Embed Size (px)
Citation preview

J. clin. Path., 23, Suppl. (Roy. Coll. Path.), 4, 16-31
Electrolyte and body water changesafter trauma
CECIL T. G. FLEARFrom the University Department ofClinical Biochemistry, Royal Victoria Infirmary, Newcastle upon Tyne
It has been known for a long time now thatpatients commonly gain sodium and chlorideafter injuries and surgery. It is also well knownthat despite this fact, the level of sodium inextracellular fluid very often falls, and is ascribedto a relatively greater gain of water than of salt,with resulting dilution of body fluids. However,the not uncommon associated rise in potassiumand bicarbonate levels is difficult to reconcile withsimple dilution. We know too that patients losepotassium after trauma and that more is lostthan might be expected to be brought about bythe 'catabolism' of tissue which trauma provokes.Why these changes arise is much less certain
than that they do. The gains of sodium andchloride, and also the losses of potassium, areusually held to be brought about by alteredkidney activity in turn caused by increased pro-duction of aldosterone. Imbalance betweenprotein anabolism and catabolism is said to beproduced by increased secretion of gluco-corticoids. But surgery in patients previouslyadrenalectomized provokes the same gains ofNaand Cl and losses of K and nitrogen even thoughthe patients receive the same maintenance dose ofsteroid after surgery as before.There is uncertainty too about the extent and
nature of any change in the volume of extra-cellular fluid during and after trauma. Fromdilution studies, variously carried out, somehave concluded that the volume of extracellularfluid expands, others that it shrinks, others thatit remains unchanged.
I think that present uncertainties on all scoresarise from inadequacies in our knowledge ofcellular happenings provoked by trauma-oftheir nature and of their consequences. Measure-ments show that cardiac output falls duringsurgery (McNeill and Mackenzie, unpublishedobservations), even when every attempt is made
fully to replace blood as it is lost, and whenindices such as central venous pressure, systemicblood pressure, pulse rate, and measurement ofblood loss on swabs suggest that this has beenachieved. Moreover, even uncomplicated surgeryis accompanied by hypoxia (Nunn, 1965; Nunnand Freeman, 1964). This, together with impairedor altered tissue perfusion from blood loss, etc,is likely to provoke widespread cell changeswhich, with directly incurred local tissue celldamage, may be largely responsible for theaccumulation of Na, Cl, and water, for loss ofpotassium, for hyponatraemia, and for otherchanges in the composition of extracellular fluid,and possibly also for the imbalance between tissueprotein anabolism and catabolism. Postoperativecomplications, such as further blood loss, in-flammatory accumulation of protein and fluidin the peritoneum or elsewhere, infection andsepsis, will directly or indirectly accentuate thecellular changes and so the changes in body waterand electrolytes.
Regulation of Body Water and Electrolytes inHealth
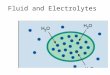
Most of our body water is inside cells. Some isoutside (Fig. 1,1). Virtually all is osmoticallyavailable (Bernal, 1965; Olmstead, 1966; Netter,1969), dissolving solutes, so that it is presentthroughout effectively as a dilute solution whichis stirred (Fig. 1,2) by the circulation (Table I)and by diffusion across capillary walls, throughinterstitial fluid, across outer cell membranes,and within cells. Over the very short distancesinvolved (cell diameter 20-50 ,u; cell to capillary,approximately 25.4 ,t) diffusion will ensurevirtually instantaneous mixing (Robinson, 1960;
on March 31, 2020 by guest. P
rotected by copyright.http://jcp.bm
j.com/
J Clin P
athol: first published as 10.1136/jcp.s3-4.1.16 on 1 January 1970. Dow
nloaded from

Electrolyte and body water changes after trauma
1 2
Vj c 2V0
approx. [Na]i = 10 [Nla]= 140imeq./1 [K]j = 160 [K]0= 4
[Osm]i - [Osm]0290 mOsm/lg
Og = QI
iQ,
Qg = 0l
5
Na+ _i. Na+Cl- cl-K +_- K +
zA- D
Q,Fig. 1 Simplified presentation of the most importantfeatures of water and electrolyte balance.
Landis and Pappenheimer, 1963; Langer, 1968).1Fluid inside cells has the same osmolality as thatoutside. Osmotic transients which arise duringthe course of metabolic activity are cancelled byappropriate shifts of water which diffuses intoand out of cells vastly more quickly than solutes(Robinson, 1965).The principal solutes in body fluids are salts
of Na and K. These two cations and their anionshave, however, an asymmetrical distributionbetween fluid inside and outside cells (Fig. 1,3).
All gain water and electrolytes in food anddrink and all lose both in urine, faeces, and sweat.Water is of course also produced within cells asa result of cellular respiration and lost throughthe lungs and in insensible perspiration. Never-theless, so long as adults are healthy the amountsof both water and electrolytes in the body are
'It can be calculated (after Jacobs, 1935) that for a solute diffus-ing into a pure solvent from the plane boundary of a solutionmaintained at a fixed composition, the concentration 25-4 ji fromthe boundary should reach 99% of its steady state value in 0-294seconds. In this time, and under resting conditions, plasma flowthrough 100 g of skeletal muscle would be approximately 1-5 x10-' ml.Landisand Pappenheimer(1963) havecalculated thatbydiffusion
plasma water exchanges 120 times per minute with the interstitialwater immediately surrounding the capillaries (for K and Na, thecorresponding exchanges are 90 and 60 times plasma content).Alternatively these exchanges amount respectively to 40, 30, and20 times the resting plasma flow in skeletal muscle. (Clearance isless.)
fairly constant week in week out. This constancyis dynamically attained from adjustment of lossesin urine in the light of various gains and thevarious other losses in sweat, faeces, and so on(Fig. 1,4). Constancy is achieved in a stepwisefashion. Small deficits provoke net gains and viceversa. The kidneys are assisted in this process bya number of neurohumoral servomechanismswhich signal shortages or excesses and theircancellation.But self maintenance of body fluids entails
more than simply balancing the quantities gainedand lost. This alone would not preserve theasymmetrical distribution of Na and K in bodyfluids; nor would it ensure that the volumes ofextra- and intracellular fluid remain fairly
Plasma Flow Multiples ofper Minute Extracellular(1.) Fluid per
Minute
Cardiac output at rest 3.0 0.221Skeletal muscle blood flow
Atrest 0-6 0l10 1'During vigorous exercise" 22-0 3-68 f
Table I Movement of body water in healthy adults'Extracellular fluid 14-1'Extracellular fluid = 20% wet muscle weight.3Wade and Bishop (1962)
3
17 on M
arch 31, 2020 by guest. Protected by copyright.
http://jcp.bmj.com
/J C
lin Pathol: first published as 10.1136/jcp.s3-4.1.16 on 1 January 1970. D
ownloaded from

Cecil T. G. Flear
constant, which they both do, in the face of thecontinual interchange of water and ions thattakes place between fluid compartments.
Attainment of Cell Balance and Its Significance forHomeostasis
Constancy in the total volume of intracellularfluid derives from the separate regulation of thevolume of fluid in each of the millions of cellswithin the body. This is achieved as a result ofeach cell keeping its total solute content constant.And this involves attaining a balance between(1) the rate at which Na leaks into cells from theextracellular fluid and the rate at which it isactively transported, or pumped out again(Post and Jolly, 1957; Robinson, 1960, 1965;Tosteson and Hoffman, 1960; Tosteson, 1964;Stein, 1967) and (2) the metabolic productionand 'disposal' of non-diffusible organic solutes.The way in which both balances are achieved
has been little studied. Possibly in man the latterbalance derives largely as a byproduct of regula-tion of cell metabolism. But it is known thatchange in osmolality of extracellular fluid incertain euryhaline inhabitants of estuaries (causedby the changing tides) provokes specific changesin metabolism which drastically alter the amountsof organic solutes in cells, so that the osmolalityof cell fluids also changes and in a parallel direc-tion (Brown and Stein, 1960; Lockwood, 1963;Potts and Parry, 1964; Schoffeniels, 1967).This minimizes the shifts of water between extra-and intracellular fluid caused by the change inosmolality of extracellular fluid, and Wolf andhis colleagues (McDowell, Wolf, and Steer, 1955)have suggested that similar changes may takeplace in response to fluctuation in osmoticpressure of extracellular fluid in the cells ofterrestrial animals.The way in which the Na pumps are regulated
is poorly understood (Mullins and Frumento,1963; Woodbury, 1963, 1965; Mullins and Awad,1965; Langer, 1968). They are, however, collec-tively responsible for the asymmetrical distribu-tion of Na and K throughout the body.' Theyare also partly responsible for regulating theoverall daily balance of K, for the amount of Kin each cell, and so in the body as a whole, isdetermined by the quantity of non-diffusibleanions (zA-) it contains (Fig. 1,5). Conway(1957) considers that the anions which requireK as counterions, are largely metabolites such asATP, creatine phosphate, hexose, and triose
'Why cells possess Na pumps rather than K pumps is of coursean intriguing point. It is perhaps relevant that, since cells arenormally some 50 to 100 times more permeable to K than Na(probably because the hydrated Na ion is larger than that of K(Conway, 1957), although see Mullins, 1959, 1960; Diamond andWright, 1969), pumping K out from cells as fast as it diffused infrom the extracellular fluid would require a greater expenditure ofenergy than pumping out Na.
18
phosphates. Ling, on the other hand, suggests(Ling and Ochsenfeld. 1966) that K acts ascounterions to negative charges on structuralproteins. If Na accumulates in a cell, less K isneeded to maintain electrical neutrality. Theother main factor determining the amountof K in cells (and in the body) is the balancebetween production and disposal of non-diffusibleorganic solutes, since many of these are poly-valent anions.
Thus, the overall daily balance of K as wellas water derives from the balance attained at thecell level. The kidneys play a subservient role:they get rid of unavoidable dietary gains.2 Butsince K enters cells and is not extruded, it is notimmediately obvious why the daily provision ofK and diffusible anions fails to provoke a pro-gressive accumulation within cells. It can, how-ever, be shown that unless there is an increase inthe quantity of non-diffusible anions in cells,for cells to gain a substantial amount of potass-ium is only possible if there is a very markedincrease in the level of K in extracellular fluid.This is prevented by the spilling of K in urine,which is probably brought about by an increasedsecretion of aldosterone triggered by a risinglevel of K. The first observed effect when verysmall amounts of aldosterone are infused is onthe excretion ofK rather than of Na (Ross, 1965).
In contrast, the kidneys normally play thedominant role in maintaining a constant bodycontent of Na. In this they are guided by neuro-humoral servomechanisms activated by changein volume of extracellular fluid. Probably themost important factor here is the so-called 'thirdfactor'. This increases in response to expansionof extracellular fluid and brings about an increasein urinary loss of Na. Shrinkage of extracellularfluid provokes a decrease in the 'third factor' andalso increases secretion of aldosterone; excretionof Na is lessened. But for Na, too, inasmuch asgeneral body cells normally actively extrude Naas fast as it leaks in from the extracellular fluid,cell balance can be considered to play a part inthe regulation of body content. By the sametoken, in achieving a balance of Na and in themolar quantity of non-diffusible solutes con-strained within them, the body's cells contributetowards maintaining a fairly constant volume ofextracellular fluid.
In a balanced state there are at least threeequilibria across all cell membranes and withinfluids: (1) osmotic equilibrium, (2) electricalneutrality, and (3) double Donnan equilibrium.
iThe foods eaten by man are mostly plant and animal tissues orproducts made from them. Possibly not all urinary losses of Karise as spillage ofunavoidable dietary gains. There is, for instance,good evidence that first thing in the morning muscles lose a sub-stantial amount of K (Andres, Cader, Goldman, and Zierler,1957). Of course these and any other losses that may arise will becorrected promptly because the cells still have the usual amountof anions. With K available from the diet, cells simply eject theexcess of Na that accumulated to balance the deserted anions,and K is attracted into them by the electrical gradient createdduring the process of extrusion.
on March 31, 2020 by guest. P
rotected by copyright.http://jcp.bm
j.com/
J Clin P
athol: first published as 10.1136/jcp.s3-4.1.16 on 1 January 1970. Dow
nloaded from

Electrolyte and body water changes after trauma
Their resolution' in the light of overall dailybalances, and the outcome of exchanges betweencells and extracellular fluid are responsible forsetting the composition of extracellular fluid.The relative constancy of this composition inhealth derives as much therefore from the activityof general tissue cells in resolving their individualbalance problems as from the kidneys in regulat-ing overall balance (Fig. 1,6). At rest about 80%of the blood ejected with each beat of the heartgoes elsewhere than to the kidneys. Duringexercise this proportion ipcreases substantially.Just as cells help to keep the concentration ofhydrogen ions fairly constant in extracellularfluid, so too they absorb the first impact ofchange in other components of extracellularfluid thereby damping down any such change.For instance, a net loss of K from one tissue willprovoke an increase in the level of K in bloodleaving it, and so in blood which is supplied toother tissues. Cells in these other tissues in theprocess of coming into equilibrium with the newextracellular fluid levels will gain some K,thereby reducing its level. To some extent suchchanges take place during normal activity. Musclecells, for instance, gain Na and lose K when theycontract. Changes in metabolism are likely toalter the steady state levels of non-diffusible inter-mediates, and so to provoke net shifts of waterand K. But these changes are not sustained, norare they likely at any one time to involve muchtissue in a concerted fashion. In a sense theymay be considered to be stochastic, and this willresult in their being at least partly self cancelling.For instance, muscles are not used at the sametime, and in those being used, muscle fibres donot all contract and-relax in unison. Some relaxwhilst others contract, so that some cells will begaining K whilst others are losing it (Flear, 1967).
Imbalance at the Cell Level and Its Consequences
Sustained imbalance causes a cell to gain, or lose,water and ions, and, when enough cells areinvolved, a substantial change in body contentwill result. Abrupt changes will also alter thevolume of extracellular fluid.
Normally, the actual composition of extra-cellular fluid fluctuates about a point of balanceas a result of net exchanges of water and ionsacross cell membranes in response to the exig-encies of tissue function and metabolic activity.This will also be the case when a new balanced
'In view of its complexity the outcome of simultaneous resolu-tion of these equilibria in response to a given perturbation is notalways immediately obvious. But equations can be derived whichexpress the requirements for each of the separate equilibria. Theycan be combined in various ways (Boyle and Conway, 1941;Conway, 1945, 1957; Leaf, 1956; Adrian, 1960; Brown and Stein,1960) to give a number of additional equations that predict theoutcome of changes.
19
state has been established. When, however,imbalance involves progressively more cells, orbecomes progressively more marked, the com-position of the extracellular fluid will showprogressive changes. In labile situations withrapid alterations in the extent or direction ofimbalance in affected cells, or in the number ofcells involved, the volume and composition ofextracellular fluid will oscillate markedly abouta rather tenuous point of balance.
Cellular Happenings
When Na leaks into cells faster than it is baledout cells accumulate Na, Cl, and water. At thesame time they will lose K. The more abnormallypermeable the membranes, the less potassiumwill be lost during the early stages of the changingstate and the more the cells will swell. When abalanced state is re-established with cells neithergaining nor losing overall there is a substantialresidual loss of K and only a slight net gain ofwater (Stein, 1967).When cells swell abruptly at the expense of
extracellular fluid, that fluid shrinks. The levelof Na in extracellular fluid will not change, sincecells have gained virtually isosmotic NaCI, unlesscell membranes are so leaky that hitherto non-diffusible molecules escape into the extracellularfluid. When this happens the level of Na through-out the extracellular fluid [Na]0, will fall andthere will be hyponatraemia (Singh and Flear,1968). The reason for the fall is that in this situa-tion less water will be gained by cells than in theformer, and so the saline that is gained will havea higher Na level than that in the extracellularfluid. Osmolality will not fall. There has beenneither loss of solute from the body, nor dilutionof body fluids from gain of water. The volumeof extracellular fluid still shrinks when thishappens but to a lesser extent.
If at the same time that cells accumulate Na,more solutes are generated within cells whichthen escape through leaky membranes, cells willstill swell and the extracellular fluid shrink.Hyponatraemia, too, still develops.
In all of these situations the level ofK through-out the extracellular fluid is not diluted. On thecontrary hyperkalaemia is extremely likely todevelop. Accumulation of saline within cellsincreases the concentration of chloride in cellfluid. The product of the concentrations of Kand chloride thereupon rises. For the establish-ment of a new balanced state across cell mem-branes, the similar product of these concentra-tions in extracellular fluid must rise, since in abalanced state these two products are equal,as a consequence of a double Donnan equili-brium (equation 3, Appendix). In the presenceof an hyponatraemia the level of chloride inextracellular fluid must be low, and so an increase
on March 31, 2020 by guest. P
rotected by copyright.http://jcp.bm
j.com/
J Clin P
athol: first published as 10.1136/jcp.s3-4.1.16 on 1 January 1970. Dow
nloaded from

Cecil T. G. Flear
changing statehuman sk. muscle
+
plasma 0°C [Na] p=155 mM.
Fig. 2 Gains ofNa and water and loss ofK bypieces ofhuman skeletal muscle during thefirst40 min after immersion in coldplasma (0°C). Tenpieces of rectus abdominis (20-50 mg) from a givenperson were put into plasma taken from the sameperson. Duplicates were removed at each of thespecified time intervals. The experiment was repeatedusing samples from six patients. Values are thuseach the mean of 12. The level ofNa in all plasmaswas brought to 155 mM by addition of the appropriatevolume ofMNaCI. Gains and losses in each patientwere calculated by comparing with average com-
position ofsix samples not immersed in plasma.Data from Flear and Rajasingham (1969).
in the 'KCl product' is achieved by an increasein the concentration of K.The consequences of imbalance between leak
and pump rates can be predicted from what isknown of the factors governing the partition ofwater and ions in tissues. Knowledge of thesefactors has been derived largely from studies offrog skeletal muscle, squid nerves, and so on
in vitro. But it has been shown to apply to tissuesfrom other animals and to situations in situ(Kusel and Netter, 1952; Lukowsky and Netter,1952; Mond, 1955) as well. Many of us considerthat this understanding is directly relevant tohuman tissue also, and Fig. 2 shows what hap-pens to pieces of human rectus abdominis whenimbalance between influx and reflux of Na isprovoked by putting them into human plasmaat 0°C. In this situation there is also an increasein organic anions (zA-) in the tissue, so thatpieces show rather more residual gain of waterthan they would otherwise.The various situations considered above were
selected because anoxia, in addition to impairingNa pumps, is known to provoke breakdown of
[Na]j 50'[Na]j 8o
[Na]j 120
0 0l 05 [0
relative permeabilities Na/K
Fig. 3 Shrinkage ofextracellular fluid during abruptswelling of cells caused by imbalance between therate at which Na leaks into cells and the rate atwhich it is actively transported out again (seeAppendix for details of calculation).
Cells swell more when the permeability of theirmembranes increases, indicated by an increase in theratio ofpermeabilities towards Na and K. In theexamples shown, 10, 20, and 30% of the body'scells are assumed to swvell. It is further assumed thatthese cells can pump Na at a rate just insufficientto balance the passive influx when the Na concentra-tion in cell water is 120, 80, and 50 m-equiv per
litre respectively. A concentration ofNa in extra-cellular fluid of 140 m-equiv per litre is assumed.
compounds such as ATP and creatine phosphate,thereby increasing the molar quantity of soluteswithin cells. It is also known to increase perme-
ability of cell membranes with resultant escapeof some, at any rate, of the breakdown products.
Effects of Abrupt Inbalance on Volume of Extra-cellular Fluid
It is possible to go further and to calculate thesort of impact that abrupt and extensive cellularswelling will have on the volume of extracellularfluid.
During the changing state of Na gain, elec-trical neutrality is maintained in intra- and extra-cellular fluid, and in the fluid shifting from onecompartment to the other. Osmotic equilibriumis also maintained. On the other hand, thepotential across outer membranes of cellsinvolved is a diffusion potential rather than aDonnan potential. These conditions also can beembodied in suitable equations which can be
Na mEq.
H200 .
shrinkg
oK0 H20 excq.
K mEq.
20 on M
arch 31, 2020 by guest. Protected by copyright.
http://jcp.bmj.com
/J C
lin Pathol: first published as 10.1136/jcp.s3-4.1.16 on 1 January 1970. D
ownloaded from

Electrolyte and body water changes after trauma
changing state[Na] i=120: 10% cells
400
K lossmEq.
relative permeabilities Na/
Fig. 4 Potassium loss calculated to accompanyswelling of 10% of the body's cells. It is assumedthat affected cells can only pump Na at a rate justless than the influx from an external concentrationlevel of 140 to an internal concentration of120 m-equiv per litre.
Percentage Weight of Extracellular CalculatedSkeletal Muscle Skeletal Muscle Fluid of Shrinkage of
(kg) Skeletal ExtracellularMuscle (kg) Fluid (kg)
20 5-88 1-18 3-3kg[Na],= 120
100 29-4 5-88 2-2 kg [Na]l= 80
Table IL Calculated shrinkage of extracellular fluidwhen 10% of body cells swell abruptly as result ofimbalance between influx and efflux of Nal
'For comparison the volume of extracellular fluid is shown in20% of the body's skeletal muscle mass (this corresponds to 10%of body's cells) and in the whole of skeletal muscle mass.
Skeletal muscle = 42% body weight, body weight = 70 kg,and extracellular fluid = 20% wet weight.
combined (Appendix) so that it becomes possibleto predict the change in the volume of fluid incells, their content of K, and so on. In this waywe can calculate the sort of shrinkage in thevolume of extracellular fluid that would accom-pany the abrupt onset of imbalance involving 10,20, and 30% of the body's cell mass.The results of our calculations are shown in
Figure 3. They reveal that the more abnormallypermeable cell membranes are the more cellsswell and so the more the extracellular fluidshrinks. If 30% of the body's cells are involvedthe extracellular fluid shrinks to about one halfof its normal volume: when permeability is in-creased and Na pumps can only sustain pumpingat a rate nearly equal to the passive influx thatwould obtain when the concentration of Na inextracellular water [Na]0 is 140; in cell water
[Na]i 50 m-equiv/litre. The loss of K that wouldresult if 10% of the cell mass were involved isshown in Figure 4. It is substantial.
Swelling to the extent shown in Fig. 3 could,if only skeletal muscle were involved, take placeat the expense of its own tissue extracellularfluid but only if the cells involved were diffuselydistributed throughout the whole of the skeletalmuscle mass in the body (Table II). However,even were this the case, the shift of water frominterstitial fluid into cells would increase theconcentration of protein in interstitial fluid, suffi-ciently to provoke transfer of fluid from capil-laries and so to shrink the plasma volume.1 Ifthese changes are abrupt it is unlikely that theplasma will be re-expanded at the expense ofinterstitial fluid in unaffected tissues since a rapidfall in plasma volume will provoke circulatorydeterioration. The impact of this on the balanceof forces responsible for the regulation of theexchange of fluid across the capillary in varioustissues cannot be predicted.
Findings after Trauma Explained by Such CellularEvents
In my opinion cellular events such as those justdescribed could account for all the changes inbody water and electrolyte that arise aftertrauma. If this is so, present uncertainties con-cerning these changes resolve.
If gains of Na, chloride, and water and lossesof K after trauma are direct consequences ofimbalance in the exchange of water and solutesbetween cells and the fluid bathing them, the factthat such changes arise after surgery in personspreviously adrenalectomized and maintainedon constant dosage of cortisone and in patientsreceiving an aldosterone antagonist does not callfor special comment. In all patients cell imbalanceis the direct outcome of factors increasing thepermeability of cell membranes, depressing Napumnping, or altering the molar quantity of non-diffusible solutes constrained within cells and inthe balance of anionic and cationic charges onthese solutes. These events are not dependentupon increased production and release of aldo-sterone or cortisone, and it is unnecessary tospeculate on a 'permissive' role played by thesesteroids in events after trauma. The increase inaldosterone seen after trauma in persons withintact adrenals is secondary to shrinkage ofextracellular fluid caused by cellular swelling,inadequate replacement of blood lost, and so on.
'If skeletal muscle cells abruptly gain 3-3 kg water (Table II),and if interstitial fluid is initially 5 5 kg (extracellular fluid lessplasma volume where plasma volume is 1 % wet weight) the finalinterstitial fluid will drop to 2-2 kg. If the level of albumin ininterstitial fluid was initially 1-4 g per 100 ml it would become(5-5/2-2) 1-4 g per 100 ml. Its protein osmotic pressure would thenchange from approximately 4 mm Hg to 12 mm Hg (see Fig.3,1 of Landis and Pappenheimer, 1963).
21 on M
arch 31, 2020 by guest. Protected by copyright.
http://jcp.bmj.com
/J C
lin Pathol: first published as 10.1136/jcp.s3-4.1.16 on 1 January 1970. D
ownloaded from

Cecil T. G. Flear
It neither initiates nor is it responsible for reten-tion of Na, chloride, and water, or for loss of Kalthough it reinforces these events. The effectsof adequate blood replacement in abolishingretention of Na, Cl, and water and in lesseninglosses of nitrogen etc (Flear and Clark, 1955)are attributable in part to preventing or switchingoff the secondary hormonal response but largelyto its direct effect on tissue perfusion.
There are good reasons for believing that lossof nitrogen too is an outcome of cell swelling,rather than the result of increased release ofcortisone. It is known that certain euryhalineanimals excrete more nitrogen when the cellsswell in response to an increase in the fresh waterof their surroundings (Schoffeniels, 1967). Lossof nitrogen from slices of tissue in vitro is alsoknown to be closely related to uptake of fluidby their cells (Adrian, 1960; Aebi, 1952 and 1953).We do not know whether the increased loss ofnitrogen which follows trauma derives from in-creased protein catabolism or diminished proteinanabolism. Certainly, as Christensen (1948) firstpointed out, a fall in the concentration of aminoacids in cell fluid would be expected to lessenanabolism and the concentration of amino acidswill fall when cells swell for several reasons,namely, dilution from gain of water, diminishedability of cells to accumulate amino acids whenNa pumping is impaired, and some escape fromcells through abnormally leaky membranes.Just as the increased secretion of aldosteronemay be provoked by cellular happenings aftertrauma, it seems possible that release of cortisonemay be provoked by a transient fall in bloodglucose due to its entering cells whose membraneshave become unusually permeable.'
It is difficult to account for the abrupt onset ofthe hyponatraemia sometimes seen on any otherbasis. This is true also of the rise in K levelwhich not uncommonly accompanies a markedfall in plasma Na. The coexistence of Na reten-tion and fall in plasma Na is no longer para-doxical, nor is it necessary to suppose that therewas a still greater gain of water. In many instancesassociated loss of weight has caused this latterassumption to appear doubtful. Others as well asmyself have seen marked swings in plasma levelsof Na, K, and chloride after major surgery orinjuries unaccountable for by variations in gainsor losses of water or electrolyte. In my experiencethese are most likely to arise where complica-tions supervene. They may take place over amatter of hours or over a period of several days.These findings have no longer to be ignored ordismissed as 'laboratory error'.The differing and apparently contradictory
findings that have been reported concerningchanges in the volume of extracellular fluid after
'Egdahl, Mack, and Herman (1969) consider hypovolaemia tobe the best stimulus for increased secretion of ACTH. They findthat endotoxins also stimulate but less consistently.
trauma (Flear, McNeill, Greener, and Singh,1969a) are seen to derive partly from the fluctua-tions in this volume at different times aftertrauma, since events after trauma are dynamic,and measurements by different investigators havenot all been made at the same time. In part theyderive also from methodological considerations.Variable increase in membrane permeabilitymeans that markers used to measure the volumeof extracellular fluid may now penetrate intocells and may also take longer to equilibratewithin the body. Different investigators have notall used the same marker; nor indeed have theyallowed the same period for equilibration.
Singh's investigations (1965) of patientsseverely ill with intestinal obstruction established(1) that on admission two to nine days afteronset the thiocyanate space was commonly raised(11 of 13 patients). These patients had all beenat home untreated and taking little or nothingby mouth; clinically they were dehydrated andone would therefore have expected the volume ofextracellular fluid to be low. (2) After operationthe thiocyanate space rose still further in 11 of13 patients. The increase exceeded any gain ofweight over the same interval. Some did not gainweight. It seems probable, as Singh suggested,that the thiocyanate space in these patients over-estimates the volume of extracellular space dueto its entering into cells as a result of cell mem-branes being abnormally permeable. Singh (1965)also found substantial increases (from 2 to 7 kg)in thiocyanate space during the first 24 hours afterother major operations. These increases weresome 2.5 to 4 kg in excess of the carefully mea-sured water balances over the same 24-hourperiod.2 He interpreted these findings similarly.
Hinton and Allison (1969) have found thatintravenous infusions of insulin and glucosesignificantly reduce protein breakdown andpotassium loss in severely burnt patients, andpromote large diureses of Na and chloride. Theysuggest that these effects result from the actionof insulin in enhancing Na extrusion from cells(Kernan, 1962) and causing them to gain K.Glucose alone is ineffective. These findingswould be difficult to explain otherwise.
Direct Evidence for Cellular Events after Trauma
There is more direct evidence for the occurrenceof these cellular happenings after trauma.
EVIDENCE FROM TISSUE COMPOSITIONCollectively, skeletal muscle comprises some 50%of the body's cell mass. It must therefore beinvolved in any major change in the content of
2Water balance was determined as intake minus urine plus1.5 litres (estimated insensible loss and evaporative water lossfrom viscera during operation). No allowance was made for waterof metabolism.
2.2 on M
arch 31, 2020 by guest. Protected by copyright.
http://jcp.bmj.com
/J C
lin Pathol: first published as 10.1136/jcp.s3-4.1.16 on 1 January 1970. D
ownloaded from

Electrolyte and body water changes after trauma
-~0.78
~0.5
0~~~~~~~~~~~~
0 0.1 0.2
0.082 0.132
Fig. 5 Calculated average values for ext
fluid in muscle taken at the end of operati,10 patients undergoing major surgery.
The calculated average value (equationfor extracellular fluid in biopsies taken at
beginning of operation was 0-78 kg per kgdry weight of tissue. Values for extracelluat the end of operation were calculated us
9 (Appendix) and assuming values for b (tof the permeabilities PNa/PK) of 0.01 (no0-02, 0-05, 0-10, 0-125, 0-25 (increased pepof cell membranes to sodium).
water and electrolyte in the body. M(composition and volume of extracewill only be altered after trauma as
perturbation in cell metabolism or
the permeability of cell membranesstantial mass of tissue is so affectwould almost certainly have to be insuch changes. We therefore analysmuscle from 10 patients undergcvascular surgery (Flear, Pickering, ar
1969b). Three biopsies of rectus abdctaken from each patient, one at thand two at the end of the operation. Cwas taken from a portion of muscle thunder a retractor for much of the(traumatized muscle), and one fromthe same muscle that had not beenhandled (non-traumatized muscle).were cut from each biopsy from ea
Each slice was individually analysedCl, water, and fat. In all patientsabdominis gained water, Na, and Clduring operation. Similar changesnoted after injury in muscle froi(Fuhrman, 1960).Our detailed findings could only b
as resulting from cellular changes-in exchanges of water and electrolytesnet gains of Na and Cl and water anctogether with the escape of some prevdiffusible solutes from cells.
Interpretation of our findings was
detailed individual evaluation of thein composition noted between samplthe biopsies in each patient. From this
whether or not the differences reflect varyingproportions of cell and extracellular phases
Untraumatised (solids as well as fluid) in each sample. Wheredifferences do not arise in this way, but reflectdifferences in average composition of cells in
Traumatised slices, we consider it reasonable to presume thatthe difference in cell and tissue compositionreflects the varying impact throughout the tissuesampled (and in this instance also between tissuesampled at the beginning and end of operations)of the major cellular changes caused by trauma.
0.3 b What these are may be revealed by additionalscrutiny of the data. Certain patterns in the rela-tionships between sample contents of water,
racellular Na, K, and Cl have been carefully defined corre-fon from sponding to particular cellular changes. These
are based on the detailed consequences of speci-8, Appendix) fied changes in cell composition which are pre-the dicted with the aid of the equations set out inrfat-free the Appendix.' Data are therefore examined totlar fluid see which of these patterns is displayed. It ising equation necessary to adopt this approach because unlessrmal) and a balanced state obtains across cell membranesrmeability the volume of extracellular fluid in pieces of
skeletal muscle cannot be calculated2 so that it isnot possible to partition total content into cellularand extracellular contributions. In a changingstate with cells gaining Na the volume of extra-relover,the cellular fluid calculated for a sample of given
auarr ltuof composition depends upon the permeabilityincrease in
assumed (Appendix). We were, however, able to
yif a sub-n conclude from our appraisal of patterns dis-
ed Muscle played that (1) a balanced state had not been re-ed. Mucle attained at the end of operation as cells were stillned skeletal gaining Na; (2) on average skeletal musclese.gsmaje.a showed neither net gain nor loss of extracellulardi majNiolr fluid. We were, therefore, able to say what the)i McNeil, average volume of extracellular fluid was at thec beginning end of operation, since a reasonable estimate)ne of these
could be made of its average value in samples)ne of these taken at the beginning of the operation. As aoperation result, we were able to deduce that permeabilityoperation of cell membranes increased during operation,
an area of and that the extent of this increase was moreexposed or marked in the area of muscle that had beenTen tliets damaged during the operation (Fig. 5).lch patent. Further investigations will be necessary tofor Na, K, establish for how long after operations cellsthe rectus continue to gain Na and lose K. We also need toand lost Khave beenm animals 'The derivation of some of these patterns has been presented
and discussed elsewhere (Flear, 1969; Flear et al, 1969b).'The simple calculation of extracellular fluid used by Graham,
e explained Lamb, and Linton (1967) is unreliable. It is necessary to assume a
-imbalances value for the resting membrane potential; Graham suggests- 85 mV. It can, for instance, be shown that the membranes leading to potential will be very much less than 85 mV during a period ofA loss of K, continuing net gain of Na provoked or accompanied by increase
in cell membrane permeability. Moreover the membrane potential,iously non- has been found at times to be very low after a period of net gain
of Na by cells (in vitro) when a balanced state may be presumedased on a to be re-established (Cross, Keynes, and Rybova, 1965). Finally,baseu on a calculations from our own data suggest that membrane potentialsdifferences vary substantially in alimentary disease (TableIII). They suggest,es from all too, that there is a marked fall in potential during major opera-tions. Calculations from data in reference (Flear et al, 1969b)we deduced indicate an average fall of 50 mV.
23 on M
arch 31, 2020 by guest. Protected by copyright.
http://jcp.bmj.com
/J C
lin Pathol: first published as 10.1136/jcp.s3-4.1.16 on 1 January 1970. D
ownloaded from

Cecil T. G. Flear
in vitro dilution
5 15 25 35 45
mins.
Fig. 6 Sodium content ofbundles offibres dissectedat the times indicatedfrom a biopsy of rectusabdominis immersed in an oxygenated saline at22°C (for composition see unpublished data ofGreener and Flear). Each point is the mean offour or five bundles.
establish whether skeletal muscle in general, ie,from parts of the body remote from the area ofoperation, show these same changes.
Swedish investigators report loss of 3% Kfrom adductor muscle in the thigh of dogsshocked by blood loss (Hagberg, Haljamae, andRockert, 1968). This compares with an averageloss of 3-5 % K in our patients (Flear et al, 1969).The interval between taking control and post-shock samples was two and a quarter hours, atime which is very similar to the average durationof our operations. They suggest that loss of Kactually is more than this but that most of thelost K stays in the interstitial fluid and so isanalysed (Hagberg et al, 1968). They find thatsingle muscle fibres contain 10 to 50% less Kafter shock than before bleeding, and thatbundles of fibres contain from 0 to 35% less Kafter shock (Hagberg, Haljamae, and Rockert,1967). Individual fibres and bundles alike weredissected with the biopsy immersed in Ringersaline at 20°C which had been oxygenated for10 minutes with 95% 02, 5% CO2 (Haljamae,1967). The Swedish investigators consider thatthis greater loss of K resulted from washingaway K trapped in interstitial fluid. Dr Greenerand I, however, find that bundles of humanskeletal muscle fibres dissected in similar Ringersolution (at 22°C) and gassed for one hourrapidly gain Na and water and lose K (Fig. 6).Their dissections took from five to 40 minutes.Our biopsies were taken at the beginning ofoperations, when there could be no question ofsignificant blood loss or shock. We believe thatthe greater loss of K found in bundles and fibres
Fig. 7 Relationship between osmolality (correctedfor osmotic contribution made by urea and glucose)- [Osm]wc, (mOsm/kg plasma water) and the levelofNa in plasma - [Na]w, in m-equiv per kg plasmawater.
from biopsies taken after shock, compared withbundles taken similarly from pre-shock biopsies,reflects an increase in cell membrane permeabilitybrought about by shock. Consequently Na entersmore rapidly into post-shock fibres and loss of Kwill be more marked.
EVIDENCE FROM RELATIONSHIP BETWEENPLASMA OSMOLALITY AND LEVEL OFSODIUMIt was pointed out earlier that when a sufficientmass of tissue cells gain Na and chloride and atthe same time some previously non-diffusiblesolutes escape from cells the level of Na through-out extracellular fluid will fall without a fall inosmolality. Dr Singh and I have found that thisis in fact what happens when shock is experi-mentally induced in dogs by blood loss and aftersurgery in man.We first defined the relationship between
osmolality and the plasma level of Na whenchange in both is brought about by dilution(Singh and Flear, 1968). Since levels of urea andglucose can vary substantially in patients,osmolality was corrected by subtracting theosmotic contribution made by each of thesesubstances. Osmolality and the level of Na weremeasured in plasma (also urea, glucose, andwater) before and after diluting it in vitro byadding varying amounts of 5% dextrose. Similardilution studies were made by infusing 5%dextrose intravenously and taking samples ofblood from a vein some 3 to 6 inches proximalto the point of insertion of the drip needles. The
300
0)
0)
zulE
200
100
24 on M
arch 31, 2020 by guest. Protected by copyright.
http://jcp.bmj.com
/J C
lin Pathol: first published as 10.1136/jcp.s3-4.1.16 on 1 January 1970. D
ownloaded from

25Electrolyte and body water changes after trauma
blood loss shock
[Osm]c
120 140 160 120 140 160
[Na]W
Fig. 8 Relationship between plasma levels ofNa(m-equiv/kg plasma water: [Na],,,) and osmolalitycorrectedfor urea and glucose (mOsm/kg plasmawater: [Osm]wc) before (open circles) and duringthe development of blood loss shock in two dogs.The relationship caused by dilution is projectedfromthe pre-shock values. The 95% confidence limitswithin which observations should be, if the fall inNa is caused by dilution, are also indicated (un-published data, Singh and Flear).
2201""
Fig. 9 Plasma levels ofNa and osmolality (seeFig. 8) before surgery and at intervals thereafter for
three weeks. The band is derived by fitting theaverage slope ofdilution to the outermost valuesobserved in a group of50 healthy subjects (Singhand Flear, unpublished data).
rate of infusion was varied. Close agreement was
noted between findings with both methods, andbetween the relationship found in differentsubjects. Figure 7 shows the relationship notedwhen plasma from one subject was diluted in vitro.
Observations were then made in 12 dogs beforeand while inducing shock by bleeding (Singh andFlear, 1969). Systolic blood pressure was droppedin all cases to 40 mm Hg or less and held at thatlevel for one or more hours. Total amounts ofblood removed ranged from 1-8 to 2.8 litres.Findings were essentially the same in all dogs:the plasma Na level fell without a comparablefall in osmolality. Figure 8 shows data from twodogs. The oblique line depicts the relationshipbetween Na and osmolality when change in bothis brought about by dilution. This r ;lationshipwas determined in all instances on blood takenbefore shock was induced. Figure 9 presentssimilar findings from a patient before, during,and after colectomy. We have similar findingsafter surgery in many other patients. Neither inexperimental shock nor after surgery can thelack of a comparable fall in osmolality beexplained by the rise in K level commonly seen.
These findings clearly demonstrate that hypo-natraemia in shock is largely provoked byimbalance across cell membranes. This factoris of major importance also in the onset of hypo-natraemia after surgery. Here, dilution alsocontributes to its onset and at times may be theoverriding factor. This was Wynn's experience(Wynn and Robb, 1954; Wynn and Houghton,1957). Most of his patients had received very
generous transfusions of 5% dextrose, to thehazards of which he directed attention. PlasmaK levels fell in his patients.
OTHER EVIDENCE
A final piece of evidence that trauma provokescellular gain ofNa is afforded by the observationsthat resting membrane potentials of skeletalmuscle fibres, measured in situ, fall during theonset of shock induced in rats by bleeding (Shiresand Carrico, 1966). More recently Shires and hiscolleagues also found that in man too membranepotentials (measurements in situ) were low whenpatients were admitted in traumatic shock, andthat with recovery the potentials rose (Shires,1969).We have confirmed in vitro that while human
skeletal muscle cells are accumulating Na, theirmembrane potential falls (Table III), and that
Controls Peptic Regional Ulcerative(3) Ulcer Enteritis Colitis
(13) (12) (14)
Maximum variation(mV) within onemuscle in a givensubject 10 23 19 36Highest and lowestpotentials (m/V)noted in each group 87-99 60-100 81-104 80-124
Table ILL Variation in calculated membrane potentials(rectus abdominis) in controls and in patients withalimentary diseasel
'Unpublished data of Flear, Florence, and Williams calculatedfrom tissue contents of K, Cl, water; and concentrations K and Clin extracellular water. Equation 8 in the appendix was used.Biopsies were all taken at the beginning of operations.The potential calculated for each sample of tissue is the average
for all fibres in the sample. These potentials will therefore showless variation than would be apparent from actual measurementson individual fibres due to the smoothing effect of averaging.From 10 to 30 observations were made in each individual.
on March 31, 2020 by guest. P
rotected by copyright.http://jcp.bm
j.com/
J Clin P
athol: first published as 10.1136/jcp.s3-4.1.16 on 1 January 1970. Dow
nloaded from

Cecil T. G. Flear
after re-establishment of a new balanced state ofnet Na gain the potentials are also subnormal(Flear, Greener, and Ferry, 1970).
Causes of Cellular Happenings after Trauma
3230
[Osm]I 21
end1
L0
In addition to the evidence presented in the FV_previous section of imbalance in exchanges of 240 _water and electrolytes between cells and extra- Kcellular fluid, there is substantial evidence that 120trauma disturbs the metabolism of cells. Theamounts of a number of metabolites are alteredin injured tissues (Stoner and Threlfall, 1960; Fig. 10 Pla.Tremoliere and Derache, 1960), and these Fig. 8) beforechanges must affect the molar quantity of non- by injecting i,diffusible solutes constrained within cells, the of killed E. cbalance of charges on them, and consequentlyalso the amounts of water and cations in cells.Probably all cellular events after trauma are
partly caused by hypoxia and tissue under- 1600perfusion. Ischaemia of skeletal muscle, heart, 82(Lamprecht, 1967) is known to affect levels Br cpmof ATP etc in tissues in animals and man.
Whilst the shift to anaerobic metabolism in themyocardium following the- sudden arrest of the 1400coronary blood flow octurs in an oscillatoryfashion (Wollenberger and Krause, 1968), itis likely to be accompanied by similar fluctua-tions in the amounts of various compounds withincells. Moreover, ischaemia of the lower limbs 1200in animals provokes breakdown of glycogen inthe liver and skeletal muscle elsewhere in thebody. Anoxaemia induced by breathing nitrogenprovokes a considerable upset of metabolism,as judged by the resultant rapid increase in Fig. 11 Cososmolality of body fluids (Battaglia, Meschia, injectingsuHelligers, and Baron, 1958). It has also been into a dogtoshown to increase the permeability of cells in SO mm Hsitu and in vitro, and in vitro to inhibitextrusion of Na and to provoke breakdown oflabile compounds. Elsewhere I have presentedevidence from patients that skeletal muscle cells Other evidcin ischaemic limbs show changes similar to those on 16 dogsinduced by anoxia in vitro (Flear, 1969: see Fig. 9). given E. cMore recently in nine patients operated upon by Flear, and ]Mr Donald Ross, Dr Singh and I noted striking 82Br by injegains of Na and Cl by ventricular heart muscle Over a con
cells, losses of K, and variation in the amounts in plasma v
of non-diffusible anions following peroperative 82Br countanoxic arrest of the heart (Singh, Flear, and Ross, of the dog1969). Losses of K, for instance, ranged from 35 We concluto 168 m-equiv per kilogram fat-free dry weight, extracellulaan average loss of 30 %. that it cont
Other factors may also provoke imbalance in endotoxinsthe exchange of water and electrolytes across cell tion of wa
membranes. My colleagues and I have evidence branes. Mthat E. coli endotoxin provokes a considerable variabilityincrease in the permeability of a substantial mass which, we
of body cells. Figure 10 presents observations to will gain sithis effect on the plasma levels of Na and osmol- it is thereality in two dogs before and after they had been shrinkage ishocked by infusing a killed suspension of E. coli. However, t
dotoxic shock
2
1~~~~~~~~~~~~~~~~
140 160 120 140 160
[HaW~ma levels ofNa and osmolality (seee and after inducing shock in two dogsintravenously aliquots ofa suspension,oli in water.
endotoxin. . ~ .0
0 _
% %*0
Vi0 50 100 150 200
time -mins250
unts of 82Br in plasma before and aftericient E. coli endotoxin (strain CA 38)7 lower systolic bloodpressure to
ence to the same effect was obtained;, splenectomized 10 days before beingcoli endotoxin intravenously (Batey,McNeill, 1970). These dogs were givenection some 12 to 15 hours previously.itrol period the concentration of 82Brwas constant. During shock, the meanper millilitre of plasma in all but one
,s rose above control levels (Fig. 11).ide that this is due to shrinkage ofar fluid around the quantity of 82Brtains. This effect would be expected ifcaused disequilibration in the parti-
Lter and electrolytes across cell mem-
[oreover, after giving endotoxin thebetween plasma counts increased
believe, supports our inference. Cells,ome 82Br along with Na and Cl, andfore not possible to quantitate thein extracellular fluid by this technique.the haematocrit was seen to rise after
1 *v *a a
26
l l
on March 31, 2020 by guest. P
rotected by copyright.http://jcp.bm
j.com/
J Clin P
athol: first published as 10.1136/jcp.s3-4.1.16 on 1 January 1970. Dow
nloaded from

Electrolyte and body water changes after trauma
injecting endotoxin, affording evidence thatplasma volume shrinks.
E. coli endotoxin has also been observed toincrease the permeability of skeletal muscle toNa. Uptake of Na by frog sartorius muscle (at0°C) is faster from plasma taken from dogsduring endotoxic shock than from plasma takenbefore injecting endotoxin (Greener, Singh, andFlear, 1969). When the findings were presentedto the Surgical Research Society they weremerely suggestive. They have since been expandedand shown to be significant. Evidence has alsobeen obtained that E. coli endotoxin similarlyincreases the uptake of Na in vitro by humanskeletal muscle (at 0'C) from the person's ownplasma, and also that it provokes striking changesin the molar quantity of non-diffusible organicsolutes constrained within cells. It is of relatedinterest that, according to Kovach and Fonyo(1960), Boda found that serum and cerebrospinalfluid in infants in toxicosis inhibits glucose up-take by rat diaphragm.
Finally, Graber and I (ms. only) concludedfrom detailed balance studies that the abruptclinical onset of endotoxic shock in a youngwoman was associated with cell gain of Na andCl and loss of K. These changes continued untilthe toxic period, as well as the period of circul-atory instability, had been successfully negotiated.During recovery they reversed. It was also notedthat the marked swings in plasma Na could notbe explained as simple consequences of gains orlosses of water and cations (Table IV).
Phase (No.) Extracellular Water(m-equiv/days)
Circulatory collapse (1) 205Toxic (2) 44Recov_ry (3) 18
(4) 6(5) 4
Table IV Discrepancy between observed and expectedbalance ofNa + Kfrom daily water balance' and theconcentration (m-equiv (Na + K)16) extracellular waterin one patient'Graber and Flear (1970).
The possibility that other factors associatedwith trauma and its sequelae may provoke im-balance in exchanges across cell membranesshould not be ignored. For instance, cate-cholamines increase and there is evidence thatthey affect membranes and Na pumps.
Wider Relevance of Cenular Events and ofInvestigations of Trauma
I would conclude by affirming my belief that thechanges in water and electrolytes, and in nitrogenmetabolism also, that we see after trauma are theresult of perturbations or sustained dislocationsof cell metabolism and the permeability of their
27
membranes. I suggest that increased productionand release of adrenal steroids, and po3siblyADH, are consequences rather than causes ofdisturbances in water and electrolytes. If this isso, then I believe that it raises questions oftherapeutic importance, and also that researchesinto events which follow trauma perhaps assumeadded significance.
QUESTIONS OF THERAPEUTIC RELEVANCEPerhaps the most obvious of these concerns whatcan be done to lessen or prevent, and also toreverse, changes in membrane permeability,active transport of Na, in the molar quantity ofnon-diffusible solutes, including anions, or tolessen their consequences for water and electro-lyte partition.As we have seen, the consequences of cellular
events vary. Abrupt swelling of cells causes anequally rapid shrinkage of extracellular fluid,and it seems probable that these events could bethe mechanism underlying the abrupt onset ofshock which may develop in septic and toxicsituations. Shrinkage of plasma as a result ofshift of fluid into the interstitial fluid, provokedby the concentrating effect on interstitial fluidprotein levels of cell swelling, is unlikely to beseriously opposed by an increase in tissue pressure.Observations on skin (Guyton and Coleman,1967), have shown that after an initial increase inpressure tissue becomes strikingly pliant so thatmarked swelling is tolerated with little or nofurther rise in pressure. In the case of skeletalmuscle, cell swelling will produce a fall in restingmembrane potential, whence loss of muscle tone,so that tissue pressure may even fall.The use of hypertonic saline has been suggested
as a means of lessening cellular swelling (Brooks,Williams, Manley, and Whiteman, 1963);efficacy is lessened or even cancelled by the greaterinflux of Na into cells caused by a raised level ofNa in extracellular fluid. Probably salt-freealbumin would be the most appropriate infusionfluid. It would lessen plasma volume shrinkageand not increase the influx of sodium.
It seems likely that tissue underperfusion inshock will promote a vicious spiral of cell swell-ing, shrinkage of plasma volume, or furtherunderperfusion. I would suggest that the reducedblood flow to skeletal muscle seen in shock isthus of greater significance than has been thoughthitherto since it may provoke such a spiral.Accumulation of Na in heart muscle cells is of
special significance. It causes a fall in restingmembrane potential. A slight fall increases theexcitability of fibres, and if this happens un-evenly throughout the myocardium it may causearrhythmia. Greater fall causes a fibre to becomeinexcitable, and, if a sufficient proportion of thehearts fibres are so affected, cardiac output mustfall. I believe that these events underly complica-tions not infrequently seen after heart surgery,
on March 31, 2020 by guest. P
rotected by copyright.http://jcp.bm
j.com/
J Clin P
athol: first published as 10.1136/jcp.s3-4.1.16 on 1 January 1970. Dow
nloaded from

Cecil T. G. Flear
particularly the complication associated withanoxic arrest of the heart.
Hinton and Allison (1969) report the successfuluse of infusions of insulin and glucose aftertrauma and suggest that insulin is the activefactor promoting extrusion of sodium from cells.Kernan (1962) presented evidence for such anaction by insulin, and it would seem worthassessing its value after heart surgery. It has ofcourse been used for this purpose after myocardialinfarction (Sodi-Pallares, Bisteni, Medrano,Micheli, Ponce de Leon, Calva, Fishleder,Testelli, and Miller, 1966; Sodi-Pallares, Poncede Leon, Bisteni, and Medrano, 1969), inrefractory congestive heart failure (Brenner,1960), and in terminal carcinoma associated withhyponatraemia (Neufeld, 1962).My interpretation of events after trauma raises
the question of what should be our attitude to-wards hyponatraemia and hyperkalaemia. Ifthese disturbances arise as outlined serious con-sideration should be paid to the possibility thatthey are more appropriate levels than 'normal'ones would be in the prevailing circumstances. Ifthis is so attempts should not be made to alterthem directly. Elsewhere, discussing events incongestive heart failure, it was pointed out thatboth low Na and high K levels facilitate transportof Na from cells (Flear, 1969).
WIDER SIGNIFICANCE OF RESEARCH INTOEVENTS AFTER TRAUMAThere is an increasing awareness of the relevanceof changes in cell membranes and Na pumps, inmany situations. Some hormones (Christensen,1962) and drugs (Burgen, 1967) act at this level,and disturbances in active transport of Na from
erythrocytes have been reported in uraemia (Welt,Sachs, and McManus, 1964; Welt, Smith, Dunn,Czerwinski, Proctor, Cole, Balfe, and Gitelman,1967; Smith, Welt, and Czerwinski, 1967),malaria (Dunn, 1968), and thyrotoxicosis (Smithand Samuel, 1970). Elsewhere I have discussedevidence for alterations in permeability, etc, inprogressive heart failure (Flear, 1966, 1969).
Insight gained into disturbances at this levelafter trauma will thus be of wide interest. More-over, trauma offers the advantage of being adisturbance with a discrete and known abruptonset in time. It has the feature too of arising inboth the previously healthy and the chronicallyill: in both its consequences may be reversible.It does of course pose the problem of research inthe midst of intense activity directed at thesurvival of the patient.
Researches have been supported by the RoyalSociety, the Scientific and Research Sub-Com-mittee of the Royal Victoria Infirmary, Newcastle,and Muscular Dystrophy Group of the UnitedKingdom, The Wellcome Foundation, and GeigyPharmaceuticals.My thanks are due to Miss Barbara James and
others of the University Department of Photo-graphy for help with the figures. I should alsolike to record my appreciation of the many help-ful discussions with many colleagues, in par-ticular, George Graber, Ian McNeill, ChanderMohan Singh, and John Greener.Dr Harry Richardson (Department of Micro-
biology, University of Newcastle) kindly suppliedthe suspension of killed E. coli.
Appendix
Prediction of the effect of variation in cellular content of sodiumupon the amount of water and potassium in cells
During a Balanced State
This is a state of zero net flux of water and ions,and here the partition is in accord with the follow-ing equilibria.
Electrical neutralityzA
[K]i -- [Na]i = [Cl]i- . . . 1
for osmotic balance across the cell membraneA
[K]i + [Na]i + [C]i + = C . . . 2
for a Donnan equilibrium[K]u [Cli = [K]o [Cl]o . . . 3where [ ]i denote concentrations in cell water;[ ]0 in extracelluJar water.
28 on M
arch 31, 2020 by guest. Protected by copyright.
http://jcp.bmj.com
/J C
lin Pathol: first published as 10.1136/jcp.s3-4.1.16 on 1 January 1970. D
ownloaded from

Electrolyte and body water changes after trauma
V represents the volume of cell water, and istaken as unity in the initial resting circumstance.A represents mols of non-diffusible material
in V.zA represents the sum of the electrical charges
on A, represented as milliequivalents of anions(z is average valence of A).C represents total external concentration less
that due to freely diffusible substances, such asurea, present in equal concentration on eitherside of cell membranes. Equations 1, 2, and 3 maybe combined to give an equation which can beused to predict the outcome of changes in one ormore of the terms in equations 1, 2, and 3.
V2 {C2 - 4[K]o [Cl]0 + 2[Na]i C} +V {2[Na]i A(I - z) - 2CA} +
A2(1 - Z2) = 0 . . . 4Equation 4 can be used with equation 5 below,
to predict the effects of cell content and concen-tration of Na, K, chloride on A and zA and oncell content of water of (1) gain of sodium bycells, (2) change in A, and (3) change in zA eithersingly or jointly (Flear, 1969). The procedure isto substitute the postulated new value of [Nalietc in equation 4 and solve for V.The value [Cl]I is then derived from the
equation2[Cl]1= C-(A/V) (1 -z) . . . 5
obtained by subtracting equation 1 from equation2. [K]i is then obtained using equation 3; A/Vfrom equation 2, etc.
During a Changing State of Net Na Gain by Cells
Here equations 1 and 2 obtain, but as the poten-tial across cell membranes is a diffusion potentialequation 3 is invalid. Instead we may write
[K]i (Cl]i = [K]o [C1]0 + b[Na]0 [Cl]o-b[Na]i [Cl]1 . . . 6
where b is the ratio of the permeability factorsfor K and Na (PK/PNa).Combining equations 1, 2, and 6 one can
obtain
V2 {C2 -2(1 - b) [Na]iC - 4([K]o [Cl]o+b[Na]0 [Cl]0)}+2V {(1 -b) ]Na]i A-(1 -b) [Na]izA -CA}
+ A2(1 - (Z)2) = 0 . . . 7which is analogous to equation 4.
It has been used to predict the consequencesof imbalance between influx and efflux of sodium,first assuming no change in the permeability ofcell membranes (b = 0-01) and subsequentlyassuming the following values for b = 0.1, 0.5,and 1-0, that is, assuming that cell membranesbecome abnormally permeable.
Separate calculations were made for the three
29
situations where the rate of Na pumping is nearlyequal to the passive influx that would obtainwhen [Na]0 = 140 m-equiv/l and [Na]i was 50,80, and 120 m-equiv/l. The procedure was asindicated in previous sections. The assumed initialcomposition of cellular and extracellular fluidswas
[Nali = 10; [K]I [Cl]i = 500; [K]i = 160;A = 117; zA =-167; [Na]0 = 140; [K]o =5; [Cl]o = 100; C = 290.
The composition of extracellular fluid waspresumed to remain constant.The effects were then calculated of abrupt
swelling of cells, of the order calculated to arisein the above situations, on the volume of extra-cellular fluid, and on body K, in a 70 kg adultwhose 1H20= 60% body weight, 2 intracellularfluid = 301 £ extracellular fluid - 121.
These effects were calculated assuming (1) 10 %,(2) 20 %, and (3) 30% of the body's cell mass wasimplicated. Skeletal muscle was presumed tocomprise 50% of the body cell mass. Pertinentresults are presented in Figures 3 and 4.
Calculation of Extracellular Fluid Volume fromTissue Contents of Na, K, Cl, and Water
Conway (1957) first showed that, providing abalanced state obtained, the volume of extra-cellular fluid could be calculated from tissuecontents of K, Cl, and water, plus the concen-trations of K and Cl in extracellular water usingthe equationVo = (KtClt -[K]o [Cl]0 Vt2)/(Kt[Cl]0 + Clt[K]o - 2Vt[K]o [Cl]0) . . . 8
where Vo = kilograms of extracellular fluid perkilogram fat-free dry weight of tissue.Kt and Clt denote tissue contents in milli-
equivalents per kilogram fat-free dry weight oftissue
Vt = kilograms of tissue water per kilogramfat-free dry weight.
Other symbols are defined above. (See refer-ence for a discussion of the derivation of theformula used. It is based on equation 3 above.)
In similar fashion it can be shown that during achanging state of sodium gain, the volume ofextracellular fluid can be calculated from tissuecontents of Na, K, Cl, and water, and the con-centration of these ions in extracellular water:V0 = {(KtClt -[K]o [Cl]o Vt2) +b(NatClt - [Na]0 Cl0 Vt2)} /{(Kt[Cl]o + Clt[K]o - 2Vt[K]o [Cl]0) +b(Nat[C1]0 + Clt[Na]0 - 2Vt[Na]o [Cl]o}
.9Nat = milliequivalents Na per kilogram fat-
free dry weight of tissue.
on March 31, 2020 by guest. P
rotected by copyright.http://jcp.bm
j.com/
J Clin P
athol: first published as 10.1136/jcp.s3-4.1.16 on 1 January 1970. Dow
nloaded from

Cecil T. G. Flear 3
References
Adrian, R. H. (1960). Potassium chloride movement and themembrane potential of frog muscle. J. Physiol. (Lond.),151, 154-185.
Aebi, H. (1952). Ueber die chemische und morphologischeNatur der Stickstoffabgabe von Gewebsschnitten imWarburg-versuch. Biochim. biophys. Acta. (Amst.), 9,443-456.
Aebi, H. (1953). Elektrolyte Akkumulierung und Osmoregulationin Gewebesschnitten. Helv. physiol. Acta, 11, 96-121.
Andres, R., Cader, G., Goldman, S. P., and Zierler, K. L. (1957).Net potassium movement between resting muscle andplasma in man in the basal state and during the night. J.clin. Invest., 36, 723-729.
Batey, N. R., Flear, C. T. G., and McNeill, I. F. (1970). The effectof E. coli endotoxin on extracellular fluid volume. Com-munication to the Surgical Research Society Meeting,Glasgow, January, 1970.
Battaglia, F. C., Meschia, G., Hellegers, A., and Barron, D. H.(1958). The effects of acute hypoxia on the osmotic pres-sure of the plasma. Quart. J. exp. Physiol., 43, 197-208.
Bernal, J. D. (1965). The structure of water and its biologicalimplications, pp. 17-31. In The State and Movement ofWater in Living Organisms. S.E.B. Symposium XIX 1965.
Boyle, P. J., and Conway, E. J. (1941). Potassium accumulationin muscle and associated changes. J. Physiol. (Lond.), 100,1-63.
Brenner, 0. (1960). The management of congestive heart failure.In Modern Trends in Cardiology, edited by E. MorganJones, p. 82. Butterworths, London.
Brooks, D. K., Williams, W. G., Manky, R. W., and Whiteman,P. (1963). Osmolar and electrolyte changes in haemorrhagicshock-hypertonic solutions in the prevention of tissuedamage. Lancet, 1, 521-527.
Brown, F., and Stein, W. D. (1960). Balance of water, electrolytes,and nonelectrolytes. In Comparative Biochemistry, Vol. 2,edited by M. Florkin, and H. S. Mason, pp. 403-470.Academic Press, New York.
Burgen, A. S. V. (1967). The effect of drugs on membrane trans-port. Proc. roy. Soc. Med., 60, 329-334.
Christensen, H. N. (1948). The distribution of amino acidsbetween cellular and extracellular fluids. Relation togrowth. Bull. New Engi. med. Cent., 10, 108-1 1 1.
Christensen, H. N. (1962). Biological Transport. Benjamin, NewYork.
Conway, E. J. (1945). The physiological significance of inorganiclevels in the internal medium of animals. Biol. Rev., 20,56-72.
Conway, E. J. (1957). Nature and significance of concentrationrelations of potassium and sodium ions in skeletal muscle.Physiol. Rev., 37, 84-132.
Cross, S. B., Keynes, R. D., and Rybova, R. J. (1965). Thecoupling of sodium efflux and potassium influx in frogmuscle. J. Physiol. (Lond.), 181, 865-888.
Diamond, J. M., and Wright, E. M. (1969). Biological membranes:the physical basis of ions and nonelectrolyte selectivity.Ann. Rev. Physiol., 31, 582-646.
Dunn, M. J. (1968). Aberrations of red blood cell sodiumtransport during malarial infection. (Abstr.) Clin. Res.,16, 382.
Egdahl, R. H., Mack, E., and Herman, A. (1969). The effect ofhaemorrhage on adrenocortical perfusion and function.(Abstr.). Brit. J. Surg., 56, 703.
Flear, C. T. G., and Rajasingham, M. S. Unpublished observa-tions.
Flear, C. T. G. (1966). Disturbance of volume and compositionof body fluids in congestive heart failure. In Electrolytesand Cardiovascular Diseases, edited E. Bajasz, vol. 2,p. 357. Karger, Basel, New York.
Flear, C. T. G. (1969). Alterations in water and electrolyte dis-tribution in congestive heart failure and their significance.Ann. N. Y. Acad. Sci., 156, 421-444.
Flear, C. T. G. (1967). Significance of potassium and sodiummasses in cardiac disease. In Compartments Pools andSpaces in Medical Physiology, edited by P-E. E. Bergner,p. 53, and C. C. Lushbaugh (see pp. 53, and 123-138). U.S.Atomic Energy Commission Division of Technical In-formation, 1967.
Flear, C. T. G., and Clark, R. (1955). The influence of blood lossand blood transfusion upon changes in the metabolism ofwater, electrolytes, and nitrogen following civilian trauma.Clin. Sci., 14, 575-599.
Flear, C. T. G., Greener, J. S., and Ferry, C. B. (1970). Activetransport of sodium from human skeletal muscle: an invitro technique and findings. In 5th Symposiuim on CurrentResearch in Muscular Dystrophy and Related Diseases.London.
Flear, C. T. G., McNeill, I. F., Greener, J. S., and Singh, C. M.(1969a). Crystalloid administration in shock and surgicaltrauma. (Letter.) Lancet, 2, 155.
Flear, C. T. G., Pickering, J., and McNeill I. F. (1969b). Observa-tions on water and electrolyte changes in skeletal muscleduring major surgery. J. Surg. Res., 9, 369-382.
Fuhrman, F. A. (1960). Electrolytes and glycogen in injuredtissues. In The Biochemical Response to Injurjy, edited byH. B. Stoner and C. J. Threlfall, p. 5. Blackwell, Oxford.
Graber, I. G., and Flear, C. T. G. (1970). Water electrolyte andnitrogen balance studies in the successful management ofa case of severe endotoxic shock in the puerperium, com-plicated by staphylococcal enteritis. (In manuscript.)
Graham, J. A., Lamb, J. F., and Linton, A. L. (1967). Measure-ment of body water and intracellular electrolytes by meansof muscle biopsy. L.,ncet, 2, 1172-1176.
Greener, J., Singh, C. M., and Flear, C. T. G. (1969). Doesplasma from shocked or uraemic dogs contain a factorwhich increases permeability of skeletal muscle-cell mem-brane. (Abstr.) Brit. J. Surg., 56, 623.
Guyton, A. C., and Coleman, T. G. (1967). Long term regulationof the circulation: interrelationships with body fluidvolumes. In The Physical Bases of Circulatory Transport,edited by E. B. Reeve and A. C. Guyton, ch. II. Saunders,Philadelphia, London.
Haljamae, H. (1967). Shock reactions in skeletal muscle. I. Amethod for determining the potassium content of singleskeletal muscle cells. Acta chir. scand.. 133, 259-363.
Hagberg, S., Haljamae, H., and Rockert, H. (1967). Shock re-actions in skeletal muscle, II. Intracellular potassium ofskeletal muscle before and after induced haemorrhagicshock. Acta chir. scand., 133, 265-268.
Hagberg, S., Haljamae, H., and Rockert, H. (1968). Shock re-actions in skeletal muscle. III. The electrolyte content oftissue fluid and blood plasma before and after inducedhaemorrhagic shock. Ann. Surg., 168, 243-248.
Hinton, P., and Allison, S. P. (1969). Crystalloid administrationin shock and surgical trauma. Lancet, 2, 594-595.
Jacobs, M. H. (1935). In Diffusion Process, 1967. Springer-Verlag,Berlin and New York. (Reprinted from Ergebnisse derBiologie Zwolfter Band, 1935.)
Kernan, R. P. (1962). The role of lactate in the active excretion ofsodium by frog muscle. J. Phvsiol. (Lond.), 162, 129-137.
Kovach, A. G. B., and Fonyo, A. (1960). Metabolic responses toinjury in cerebral tissue. In The Biochemical Response toInjury, edited by H. B. Stoner and C. J. Threfall, p. 129.Blackwell, Oxford.
Kusel, H., and Netter, H. (1952). Beweis der Conway-Theorieuber die lonenverteilung im Muskel. Biochem. Z., 323,39-52.
Lamprecht, W. (1967). Substrate and energy metabolism in heartfailure. In an International Symposium on CoronaryCirculation and Energetics of the Myocardium, Milan,1966, p. 186. Karger, Basel, New York.
Landis, E. M., and Pappenheimer, J. R. (1963). Exchange ofsubstances through the capillary walls. Ch. 29 in Handbookof Phvsiology. Circulation Vol. 2. American PhysiologicalSociety.
Langer, G. A. (1968). Ion fluxes in cardiac excitation and con-traction and their relation to myocardial contractility.Physiol. Rev.. 48, 708-757.
Leaf, A. (1956). On the mechanism of fluid exchange of tissuesin vitro. Biochem. J., 62, 241-248.
Ling, G. N., and Ochsenfeld, M. M. (1966). Studies on ionaccumulation in muscle cells. J. gen. Physiol., 49, 819-843.
Lockwood, A. P. M. (1963). Animal Body Fluids and Their Regula-tion. Heinemann, London.
Lukowsky, G., and Netter, H. (1952). Zur Theorie der Kalium-schwelle an der Muskulatur. Biochem. Z., 323, 53-66.
McDowell, M. E., Wolf, A. V., and Steer, A. (1955). Osmoticvolumes of distribution. Idiogenic changes in osmoticpressure associated with administration of hypertonicsolutions. Amer. J. Physiol., 180, 545-558.
McNcill, 1. F., and MacKenzie, A. Unpublished observations.Mond, R. (1955). Transport von K, Cl Wasser durch die Muskel-
fasermembran. Pflugers Arch. ges. Physiol., 261, 243-248.Mullins, L. J. (1959). The penetration ofsome cations into muscle.
J. gen. Physiol., 42, 817-829.Mullins, L. J. (1960). An analysis of pore size in excitable mem-
branes. J. gen. Physiol., 43, Suppl. 1, 105-117.Mullins, L. J., and Frumento, A. S. (1963). The concentration
dependence of sodium efflux from muscle. J. gen. Physiol.,46, 629-654.
Mullins, L. J., and Awad, M. Z. (1965). The control of themembrane potential of muscle fibers by the sodium pump.J. gen. Pkvsiol., 48, 761-775.
Netter, H. (1969). Theoretical Biochemistry, Oliver and Boyd,Edinburgh.
on March 31, 2020 by guest. P
rotected by copyright.http://jcp.bm
j.com/
J Clin P
athol: first published as 10.1136/jcp.s3-4.1.16 on 1 January 1970. Dow
nloaded from

Electrolyte and body water changes after trauma 31
Neufeld, 0. (1962). Insulin therapy in terminal cancer: a pre-liminary report. Amer. J. geriat. Soc., 10, 274-276.
Nunn, J. F. (1965). Influence of age and other factors on hypo-xaemia in the postoperative period. Lancet, 2, 466-468.
Nunn, J. F., and Freeman J. (1964). Problems of oxygenationand oxygen transport during haemorrhage. Anaesthesia,19, 206-216.
Olmstead, E. G. (1966). Mammalian Cell Water. Kimpton,London.
Post, R. L., and Jolly, P. C. (1957). The linkage of Na, K, andammonium active transport across the human erythrocytemembrane. Biochim. biophys. Acta (Anmst.), 25, 118-128.
Potts, W. T. W., and Parry, G. (1964). Osmotic and Ionic Regula-tion in Animals. Pergamon Press, Oxford.
Robinson, J. R. (1960). Body fluid dynamics. In Mineral Meta-bolism, vol. 1, pt A, edited by C. F. Comar and F. Bronner,pp. 205-248. Academic Press, New York and London.
Robinson, J. R. (1965). Water regulation in mammalian cells.In The State and Movement of Water in Living Organisms,S.E.B. Symposium No. xix, pp. 237-258. C.U.P.
Ross, E. J. (1965). Aldosterone and its antagonists. Pharmacol.Ther., 6, 65-106.
Schoffeniels, E. (1967). Cellular .4spects ofMembrane Permeability.Pergamon Press, Oxford.
Shires, T. (1969). Personal communication.Shires, T., and Carrico, C. J. (1966). Current Status of the Shock
Problem. Curr. Probl. Surg.Singh, C. M. (1965). Effect of major surgery on blood volume and
thiocyanate space. MS Thesis, Christian Medical College,Ludhiana Punjab, India.
Singh, C. M., and Flear, C. T. G. (1968). Why does the plasmasodium level fall after surgery? Brit. J. Surg., 55, 858.(Abstr.)
Singh, C. M., and Flear, C. T. G. (1969). Experimental haemor-rhagic and endotoxic shock in dogs; evidence for imbalancein exchanges of water and solutes across cell membranes.(Abstr.) Brit. J. Surg., 56, 703.
Singh, C. M., Flear, C. T. G., and Ross, D. (1969). Anoxic arrestduring heart surgery, its effects on water and electrolytecontents of the human heart. 2nd Annual Meeting. Inter-national Study Group for Research in Cardiac Metabolism,Milan/Gargnano, Sept. 1969. (To be published in full in aseparate volume of Rendiconti dell'Instituto Lombardo diScienze e Lettere.)
Smith, E. K. M., and Samuel, P. D. (1970). Abnormalities in thesodium pump of erythrocytes from patients with hyper-thyroidism. Clin. Sci., 38, 49-00.
Smith, E. K. M., and Welt, L. G. (1967). A reversible defect ofmembrane ATPase in the red cells of uremic patients.(Abstr.) Clin. Res., 15, 51.
Sodi-Pallares, D., Bisteni, A., Medrano, G. A., Micheli, A., dePonce Leon J. de, Calva, E., Fishleder, B. L., Testelli,M. R., and Miller, B. L. (1966). The polarizing treatmentin cardiovascular conditions. Experimental basis andclinical applications. In Electrolytes and CardiovascularDiseases, vol. 2, edited by E. Bajusz, p. 198. Karger, Basel.
Sodi-Pallares, D., Ponce de LUon. J., de Bisteni, A., and Medrano,G. A. (1969). Potassium, glucose, and insulin in myocardialinfarction. Lancet, 1, 1315-1316.
Stein, W. D. (1967). The Movement of Molecules Across CellMembranes. Academic Press, New York and London.
Stoner, H. B., and Threlfall, C. J. (1960). The effect of limbischaemia on carbohydrate distribution and energy trans-formation. In The Biochemical Response to Injury, editedby H. B. Stoner and C. J. Threlfall, pp. 105-128.Blackwell, Oxford.
Tosteson, D. C. (1964). Regulation of cell volume by sodium andpotassium transport. In The Cellular Functions ofMembraneTransport, pp. 3-22. Prentice Hall.
Tosteson, D. C., and Hoffman, J. F. (1960). Regulation of cellvolume by active cation transport in high and low potass-ium sheep red cells. J. gen. Physiol., 44, 169-194.
Tremoliere, S. J., and Derache, R. (1960). Metabolisme descomposes phosphores et specialement des nucleotides dansle tissu traumatise. In The Biochemical Response to Injury.edited by H. B. Stoner and C. J. Threlfall, p. 23. Blackwell,Oxford.
Wade, 0. L., and Bishop, J. M. (1962). Cardiac Output andRegional Blood Flowv. Blackwell Scientific Publications,Oxford.
Welt, L. G., Sachs, J. R., and McManus, T. J. (1964). An iontransport defect in erythrocytes from uremic patients.Trans. Ass. Amer. Phycns, 77, 169-181.
Welt, L. G., Smith, E. K. H., Dunn, M. J., Czerwinski, A.,Proctor, H., Cole, C., Balfe, J. W., and Gitelman, H. J.(1967). Membrane transport defect: the sick cell. Trans.Ass. Amer. Phycns, 80, 217-226.
Wollenberger, A., and Krause, E. G. (1968). Metabolic controlcharacteristics of the acutely ischemic myocardium.Amer. J. Cardiol, 22, 349-359.
Woodbury, J. W. (1963). Interrelationships between ion transportmechanisms and excitatory events. Fed. Proc., 22, 31-35.
Woodbury, J. W. (1965). The cell membrane: ionic and potentialgradients and active transport. In Physiology and Bio-physics, edited by T. C. Ruch and H. D. Patton, pp. 1-25.Saunders, Philadelphia and London.
Wynn, V., and Houghton, B. J. (1957). Observations in man uponthe osmotic behaviour of the body cells after trauma.Quart. J. Med. (N.S.), 26, 375-392.
Wynn, V., and Rob, C. G. (1954). Water intoxication: differentialdiagnosis of the hypotonic syndromes. Lancet, 1. 587-594.
on March 31, 2020 by guest. P
rotected by copyright.http://jcp.bm
j.com/
J Clin P
athol: first published as 10.1136/jcp.s3-4.1.16 on 1 January 1970. Dow
nloaded from