2015
[30] R Lin K Yu X Li et al ldquoElectroacupuncture ameliorates post-stroke learning and memory through minimizing ultrastruc-tural brain damage and inhibiting the expression of MMP-2andMMP-9 in cerebral ischemia-reperfusion injured ratsrdquoMol-ecular Medicine Reports vol 14 no 1 pp 225ndash233 2016
[31] X Feng S Yang J Liu et al ldquoElectroacupuncture amelioratescognitive impairment through inhibition of NF-120581B-mediatedneuronal cell apoptosis in cerebral ischemia-reperfusion inj-ured ratsrdquo Molecular Medicine Reports vol 7 no 5 pp 1516ndash1522 2013
[32] Z K Wang N I Guang-Xia K Liu et al ldquoResearch on thechanges of IL-1 receptor and TNF-120572 receptor in rats with cere-bral ischemia reperfusion and the chronergy of acupunctureinterventionrdquo Zhongguo Zhen Jiu vol 32 no 11 pp 1012ndash10182012
[33] X S LiuM Chopp R L Zhang and Z G Zhang ldquoMicroRNAsin cerebral ischemia-induced neurogenesisrdquo Journal of Neu-ropathology amp Experimental Neurology vol 72 no 8 pp 718ndash722 2013
[34] X He T Yan R Chen and D Ran ldquoAcute effects of electro-acupuncture (EA) on hippocampal long term potentiation(LTP) of perforant path-dentate gyrus granule cells synapserelated to memoryrdquo Acupuncture and Electro-TherapeuticsResearch vol 37 no 2-3 pp 89ndash101 2012
[35] W Dong W Guo X Zheng et al ldquoElectroacupunctureimproves cognitive deficits associated with AMPK activation inSAMP8 micerdquo Metabolic Brain Disease vol 30 no 3 pp 777ndash784 2015
[36] B Milner and D Klein ldquoLoss of recent memory after bilateralhippocampal lesions memory and memoriesmdashlooking backand looking forwardrdquo Journal of Neurology Neurosurgery ampPsychiatry vol 87 no 3 article 230 2016
[37] A Veronica Witte L Kerti D S Margulies and A FloelldquoEffects of resveratrol on memory performance hippocampalfunctional connectivity and glucose metabolism in healthyolder adultsrdquo Journal of Neuroscience vol 34 no 23 pp 7862ndash7870 2014
[38] T Bonhoeffer and R Yuste ldquoSpine motility phenomenologymechanisms and functionrdquoNeuron vol 35 no 6 pp 1019ndash10272002
Neural Plasticity 11
[39] H Qiao M Li C Xu H Chen S An and X Ma ldquoDendriticspines in depression what we learned from animal modelsrdquoNeural Plasticity vol 2016 Article ID 8056370 26 pages 2016
[40] S C McQuown R M Barrett D P Matheos et al ldquoHDAC3 isa critical negative regulator of long-term memory formationrdquoJournal of Neuroscience vol 31 no 2 pp 764ndash774 2011
[41] K-O Lai and N Y Ip ldquoStructural plasticity of dendritic spinesthe underlying mechanisms and its dysregulation in brain dis-ordersrdquo Biochimica et Biophysica ActamdashMolecular Basis of Dis-ease vol 1832 no 12 pp 2257ndash2263 2013
[42] C H Bailey E R Kandel and K M Harris ldquoStructural com-ponents of synaptic plasticity and memory consolidationrdquo ColdSpring Harbor Perspectives in Biology vol 7 no 7 Article IDa021758 2015
[43] C E Brown P Li J D Boyd K R Delaney and T HMurphy ldquoExtensive turnover of dendritic spines and vascularremodeling in cortical tissues recovering from strokerdquo Journalof Neuroscience vol 27 no 15 pp 4101ndash4109 2007
[44] R Mostany T G Chowdhury D G Johnston S A PortonovoS T Carmichael and C Portera-Cailliau ldquoLocal hemodynam-ics dictate long-term dendritic plasticity in peri-infarct cortexrdquoJournal of Neuroscience vol 30 no 42 pp 14116ndash14126 2010
[45] J George C Soares A Montersino J-C Beique and G MThomas ldquoPalmitoylation of LIM kinase-1 ensures spine-specificactin polymerization and morphological plasticityrdquo eLife vol2015 no 4 Article ID e06327 2015
[46] S Bicker M Lackinger K Weiszlig and G Schratt ldquoMicroRNA-132 -134 and -138 a microRNA troika rules in neuronaldendritesrdquo Cellular and molecular life sciences CMLS vol 71no 20 pp 3987ndash4005 2014
[47] H-C Tai and E M Schuman ldquoMicroRNA microRNAs reachout into dendritesrdquoCurrent Biology vol 16 no 4 pp R121ndashR1232006
Submit your manuscripts athttpswwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
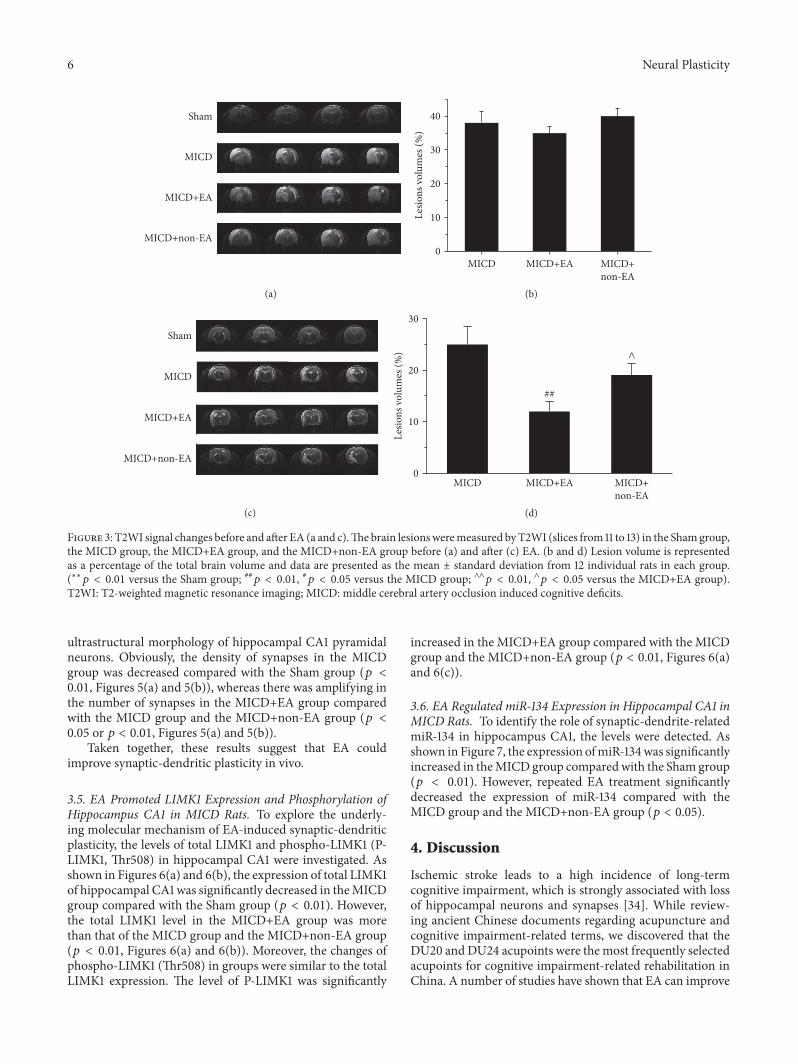
6 Neural Plasticity
MICD+non-EA
MICD+EA
MICD
Sham
(a)
MICD+non-EA
MICD+EAMICD
Lesio
ns v
olum
es (
)
0
10
20
30
40
(b)
MICD+non-EA
MICD+EA
MICD
Sham
(c)
MICD+non-EA
MICD+EAMICD
Lesio
ns v
olum
es (
)
0
10
20
30
and
(d)
Figure 3 T2WI signal changes before and after EA (a and c)Thebrain lesionsweremeasured byT2WI (slices from 11 to 13) in the Shamgroupthe MICD group the MICD+EA group and the MICD+non-EA group before (a) and after (c) EA (b and d) Lesion volume is representedas a percentage of the total brain volume and data are presented as the mean plusmn standard deviation from 12 individual rats in each group(lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD group andand119901 lt 001 and119901 lt 005 versus the MICD+EA group)T2WI T2-weighted magnetic resonance imaging MICD middle cerebral artery occlusion induced cognitive deficits
ultrastructural morphology of hippocampal CA1 pyramidalneurons Obviously the density of synapses in the MICDgroup was decreased compared with the Sham group (119901 lt001 Figures 5(a) and 5(b)) whereas there was amplifying inthe number of synapses in the MICD+EA group comparedwith the MICD group and the MICD+non-EA group (119901 lt005 or 119901 lt 001 Figures 5(a) and 5(b))
Taken together these results suggest that EA couldimprove synaptic-dendritic plasticity in vivo
35 EA Promoted LIMK1 Expression and Phosphorylation ofHippocampus CA1 in MICD Rats To explore the underly-ing molecular mechanism of EA-induced synaptic-dendriticplasticity the levels of total LIMK1 and phospho-LIMK1 (P-LIMK1 Thr508) in hippocampal CA1 were investigated Asshown in Figures 6(a) and 6(b) the expression of total LIMK1of hippocampal CA1was significantly decreased in theMICDgroup compared with the Sham group (119901 lt 001) Howeverthe total LIMK1 level in the MICD+EA group was morethan that of the MICD group and the MICD+non-EA group(119901 lt 001 Figures 6(a) and 6(b)) Moreover the changes ofphospho-LIMK1 (Thr508) in groups were similar to the totalLIMK1 expression The level of P-LIMK1 was significantly
increased in the MICD+EA group compared with the MICDgroup and the MICD+non-EA group (119901 lt 001 Figures 6(a)and 6(c))
36 EA Regulated miR-134 Expression in Hippocampal CA1 inMICD Rats To identify the role of synaptic-dendrite-relatedmiR-134 in hippocampus CA1 the levels were detected Asshown in Figure 7 the expression ofmiR-134was significantlyincreased in theMICD group comparedwith the Sham group(119901 lt 001) However repeated EA treatment significantlydecreased the expression of miR-134 compared with theMICD group and the MICD+non-EA group (119901 lt 005)
4 Discussion
Ischemic stroke leads to a high incidence of long-termcognitive impairment which is strongly associated with lossof hippocampal neurons and synapses [34] While review-ing ancient Chinese documents regarding acupuncture andcognitive impairment-related terms we discovered that theDU20 andDU24 acupoints were themost frequently selectedacupoints for cognitive impairment-related rehabilitation inChina A number of studies have shown that EA can improve
Neural Plasticity 7
MICD+non-EAMICD+EA
MICDSham
(a)
MICD+non-EA
MICD+EA
MICD
Sham
(b)
MICD+non-EA
MICD+EAMICDSham0
5
10
15
20
lowastlowastandand
Spin
e den
sity
(10
um)
(c)
Figure 4 The structures and densities of dendritic spine in the hippocampus of rats (a) Representative images of dendritic spine in thehippocampus of each group (Golgi staining times50) (b) Representative images of dendritic spine density and morphology from the pyramidalcell layer of hippocampal CA1 area in each group (Golgi staining times1000) (c)The density of dendritic spine was analyzed in hippocampal CA1pyramidal cells in each group (119899 = 6group lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD group andand119901 lt 001and119901 lt 005 versus the MICD+EA group) All experiments were repeated three times
learning and memory ability and stimulate consciousness[30 31 35]These results indicated that EA could be a comple-mentary therapy for cognitive impairment after stroke Thepresent study found that EA at theDU20 andDU24 acupointsshortened time to find the platform and increased the timesof crossing the position of the platform compared to untreat-ment or nonacupoint EA (stimulation control) in Morriswater maze test suggesting that EA could improve spatialreference learning and memory ability in MCAO-inducedcognitive deficit (MICD) rats Moreover it is worth mention-ing that the path length in the Morris water maze in the fourgroups showed no significant difference indicating that thetime spending ofMICD rats to find the objectwas not affectedby themotor functionHowever themechanisms of cognitivetreatment involved are far from being fully elucidated
Firstly the cognitive deficit-related brain regions weredetermined by a small animal MRI in rats with ischemicstroke T2-weighted imaging showed that MICD caused thelesions of left cortex hippocampus corpus striatum andthalamus regions before EA It had beennoticed that the brain
lesion exhibited amild spontaneous recovery performance inMICD rats However the lesion regions comprising approxi-mately 23of thewhole brain volumewere reduced to 15byEA treatment indicating that EA could attenuate cortex andhippocampus corpus striatum and thalamus region lesionsin theMICD rats Some of these regions such as the hippoca-mpus and cortex are essential for regulating learning andmemory behaviors including spatial exploration [36] Studieshave confirmed that the hippocampus regions play a veryimportant role in learning and memory through their spe-cific structure and location which connect with other brainregions together [37] Furthermore in order to identify thehippocampus function in cognitive deficit the hippocampalmorphology staining was observed
Using Golgi staining and electron microscopy we demo-nstrated that the density of dendritic spine and the number ofsynapses in hippocampal CA1 pyramidal cells were obviouslyreduced at Day 14 after MICD However EA can rescue theloss of dendritic spine and synapses in hippocampal CA1region Moreover EA promoted synaptic-dendritic spine
8 Neural Plasticity
MICD+non-EAMICD+EA
MICDSham
(a)
MICD+non-EA
MICD+EAMICDSham0
2
4
6
8
10
The n
umbe
r of s
ynap
ses (10cm
2)
lowastlowast andand
(b)
Figure 5The number of synapses in hippocampal CA1 pyramidal cells (a) Representative electronmicrographic images of synapses (arrows)in the CA1 region of each group (magnification times50000) (b) Histogram shows the significant difference in the mean plusmn SEM of the numberof synapses in each group (119899 = 5group lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD group andand119901 lt 001and119901 lt 005 versus the MICD+EA group)
MICD+non-EAMICD+EAMICD
120573-Actin
P-LIMK1
T-LIMK1
Sham
(a)
MICD+non-EA
MICD+EAMICDSham00
02
04
06
08
10
Tota
l LIM
K1120573
-act
in
lowastlowastandand
(b)
MICD+non-EA
MICD+EAMICDSham00
02
04
06
08
10
lowastlowastandand
P-LI
MK1
T-L
IMK1
(c)
Figure 6 EA amplified the LIMK1 level in the hippocampus CA1 pyramidal cells (a) The expression of total LIMK1 and phosphorylatedLIMK1was evaluated usingwestern blotting (b and c)Histogram shows significant difference of the levels of P-LIMK1 and phosphor-LIMLK1in each group Data are means plusmn SEM (119899 = 6group lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD groupandand119901 lt 001 and119901 lt 005 versus the MICD+EA group)
Neural Plasticity 9
0
2
4
6
8
10
12
miR
-134
relat
ive e
xpre
ssio
n to
U6 lowastlowast
and
MICD+non-EA
MICD+EAMICDSham
Figure 7 Changes of miR-134 expression in the hippocampusCA1 cells The changes of miR-134 expression level were measuredusing quantitative RT-PCR U6 was used as internal control forquantification of miRNAs Data are mean plusmn SEM (119899 = 6grouplowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus theMICD group andand119901 lt 001 and119901 lt 005 versus the MICD+EA group)
regeneration Dendritic spine is the tiny protrusions withmulti-dense body and ion channels on the surface of varioustypes of neurons which serve as cellular substrates of brainconnectivity and the major sites of information processingin the brain [38 39] Evidence is increasing that synaptic-dendritic plasticity enables the encoding of memory [40]Indeed themodification on dendritic spine number size andshape is of importance for the plasticity of synapses accom-panied by hippocampus-dependent learning and memory[41 42] Studies have well described that dendritic spineremodeling in ischemia surrounding areas may contribute tocognitive functional recovery after stroke [43 44]
Increasing evidence showed that LIMK1 protein is acti-vated at dendritic spine membranes [45] LIMK1 positivecells were located in the CA1 region of the hippocampuswhich is important in the regulation of the spinemorphologyand synaptic function in vivo A prominent form of longlasting synaptic plasticity is thought to be critical to memoryformation [26] The present study found that the expressionof total LIMK1 and phospho-LIMK1 in hippocampal CA1wassignificantly decreased in the MICD rats however EA couldenhance the total LIMK1 and phospho-LIMK1 levels to pro-mote synaptic-dendritic plasticity in hippocampal CA1 area
In addition we have identified a dendritically localizedmiRNA that regulated the expression of the synaptic LIMK1protein thereby controlling dendritic pine sizemiR-134 is thefirst discovered dendritic microRNA enriched in the neu-ronal dendrites of rat hippocampal neurons and negativelycontrols the size of dendritic spines [46] This effect ismediated by miR-134 inhibition of the translation of LIMK1mRNA [47] The present study showed that the expression ofmiR-134 was obviously increased in theMICD rats Howeverrepeated EA treatment could relieve the upregulation of miR-134 expression in hippocampal CA1 area
In conclusion the study indicated that EA at the DU20and DU24 acupoints could ameliorate cognitive impairmentof MICD rats through the regulation of synaptic-dendritic
plasticity of the hippocampal CA1 area and the expressionand phosphorylation of synaptic LIMK1 protein were acti-vated to control dendritic spine number size and shapeFurthermore the mechanism of LIMK1 increase induced byEA treatmentmay be associated withmiR-134 which is local-ized in hippocampal CA1 by negatively regulating LIMK1 toenhance synaptic-dendritic plasticity in the recovery stage ofischemic stroke
Competing Interests
The authors declare no financial or commercial conflict ofinterests
Authorsrsquo Contributions
Weilin Liu Jie Wu and Jia Huang contributed equally to thiswork
Acknowledgments
The present study was supported by the Natural ScienceFoundation of China (no 81403450) and the Natural ScienceFoundation of Fujian Province (no 2016J01382)
References
[1] V L Roger ldquoHeart disease and stroke statisticsmdash2011 update areport from the American Heart Associationrdquo Circulation vol131 no 4 pp 29ndash322 2014
[2] T E Nichols and A P Holmes ldquoNonparametric permutationtests for functional neuroimaging a primer with examplesrdquoHuman Brain Mapping vol 15 no 1 pp 1ndash25 2002
[3] T K Tatemichi D W Desmond Y Stern M Paik M Sanoand E Bagiella ldquoCognitive impairment after stroke frequencypatterns and relationship to functional abilitiesrdquo Journal ofNeurology Neurosurgery amp Psychiatry vol 57 no 2 pp 202ndash207 1994
[4] F Liu Z-M Li Y-J Jiang and L-D Chen ldquoA meta-analysis ofacupuncture use in the treatment of cognitive impairment afterstrokerdquo Journal of Alternative amp Complementary Medicine vol20 no 7 pp 535ndash544 2014
[5] W Li R Huang R A Shetty et al ldquoTransient focal cerebralischemia induces long-termcognitive function deficit in an exp-erimental ischemic stroke modelrdquo Neurobiology of Disease vol59 pp 18ndash25 2013
[6] P S Sachdev X Chen H Brodaty CThompson A Altendorfand W Wen ldquoThe determinants and longitudinal course ofpost-stroke mild cognitive impairmentrdquo Journal of the Inter-national Neuropsychological Society vol 15 no 6 pp 915ndash9232009
[7] A-J Liu J-H Li H-Q Li et al ldquoElectroacupuncture foracute ischemic stroke ameta-analysis of randomized controlledtrialsrdquoThe American Journal of Chinese Medicine vol 43 no 8pp 1541ndash1566 2015
[8] X Ding C-Y Li Q-S Wang et al ldquoPatterns in default-modenetwork connectivity for determining outcomes in cognitivefunction in acute stroke patientsrdquo Neuroscience vol 277 no 10pp 637ndash646 2014
10 Neural Plasticity
[9] H Alipanahzadeh M Soleimani S Soleimani Asl B Pourhey-dar A Nikkhah and M Mehdizadeh ldquoTransforming growthfactor-alpha improves memory impairment and neurogenesisfollowing ischemia reperfusionrdquo Cell Journal vol 16 no 3 pp315ndash324 2014
[10] H Eichenbaum ldquoHippocampus cognitive processes and neuralrepresentations that underlie declarative memoryrdquoNeuron vol44 no 1 pp 109ndash120 2004
[11] T Kadar S Dachir B Shukitt-Hale and A Levy ldquoSub-regionalhippocampal vulnerability in various animal models leading tocognitive dysfunctionrdquo Journal of Neural Transmission vol 105no 8-9 pp 987ndash1004 1998
[12] J B Pereira C Valls-Pedret E Ros et al ldquoRegional vulnera-bility of hippocampal subfields to aging measured by structuraland diffusion MRIrdquo Hippocampus vol 24 no 4 pp 403ndash4142014
[13] S Knafo G Ariav E Barkai and F Libersat ldquoOlfactory learn-ing-induced increase in spine density along the apical dendritesof CA1 hippocampal neuronsrdquo Hippocampus vol 14 no 7 pp819ndash825 2004
[14] S Schacher and J-Y Hu ldquoThe less things change the more theyare different contributions of long-term synaptic plasticity andhomeostasis to memoryrdquo Learning and Memory vol 21 no 3pp 128ndash134 2014
[15] K Park H Heo M E Han et al ldquoLearning-induced synapticpotentiation in implanted neural precursor cell-derived neu-ronsrdquo Scientific Reports vol 5 article 17796 2015
[16] S Knafo F Libersat and E Barkai ldquoDynamics of learning-induced spine redistribution along dendrites of pyramidalneurons in ratsrdquo European Journal of Neuroscience vol 21 no4 pp 927ndash935 2005
[17] V C Foletta N Moussi P D Sarmiere J R Bamburg and OBernard ldquoLIM kinase 1 a key regulator of actin dynamics iswidely expressed in embryonic and adult tissuesrdquo ExperimentalCell Research vol 294 no 2 pp 392ndash405 2004
[18] M Endo K Ohashi Y Sasaki et al ldquoControl of growth conemotility andmorphology by LIMkinase and slingshot via phos-phorylation and dephosphorylation of cofilinrdquo Journal of Neu-roscience vol 23 no 7 pp 2527ndash2537 2003
[19] C Jarrold A D Baddeley and C Phillips ldquoLong-termmemoryfor verbal and visual information in Down syndrome andWilliams syndrome performance on the doors and people testrdquoCortex vol 43 no 2 pp 233ndash247 2007
[20] Y Meng Y Zhang V Tregoubov et al ldquoAbnormal spinemorphology and enhanced LTP in LIMK-1 knockout micerdquoNeuron vol 35 no 1 pp 121ndash133 2002
[21] Z Todorovski S Asrar J Liu et al ldquoLIMK1 regulates long-termmemory and synaptic plasticity via the transcriptional factorCREBrdquo Molecular and Cellular Biology vol 35 no 8 pp 1316ndash1328 2015
[22] G M Schratt F Tuebing E A Nigh et al ldquoA brain-specificmicroRNA regulates dendritic spine developmentrdquo Nature vol439 no 7074 pp 283ndash289 2006
[23] F Capitano J Camon V Ferretti et al ldquomicroRNAs modulatespatial memory in the hippocampus and in the ventral striatumin a region-specific mannerrdquo Molecular Neurobiology vol 53no 7 pp 4618ndash4630 2015
[24] J GaoW-YWang Y-WMao et al ldquoA novel pathway regulatesmemory and plasticity via SIRT1 andmiR-134rdquoNature vol 466no 7310 pp 1105ndash1109 2010
[25] J Remenyi M W M van den Bosch O Palygin et al ldquomiR-132212 knockout mice reveal roles for these miRNAs in regu-lating cortical synaptic transmission and plasticityrdquo PLoS ONEvol 8 no 4 Article ID e62509 p 65 2013
[26] L An J LiuWW Li et al ldquoDistribution of LIM domain kinase1 in the olfactory bulb cerebral cortex hippocampus and cere-bellum of the AppPS+minus micerdquo Genetics amp Molecular Researchvol 14 no 4 pp 17244ndash17251 2015
[27] R Lin Y Wu J Tao et al ldquoElectroacupuncture improves cog-nitive function through Rho GTPases and enhances dendriticspine plasticity in rats with cerebral ischemia-reperfusionrdquoMolecular Medicine Reports vol 13 no 3 pp 2655ndash2660 2016
[28] P Castaneda M Munoz G Garcıa-Rojo et al ldquoAssociation ofN-cadherin levels and downstream effectors of Rho GTPaseswith dendritic spine loss induced by chronic stress in rathippocampal neuronsrdquo Journal of Neuroscience Research vol93 no 10 pp 1476ndash1491 2015
[29] A Romarowski M A Battistone F A La Spina et al ldquoPKA-dependent phosphorylation of LIMK1 and Cofilin is essentialformouse sperm acrosomal exocytosisrdquoDevelopmental Biologyvol 405 no 2 pp 237ndash249 2015
[30] R Lin K Yu X Li et al ldquoElectroacupuncture ameliorates post-stroke learning and memory through minimizing ultrastruc-tural brain damage and inhibiting the expression of MMP-2andMMP-9 in cerebral ischemia-reperfusion injured ratsrdquoMol-ecular Medicine Reports vol 14 no 1 pp 225ndash233 2016
[31] X Feng S Yang J Liu et al ldquoElectroacupuncture amelioratescognitive impairment through inhibition of NF-120581B-mediatedneuronal cell apoptosis in cerebral ischemia-reperfusion inj-ured ratsrdquo Molecular Medicine Reports vol 7 no 5 pp 1516ndash1522 2013
[32] Z K Wang N I Guang-Xia K Liu et al ldquoResearch on thechanges of IL-1 receptor and TNF-120572 receptor in rats with cere-bral ischemia reperfusion and the chronergy of acupunctureinterventionrdquo Zhongguo Zhen Jiu vol 32 no 11 pp 1012ndash10182012
[33] X S LiuM Chopp R L Zhang and Z G Zhang ldquoMicroRNAsin cerebral ischemia-induced neurogenesisrdquo Journal of Neu-ropathology amp Experimental Neurology vol 72 no 8 pp 718ndash722 2013
[34] X He T Yan R Chen and D Ran ldquoAcute effects of electro-acupuncture (EA) on hippocampal long term potentiation(LTP) of perforant path-dentate gyrus granule cells synapserelated to memoryrdquo Acupuncture and Electro-TherapeuticsResearch vol 37 no 2-3 pp 89ndash101 2012
[35] W Dong W Guo X Zheng et al ldquoElectroacupunctureimproves cognitive deficits associated with AMPK activation inSAMP8 micerdquo Metabolic Brain Disease vol 30 no 3 pp 777ndash784 2015
[36] B Milner and D Klein ldquoLoss of recent memory after bilateralhippocampal lesions memory and memoriesmdashlooking backand looking forwardrdquo Journal of Neurology Neurosurgery ampPsychiatry vol 87 no 3 article 230 2016
[37] A Veronica Witte L Kerti D S Margulies and A FloelldquoEffects of resveratrol on memory performance hippocampalfunctional connectivity and glucose metabolism in healthyolder adultsrdquo Journal of Neuroscience vol 34 no 23 pp 7862ndash7870 2014
[38] T Bonhoeffer and R Yuste ldquoSpine motility phenomenologymechanisms and functionrdquoNeuron vol 35 no 6 pp 1019ndash10272002
Neural Plasticity 11
[39] H Qiao M Li C Xu H Chen S An and X Ma ldquoDendriticspines in depression what we learned from animal modelsrdquoNeural Plasticity vol 2016 Article ID 8056370 26 pages 2016
[40] S C McQuown R M Barrett D P Matheos et al ldquoHDAC3 isa critical negative regulator of long-term memory formationrdquoJournal of Neuroscience vol 31 no 2 pp 764ndash774 2011
[41] K-O Lai and N Y Ip ldquoStructural plasticity of dendritic spinesthe underlying mechanisms and its dysregulation in brain dis-ordersrdquo Biochimica et Biophysica ActamdashMolecular Basis of Dis-ease vol 1832 no 12 pp 2257ndash2263 2013
[42] C H Bailey E R Kandel and K M Harris ldquoStructural com-ponents of synaptic plasticity and memory consolidationrdquo ColdSpring Harbor Perspectives in Biology vol 7 no 7 Article IDa021758 2015
[43] C E Brown P Li J D Boyd K R Delaney and T HMurphy ldquoExtensive turnover of dendritic spines and vascularremodeling in cortical tissues recovering from strokerdquo Journalof Neuroscience vol 27 no 15 pp 4101ndash4109 2007
[44] R Mostany T G Chowdhury D G Johnston S A PortonovoS T Carmichael and C Portera-Cailliau ldquoLocal hemodynam-ics dictate long-term dendritic plasticity in peri-infarct cortexrdquoJournal of Neuroscience vol 30 no 42 pp 14116ndash14126 2010
[45] J George C Soares A Montersino J-C Beique and G MThomas ldquoPalmitoylation of LIM kinase-1 ensures spine-specificactin polymerization and morphological plasticityrdquo eLife vol2015 no 4 Article ID e06327 2015
[46] S Bicker M Lackinger K Weiszlig and G Schratt ldquoMicroRNA-132 -134 and -138 a microRNA troika rules in neuronaldendritesrdquo Cellular and molecular life sciences CMLS vol 71no 20 pp 3987ndash4005 2014
[47] H-C Tai and E M Schuman ldquoMicroRNA microRNAs reachout into dendritesrdquoCurrent Biology vol 16 no 4 pp R121ndashR1232006
Submit your manuscripts athttpswwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

Neural Plasticity 7
MICD+non-EAMICD+EA
MICDSham
(a)
MICD+non-EA
MICD+EA
MICD
Sham
(b)
MICD+non-EA
MICD+EAMICDSham0
5
10
15
20
lowastlowastandand
Spin
e den
sity
(10
um)
(c)
Figure 4 The structures and densities of dendritic spine in the hippocampus of rats (a) Representative images of dendritic spine in thehippocampus of each group (Golgi staining times50) (b) Representative images of dendritic spine density and morphology from the pyramidalcell layer of hippocampal CA1 area in each group (Golgi staining times1000) (c)The density of dendritic spine was analyzed in hippocampal CA1pyramidal cells in each group (119899 = 6group lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD group andand119901 lt 001and119901 lt 005 versus the MICD+EA group) All experiments were repeated three times
learning and memory ability and stimulate consciousness[30 31 35]These results indicated that EA could be a comple-mentary therapy for cognitive impairment after stroke Thepresent study found that EA at theDU20 andDU24 acupointsshortened time to find the platform and increased the timesof crossing the position of the platform compared to untreat-ment or nonacupoint EA (stimulation control) in Morriswater maze test suggesting that EA could improve spatialreference learning and memory ability in MCAO-inducedcognitive deficit (MICD) rats Moreover it is worth mention-ing that the path length in the Morris water maze in the fourgroups showed no significant difference indicating that thetime spending ofMICD rats to find the objectwas not affectedby themotor functionHowever themechanisms of cognitivetreatment involved are far from being fully elucidated
Firstly the cognitive deficit-related brain regions weredetermined by a small animal MRI in rats with ischemicstroke T2-weighted imaging showed that MICD caused thelesions of left cortex hippocampus corpus striatum andthalamus regions before EA It had beennoticed that the brain
lesion exhibited amild spontaneous recovery performance inMICD rats However the lesion regions comprising approxi-mately 23of thewhole brain volumewere reduced to 15byEA treatment indicating that EA could attenuate cortex andhippocampus corpus striatum and thalamus region lesionsin theMICD rats Some of these regions such as the hippoca-mpus and cortex are essential for regulating learning andmemory behaviors including spatial exploration [36] Studieshave confirmed that the hippocampus regions play a veryimportant role in learning and memory through their spe-cific structure and location which connect with other brainregions together [37] Furthermore in order to identify thehippocampus function in cognitive deficit the hippocampalmorphology staining was observed
Using Golgi staining and electron microscopy we demo-nstrated that the density of dendritic spine and the number ofsynapses in hippocampal CA1 pyramidal cells were obviouslyreduced at Day 14 after MICD However EA can rescue theloss of dendritic spine and synapses in hippocampal CA1region Moreover EA promoted synaptic-dendritic spine
8 Neural Plasticity
MICD+non-EAMICD+EA
MICDSham
(a)
MICD+non-EA
MICD+EAMICDSham0
2
4
6
8
10
The n
umbe
r of s
ynap
ses (10cm
2)
lowastlowast andand
(b)
Figure 5The number of synapses in hippocampal CA1 pyramidal cells (a) Representative electronmicrographic images of synapses (arrows)in the CA1 region of each group (magnification times50000) (b) Histogram shows the significant difference in the mean plusmn SEM of the numberof synapses in each group (119899 = 5group lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD group andand119901 lt 001and119901 lt 005 versus the MICD+EA group)
MICD+non-EAMICD+EAMICD
120573-Actin
P-LIMK1
T-LIMK1
Sham
(a)
MICD+non-EA
MICD+EAMICDSham00
02
04
06
08
10
Tota
l LIM
K1120573
-act
in
lowastlowastandand
(b)
MICD+non-EA
MICD+EAMICDSham00
02
04
06
08
10
lowastlowastandand
P-LI
MK1
T-L
IMK1
(c)
Figure 6 EA amplified the LIMK1 level in the hippocampus CA1 pyramidal cells (a) The expression of total LIMK1 and phosphorylatedLIMK1was evaluated usingwestern blotting (b and c)Histogram shows significant difference of the levels of P-LIMK1 and phosphor-LIMLK1in each group Data are means plusmn SEM (119899 = 6group lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD groupandand119901 lt 001 and119901 lt 005 versus the MICD+EA group)
Neural Plasticity 9
0
2
4
6
8
10
12
miR
-134
relat
ive e
xpre
ssio
n to
U6 lowastlowast
and
MICD+non-EA
MICD+EAMICDSham
Figure 7 Changes of miR-134 expression in the hippocampusCA1 cells The changes of miR-134 expression level were measuredusing quantitative RT-PCR U6 was used as internal control forquantification of miRNAs Data are mean plusmn SEM (119899 = 6grouplowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus theMICD group andand119901 lt 001 and119901 lt 005 versus the MICD+EA group)
regeneration Dendritic spine is the tiny protrusions withmulti-dense body and ion channels on the surface of varioustypes of neurons which serve as cellular substrates of brainconnectivity and the major sites of information processingin the brain [38 39] Evidence is increasing that synaptic-dendritic plasticity enables the encoding of memory [40]Indeed themodification on dendritic spine number size andshape is of importance for the plasticity of synapses accom-panied by hippocampus-dependent learning and memory[41 42] Studies have well described that dendritic spineremodeling in ischemia surrounding areas may contribute tocognitive functional recovery after stroke [43 44]
Increasing evidence showed that LIMK1 protein is acti-vated at dendritic spine membranes [45] LIMK1 positivecells were located in the CA1 region of the hippocampuswhich is important in the regulation of the spinemorphologyand synaptic function in vivo A prominent form of longlasting synaptic plasticity is thought to be critical to memoryformation [26] The present study found that the expressionof total LIMK1 and phospho-LIMK1 in hippocampal CA1wassignificantly decreased in the MICD rats however EA couldenhance the total LIMK1 and phospho-LIMK1 levels to pro-mote synaptic-dendritic plasticity in hippocampal CA1 area
In addition we have identified a dendritically localizedmiRNA that regulated the expression of the synaptic LIMK1protein thereby controlling dendritic pine sizemiR-134 is thefirst discovered dendritic microRNA enriched in the neu-ronal dendrites of rat hippocampal neurons and negativelycontrols the size of dendritic spines [46] This effect ismediated by miR-134 inhibition of the translation of LIMK1mRNA [47] The present study showed that the expression ofmiR-134 was obviously increased in theMICD rats Howeverrepeated EA treatment could relieve the upregulation of miR-134 expression in hippocampal CA1 area
In conclusion the study indicated that EA at the DU20and DU24 acupoints could ameliorate cognitive impairmentof MICD rats through the regulation of synaptic-dendritic
plasticity of the hippocampal CA1 area and the expressionand phosphorylation of synaptic LIMK1 protein were acti-vated to control dendritic spine number size and shapeFurthermore the mechanism of LIMK1 increase induced byEA treatmentmay be associated withmiR-134 which is local-ized in hippocampal CA1 by negatively regulating LIMK1 toenhance synaptic-dendritic plasticity in the recovery stage ofischemic stroke
Competing Interests
The authors declare no financial or commercial conflict ofinterests
Authorsrsquo Contributions
Weilin Liu Jie Wu and Jia Huang contributed equally to thiswork
Acknowledgments
The present study was supported by the Natural ScienceFoundation of China (no 81403450) and the Natural ScienceFoundation of Fujian Province (no 2016J01382)
References
[1] V L Roger ldquoHeart disease and stroke statisticsmdash2011 update areport from the American Heart Associationrdquo Circulation vol131 no 4 pp 29ndash322 2014
[2] T E Nichols and A P Holmes ldquoNonparametric permutationtests for functional neuroimaging a primer with examplesrdquoHuman Brain Mapping vol 15 no 1 pp 1ndash25 2002
[3] T K Tatemichi D W Desmond Y Stern M Paik M Sanoand E Bagiella ldquoCognitive impairment after stroke frequencypatterns and relationship to functional abilitiesrdquo Journal ofNeurology Neurosurgery amp Psychiatry vol 57 no 2 pp 202ndash207 1994
[4] F Liu Z-M Li Y-J Jiang and L-D Chen ldquoA meta-analysis ofacupuncture use in the treatment of cognitive impairment afterstrokerdquo Journal of Alternative amp Complementary Medicine vol20 no 7 pp 535ndash544 2014
[5] W Li R Huang R A Shetty et al ldquoTransient focal cerebralischemia induces long-termcognitive function deficit in an exp-erimental ischemic stroke modelrdquo Neurobiology of Disease vol59 pp 18ndash25 2013
[6] P S Sachdev X Chen H Brodaty CThompson A Altendorfand W Wen ldquoThe determinants and longitudinal course ofpost-stroke mild cognitive impairmentrdquo Journal of the Inter-national Neuropsychological Society vol 15 no 6 pp 915ndash9232009
[7] A-J Liu J-H Li H-Q Li et al ldquoElectroacupuncture foracute ischemic stroke ameta-analysis of randomized controlledtrialsrdquoThe American Journal of Chinese Medicine vol 43 no 8pp 1541ndash1566 2015
[8] X Ding C-Y Li Q-S Wang et al ldquoPatterns in default-modenetwork connectivity for determining outcomes in cognitivefunction in acute stroke patientsrdquo Neuroscience vol 277 no 10pp 637ndash646 2014
10 Neural Plasticity
[9] H Alipanahzadeh M Soleimani S Soleimani Asl B Pourhey-dar A Nikkhah and M Mehdizadeh ldquoTransforming growthfactor-alpha improves memory impairment and neurogenesisfollowing ischemia reperfusionrdquo Cell Journal vol 16 no 3 pp315ndash324 2014
[10] H Eichenbaum ldquoHippocampus cognitive processes and neuralrepresentations that underlie declarative memoryrdquoNeuron vol44 no 1 pp 109ndash120 2004
[11] T Kadar S Dachir B Shukitt-Hale and A Levy ldquoSub-regionalhippocampal vulnerability in various animal models leading tocognitive dysfunctionrdquo Journal of Neural Transmission vol 105no 8-9 pp 987ndash1004 1998
[12] J B Pereira C Valls-Pedret E Ros et al ldquoRegional vulnera-bility of hippocampal subfields to aging measured by structuraland diffusion MRIrdquo Hippocampus vol 24 no 4 pp 403ndash4142014
[13] S Knafo G Ariav E Barkai and F Libersat ldquoOlfactory learn-ing-induced increase in spine density along the apical dendritesof CA1 hippocampal neuronsrdquo Hippocampus vol 14 no 7 pp819ndash825 2004
[14] S Schacher and J-Y Hu ldquoThe less things change the more theyare different contributions of long-term synaptic plasticity andhomeostasis to memoryrdquo Learning and Memory vol 21 no 3pp 128ndash134 2014
[15] K Park H Heo M E Han et al ldquoLearning-induced synapticpotentiation in implanted neural precursor cell-derived neu-ronsrdquo Scientific Reports vol 5 article 17796 2015
[16] S Knafo F Libersat and E Barkai ldquoDynamics of learning-induced spine redistribution along dendrites of pyramidalneurons in ratsrdquo European Journal of Neuroscience vol 21 no4 pp 927ndash935 2005
[17] V C Foletta N Moussi P D Sarmiere J R Bamburg and OBernard ldquoLIM kinase 1 a key regulator of actin dynamics iswidely expressed in embryonic and adult tissuesrdquo ExperimentalCell Research vol 294 no 2 pp 392ndash405 2004
[18] M Endo K Ohashi Y Sasaki et al ldquoControl of growth conemotility andmorphology by LIMkinase and slingshot via phos-phorylation and dephosphorylation of cofilinrdquo Journal of Neu-roscience vol 23 no 7 pp 2527ndash2537 2003
[19] C Jarrold A D Baddeley and C Phillips ldquoLong-termmemoryfor verbal and visual information in Down syndrome andWilliams syndrome performance on the doors and people testrdquoCortex vol 43 no 2 pp 233ndash247 2007
[20] Y Meng Y Zhang V Tregoubov et al ldquoAbnormal spinemorphology and enhanced LTP in LIMK-1 knockout micerdquoNeuron vol 35 no 1 pp 121ndash133 2002
[21] Z Todorovski S Asrar J Liu et al ldquoLIMK1 regulates long-termmemory and synaptic plasticity via the transcriptional factorCREBrdquo Molecular and Cellular Biology vol 35 no 8 pp 1316ndash1328 2015
[22] G M Schratt F Tuebing E A Nigh et al ldquoA brain-specificmicroRNA regulates dendritic spine developmentrdquo Nature vol439 no 7074 pp 283ndash289 2006
[23] F Capitano J Camon V Ferretti et al ldquomicroRNAs modulatespatial memory in the hippocampus and in the ventral striatumin a region-specific mannerrdquo Molecular Neurobiology vol 53no 7 pp 4618ndash4630 2015
[24] J GaoW-YWang Y-WMao et al ldquoA novel pathway regulatesmemory and plasticity via SIRT1 andmiR-134rdquoNature vol 466no 7310 pp 1105ndash1109 2010
[25] J Remenyi M W M van den Bosch O Palygin et al ldquomiR-132212 knockout mice reveal roles for these miRNAs in regu-lating cortical synaptic transmission and plasticityrdquo PLoS ONEvol 8 no 4 Article ID e62509 p 65 2013
[26] L An J LiuWW Li et al ldquoDistribution of LIM domain kinase1 in the olfactory bulb cerebral cortex hippocampus and cere-bellum of the AppPS+minus micerdquo Genetics amp Molecular Researchvol 14 no 4 pp 17244ndash17251 2015
[27] R Lin Y Wu J Tao et al ldquoElectroacupuncture improves cog-nitive function through Rho GTPases and enhances dendriticspine plasticity in rats with cerebral ischemia-reperfusionrdquoMolecular Medicine Reports vol 13 no 3 pp 2655ndash2660 2016
[28] P Castaneda M Munoz G Garcıa-Rojo et al ldquoAssociation ofN-cadherin levels and downstream effectors of Rho GTPaseswith dendritic spine loss induced by chronic stress in rathippocampal neuronsrdquo Journal of Neuroscience Research vol93 no 10 pp 1476ndash1491 2015
[29] A Romarowski M A Battistone F A La Spina et al ldquoPKA-dependent phosphorylation of LIMK1 and Cofilin is essentialformouse sperm acrosomal exocytosisrdquoDevelopmental Biologyvol 405 no 2 pp 237ndash249 2015
[30] R Lin K Yu X Li et al ldquoElectroacupuncture ameliorates post-stroke learning and memory through minimizing ultrastruc-tural brain damage and inhibiting the expression of MMP-2andMMP-9 in cerebral ischemia-reperfusion injured ratsrdquoMol-ecular Medicine Reports vol 14 no 1 pp 225ndash233 2016
[31] X Feng S Yang J Liu et al ldquoElectroacupuncture amelioratescognitive impairment through inhibition of NF-120581B-mediatedneuronal cell apoptosis in cerebral ischemia-reperfusion inj-ured ratsrdquo Molecular Medicine Reports vol 7 no 5 pp 1516ndash1522 2013
[32] Z K Wang N I Guang-Xia K Liu et al ldquoResearch on thechanges of IL-1 receptor and TNF-120572 receptor in rats with cere-bral ischemia reperfusion and the chronergy of acupunctureinterventionrdquo Zhongguo Zhen Jiu vol 32 no 11 pp 1012ndash10182012
[33] X S LiuM Chopp R L Zhang and Z G Zhang ldquoMicroRNAsin cerebral ischemia-induced neurogenesisrdquo Journal of Neu-ropathology amp Experimental Neurology vol 72 no 8 pp 718ndash722 2013
[34] X He T Yan R Chen and D Ran ldquoAcute effects of electro-acupuncture (EA) on hippocampal long term potentiation(LTP) of perforant path-dentate gyrus granule cells synapserelated to memoryrdquo Acupuncture and Electro-TherapeuticsResearch vol 37 no 2-3 pp 89ndash101 2012
[35] W Dong W Guo X Zheng et al ldquoElectroacupunctureimproves cognitive deficits associated with AMPK activation inSAMP8 micerdquo Metabolic Brain Disease vol 30 no 3 pp 777ndash784 2015
[36] B Milner and D Klein ldquoLoss of recent memory after bilateralhippocampal lesions memory and memoriesmdashlooking backand looking forwardrdquo Journal of Neurology Neurosurgery ampPsychiatry vol 87 no 3 article 230 2016
[37] A Veronica Witte L Kerti D S Margulies and A FloelldquoEffects of resveratrol on memory performance hippocampalfunctional connectivity and glucose metabolism in healthyolder adultsrdquo Journal of Neuroscience vol 34 no 23 pp 7862ndash7870 2014
[38] T Bonhoeffer and R Yuste ldquoSpine motility phenomenologymechanisms and functionrdquoNeuron vol 35 no 6 pp 1019ndash10272002
Neural Plasticity 11
[39] H Qiao M Li C Xu H Chen S An and X Ma ldquoDendriticspines in depression what we learned from animal modelsrdquoNeural Plasticity vol 2016 Article ID 8056370 26 pages 2016
[40] S C McQuown R M Barrett D P Matheos et al ldquoHDAC3 isa critical negative regulator of long-term memory formationrdquoJournal of Neuroscience vol 31 no 2 pp 764ndash774 2011
[41] K-O Lai and N Y Ip ldquoStructural plasticity of dendritic spinesthe underlying mechanisms and its dysregulation in brain dis-ordersrdquo Biochimica et Biophysica ActamdashMolecular Basis of Dis-ease vol 1832 no 12 pp 2257ndash2263 2013
[42] C H Bailey E R Kandel and K M Harris ldquoStructural com-ponents of synaptic plasticity and memory consolidationrdquo ColdSpring Harbor Perspectives in Biology vol 7 no 7 Article IDa021758 2015
[43] C E Brown P Li J D Boyd K R Delaney and T HMurphy ldquoExtensive turnover of dendritic spines and vascularremodeling in cortical tissues recovering from strokerdquo Journalof Neuroscience vol 27 no 15 pp 4101ndash4109 2007
[44] R Mostany T G Chowdhury D G Johnston S A PortonovoS T Carmichael and C Portera-Cailliau ldquoLocal hemodynam-ics dictate long-term dendritic plasticity in peri-infarct cortexrdquoJournal of Neuroscience vol 30 no 42 pp 14116ndash14126 2010
[45] J George C Soares A Montersino J-C Beique and G MThomas ldquoPalmitoylation of LIM kinase-1 ensures spine-specificactin polymerization and morphological plasticityrdquo eLife vol2015 no 4 Article ID e06327 2015
[46] S Bicker M Lackinger K Weiszlig and G Schratt ldquoMicroRNA-132 -134 and -138 a microRNA troika rules in neuronaldendritesrdquo Cellular and molecular life sciences CMLS vol 71no 20 pp 3987ndash4005 2014
[47] H-C Tai and E M Schuman ldquoMicroRNA microRNAs reachout into dendritesrdquoCurrent Biology vol 16 no 4 pp R121ndashR1232006
Submit your manuscripts athttpswwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
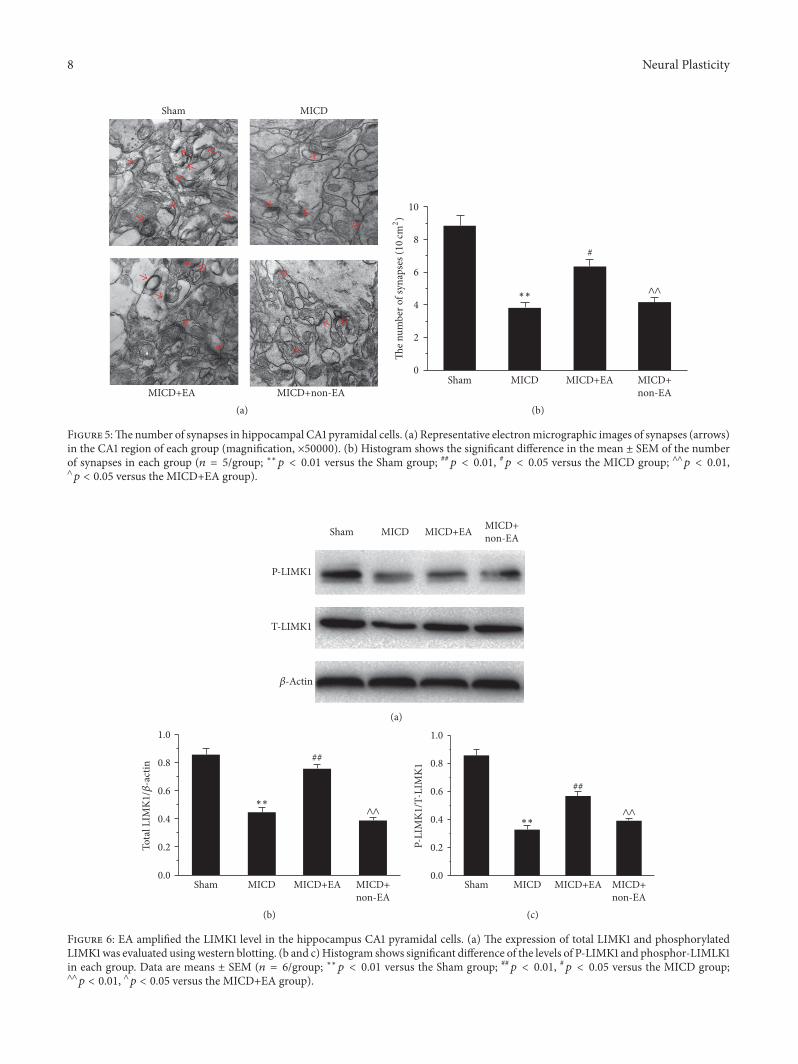
8 Neural Plasticity
MICD+non-EAMICD+EA
MICDSham
(a)
MICD+non-EA
MICD+EAMICDSham0
2
4
6
8
10
The n
umbe
r of s
ynap
ses (10cm
2)
lowastlowast andand
(b)
Figure 5The number of synapses in hippocampal CA1 pyramidal cells (a) Representative electronmicrographic images of synapses (arrows)in the CA1 region of each group (magnification times50000) (b) Histogram shows the significant difference in the mean plusmn SEM of the numberof synapses in each group (119899 = 5group lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD group andand119901 lt 001and119901 lt 005 versus the MICD+EA group)
MICD+non-EAMICD+EAMICD
120573-Actin
P-LIMK1
T-LIMK1
Sham
(a)
MICD+non-EA
MICD+EAMICDSham00
02
04
06
08
10
Tota
l LIM
K1120573
-act
in
lowastlowastandand
(b)
MICD+non-EA
MICD+EAMICDSham00
02
04
06
08
10
lowastlowastandand
P-LI
MK1
T-L
IMK1
(c)
Figure 6 EA amplified the LIMK1 level in the hippocampus CA1 pyramidal cells (a) The expression of total LIMK1 and phosphorylatedLIMK1was evaluated usingwestern blotting (b and c)Histogram shows significant difference of the levels of P-LIMK1 and phosphor-LIMLK1in each group Data are means plusmn SEM (119899 = 6group lowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus the MICD groupandand119901 lt 001 and119901 lt 005 versus the MICD+EA group)
Neural Plasticity 9
0
2
4
6
8
10
12
miR
-134
relat
ive e
xpre
ssio
n to
U6 lowastlowast
and
MICD+non-EA
MICD+EAMICDSham
Figure 7 Changes of miR-134 expression in the hippocampusCA1 cells The changes of miR-134 expression level were measuredusing quantitative RT-PCR U6 was used as internal control forquantification of miRNAs Data are mean plusmn SEM (119899 = 6grouplowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus theMICD group andand119901 lt 001 and119901 lt 005 versus the MICD+EA group)
regeneration Dendritic spine is the tiny protrusions withmulti-dense body and ion channels on the surface of varioustypes of neurons which serve as cellular substrates of brainconnectivity and the major sites of information processingin the brain [38 39] Evidence is increasing that synaptic-dendritic plasticity enables the encoding of memory [40]Indeed themodification on dendritic spine number size andshape is of importance for the plasticity of synapses accom-panied by hippocampus-dependent learning and memory[41 42] Studies have well described that dendritic spineremodeling in ischemia surrounding areas may contribute tocognitive functional recovery after stroke [43 44]
Increasing evidence showed that LIMK1 protein is acti-vated at dendritic spine membranes [45] LIMK1 positivecells were located in the CA1 region of the hippocampuswhich is important in the regulation of the spinemorphologyand synaptic function in vivo A prominent form of longlasting synaptic plasticity is thought to be critical to memoryformation [26] The present study found that the expressionof total LIMK1 and phospho-LIMK1 in hippocampal CA1wassignificantly decreased in the MICD rats however EA couldenhance the total LIMK1 and phospho-LIMK1 levels to pro-mote synaptic-dendritic plasticity in hippocampal CA1 area
In addition we have identified a dendritically localizedmiRNA that regulated the expression of the synaptic LIMK1protein thereby controlling dendritic pine sizemiR-134 is thefirst discovered dendritic microRNA enriched in the neu-ronal dendrites of rat hippocampal neurons and negativelycontrols the size of dendritic spines [46] This effect ismediated by miR-134 inhibition of the translation of LIMK1mRNA [47] The present study showed that the expression ofmiR-134 was obviously increased in theMICD rats Howeverrepeated EA treatment could relieve the upregulation of miR-134 expression in hippocampal CA1 area
In conclusion the study indicated that EA at the DU20and DU24 acupoints could ameliorate cognitive impairmentof MICD rats through the regulation of synaptic-dendritic
plasticity of the hippocampal CA1 area and the expressionand phosphorylation of synaptic LIMK1 protein were acti-vated to control dendritic spine number size and shapeFurthermore the mechanism of LIMK1 increase induced byEA treatmentmay be associated withmiR-134 which is local-ized in hippocampal CA1 by negatively regulating LIMK1 toenhance synaptic-dendritic plasticity in the recovery stage ofischemic stroke
Competing Interests
The authors declare no financial or commercial conflict ofinterests
Authorsrsquo Contributions
Weilin Liu Jie Wu and Jia Huang contributed equally to thiswork
Acknowledgments
The present study was supported by the Natural ScienceFoundation of China (no 81403450) and the Natural ScienceFoundation of Fujian Province (no 2016J01382)
References
[1] V L Roger ldquoHeart disease and stroke statisticsmdash2011 update areport from the American Heart Associationrdquo Circulation vol131 no 4 pp 29ndash322 2014
[2] T E Nichols and A P Holmes ldquoNonparametric permutationtests for functional neuroimaging a primer with examplesrdquoHuman Brain Mapping vol 15 no 1 pp 1ndash25 2002
[3] T K Tatemichi D W Desmond Y Stern M Paik M Sanoand E Bagiella ldquoCognitive impairment after stroke frequencypatterns and relationship to functional abilitiesrdquo Journal ofNeurology Neurosurgery amp Psychiatry vol 57 no 2 pp 202ndash207 1994
[4] F Liu Z-M Li Y-J Jiang and L-D Chen ldquoA meta-analysis ofacupuncture use in the treatment of cognitive impairment afterstrokerdquo Journal of Alternative amp Complementary Medicine vol20 no 7 pp 535ndash544 2014
[5] W Li R Huang R A Shetty et al ldquoTransient focal cerebralischemia induces long-termcognitive function deficit in an exp-erimental ischemic stroke modelrdquo Neurobiology of Disease vol59 pp 18ndash25 2013
[6] P S Sachdev X Chen H Brodaty CThompson A Altendorfand W Wen ldquoThe determinants and longitudinal course ofpost-stroke mild cognitive impairmentrdquo Journal of the Inter-national Neuropsychological Society vol 15 no 6 pp 915ndash9232009
[7] A-J Liu J-H Li H-Q Li et al ldquoElectroacupuncture foracute ischemic stroke ameta-analysis of randomized controlledtrialsrdquoThe American Journal of Chinese Medicine vol 43 no 8pp 1541ndash1566 2015
[8] X Ding C-Y Li Q-S Wang et al ldquoPatterns in default-modenetwork connectivity for determining outcomes in cognitivefunction in acute stroke patientsrdquo Neuroscience vol 277 no 10pp 637ndash646 2014
10 Neural Plasticity
[9] H Alipanahzadeh M Soleimani S Soleimani Asl B Pourhey-dar A Nikkhah and M Mehdizadeh ldquoTransforming growthfactor-alpha improves memory impairment and neurogenesisfollowing ischemia reperfusionrdquo Cell Journal vol 16 no 3 pp315ndash324 2014
[10] H Eichenbaum ldquoHippocampus cognitive processes and neuralrepresentations that underlie declarative memoryrdquoNeuron vol44 no 1 pp 109ndash120 2004
[11] T Kadar S Dachir B Shukitt-Hale and A Levy ldquoSub-regionalhippocampal vulnerability in various animal models leading tocognitive dysfunctionrdquo Journal of Neural Transmission vol 105no 8-9 pp 987ndash1004 1998
[12] J B Pereira C Valls-Pedret E Ros et al ldquoRegional vulnera-bility of hippocampal subfields to aging measured by structuraland diffusion MRIrdquo Hippocampus vol 24 no 4 pp 403ndash4142014
[13] S Knafo G Ariav E Barkai and F Libersat ldquoOlfactory learn-ing-induced increase in spine density along the apical dendritesof CA1 hippocampal neuronsrdquo Hippocampus vol 14 no 7 pp819ndash825 2004
[14] S Schacher and J-Y Hu ldquoThe less things change the more theyare different contributions of long-term synaptic plasticity andhomeostasis to memoryrdquo Learning and Memory vol 21 no 3pp 128ndash134 2014
[15] K Park H Heo M E Han et al ldquoLearning-induced synapticpotentiation in implanted neural precursor cell-derived neu-ronsrdquo Scientific Reports vol 5 article 17796 2015
[16] S Knafo F Libersat and E Barkai ldquoDynamics of learning-induced spine redistribution along dendrites of pyramidalneurons in ratsrdquo European Journal of Neuroscience vol 21 no4 pp 927ndash935 2005
[17] V C Foletta N Moussi P D Sarmiere J R Bamburg and OBernard ldquoLIM kinase 1 a key regulator of actin dynamics iswidely expressed in embryonic and adult tissuesrdquo ExperimentalCell Research vol 294 no 2 pp 392ndash405 2004
[18] M Endo K Ohashi Y Sasaki et al ldquoControl of growth conemotility andmorphology by LIMkinase and slingshot via phos-phorylation and dephosphorylation of cofilinrdquo Journal of Neu-roscience vol 23 no 7 pp 2527ndash2537 2003
[19] C Jarrold A D Baddeley and C Phillips ldquoLong-termmemoryfor verbal and visual information in Down syndrome andWilliams syndrome performance on the doors and people testrdquoCortex vol 43 no 2 pp 233ndash247 2007
[20] Y Meng Y Zhang V Tregoubov et al ldquoAbnormal spinemorphology and enhanced LTP in LIMK-1 knockout micerdquoNeuron vol 35 no 1 pp 121ndash133 2002
[21] Z Todorovski S Asrar J Liu et al ldquoLIMK1 regulates long-termmemory and synaptic plasticity via the transcriptional factorCREBrdquo Molecular and Cellular Biology vol 35 no 8 pp 1316ndash1328 2015
[22] G M Schratt F Tuebing E A Nigh et al ldquoA brain-specificmicroRNA regulates dendritic spine developmentrdquo Nature vol439 no 7074 pp 283ndash289 2006
[23] F Capitano J Camon V Ferretti et al ldquomicroRNAs modulatespatial memory in the hippocampus and in the ventral striatumin a region-specific mannerrdquo Molecular Neurobiology vol 53no 7 pp 4618ndash4630 2015
[24] J GaoW-YWang Y-WMao et al ldquoA novel pathway regulatesmemory and plasticity via SIRT1 andmiR-134rdquoNature vol 466no 7310 pp 1105ndash1109 2010
[25] J Remenyi M W M van den Bosch O Palygin et al ldquomiR-132212 knockout mice reveal roles for these miRNAs in regu-lating cortical synaptic transmission and plasticityrdquo PLoS ONEvol 8 no 4 Article ID e62509 p 65 2013
[26] L An J LiuWW Li et al ldquoDistribution of LIM domain kinase1 in the olfactory bulb cerebral cortex hippocampus and cere-bellum of the AppPS+minus micerdquo Genetics amp Molecular Researchvol 14 no 4 pp 17244ndash17251 2015
[27] R Lin Y Wu J Tao et al ldquoElectroacupuncture improves cog-nitive function through Rho GTPases and enhances dendriticspine plasticity in rats with cerebral ischemia-reperfusionrdquoMolecular Medicine Reports vol 13 no 3 pp 2655ndash2660 2016
[28] P Castaneda M Munoz G Garcıa-Rojo et al ldquoAssociation ofN-cadherin levels and downstream effectors of Rho GTPaseswith dendritic spine loss induced by chronic stress in rathippocampal neuronsrdquo Journal of Neuroscience Research vol93 no 10 pp 1476ndash1491 2015
[29] A Romarowski M A Battistone F A La Spina et al ldquoPKA-dependent phosphorylation of LIMK1 and Cofilin is essentialformouse sperm acrosomal exocytosisrdquoDevelopmental Biologyvol 405 no 2 pp 237ndash249 2015
[30] R Lin K Yu X Li et al ldquoElectroacupuncture ameliorates post-stroke learning and memory through minimizing ultrastruc-tural brain damage and inhibiting the expression of MMP-2andMMP-9 in cerebral ischemia-reperfusion injured ratsrdquoMol-ecular Medicine Reports vol 14 no 1 pp 225ndash233 2016
[31] X Feng S Yang J Liu et al ldquoElectroacupuncture amelioratescognitive impairment through inhibition of NF-120581B-mediatedneuronal cell apoptosis in cerebral ischemia-reperfusion inj-ured ratsrdquo Molecular Medicine Reports vol 7 no 5 pp 1516ndash1522 2013
[32] Z K Wang N I Guang-Xia K Liu et al ldquoResearch on thechanges of IL-1 receptor and TNF-120572 receptor in rats with cere-bral ischemia reperfusion and the chronergy of acupunctureinterventionrdquo Zhongguo Zhen Jiu vol 32 no 11 pp 1012ndash10182012
[33] X S LiuM Chopp R L Zhang and Z G Zhang ldquoMicroRNAsin cerebral ischemia-induced neurogenesisrdquo Journal of Neu-ropathology amp Experimental Neurology vol 72 no 8 pp 718ndash722 2013
[34] X He T Yan R Chen and D Ran ldquoAcute effects of electro-acupuncture (EA) on hippocampal long term potentiation(LTP) of perforant path-dentate gyrus granule cells synapserelated to memoryrdquo Acupuncture and Electro-TherapeuticsResearch vol 37 no 2-3 pp 89ndash101 2012
[35] W Dong W Guo X Zheng et al ldquoElectroacupunctureimproves cognitive deficits associated with AMPK activation inSAMP8 micerdquo Metabolic Brain Disease vol 30 no 3 pp 777ndash784 2015
[36] B Milner and D Klein ldquoLoss of recent memory after bilateralhippocampal lesions memory and memoriesmdashlooking backand looking forwardrdquo Journal of Neurology Neurosurgery ampPsychiatry vol 87 no 3 article 230 2016
[37] A Veronica Witte L Kerti D S Margulies and A FloelldquoEffects of resveratrol on memory performance hippocampalfunctional connectivity and glucose metabolism in healthyolder adultsrdquo Journal of Neuroscience vol 34 no 23 pp 7862ndash7870 2014
[38] T Bonhoeffer and R Yuste ldquoSpine motility phenomenologymechanisms and functionrdquoNeuron vol 35 no 6 pp 1019ndash10272002
Neural Plasticity 11
[39] H Qiao M Li C Xu H Chen S An and X Ma ldquoDendriticspines in depression what we learned from animal modelsrdquoNeural Plasticity vol 2016 Article ID 8056370 26 pages 2016
[40] S C McQuown R M Barrett D P Matheos et al ldquoHDAC3 isa critical negative regulator of long-term memory formationrdquoJournal of Neuroscience vol 31 no 2 pp 764ndash774 2011
[41] K-O Lai and N Y Ip ldquoStructural plasticity of dendritic spinesthe underlying mechanisms and its dysregulation in brain dis-ordersrdquo Biochimica et Biophysica ActamdashMolecular Basis of Dis-ease vol 1832 no 12 pp 2257ndash2263 2013
[42] C H Bailey E R Kandel and K M Harris ldquoStructural com-ponents of synaptic plasticity and memory consolidationrdquo ColdSpring Harbor Perspectives in Biology vol 7 no 7 Article IDa021758 2015
[43] C E Brown P Li J D Boyd K R Delaney and T HMurphy ldquoExtensive turnover of dendritic spines and vascularremodeling in cortical tissues recovering from strokerdquo Journalof Neuroscience vol 27 no 15 pp 4101ndash4109 2007
[44] R Mostany T G Chowdhury D G Johnston S A PortonovoS T Carmichael and C Portera-Cailliau ldquoLocal hemodynam-ics dictate long-term dendritic plasticity in peri-infarct cortexrdquoJournal of Neuroscience vol 30 no 42 pp 14116ndash14126 2010
[45] J George C Soares A Montersino J-C Beique and G MThomas ldquoPalmitoylation of LIM kinase-1 ensures spine-specificactin polymerization and morphological plasticityrdquo eLife vol2015 no 4 Article ID e06327 2015
[46] S Bicker M Lackinger K Weiszlig and G Schratt ldquoMicroRNA-132 -134 and -138 a microRNA troika rules in neuronaldendritesrdquo Cellular and molecular life sciences CMLS vol 71no 20 pp 3987ndash4005 2014
[47] H-C Tai and E M Schuman ldquoMicroRNA microRNAs reachout into dendritesrdquoCurrent Biology vol 16 no 4 pp R121ndashR1232006
Submit your manuscripts athttpswwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

Neural Plasticity 9
0
2
4
6
8
10
12
miR
-134
relat
ive e
xpre
ssio
n to
U6 lowastlowast
and
MICD+non-EA
MICD+EAMICDSham
Figure 7 Changes of miR-134 expression in the hippocampusCA1 cells The changes of miR-134 expression level were measuredusing quantitative RT-PCR U6 was used as internal control forquantification of miRNAs Data are mean plusmn SEM (119899 = 6grouplowastlowast119901 lt 001 versus the Sham group 119901 lt 001 119901 lt 005 versus theMICD group andand119901 lt 001 and119901 lt 005 versus the MICD+EA group)
regeneration Dendritic spine is the tiny protrusions withmulti-dense body and ion channels on the surface of varioustypes of neurons which serve as cellular substrates of brainconnectivity and the major sites of information processingin the brain [38 39] Evidence is increasing that synaptic-dendritic plasticity enables the encoding of memory [40]Indeed themodification on dendritic spine number size andshape is of importance for the plasticity of synapses accom-panied by hippocampus-dependent learning and memory[41 42] Studies have well described that dendritic spineremodeling in ischemia surrounding areas may contribute tocognitive functional recovery after stroke [43 44]
Increasing evidence showed that LIMK1 protein is acti-vated at dendritic spine membranes [45] LIMK1 positivecells were located in the CA1 region of the hippocampuswhich is important in the regulation of the spinemorphologyand synaptic function in vivo A prominent form of longlasting synaptic plasticity is thought to be critical to memoryformation [26] The present study found that the expressionof total LIMK1 and phospho-LIMK1 in hippocampal CA1wassignificantly decreased in the MICD rats however EA couldenhance the total LIMK1 and phospho-LIMK1 levels to pro-mote synaptic-dendritic plasticity in hippocampal CA1 area
In addition we have identified a dendritically localizedmiRNA that regulated the expression of the synaptic LIMK1protein thereby controlling dendritic pine sizemiR-134 is thefirst discovered dendritic microRNA enriched in the neu-ronal dendrites of rat hippocampal neurons and negativelycontrols the size of dendritic spines [46] This effect ismediated by miR-134 inhibition of the translation of LIMK1mRNA [47] The present study showed that the expression ofmiR-134 was obviously increased in theMICD rats Howeverrepeated EA treatment could relieve the upregulation of miR-134 expression in hippocampal CA1 area
In conclusion the study indicated that EA at the DU20and DU24 acupoints could ameliorate cognitive impairmentof MICD rats through the regulation of synaptic-dendritic
plasticity of the hippocampal CA1 area and the expressionand phosphorylation of synaptic LIMK1 protein were acti-vated to control dendritic spine number size and shapeFurthermore the mechanism of LIMK1 increase induced byEA treatmentmay be associated withmiR-134 which is local-ized in hippocampal CA1 by negatively regulating LIMK1 toenhance synaptic-dendritic plasticity in the recovery stage ofischemic stroke
Competing Interests
The authors declare no financial or commercial conflict ofinterests
Authorsrsquo Contributions
Weilin Liu Jie Wu and Jia Huang contributed equally to thiswork
Acknowledgments
The present study was supported by the Natural ScienceFoundation of China (no 81403450) and the Natural ScienceFoundation of Fujian Province (no 2016J01382)
References
[1] V L Roger ldquoHeart disease and stroke statisticsmdash2011 update areport from the American Heart Associationrdquo Circulation vol131 no 4 pp 29ndash322 2014
[2] T E Nichols and A P Holmes ldquoNonparametric permutationtests for functional neuroimaging a primer with examplesrdquoHuman Brain Mapping vol 15 no 1 pp 1ndash25 2002
[3] T K Tatemichi D W Desmond Y Stern M Paik M Sanoand E Bagiella ldquoCognitive impairment after stroke frequencypatterns and relationship to functional abilitiesrdquo Journal ofNeurology Neurosurgery amp Psychiatry vol 57 no 2 pp 202ndash207 1994
[4] F Liu Z-M Li Y-J Jiang and L-D Chen ldquoA meta-analysis ofacupuncture use in the treatment of cognitive impairment afterstrokerdquo Journal of Alternative amp Complementary Medicine vol20 no 7 pp 535ndash544 2014
[5] W Li R Huang R A Shetty et al ldquoTransient focal cerebralischemia induces long-termcognitive function deficit in an exp-erimental ischemic stroke modelrdquo Neurobiology of Disease vol59 pp 18ndash25 2013
[6] P S Sachdev X Chen H Brodaty CThompson A Altendorfand W Wen ldquoThe determinants and longitudinal course ofpost-stroke mild cognitive impairmentrdquo Journal of the Inter-national Neuropsychological Society vol 15 no 6 pp 915ndash9232009
[7] A-J Liu J-H Li H-Q Li et al ldquoElectroacupuncture foracute ischemic stroke ameta-analysis of randomized controlledtrialsrdquoThe American Journal of Chinese Medicine vol 43 no 8pp 1541ndash1566 2015
[8] X Ding C-Y Li Q-S Wang et al ldquoPatterns in default-modenetwork connectivity for determining outcomes in cognitivefunction in acute stroke patientsrdquo Neuroscience vol 277 no 10pp 637ndash646 2014
10 Neural Plasticity
[9] H Alipanahzadeh M Soleimani S Soleimani Asl B Pourhey-dar A Nikkhah and M Mehdizadeh ldquoTransforming growthfactor-alpha improves memory impairment and neurogenesisfollowing ischemia reperfusionrdquo Cell Journal vol 16 no 3 pp315ndash324 2014
[10] H Eichenbaum ldquoHippocampus cognitive processes and neuralrepresentations that underlie declarative memoryrdquoNeuron vol44 no 1 pp 109ndash120 2004
[11] T Kadar S Dachir B Shukitt-Hale and A Levy ldquoSub-regionalhippocampal vulnerability in various animal models leading tocognitive dysfunctionrdquo Journal of Neural Transmission vol 105no 8-9 pp 987ndash1004 1998
[12] J B Pereira C Valls-Pedret E Ros et al ldquoRegional vulnera-bility of hippocampal subfields to aging measured by structuraland diffusion MRIrdquo Hippocampus vol 24 no 4 pp 403ndash4142014
[13] S Knafo G Ariav E Barkai and F Libersat ldquoOlfactory learn-ing-induced increase in spine density along the apical dendritesof CA1 hippocampal neuronsrdquo Hippocampus vol 14 no 7 pp819ndash825 2004
[14] S Schacher and J-Y Hu ldquoThe less things change the more theyare different contributions of long-term synaptic plasticity andhomeostasis to memoryrdquo Learning and Memory vol 21 no 3pp 128ndash134 2014
[15] K Park H Heo M E Han et al ldquoLearning-induced synapticpotentiation in implanted neural precursor cell-derived neu-ronsrdquo Scientific Reports vol 5 article 17796 2015
[16] S Knafo F Libersat and E Barkai ldquoDynamics of learning-induced spine redistribution along dendrites of pyramidalneurons in ratsrdquo European Journal of Neuroscience vol 21 no4 pp 927ndash935 2005
[17] V C Foletta N Moussi P D Sarmiere J R Bamburg and OBernard ldquoLIM kinase 1 a key regulator of actin dynamics iswidely expressed in embryonic and adult tissuesrdquo ExperimentalCell Research vol 294 no 2 pp 392ndash405 2004
[18] M Endo K Ohashi Y Sasaki et al ldquoControl of growth conemotility andmorphology by LIMkinase and slingshot via phos-phorylation and dephosphorylation of cofilinrdquo Journal of Neu-roscience vol 23 no 7 pp 2527ndash2537 2003
[19] C Jarrold A D Baddeley and C Phillips ldquoLong-termmemoryfor verbal and visual information in Down syndrome andWilliams syndrome performance on the doors and people testrdquoCortex vol 43 no 2 pp 233ndash247 2007
[20] Y Meng Y Zhang V Tregoubov et al ldquoAbnormal spinemorphology and enhanced LTP in LIMK-1 knockout micerdquoNeuron vol 35 no 1 pp 121ndash133 2002
[21] Z Todorovski S Asrar J Liu et al ldquoLIMK1 regulates long-termmemory and synaptic plasticity via the transcriptional factorCREBrdquo Molecular and Cellular Biology vol 35 no 8 pp 1316ndash1328 2015
[22] G M Schratt F Tuebing E A Nigh et al ldquoA brain-specificmicroRNA regulates dendritic spine developmentrdquo Nature vol439 no 7074 pp 283ndash289 2006
[23] F Capitano J Camon V Ferretti et al ldquomicroRNAs modulatespatial memory in the hippocampus and in the ventral striatumin a region-specific mannerrdquo Molecular Neurobiology vol 53no 7 pp 4618ndash4630 2015
[24] J GaoW-YWang Y-WMao et al ldquoA novel pathway regulatesmemory and plasticity via SIRT1 andmiR-134rdquoNature vol 466no 7310 pp 1105ndash1109 2010
[25] J Remenyi M W M van den Bosch O Palygin et al ldquomiR-132212 knockout mice reveal roles for these miRNAs in regu-lating cortical synaptic transmission and plasticityrdquo PLoS ONEvol 8 no 4 Article ID e62509 p 65 2013
[26] L An J LiuWW Li et al ldquoDistribution of LIM domain kinase1 in the olfactory bulb cerebral cortex hippocampus and cere-bellum of the AppPS+minus micerdquo Genetics amp Molecular Researchvol 14 no 4 pp 17244ndash17251 2015
[27] R Lin Y Wu J Tao et al ldquoElectroacupuncture improves cog-nitive function through Rho GTPases and enhances dendriticspine plasticity in rats with cerebral ischemia-reperfusionrdquoMolecular Medicine Reports vol 13 no 3 pp 2655ndash2660 2016
[28] P Castaneda M Munoz G Garcıa-Rojo et al ldquoAssociation ofN-cadherin levels and downstream effectors of Rho GTPaseswith dendritic spine loss induced by chronic stress in rathippocampal neuronsrdquo Journal of Neuroscience Research vol93 no 10 pp 1476ndash1491 2015
[29] A Romarowski M A Battistone F A La Spina et al ldquoPKA-dependent phosphorylation of LIMK1 and Cofilin is essentialformouse sperm acrosomal exocytosisrdquoDevelopmental Biologyvol 405 no 2 pp 237ndash249 2015
[30] R Lin K Yu X Li et al ldquoElectroacupuncture ameliorates post-stroke learning and memory through minimizing ultrastruc-tural brain damage and inhibiting the expression of MMP-2andMMP-9 in cerebral ischemia-reperfusion injured ratsrdquoMol-ecular Medicine Reports vol 14 no 1 pp 225ndash233 2016
[31] X Feng S Yang J Liu et al ldquoElectroacupuncture amelioratescognitive impairment through inhibition of NF-120581B-mediatedneuronal cell apoptosis in cerebral ischemia-reperfusion inj-ured ratsrdquo Molecular Medicine Reports vol 7 no 5 pp 1516ndash1522 2013
[32] Z K Wang N I Guang-Xia K Liu et al ldquoResearch on thechanges of IL-1 receptor and TNF-120572 receptor in rats with cere-bral ischemia reperfusion and the chronergy of acupunctureinterventionrdquo Zhongguo Zhen Jiu vol 32 no 11 pp 1012ndash10182012
[33] X S LiuM Chopp R L Zhang and Z G Zhang ldquoMicroRNAsin cerebral ischemia-induced neurogenesisrdquo Journal of Neu-ropathology amp Experimental Neurology vol 72 no 8 pp 718ndash722 2013
[34] X He T Yan R Chen and D Ran ldquoAcute effects of electro-acupuncture (EA) on hippocampal long term potentiation(LTP) of perforant path-dentate gyrus granule cells synapserelated to memoryrdquo Acupuncture and Electro-TherapeuticsResearch vol 37 no 2-3 pp 89ndash101 2012
[35] W Dong W Guo X Zheng et al ldquoElectroacupunctureimproves cognitive deficits associated with AMPK activation inSAMP8 micerdquo Metabolic Brain Disease vol 30 no 3 pp 777ndash784 2015
[36] B Milner and D Klein ldquoLoss of recent memory after bilateralhippocampal lesions memory and memoriesmdashlooking backand looking forwardrdquo Journal of Neurology Neurosurgery ampPsychiatry vol 87 no 3 article 230 2016
[37] A Veronica Witte L Kerti D S Margulies and A FloelldquoEffects of resveratrol on memory performance hippocampalfunctional connectivity and glucose metabolism in healthyolder adultsrdquo Journal of Neuroscience vol 34 no 23 pp 7862ndash7870 2014
[38] T Bonhoeffer and R Yuste ldquoSpine motility phenomenologymechanisms and functionrdquoNeuron vol 35 no 6 pp 1019ndash10272002
Neural Plasticity 11
[39] H Qiao M Li C Xu H Chen S An and X Ma ldquoDendriticspines in depression what we learned from animal modelsrdquoNeural Plasticity vol 2016 Article ID 8056370 26 pages 2016
[40] S C McQuown R M Barrett D P Matheos et al ldquoHDAC3 isa critical negative regulator of long-term memory formationrdquoJournal of Neuroscience vol 31 no 2 pp 764ndash774 2011
[41] K-O Lai and N Y Ip ldquoStructural plasticity of dendritic spinesthe underlying mechanisms and its dysregulation in brain dis-ordersrdquo Biochimica et Biophysica ActamdashMolecular Basis of Dis-ease vol 1832 no 12 pp 2257ndash2263 2013
[42] C H Bailey E R Kandel and K M Harris ldquoStructural com-ponents of synaptic plasticity and memory consolidationrdquo ColdSpring Harbor Perspectives in Biology vol 7 no 7 Article IDa021758 2015
[43] C E Brown P Li J D Boyd K R Delaney and T HMurphy ldquoExtensive turnover of dendritic spines and vascularremodeling in cortical tissues recovering from strokerdquo Journalof Neuroscience vol 27 no 15 pp 4101ndash4109 2007
[44] R Mostany T G Chowdhury D G Johnston S A PortonovoS T Carmichael and C Portera-Cailliau ldquoLocal hemodynam-ics dictate long-term dendritic plasticity in peri-infarct cortexrdquoJournal of Neuroscience vol 30 no 42 pp 14116ndash14126 2010
[45] J George C Soares A Montersino J-C Beique and G MThomas ldquoPalmitoylation of LIM kinase-1 ensures spine-specificactin polymerization and morphological plasticityrdquo eLife vol2015 no 4 Article ID e06327 2015
[46] S Bicker M Lackinger K Weiszlig and G Schratt ldquoMicroRNA-132 -134 and -138 a microRNA troika rules in neuronaldendritesrdquo Cellular and molecular life sciences CMLS vol 71no 20 pp 3987ndash4005 2014
[47] H-C Tai and E M Schuman ldquoMicroRNA microRNAs reachout into dendritesrdquoCurrent Biology vol 16 no 4 pp R121ndashR1232006
Submit your manuscripts athttpswwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

10 Neural Plasticity
[9] H Alipanahzadeh M Soleimani S Soleimani Asl B Pourhey-dar A Nikkhah and M Mehdizadeh ldquoTransforming growthfactor-alpha improves memory impairment and neurogenesisfollowing ischemia reperfusionrdquo Cell Journal vol 16 no 3 pp315ndash324 2014
[10] H Eichenbaum ldquoHippocampus cognitive processes and neuralrepresentations that underlie declarative memoryrdquoNeuron vol44 no 1 pp 109ndash120 2004
[11] T Kadar S Dachir B Shukitt-Hale and A Levy ldquoSub-regionalhippocampal vulnerability in various animal models leading tocognitive dysfunctionrdquo Journal of Neural Transmission vol 105no 8-9 pp 987ndash1004 1998
[12] J B Pereira C Valls-Pedret E Ros et al ldquoRegional vulnera-bility of hippocampal subfields to aging measured by structuraland diffusion MRIrdquo Hippocampus vol 24 no 4 pp 403ndash4142014
[13] S Knafo G Ariav E Barkai and F Libersat ldquoOlfactory learn-ing-induced increase in spine density along the apical dendritesof CA1 hippocampal neuronsrdquo Hippocampus vol 14 no 7 pp819ndash825 2004
[14] S Schacher and J-Y Hu ldquoThe less things change the more theyare different contributions of long-term synaptic plasticity andhomeostasis to memoryrdquo Learning and Memory vol 21 no 3pp 128ndash134 2014
[15] K Park H Heo M E Han et al ldquoLearning-induced synapticpotentiation in implanted neural precursor cell-derived neu-ronsrdquo Scientific Reports vol 5 article 17796 2015
[16] S Knafo F Libersat and E Barkai ldquoDynamics of learning-induced spine redistribution along dendrites of pyramidalneurons in ratsrdquo European Journal of Neuroscience vol 21 no4 pp 927ndash935 2005
[17] V C Foletta N Moussi P D Sarmiere J R Bamburg and OBernard ldquoLIM kinase 1 a key regulator of actin dynamics iswidely expressed in embryonic and adult tissuesrdquo ExperimentalCell Research vol 294 no 2 pp 392ndash405 2004
[18] M Endo K Ohashi Y Sasaki et al ldquoControl of growth conemotility andmorphology by LIMkinase and slingshot via phos-phorylation and dephosphorylation of cofilinrdquo Journal of Neu-roscience vol 23 no 7 pp 2527ndash2537 2003
[19] C Jarrold A D Baddeley and C Phillips ldquoLong-termmemoryfor verbal and visual information in Down syndrome andWilliams syndrome performance on the doors and people testrdquoCortex vol 43 no 2 pp 233ndash247 2007
[20] Y Meng Y Zhang V Tregoubov et al ldquoAbnormal spinemorphology and enhanced LTP in LIMK-1 knockout micerdquoNeuron vol 35 no 1 pp 121ndash133 2002
[21] Z Todorovski S Asrar J Liu et al ldquoLIMK1 regulates long-termmemory and synaptic plasticity via the transcriptional factorCREBrdquo Molecular and Cellular Biology vol 35 no 8 pp 1316ndash1328 2015
[22] G M Schratt F Tuebing E A Nigh et al ldquoA brain-specificmicroRNA regulates dendritic spine developmentrdquo Nature vol439 no 7074 pp 283ndash289 2006
[23] F Capitano J Camon V Ferretti et al ldquomicroRNAs modulatespatial memory in the hippocampus and in the ventral striatumin a region-specific mannerrdquo Molecular Neurobiology vol 53no 7 pp 4618ndash4630 2015
[24] J GaoW-YWang Y-WMao et al ldquoA novel pathway regulatesmemory and plasticity via SIRT1 andmiR-134rdquoNature vol 466no 7310 pp 1105ndash1109 2010
[25] J Remenyi M W M van den Bosch O Palygin et al ldquomiR-132212 knockout mice reveal roles for these miRNAs in regu-lating cortical synaptic transmission and plasticityrdquo PLoS ONEvol 8 no 4 Article ID e62509 p 65 2013
[26] L An J LiuWW Li et al ldquoDistribution of LIM domain kinase1 in the olfactory bulb cerebral cortex hippocampus and cere-bellum of the AppPS+minus micerdquo Genetics amp Molecular Researchvol 14 no 4 pp 17244ndash17251 2015
[27] R Lin Y Wu J Tao et al ldquoElectroacupuncture improves cog-nitive function through Rho GTPases and enhances dendriticspine plasticity in rats with cerebral ischemia-reperfusionrdquoMolecular Medicine Reports vol 13 no 3 pp 2655ndash2660 2016
[28] P Castaneda M Munoz G Garcıa-Rojo et al ldquoAssociation ofN-cadherin levels and downstream effectors of Rho GTPaseswith dendritic spine loss induced by chronic stress in rathippocampal neuronsrdquo Journal of Neuroscience Research vol93 no 10 pp 1476ndash1491 2015
[29] A Romarowski M A Battistone F A La Spina et al ldquoPKA-dependent phosphorylation of LIMK1 and Cofilin is essentialformouse sperm acrosomal exocytosisrdquoDevelopmental Biologyvol 405 no 2 pp 237ndash249 2015
[30] R Lin K Yu X Li et al ldquoElectroacupuncture ameliorates post-stroke learning and memory through minimizing ultrastruc-tural brain damage and inhibiting the expression of MMP-2andMMP-9 in cerebral ischemia-reperfusion injured ratsrdquoMol-ecular Medicine Reports vol 14 no 1 pp 225ndash233 2016
[31] X Feng S Yang J Liu et al ldquoElectroacupuncture amelioratescognitive impairment through inhibition of NF-120581B-mediatedneuronal cell apoptosis in cerebral ischemia-reperfusion inj-ured ratsrdquo Molecular Medicine Reports vol 7 no 5 pp 1516ndash1522 2013
[32] Z K Wang N I Guang-Xia K Liu et al ldquoResearch on thechanges of IL-1 receptor and TNF-120572 receptor in rats with cere-bral ischemia reperfusion and the chronergy of acupunctureinterventionrdquo Zhongguo Zhen Jiu vol 32 no 11 pp 1012ndash10182012
[33] X S LiuM Chopp R L Zhang and Z G Zhang ldquoMicroRNAsin cerebral ischemia-induced neurogenesisrdquo Journal of Neu-ropathology amp Experimental Neurology vol 72 no 8 pp 718ndash722 2013
[34] X He T Yan R Chen and D Ran ldquoAcute effects of electro-acupuncture (EA) on hippocampal long term potentiation(LTP) of perforant path-dentate gyrus granule cells synapserelated to memoryrdquo Acupuncture and Electro-TherapeuticsResearch vol 37 no 2-3 pp 89ndash101 2012
[35] W Dong W Guo X Zheng et al ldquoElectroacupunctureimproves cognitive deficits associated with AMPK activation inSAMP8 micerdquo Metabolic Brain Disease vol 30 no 3 pp 777ndash784 2015
[36] B Milner and D Klein ldquoLoss of recent memory after bilateralhippocampal lesions memory and memoriesmdashlooking backand looking forwardrdquo Journal of Neurology Neurosurgery ampPsychiatry vol 87 no 3 article 230 2016
[37] A Veronica Witte L Kerti D S Margulies and A FloelldquoEffects of resveratrol on memory performance hippocampalfunctional connectivity and glucose metabolism in healthyolder adultsrdquo Journal of Neuroscience vol 34 no 23 pp 7862ndash7870 2014
[38] T Bonhoeffer and R Yuste ldquoSpine motility phenomenologymechanisms and functionrdquoNeuron vol 35 no 6 pp 1019ndash10272002
Neural Plasticity 11
[39] H Qiao M Li C Xu H Chen S An and X Ma ldquoDendriticspines in depression what we learned from animal modelsrdquoNeural Plasticity vol 2016 Article ID 8056370 26 pages 2016
[40] S C McQuown R M Barrett D P Matheos et al ldquoHDAC3 isa critical negative regulator of long-term memory formationrdquoJournal of Neuroscience vol 31 no 2 pp 764ndash774 2011
[41] K-O Lai and N Y Ip ldquoStructural plasticity of dendritic spinesthe underlying mechanisms and its dysregulation in brain dis-ordersrdquo Biochimica et Biophysica ActamdashMolecular Basis of Dis-ease vol 1832 no 12 pp 2257ndash2263 2013
[42] C H Bailey E R Kandel and K M Harris ldquoStructural com-ponents of synaptic plasticity and memory consolidationrdquo ColdSpring Harbor Perspectives in Biology vol 7 no 7 Article IDa021758 2015
[43] C E Brown P Li J D Boyd K R Delaney and T HMurphy ldquoExtensive turnover of dendritic spines and vascularremodeling in cortical tissues recovering from strokerdquo Journalof Neuroscience vol 27 no 15 pp 4101ndash4109 2007
[44] R Mostany T G Chowdhury D G Johnston S A PortonovoS T Carmichael and C Portera-Cailliau ldquoLocal hemodynam-ics dictate long-term dendritic plasticity in peri-infarct cortexrdquoJournal of Neuroscience vol 30 no 42 pp 14116ndash14126 2010
[45] J George C Soares A Montersino J-C Beique and G MThomas ldquoPalmitoylation of LIM kinase-1 ensures spine-specificactin polymerization and morphological plasticityrdquo eLife vol2015 no 4 Article ID e06327 2015
[46] S Bicker M Lackinger K Weiszlig and G Schratt ldquoMicroRNA-132 -134 and -138 a microRNA troika rules in neuronaldendritesrdquo Cellular and molecular life sciences CMLS vol 71no 20 pp 3987ndash4005 2014
[47] H-C Tai and E M Schuman ldquoMicroRNA microRNAs reachout into dendritesrdquoCurrent Biology vol 16 no 4 pp R121ndashR1232006
Submit your manuscripts athttpswwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

Neural Plasticity 11
[39] H Qiao M Li C Xu H Chen S An and X Ma ldquoDendriticspines in depression what we learned from animal modelsrdquoNeural Plasticity vol 2016 Article ID 8056370 26 pages 2016
[40] S C McQuown R M Barrett D P Matheos et al ldquoHDAC3 isa critical negative regulator of long-term memory formationrdquoJournal of Neuroscience vol 31 no 2 pp 764ndash774 2011
[41] K-O Lai and N Y Ip ldquoStructural plasticity of dendritic spinesthe underlying mechanisms and its dysregulation in brain dis-ordersrdquo Biochimica et Biophysica ActamdashMolecular Basis of Dis-ease vol 1832 no 12 pp 2257ndash2263 2013
[42] C H Bailey E R Kandel and K M Harris ldquoStructural com-ponents of synaptic plasticity and memory consolidationrdquo ColdSpring Harbor Perspectives in Biology vol 7 no 7 Article IDa021758 2015
[43] C E Brown P Li J D Boyd K R Delaney and T HMurphy ldquoExtensive turnover of dendritic spines and vascularremodeling in cortical tissues recovering from strokerdquo Journalof Neuroscience vol 27 no 15 pp 4101ndash4109 2007
[44] R Mostany T G Chowdhury D G Johnston S A PortonovoS T Carmichael and C Portera-Cailliau ldquoLocal hemodynam-ics dictate long-term dendritic plasticity in peri-infarct cortexrdquoJournal of Neuroscience vol 30 no 42 pp 14116ndash14126 2010
[45] J George C Soares A Montersino J-C Beique and G MThomas ldquoPalmitoylation of LIM kinase-1 ensures spine-specificactin polymerization and morphological plasticityrdquo eLife vol2015 no 4 Article ID e06327 2015
[46] S Bicker M Lackinger K Weiszlig and G Schratt ldquoMicroRNA-132 -134 and -138 a microRNA troika rules in neuronaldendritesrdquo Cellular and molecular life sciences CMLS vol 71no 20 pp 3987ndash4005 2014
[47] H-C Tai and E M Schuman ldquoMicroRNA microRNAs reachout into dendritesrdquoCurrent Biology vol 16 no 4 pp R121ndashR1232006
Submit your manuscripts athttpswwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

Submit your manuscripts athttpswwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014





























