Embed Size (px)
Citation preview

Embryonic muscle development in direct and indirect developing marine
flatworms (Platyhelminthes, Polycladida)
D. Marcela Bolanosa and Marian K. Litvaitisb,�
a,bDepartments of Zoology and Natural Resources, University of New Hampshire, Durham, NH 03824, USA�Author for correspondence (email: [email protected])
SUMMARY We compared embryonic myogenesis of thedirect-developing acotylean polyclad Melloplana ferrugineawith that of Maritigrella crozieri, a cotylean that develops via alarval stage. Fluorescently labeled F-actin was visualized withlaser confocal microscopy. Developmental times are reportedas percentages of the time from oviposition to hatching: 7 daysfor M. crozieri and 22 days for M. ferruginea. The epitheliumbegan to form at 30% development in M. crozieri and at 15%development in M. ferruginea. Random myoblasts appearedin peripheral areas of the embryo at 36% and 22--30%development in M. crozeri and M. ferruginea, respectively.Circular and longitudinal muscle bands formed synchronouslyat 37--44% development in M. crozieri; yolk obscuredobservations of early myogenesis in M. ferruginea. An orthog-
onal muscle grid was established by 45--50% development inboth species. Diagonal muscles developed in M. ferruginea at60--71% development. Hence, juveniles of this species hatchwith the same basic body-wall musculature as adults. Larvaeof M. crozieri did not hatch with diagonal muscles; thesemuscles are acquired postmetamorphosis. Additionally, aspecialized musculature developed in the larval lobes of M.crozieri. Oral musculature was complex and established by72% development in both species. Our results are comparableto the muscle differentiation reported for other indirect-developing polyclads and for direct-developing species ofmacrostomid flatworms. Furthermore, they provide additionalsupport that the orthogonal muscle pattern of circular andlongitudinal muscles is a symplesiomorphy of Spiralia.
INTRODUCTION
Rhabditophora comprise a diverse group of flatworms
characterized by highly variable body shapes that are mostly
due to the body-wall musculature. Functionally, their
body-wall musculature is not only linked to body shape but
also to locomotion, feeding, copulation, and egg-laying
(Hyman 1951). Fluorescent dye-conjugated phalloidin
has been used to study the muscle organization of free-
living rhabditophorans such as Macrostomum hystricinum
marinum Rieger 1977 and Hoploplana inquilina Wheeler
1894 (Rieger et al. 1994; Reiter et al. 1996), facultative
parasites, such as Urastoma cyprinae (Graff 1882) (Hooge
and Tyler 1999), and obligate endoparasites such as Fasciola
hepatica Linne 1758, Diplostomum pseudospathaceum
Niewiadomska 1984 and Schistosoma mansoni Sambon 1907
(Czubaj and Niewiadomska 1997; Mair et al. 1998, 2000,
2003).
Overall, the rhabditophoran body-wall musculature is
composed of layers of outer circular and inner longitudinal
fibers organized into an orthogonal, grid-like pattern (Hyman
1951; Prudhoe 1985). This organization is considered the
ground pattern for many flattened, vermiform invertebrates
(all lineages within Platyhelminthes, Acoelomorpha, Nemer-
tea) (Hyman 1951; Rieger et al. 1991, 1994; Hooge 2001).
Additionally, the rhabditophoran body wall may contain one
or several sheets of diagonally oriented muscle fibers located
between and below the circular and longitudinal layers (Ho-
oge 2001). However, these diagonal layers may be reduced or
even be completely absent (for a comprehensive review of
adult flatworm body-wall musculature, see Hooge and Tyler
2004). The orientation of the diagonal fibers is such that they
angle clockwise and counterclockwise around the animal (Ri-
eger et al. 1991), crossing over each other and forming a
second grid, offset by about 551 from the orthogonal pattern.
Finally, a parenchymal musculature formed by muscle fibers
traversing the central parenchyma and consisting of well-de-
veloped dorsoventral, transverse, and ventral longitudinal
muscles is present (Hyman 1951).
Although descriptions of flatworm muscle organization
have received considerable attention, studies of pattern for-
mation during developmental myogenesis remain scarce (Re-
iter et al. 1996; Younossi-Hartenstein and Hartenstein 2000;
Hartenstein and Jones 2003), and are mostly focused on the
EVOLUTION & DEVELOPMENT 11:3, 290 –301 (2009)
DOI: 10.1111/j.1525-142X.2009.00331.x
& 2009 Wiley Periodicals, Inc.290

emerging model species, M. h. marinum (Rieger et al. 1991,
1994; Reiter et al. 1996; Morris et al. 2004). In a comparative
study on the differentiation of body-wall musculature inM. h.
marinum and H. inquilina, Reiter et al. (1996) found that
muscle development varied depending on the life cycle that is
if worms hatched as miniature adults or as larvae.H. inquilina
is an acotylean polyclad with indirect development via a
Muller’s larva; M. h. marinum belongs to the direct-develop-
ing Macrostomida. Both Polycladida and Macrostomida rep-
resent early lineages within the Rhabditophora (Carranza et
al. 1997; Litvaitis and Rohde 1999; Baguna and Riutort
2004). Furthermore, they both belong to the Archoophora, a
grouping based on an organizational grade derived from the
homocellular arrangement of female gonads and the produc-
tion of entolecithal eggs (Hyman 1951). This organizational
grade contrasts with the Neoophora, a group characterized by
heterocellular gonads and ectolecithal eggs and include all
remaining rhabditophorans (Hyman 1951). The terms, how-
ever, carry no systematic value and are best used for devel-
opmental descriptions only.
In the larvae of H. inquilina muscle development was
strongly dependent on two founder muscles bands that were
laid down in a bilaterally symmetrical pattern along the lon-
gitudinal axis of the animal. Following this initial muscle
guide, two circular rings of muscles developed: one anteriorly,
demarcating the rostral end from the trunk region, the other
more posteriorly at the junction between trunk and tail end
(Reiter et al. 1996). This is in contrast to observations in the
direct-developing M. h. marinum, where several longitudinal
muscle bands could be seen to which many circular fibers
attached at right angles (Reiter et al. 1996). In this direct-
developing pattern, no initial bilateral symmetry or trunk de-
marcations are evident. It is tempting to speculate that such
observed differences in muscle development are related to
differences in developmental mode (direct vs. indirect). How-
ever, at present, there is no evidence to support or refute such
speculations.
In this study, we examined embryonic muscle differentia-
tion in Maritigrella crozieri (Hyman 1939) and Melloplana
ferruginea (Schmarda 1859), providing the first comparative
analysis on pattern formation during polyclad myogenesis.
M. crozieri is characterized by a Muller’s larva and is grouped
into the suborder Cotylea (polyclads with a ventral sucker,
Lang 1884), whereas M. ferruginea is a direct-developing
acotylean (polyclads without a ventral sucker, Lang 1884).
The main purpose of this study was to provide new devel-
opmental and morphological data that contribute to the un-
derstanding of body-wall muscle formation during
embryogenesis, and to determine whether observed differ-
ences in muscle development are related to differences in de-
velopmental mode. Additionally, we also compared our
findings to muscle development in other flatworms and other
Spiralia.
MATERIALS AND METHODS
CollectionAdult worms of M. crozieri were collected from submerged hang-
ing lines at Little Jim’s Marina, Fort Pierce, FL, USA (27128.420N,
80118.40W), where they were associated with their prey, the ascid-
ian Ecteinascidia turbinata Herdermann 1880. Adult specimens of
M. ferruginea were collected from under rocks in the intertidal and
shallow subtidal zones at Peanut Island (261 46.4280N; 801
82.6080W) on the Atlantic coast of Florida, USA. Animals were
lifted off the substrate using a soft paintbrush and were placed
individually into small plastic bags filled with seawater. Reproduc-
tive maturity of individuals was determined by inspecting the ven-
tral sides of the worms for eggs. When present, eggs are easily
visible in the oviducts. Egg-bearing specimens were acclimated at
221C for 2 days in individual plastic bags containing Millipore-
filtered seawater.
CultureAfter acclimatization, mature specimens were placed into Petri
dishes and their uteri were punctured with small tungsten needles.
This caused the release of zygotes without egg capsules (naked
embryos). The embryos were transferred into small gelatin-coated
Petri dishes of Millipore-filtered seawater that contained 200mg/ml
streptomycin and 60mg/ml penicillin to prevent microbial or fungal
growth and maintained at 221C. Embryonic development was fol-
lowed until the formation of larvae and juveniles. Embryos were
observed under a Leica DMLB microscope (Leica, Bannockburn,
IL, USA) equipped with a Nikon CoolPix 8700 (Nikon, Melville,
NY, USA).
Phalloidin staining and confocal microscopyEmbryos, larvae, and juveniles were fixed at different developmen-
tal stages at room temperature in 4% paraformaldehyde in 0.01M
phosphate buffer (PBS; pH 7.4) for 45min. Specimens were rinsed
(3 � ) for 15min with 0.01M PBS, permeabilized for 1h in 0.2%
Triton X-100 in PBS, stained for 2h or overnight with Alexa Fluor
488 phalloidin (Molecular Probes, Eugene, OR, USA), and rinsed
twice for 10min with PBS. Specimens were mounted on glass slides
in Gel/Mount (Biomeda Corp., Electron Microscopy Sciences,
Hatfield, PA, USA), and digital image acquisition and analysis
were performed using a Zeiss LSM 510 confocal microscope (Zeiss,
Thornwood, NY, USA). Samples were excited with a 488nm
multi-line argon-ion laser and emitted fluorescence was collected
using a 505–530nm bandpass filter. Measurements of the speci-
mens were taken with an ocular micrometer before fixation. For
each species, at least four specimens were examined for each de-
velopmental stage.
RESULTS
To conform to published studies on embryonic myogenesis in
archoophorans (Reiter et al. 1996; Ladurner and Rieger
2000), we define developmental stages as a percentage of total
Embryonic muscle development in marine £atworms 291Bolan� os and Litvaitis

developmental time, where 100% represents the time from
oviposition to hatching. However, we also include time since
oviposition in hours to allow for comparisons with studies of
other spiralians (Maslakova et al. 2004; Bergter and Paululat
2007). Total developmental time forM. crozieri was 6–7 days
and 18–22 days for M. ferruginea. These time frames are
comparable to other studies that also recorded much longer
developmental times for acotylean than cotylean polyclads
(Rawlinson et al. 2008; D. Bolanos & M. Litvaitis,
unpublished data).
Muscle differentiation in Maritigrella crozieri
Cleavage begins before oviposition. After oviposition (0%
development), zygotes (average diameter5134mm; n550)
underwent holoblastic, spiral cleavage, eventually dividing the
embryos into 64 cells. At about 17% of development (28h),
the embryos contained small amounts of yolk, which emitted
a dull autofluorescence. Gastrulation began at approximately
30% of development (50h). The surfaces of the embryos be-
came irregularly shaped, and a faintly staining, polygonal
pattern outlining the epithelial cells became visible (Fig. 1A).
Actin filaments inserting into the zonulae adhaerentes were
evident (Fig. 1B). Additional actin was distributed across the
embryo surface but was present in higher concentrations in
some regions (Fig. 1A). No muscle fibers were observed im-
mediately after epiboly. However, shortly after gastrulation
(approximately 36% development, 60h), a few small myo-
blasts were randomly dispersed throughout the embryo.
Once gastrulation was complete, the first cilia appeared
and minimal movement of the embryos was observed (37–
43% of development, 62–72h). A brightly staining, primary
circular muscle band was recognized (Fig. 1C), and unorga-
nized muscle fibers were distinguishable in the center of the
developing embryo. During this stage, many individual mus-
cle fibers appeared. At about 43–44% of development (72–
74h, day 4), the embryo still appeared spherical but muscle
fibers began to form concentrically around the apical organ
and in the oral region (Fig. 1D). Ciliary action caused the
embryos to slowly rotate within the egg capsules. Secondary
circular fibers derived by branching from the primary circular
band were detectable, although still no contractions were ob-
served at this point (Fig. 1E). Additionally, unorganized
weakly staining fibers could be distinguished deep within the
embryo. Also during this stage, a primary longitudinal muscle
formed and extended whip-like along the entire length of
the embryo, from anterior to posterior back again (Fig. 1E).
Presumably, this looped fiber extended into the area of a
future larval lobe.
At 45–52% of development (75–88h, day 4), a simple spi-
ral of circular muscle extending around the apical organ was
evident (Fig. 2A). During this time, additional longitudinal
muscles began to develop; they could be distinguished as
faintly staining branches (Fig. 2B). Thicker, circular fibers
encircled the periphery of the embryo, bifurcating and form-
Fig. 1. Confocal projections of the earlyembryonic development of Maritigrellacrozieri stained with phalloidin. (A) Em-bryo surface at about 30% of develop-ment, showing polygonal outlines ofepidermal cells. Scale bar550mm. (B)Higher magnification of embryo surface,arrowheads indicate actin in the zonulaeadhaerentes. Scale bar510mm. (C) Api-cal view of postgastrulation embryo,showing primary circular muscle band(pcm). Scale bar550mm. (D) Dorsolat-eral view of embryo at about 43%of development, showing concentric or-ganization of musculature surroundingapical organ (ao) and developing circularmusculature of mouth (m). (E) Lateralview of embryo showing primary longi-tudinal muscle (plm) extending whip-likealong the entire length, pcm delineatingextent of ao musculature, and orthogonalnetwork of circular (cm) and longitudinalmuscles (lm); asterisk indicates apexof embryo; ep, epidermis. Scale bars5
25mm.
292 EVOLUTION & DEVELOPMENT Vol. 11, No. 3, May--June 2009
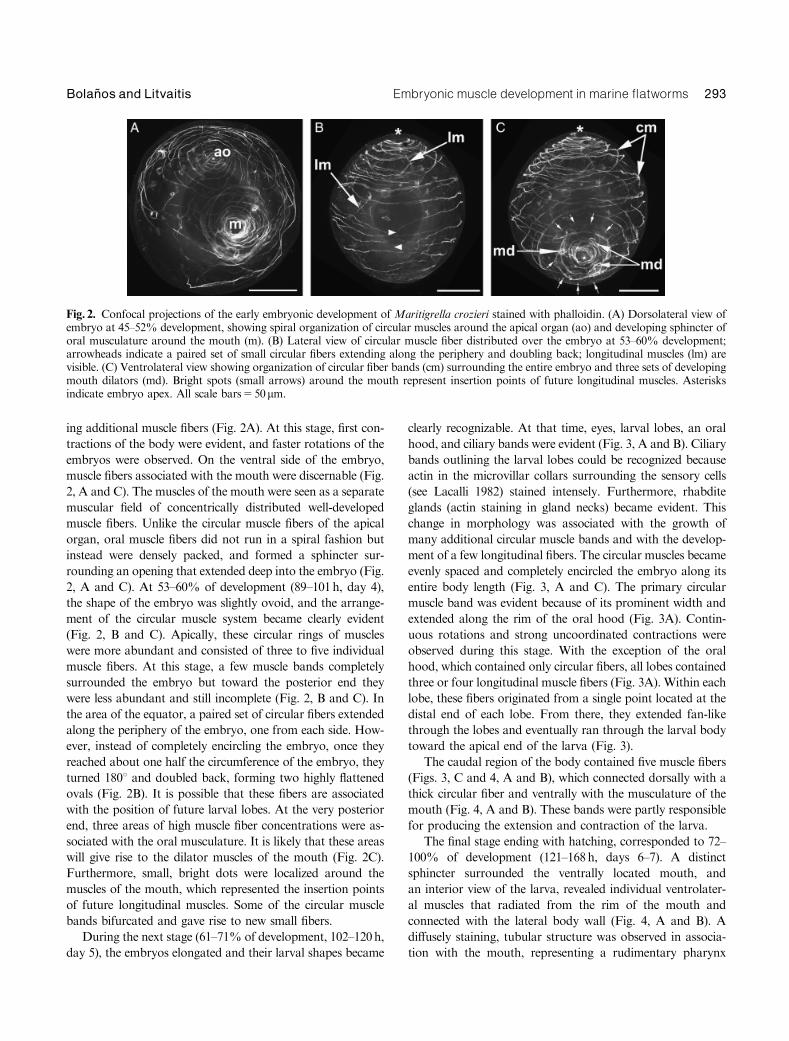
ing additional muscle fibers (Fig. 2A). At this stage, first con-
tractions of the body were evident, and faster rotations of the
embryos were observed. On the ventral side of the embryo,
muscle fibers associated with the mouth were discernable (Fig.
2, A and C). The muscles of the mouth were seen as a separate
muscular field of concentrically distributed well-developed
muscle fibers. Unlike the circular muscle fibers of the apical
organ, oral muscle fibers did not run in a spiral fashion but
instead were densely packed, and formed a sphincter sur-
rounding an opening that extended deep into the embryo (Fig.
2, A and C). At 53–60% of development (89–101h, day 4),
the shape of the embryo was slightly ovoid, and the arrange-
ment of the circular muscle system became clearly evident
(Fig. 2, B and C). Apically, these circular rings of muscles
were more abundant and consisted of three to five individual
muscle fibers. At this stage, a few muscle bands completely
surrounded the embryo but toward the posterior end they
were less abundant and still incomplete (Fig. 2, B and C). In
the area of the equator, a paired set of circular fibers extended
along the periphery of the embryo, one from each side. How-
ever, instead of completely encircling the embryo, once they
reached about one half the circumference of the embryo, they
turned 1801 and doubled back, forming two highly flattened
ovals (Fig. 2B). It is possible that these fibers are associated
with the position of future larval lobes. At the very posterior
end, three areas of high muscle fiber concentrations were as-
sociated with the oral musculature. It is likely that these areas
will give rise to the dilator muscles of the mouth (Fig. 2C).
Furthermore, small, bright dots were localized around the
muscles of the mouth, which represented the insertion points
of future longitudinal muscles. Some of the circular muscle
bands bifurcated and gave rise to new small fibers.
During the next stage (61–71% of development, 102–120h,
day 5), the embryos elongated and their larval shapes became
clearly recognizable. At that time, eyes, larval lobes, an oral
hood, and ciliary bands were evident (Fig. 3, A and B). Ciliary
bands outlining the larval lobes could be recognized because
actin in the microvillar collars surrounding the sensory cells
(see Lacalli 1982) stained intensely. Furthermore, rhabdite
glands (actin staining in gland necks) became evident. This
change in morphology was associated with the growth of
many additional circular muscle bands and with the develop-
ment of a few longitudinal fibers. The circular muscles became
evenly spaced and completely encircled the embryo along its
entire body length (Fig. 3, A and C). The primary circular
muscle band was evident because of its prominent width and
extended along the rim of the oral hood (Fig. 3A). Contin-
uous rotations and strong uncoordinated contractions were
observed during this stage. With the exception of the oral
hood, which contained only circular fibers, all lobes contained
three or four longitudinal muscle fibers (Fig. 3A). Within each
lobe, these fibers originated from a single point located at the
distal end of each lobe. From there, they extended fan-like
through the lobes and eventually ran through the larval body
toward the apical end of the larva (Fig. 3).
The caudal region of the body contained five muscle fibers
(Figs. 3, C and 4, A and B), which connected dorsally with a
thick circular fiber and ventrally with the musculature of the
mouth (Fig. 4, A and B). These bands were partly responsible
for producing the extension and contraction of the larva.
The final stage ending with hatching, corresponded to 72–
100% of development (121–168h, days 6–7). A distinct
sphincter surrounded the ventrally located mouth, and
an interior view of the larva, revealed individual ventrolater-
al muscles that radiated from the rim of the mouth and
connected with the lateral body wall (Fig. 4, A and B). A
diffusely staining, tubular structure was observed in associa-
tion with the mouth, representing a rudimentary pharynx
Fig. 2. Confocal projections of the early embryonic development of Maritigrella crozieri stained with phalloidin. (A) Dorsolateral view ofembryo at 45–52% development, showing spiral organization of circular muscles around the apical organ (ao) and developing sphincter oforal musculature around the mouth (m). (B) Lateral view of circular muscle fiber distributed over the embryo at 53–60% development;arrowheads indicate a paired set of small circular fibers extending along the periphery and doubling back; longitudinal muscles (lm) arevisible. (C) Ventrolateral view showing organization of circular fiber bands (cm) surrounding the entire embryo and three sets of developingmouth dilators (md). Bright spots (small arrows) around the mouth represent insertion points of future longitudinal muscles. Asterisksindicate embryo apex. All scale bars550mm.
Embryonic muscle development in marine £atworms 293Bolan� os and Litvaitis

(Fig. 4, A and B). However, no specific pharyngeal muscu-
lature could be identified. Additionally, short, longitudinal
fibers, some crossing over each other, forming a distinct X
shape were located in the oral hood (Fig. 4, A and B).
At hatching, no diagonal muscle fibers were found and
the entire arrangement of the muscles consisted of sets of
circular and longitudinal fibers (Fig. 4, C and D). Two groups
of concentric muscles that overlapped each other and formed
a diagonally arranged, lattice-like pattern were observed (Fig.
4, C and D).
Muscle differentiation in Melloplana ferruginea
Embryogenesis in direct-developing polyclads generally takes
longer than that of indirect-developing species. Furthermore,
the embryos usually contain much larger amounts of yolk, a
phenomenon that can slow development in the early stages
(this study; Rawlinson et al. 2008). The embryos of M. fer-
ruginea were somewhat smaller than those of M. crozieri (av-
erage diameter5125mm; n550) and contained large
amounts of yolk. This yolk emitted a strong autofluores-
cence. The period from oviposition to the beginning of gas-
trulation corresponds to approximately 0–15% development
(0–79h). During early gastrulation (at about 14% develop-
ment, 74h), the fluorescence started to abate and at about
15% of development (79h), the polygonal outlines of epider-
mal cells were visible (Fig. 5A) and continued to be distinct
until about 16–21% of development (84–111h, day 4). The
first cilia appeared and some movement was noted at this
stage.
At 22–30% development (116–152h, days 5–6), ciliation
increased and slow rotational movements could be observed.
Additionally, the bright muscle anlagen visible on the surface
of the embryo increased in number and size (Fig. 5B). During
Fig. 3. Larval stages of Maritigrella cro-zieri at 61–71% of development. Confo-cal projection (A) and schematic diagram(B) of ventral view of Muller’s larvashowing oral hood (oh), primary circularmuscle (pcm), one lateral lobe (ll), andtwo ventrolateral lobes (vl), each contain-ing prominent longitudinal muscle fibers(arrowheads) converging at the distal endof the lobes. Small bright spots outliningthe lobes represent the ciliary band. Threeeyes (e) are visible at the apical end. Con-focal projection (C) and schematic dia-gram (D) of dorsal view of the larvashowing lateral (ll) and dorsolateral (dl)lobes, apical organ (ao) and caudal endcontaining distinct muscle fibers (arrows)that extend from the dorsal to the ventralsurface of the larva. Scale bars550mm.
294 EVOLUTION & DEVELOPMENT Vol. 11, No. 3, May--June 2009

the period of 31–44% of development (163–232h, days 7–9),
the embryo was still spherical in shape and a single eye was
present. The yolk cells migrated to the interior of the embryo
diminishing the ability to visualize individual muscle fibers.
Most myocytes appeared in an unorganized network. How-
ever, along the periphery of the embryo it was possible to
differentiate some circular fibers (Fig. 5B).
A second small eye appeared at 45–59% of development
(238–312h, days 10–13). In addition, faster rotations and first
contractions of the juveniles were observed. The body elon-
gated into a more ovoid shape, and the formation of a fine
orthogonal grid of circular and longitudinal muscles was ev-
ident (Fig. 5C). Unequally spaced circular fibers surrounded
the entire embryo, whereas a few longitudinal fibers extended
the length of the body, some of which bent diagonally at the
anterior end of the worm. Circular fibers were better devel-
oped than the longitudinal muscle fibers but a more dense
formation of longitudinal muscles was observed (Fig. 5C).
Well-developed, branched, dorsoventral muscles were visible
extending across the body of the embryo (Fig. 5D). These
muscles play an important role in locomotion and the main-
tenance of a flat body shape. Furthermore, the brain was
prominently visible in the anterior third of the embryo due to
actin staining in the neuropil (Figs. 5D and 6E). Also during
this period, the rudiments of the mouth on the ventral surface
was visible as an elongated shape consisting of aggregated and
unstructured muscle fibers (Fig. 5D).
At 60–71% of development (317–374h, days 14–16), the
growth of diagonal muscle bands on the dorsal (Fig. 6, A and
C) and ventral (Fig. 6, D and F) surfaces proceeded rapidly.
Another pair of eyes developed, and the mouth took on its
circular shape (Fig. 6, B and D–F). Dorsal, longitudinal
Fig. 4. Larval stages of Maritigrella cro-zieri at 72–100% of development. Con-focal projection (A) and schematicdiagram (B) of deep ventral view withventrolateral muscles (arrows) radiatingfrom the oral sphincter (os). Caudal mus-cle fibers (arrowheads) connect ventrallywith the mouth. Short, oral hood longi-tudinal muscles (ohl), some crossing toform an X are visible in the apical portionof the larva. Confocal projection (C) andschematic diagram (D) of lateral view ofa late-stage larva, showing two sets ofconcentric muscles arranged in a lattice-like pattern in the lateral lobes. clm, con-centric lobe musculature; ao, apical or-gan; e, eyes; dl, dorsolateral lobe; ll,lateral lobe; md, mouth dilators; oh, oralhood; ph, rudimentary tubular pharynx;rg, rhabdite glands. Scale bar550mm.
Embryonic muscle development in marine £atworms 295Bolan� os and Litvaitis

muscle fibers extended straight from the anterior to the pos-
terior end (Fig. 6, A, C and D). Dorsally, a paired set of
diagonal muscles covered the entire length of the worm,
crossing over each other at a 901 angle (Fig. 6, A–C). These
fibers were denser and closer at the anterior part of the body
becoming more spaced and still incomplete at the posterior
end (Fig. 6A). Ventrally, diagonal muscle bands covered the
anterior half of the body, crossing over each other at the body
midline and then extending in a more longitudinal orientation
along the lateral sides of the body (Fig. 6, D and F). All
diagonal muscle fibers were located between the longitudinal
and the circular muscles. Juveniles within the egg capsules
were observed to vigorously bend their anterior ends, a mo-
tion mediated by diagonal muscles. Ventral, longitudinal fi-
bers located along the body edges extended the entire length
of the juveniles, but at the posterior end behind the mouth
opening, they curved and formed U-shaped muscles (Fig. 6, D
and F).
At this stage, parenchymal muscles were present through-
out the body as disorganized, conspicuous fibers running in
varying directions (Fig. 6E). The concentric sphincter muscles
of the mouth became established as a smaller, inner and a
larger, outer ring of muscles (Fig. 7, A–C). At this stage, short
radial fibers seemed to connect the inner to the outer sphinc-
ter, however, they are most likely longitudinal muscles of the
future pharynx (Fig. 7, A–C). The ventral mouth was located
at the very posterior end. Elongation of the growing embryo
will cause the mouth to gradually move anterior to its final
midventral position characteristic of adult M. ferruginea.
Rhabdite glands could be distinguished and were more abun-
dant on the dorsal surface (Figs. 6, A and B and 7A).
The final stage corresponds to 72–100% of development
(380–528h, days 18–22). At approximately 78% of develop-
ment (411h, day 17), the juveniles had almost completed their
development but remained in their egg capsules for a few
additional days, continuously rotating and contracting until
hatching. Their bodies elongated and tapered posteriorly, ac-
quiring their typical worm shape. During that period, only a
few changes in the musculature were observed. The ventral
parenchymal muscles and the oral musculature became more
defined (Fig. 7). No changes were observed in the body-wall
musculature.
The oral musculature is complex and formed by distinct
sets of muscles (Fig. 7). A set of concentrically arranged
Fig. 5. Confocal projections of muscledifferentiation in Melloplana ferruginea.(A) Embryo at 15% of development withpolygonal outlines of epidermal cells. (B)Postgastrulation embryo at 22–30% de-velopment with bright spots of actin rep-resenting the anlagen of the body-wallmuscles. Arrows indicate short longitudi-nal muscle fibers in the periphery of theembryo. (C) Elongated embryo at 45–59% of development, showing a fine or-thogonal network of circular (cm) andlongitudinal muscles (lm). (D) Deep lat-eral view of embryo with anterior brainanlagen (b) and with branched, dorso-ventral muscles (arrowheads) traversingthe body. Aggregation of unstructuredmuscle fibers on the ventral side formingmusculature associated with the mouth(om). All scale bars525mm. rg, rhabditeglands.
296 EVOLUTION & DEVELOPMENT Vol. 11, No. 3, May--June 2009

muscles forms a larger sphincter at the ventral surface of the
worm and surrounds the mouth opening. The mouth
opening invaginates into the body wall, leading into a fun-
nel-like structure with a second, smaller sphincter located
where the funnel narrows to a tube. The second sphincter is
also composed of circularly arranged muscles. The funnel
and tube are lined by longitudinal muscles, which appear
to be arranged as radial muscles between the two sphincters
because they are splayed outwards (Fig. 7, A–C). Finally,
distinct dilator muscles extending from both sphincters
to the lateral body wall anchor the mouth (Fig. 7, A–C).
Random ventral parenchymal muscles that tended to extend
in a more longitudinal direction (Fig. 7B), eventually will at-
tach to the ventral and dorsal body wall and to organs.
Other parenchymal muscles remain suspended as individual
fibers throughout the parenchyma. During this stage, an in-
crease in the number of rhabdite glands was also observed
(Fig. 7A).
Fig. 6. Confocal projections of muscle differentiation in Melloplana ferruginea. (A) Projection showing network of longitudinal, circular,and diagonal muscles. (B) Deeper view showing oral musculature. (C) Schematic diagram of dorsal musculature. (D) Projection showingnetwork of longitudinal, circular, and diagonal muscles, and early oral musculature. (E) Deep view showing oral muscle sphincter andirregularly arranged parenchymal muscles. (F) Schematic diagram of ventral musculature. Arrowheads indicate longitudinal musclesbending in a U-shape at the posterior end. b, brain; cm, circular muscles; dm, diagonal muscles; e, eyes; lm, longitudinal muscles; om, oralmusculature; pm, parenchymal muscles; rg, rhabdite glands. All scale bars525mm.
Embryonic muscle development in marine £atworms 297Bolan� os and Litvaitis

DISCUSSION
Comparison of embryonic myogenesis inMaritigrella crozieri with other indirect developingpolyclads
In an ultrastructural and immunohistochemical study, You-
nossi-Hartenstein and Hartenstein (2000) describe muscle pre-
cursors in embryos of the polyclad Imogine mcgrathi Jennings
and Newman 1996 at about 84h (their stage 6), which develop
into circular and longitudinal fibers by the end of day 4 (their
stage 7). Considering that I. mcgrathi larvae hatched in about
7–8 days (Younossi-Hartenstein and Hartenstein 2000), these
events correspond to about 43–57% of development, a time
line that is comparable to our results for M. crozieri. Addi-
tionally, Younossi-Hartenstein and Hartenstein (2000) men-
tion the development of two distinct longitudinal muscle bands
in I. mcgrathi albeit without providing further description on
their position and fate. It is possible that these bands corre-
spond to the primary longitudinal muscle seen extending into
the area of the future larval lobes in M. crozieri.
A comparison of embryonic myogenesis of M. crozieri
with that of H. inquilina reveals similarities with respect to
primary muscle bands and the arrangement of the circular
musculature around the apical organ. According to Reiter et
al. (1996), a primary muscle grid appears at 80% of devel-
opment inH. inquilina. This orthogonal grid consists of a pair
of primary longitudinal fibers located laterally, an anteriorly
located primary circular muscle band, and a posterior pri-
mary circular muscle fiber (Reiter et al. 1996). In M. crozieri,
we also recognized the early formation of an anterior primary
circular muscle (43–44% of development); however, none of
the posterior circular muscle bands could be identified as a
primary muscle. We could, however, identify a major longi-
tudinal muscle that extended the length of the embryo and
then looped back to the anterior end. It is possible that this
muscle fiber corresponds to the primary longitudinal muscle
fiber described in H. inquilina (Reiter et al. 1996).
Also at 80% of development, the circularly arranged mus-
culature around the apical organ of H. inquilina is evident
(Reiter et al. 1996). Like in M. crozieri, it forms a simple
spiral. In both species, once the oral hood is established, this
apical muscular spiral terminates at the conspicuous primary
circular muscle band. In both species, the primary circular
muscle eventually extends along the rim of the oral hood.
A clear difference with respect to the timing of the ap-
pearance of major muscle fibers was found between the two
species. In H. inquilina, the first recognizable primary fibers
appeared only at 80% of development while the embryo was
still ovoid and had not yet acquired a typical larval form
(Reiter et al. 1996). In comparison, embryos of M. crozieri at
80% of development were clearly distinguishable as Muller’s
larva with well-developed larval lobes containing a complex
network of muscle fibers and a recognizable body-wall mus-
culature of circular and longitudinal muscles. The amount of
yolk and rearing temperatures are two factors that have been
shown to affect developmental time lines in polyclads (Raw-
linson et al. 2008; D. Bolanos & M. Litvaitis, unpublished
data). A case could be made that as an acotylean polyclad, the
embryos of H. inquilina contain large amounts of yolk, which
potentially could delay the growth of the myoblasts. How-
ever, I. mcgrathi also is an acotylean and its time line follows
the one observed in M. crozieri (Younossi-Hartenstein and
Hartenstein 2000). Although Reiter et al. (1996) do not pro-
vide culture temperature for their specimens, total develop-
mental time for H. inquilina was o4.5 days (compared with
6–7 days for M. crozieri at 221C), a fact that may invalidate a
claim of delayed development due to low temperature. Most
likely, these temporal differences are due to the fact that I.
mcgrathi and M. crozieri are subtropical species, whereas H.
inquilina is found in temperate regions.
Another difference between the two species involved the
presence of a few obliquely oriented muscle fibers in H. in-
quilina as early as 80% development (Reiter et al. 1996). We
did not observe such muscle fibers in M. crozieri, even at the
Fig. 7. Confocal projections of oralmuscle differentiation in Melloplana fe-rruginea. (A) Embryo with oral muscula-ture composed of concentric and splayedlongitudinal fibers and mouth dilators.(B) Late-stage embryo with complex oralmusculature forming a funnel-like struc-ture and with mouth dilators. (C) Sche-matic diagram of oral musculature. md,mouth dilators; om, oral musculature;pm, parenchymal muscles; rg, rhabditeglands. All scale bars525mm.
298 EVOLUTION & DEVELOPMENT Vol. 11, No. 3, May--June 2009

hatching stage. Hence, the diagonal muscles found in adult
M. crozieri are acquired at or after metamorphosis. This is
consistent with the function of such muscles. Contraction of
diagonal muscles causes lateral bending of the worms, a mo-
tion that is observed even in juveniles that are still in the egg
capsules (see section on M. ferruginea). Larvae of M. crozieri
do not require such motions and therefore do not have di-
agonal muscles.
The mouth of all polyclads is located ventrally as an inv-
agination of the body wall. The mouth opening ofM. crozieri
was surrounded by a well-developed circular muscle sphinc-
ter, consisting of at least three concentric fibers. From this
sphincter, paired ventrolateral longitudinal muscles extended
apically, and eventually connected with the lateral body wall.
Some of the ventrolateral muscles appear to connect with the
primary circular muscle fiber in the oral hood. In contrast, the
oral musculature of H. inquilina consists of an oral sphincter
surrounding the mouth from which an anteriorly located
mouth retractor muscle extends and also connects to the cir-
cular musculature of the apical organ (Reiter et al. 1996). It is
possible that this mouth retractor corresponds to one of the
ventrolateral muscles observed in M. crozieri.
Additionally, in M. crozieri an inner small muscular ring
was associated with a simple, straight pharynx outlined by
diffusely staining material. Pharyngeal muscles were not yet
developed at the larval stage. According to Ruppert (1978),
the development of a muscular pharynx occurs during larval
metamorphosis. Hence, the lack of a pharyngeal musculature
in M. crozieri reflects the planktonic life style of a larval sus-
pension feeder. The same has been observed in the larvae of I.
mcgrathi (Younossi-Hartenstein and Hartenstein 2000). This
is in contrast to the findings of Reiter et al. (1996), which
describe a complex pharyngeal musculature of radial, longi-
tudinal, and circular fibers in H. inquilina.
Finally, Reiter et al. (1996) mention changes in the orien-
tation of muscle fibers especially around the lobes and the
mouth as the larvae change in shape to a typical Muller’s
larva. However, they do not elaborate on these changes, and
no mention is made with regards to a unique double-set of
overlapping muscles as we found in the lobes of M. crozieri.
Comparison of embryonic myogenesis inMelloplana ferruginea with other direct developingRhabditophora
To date, no information is available on developmental myo-
genesis of direct-developing polyclads. Hence, we compare the
events in M. ferruginea to those of other rhabdiophorans,
especially to M. h. marinum. Time of development from ovi-
position to hatching of juvenile M. h. marinum is 4–5 days
(Reiter et al. 1996).
In addition to circular and longitudinal muscles,
rhabditophoran flatworms are characterized by a layer of di-
agonal muscles (Hyman 1951; Prudhoe 1985; Hooge 2001). In
M. ferruginea diagonal muscles appeared at about 60% de-
velopment and were well developed by 70% developmental
time. JuvenileM. ferruginea still within their egg capsules were
observed to laterally bend their anterior ends, a movement
possible because of diagonal muscles. An ultrastructural study
on muscle differentiation ofM. h. marinum times the presence
of circular, diagonal, and longitudinal muscle fibers between
50% and 64% of development (Rieger et al. 1991). Using
rhodamine–phalloidin staining, Reiter et al. (1996) show that
in M. h. marinum muscle fiber differentiation does not start
before 55% of development and that earlier stages of devel-
opment are obscured. Although we were able to identify
brightly staining actin muscle anlagen of M. ferruginea
around 21% of development, later differentiation events also
were obscured in our specimens. Comparable to M. h. mari-
num, a network of circular, longitudinal, and diagonal muscle
fibers was established between 45% and 70% of development.
We did not, however, observe ‘‘muscle lattices’’ as had been
described for M. h. marinum (Reiter et al. 1996).
Temporal congruence between the two species was also
found with respect to the development of dorsoventral muscle
fibers and of the oral musculature. In juveniles of M. fer-
ruginea the dorsoventral muscles are well developed by 60–
71% of development and function to maintain the flattened
shape of the worms. In comparison to M. crozieri, the mouth
of M. ferruginea is associated with a more developed but still
rudimentary pharynx. Two rings of circular muscles act as
sphincters and are connected to dilator muscles, which in turn
connect to the body wall. The mouth leads into a tubular
structure lined by longitudinal muscles. Because of a funnel-
shaped eversion of the mouth between the two circular
sphincters, these longitudinal muscles appear to be radially
arranged. However, this is the result of splaying out the oral
opening. In M. h. marinum on the other hand, Rieger et al.
(1994) describe distinct radial fibers associated with the phar-
ynx. In polyclads, pharyngeal musculature is important in
structuring the shape of the adult pharynx, (i.e., ruffled vs.
tubular pharynx), a differentiation that is of taxonomic im-
portance for familial distinctions (Faubel 1983, 1984; Prudhoe
1985). No gut musculature can be identified in embryos of
either M. ferruginea or M. h. marinum, a notion also con-
firmed for Macrostomum sp. (now described as M. lignano,
Ladurner et al. 2005) (Morris et al. 2004). Furthermore,
Morris et al. (2004) describe a muscle net in the anterior third
of Macrostomum sp. in which fibers follow the outer surface
of the brain as well as pass through the brain cortex and
neuropil. A similar arrangement of muscle fibers surrounding
and passing through the brain can be seen in M. ferruginea.
Muscle development of M. ferruginea can be compared
with that of free-living rhabditophorans other than macros-
tomids. However, these studies mention myogenesis only
within the context of development and organogenesis of the
Embryonic muscle development in marine £atworms 299Bolan� os and Litvaitis

entire embryo and do not involve specific muscle staining
approaches (Hartenstein and Ehlers 2000; Cardona et al.
2005). Hartenstein and Ehlers (2000) found that in the
rhabdocoel Mesostoma lingua (Abildgaard 1789) embryonic
myogenesis also results in a typical grid-like pattern of circular
and longitudinal muscles on day 7 (their stage 7; total devel-
opmental time 8 days with embryos maintained at 23–251C).
In the triclad, Schmidtea polychroa (Schmidt 1862) on the
other hand, layers other than the orthogon (second longitu-
dinal, diagonal) form during development and become part of
the body-wall musculature (Cardona et al. 2005). Because
polyclads and macrostomids are considered basal within
Rhabditophora (Carranza et al. 1997; Litvaitis and Rohde
1999; Baguna and Riutort 2004), the orthogonal muscle grid
of longitudinal and circular muscles appears to represent the
primitive arrangement within Platyhelminthes, with more
derived clades evolving additional muscle sets and/or
arrangements.
Comparisons with other Lophotrochozoa
Although Polycladida clearly belongs to the Spiralia based on
its cleavage pattern and dual origin of mesoderm (Boyer et al.
1996, 1998), a comparison of muscle development of larval
M. crozieri with that of other larvae in the clade is dependent
on the homology between polyclad larvae and trochophores.
Whereas homology among trochophores of mollusks, anne-
lids, and sipunculids has been established (Nielsen 2001; Ma-
slakova et al. 2004; Wanninger et al. 2005), homology of
polyclad larvae with lophotrochozoan larvae has not yet been
demonstrated. With this caveat in mind, we here compare the
larval musculature of M. crozieri to that of other spiralian
larvae.
Embryonic myogenesis of Capitella sp. 1 occurs sequen-
tially from anterior to posterior and results in the formation
of longitudinal, circular, and oblique body-wall muscles in
addition to specialized intrasegmental and setal sac muscle
fibers (Hill and Boyer 2001). According to Hill and Boyer
(2001), all larval muscles are retained postmetamorphosis and
develop into the adult musculature. In the polychaete Pom-
atoceros lamarckii (Quatrefages 1866) musculature associated
with the esophagus and intestine develop early and body-wall
musculature (which lacks circular muscles) is formed only
once the trochophore elongates. Again, the larval body-wall
musculature is retained in adult P. lamarckii (McDougall et
al. 2006). Unlike the weakly developed (or even absent) cir-
cular muscles bands in polychaetes where muscle formation
may be controlled by segmentation (Bergter et al. 2008), most
circular muscles in the larvae of M. crozieri appear synchro-
nously; diagonal body-wall muscles are acquired after meta-
morphosis. This is comparable to the process observed in the
sipunculid Phascolion strombus (Montagu 1804) in which cir-
cular muscles also appear synchronously, not sequentially
from anterior to posterior. Development of longitudinal mus-
cles, arranged in bands, follows during late larval develop-
ment (Wanninger et al. 2005).
Wanninger and Haszprunar (2002) describe the establish-
ment of an apical muscle grid during myogenesis in the larvae
of two species of chiton. Although the authors question un-
equivocal homology of the longitudinal fibers of the apical
grid with longitudinal fibers of the body-wall muscle grid,
they identify the apical grid as possible vestiges of the body-
wall musculature shared by the common ancestor of mollus-
can and vermiform spiralians.
From the above comparisons, it appears that in unseg-
mented spiralians the rudiments of circular muscles develop
synchronously. Our results in M. crozieri follow this pattern
supporting a close evolutionary relationship between unseg-
mented lophotrochozoans and polyclad flatworms based on
myogenesis. Furthermore, myogenetic events of body-wall
musculature in M. ferruginea also resemble the embryonic
muscle development observed in Acoela (Ladurner and Ri-
eger 2000; Semmler et al. 2008), a taxon occupying a basal
position in Bilateria.
With the present study, we have added information not
only from another indirect developing polyclad but also from
a direct developing species. Our results provide additional
support that the orthogonal muscle pattern of circular and
longitudinal muscle bands is a symplesiomorphy of Spiralia
and may already have been present in the stem species of
bilateral animals (Ladurner and Rieger 2000; Rieger and
Ladurner 2001, 2003; Semmler et al. 2008).
AcknowledgmentsThis work was supported by NSF grant DEB-0412932. A portion ofthis research was conducted at the Smithsonian Marine Station atFort Pierce, FL as part of a graduate student Link Fellowship. Weare grateful to Drs. Sigmer Quiroga and Kate Rawlinson for theirhelp in the field and for collecting embryos at different developmentalstages. Special thanks go to Dr. Mark Townley (InstrumentationCenter, UNH) for his technical assistance with the confocal micro-scope. We acknowledge NSF grant DBI-0618719 through which theconfocal microscope at UNH was acquired. This is Scientific Con-tribution No. 2378 from the New Hampshire Agricultural Experi-ment Station and Contribution No. 771 from the SmithsonianMarine Station at Fort Pierce, FL.
REFERENCES
Baguna, J., and Riutort, M. 2004. Molecular phylogeny of the Platy-helminthes. Can. J. Zool. 82: 168–193.
Bergter, A., Brubacher, J. L., and Paululat, A. 2008. Muscle formationduring embryogenesis of the polychaete Ophryotrocha diadema(Dorvilleidae)Fnew insights into annelid muscle patterns. Front. Zool.5: 1–18.
Bergter, A., and Paululat, A. 2007. Pattern of body-wall muscle differen-tiation during embryonic development of Enchytraeus coronatus (Annel-ida: Oligochaeta; Enchytraeidae). J. Morphol. 268: 537–549.
300 EVOLUTION & DEVELOPMENT Vol. 11, No. 3, May--June 2009

Boyer, B. C., Henry, J. J., and Martindale, M. Q. 1998. The cell lineage of apolyclad turbellarian embryo reveals close similarity to coelomate spi-ralians. Dev. Biol. 204: 111–123.
Boyer, B. C., Henry, J. Q., and Martindale, M. Q. 1996. Dual origins ofmesoderm in a basal spiralian: cell lineage analyses in the polyclad tur-bellarian Hoploplana inquilina. Dev. Biol. 179: 329–338.
Cardona, A., Hartenstein, V., and Romero, R. 2005. The embryonicdevelopment of the triclad Schmidtea polychroa. Dev. Genes Evol. 215:109–131.
Carranza, S., Baguna, J., and Riutort, M. 1997. Are the Platyhelminthes amonophyletic group? An assessment using 18S rRNA. Mol. Biol. Evol.14: 485–497.
Czubaj, A., and Niewiadomska, K. 1997. The muscular system of the cerc-aria of Diplostomum pseudospathaceum Niew., 1984 (Digenea): a phal-loidin-rhodamine fluorescence and TEM study. Acta Parasitol. 42: 199–218.
Faubel, A. 1983. The Polycladida, Turbellaria. Proposal and establishmentof a new system. Part I. The Acotylea. Mitt. Hamb. Zool. Mus. Inst. 80:17–121.
Faubel, A. 1984. The Polycladida, Turbellaria. Proposal and establishmentof a new system. Part II. The Cotylea. Mitt. Hamb. Zool. Mus. Inst. 81:189–259.
Hartenstein, V., and Ehlers, U. 2000. The embryonic development of therhabdocoel flatworm Mesostoma lingua (Abildgaard, 1789). Dev. GenesEvol. 210: 399–415.
Hartenstein, V., and Jones, M. 2003. The embryonic development of thebody wall and nervous system of the cestode flatworm Hymenolepsisdiminuta. Cell Tissue Res. 311: 427–435.
Hill, S. D., and Boyer, B. C. 2001. Phalloidin labeling of developingmuscle in embryos of the polychaete Capitella sp. I. Biol. Bull. 201:257–258.
Hooge, M. D. 2001. Evolution of body-wall musculature in the Platy-helminthes (Acoelomorpha, Catenulida, Rhabditophora). J. Morphol.249: 171–194.
Hooge, M. D., and Tyler, S. 1999. Musculature of the facultativeparasite Urastoma cyprinae (Platyhelminthes). J. Morphol. 241:207–216.
Hooge, M. D., and Tyler, S. 2004. Comparative morphology of the bodywall in flatworms (Platyhelminthes). Can. J. Zool. 82: 194–210.
Hyman, L. H. 1951. The Invertebrates. Platyhelminthes and Rhyn-chocoela: The Acoelomate Bilateria. Vol. II. McGraw-Hill, New York,vii1572pp.
Lacalli, T. C. 1982. The nervous system and ciliary band of Muller’s larva.Proc. R. Soc. Lond. B 217: 37–58.
Ladurner, P., and Rieger, R. 2000. Embryonic muscles development ofConvoluta pulchra (TurbellariaFAcoelomorpha, Platyhelminthes). Dev.Biol. 222: 359–375.
Ladurner, P., Scharer, L., Salvenmoser, W., and Rieger, R. M. 2005. Mac-rostomum lignano, n. sp. (Rhabditophora, Macrostomorpha): a newmodel organism among the lower Bilateria and the use of digital videomicroscopy in taxonomy of meiobenthic Platyhelminthes. J. Zool. Syst.Evol. Res. 43: 114–126.
Lang, A. 1884. Die Polycladen (Seeplanarien) des Golfes von Neapel underder angrenzenden Meeresabschnitte. Eine Monographie. Fauna FloraGolfes v. Neapel, Leipzig.
Litvaitis, M. K., and Rohde, K. 1999. A molecular test of platyhelminthphylogeny: inferences from partial 28S rDNA sequences. Invertebr. Biol.118: 42–56.
Mair, G. R., Maule, A. G., Day, T. A., and Halton, D. W. 2000. A confocalmicroscopical study of the musculature of adult Schistosoma mansoni.Parasitology 121: 163–170.
Mair, G. R., Maule, A. G., Fried, B., Day, T. A., and Halton, D. W. 2003.Organization of the musculature of schistosome cercariae. J. Parasitol.89: 623–625.
Mair, G. R., Maule, A. G., Shaw, C., Johnston, C. F., and Halton, D. W.1998. Gross anatomy of the muscle systems of Fasciola hepatica as vi-sualized by phalloidin-fluorescence and confocal microscopy. Parasito-logy 117: 75–82.
Maslakova, S. A., Martindale, M. Q., and Norenburg, J. L. 2004. Vestigialprotoroch in a basal nemertean, Carinoma tremaphoros (Nemertea; Pal-aeonemertea). Evol. Dev. 6: 219–226.
McDougall, C., Chen, W., Shimeld, S. M., and Ferrier, D. E. K. 2006. Thedevelopment of the larval nervous system, musculature and ciliary bandsof Pomatoceros lamarckii (Annelida): heterochrony in polychates. Front.Zool. 3: 16–29.
Morris, J., Nallur, R., Ladurner, P., Egger, B., Rieger, R., and Hartenstein,V. 2004. The embryonic development of the flatworm Macrostomum sp.Dev. Genes Evol. 214: 220–239.
Nielsen, C. 2001. Animal Evolution: Interrelationship of the Living Phyla.2nd Ed. Oxford University Press, Oxford, 563pp.
Prudhoe, S. 1985. A Monograph on Polyclad Turbellaria. Department ofZoology, British Museum (Natural History), Oxford University Press,Oxford, 259pp.
Rawlinson, K., Bolanos, D. M., Liana, M. K., and Litvaitis, M. K. 2008.Reproduction, development and parental care in two direct-developingflatworms (Platyhelminthes: Polycladida: Acotylea). J. Nat. Hist. 42:2173–2192.
Reiter, D., Boyer, B., Ladurner, P., Mair, G., Salvenmoser, W., and Rieger,R. 1996. Differentiation of the body wall musculature in Macrostomumhystricinum marinum and Hoploplana inquilina (Plathelminthes), as mod-els for muscle development in lower Spiralia. Roux’s Arch. Dev. Biol. 205:410–423.
Rieger, R., and Ladurner, P. 2001. Searching for the stem species of theBilateria. Belg. J. Zool. 131 (suppl. 1): 27–34.
Rieger, R., and Ladurner, P. 2003. The significance of muscle cells for theorigin of mesoderm in Bilateria. Integr. Comp. Biol. 43: 47–54.
Rieger, R., Salvenmoser, W., Legniti, A., and Tyler, S. 1994. Phalloidin-rhodamine preparations of Macrostomum hystricinum marinum(Plathelminthes): morphology and postembryonic development of themusculature. Zoomorphology 114: 133–147.
Rieger, R., et al. 1991. Organization and differentiation of the body-wallmusculature in Macrostomum (Turbellaria, Macrostomidae). Hydrobio-logia 227: 119–129.
Ruppert, E. E. 1978. A review of metamorphosis of turbellarian larvae. InF.-S. Chia and M. E. Rice (eds.). Settlement and Metamorphosis of Ma-rine Invertebrate Larvae. Elsevier, New York, pp. 65–81.
Semmler, H., Bailly, X., and Wanninger, A. 2008. Myogenesis in the basalbilaterial Symsagittifera roscoffensis (Acoela). Front. Zool. 5: 14–28.
Wanninger, A., and Haszprunar, G. 2002. Chiton myogenesis: perspectivesfor the development and evolution of larval and adult muscle systems inmollusks. J. Morphol. 251: 103–113.
Wanninger, A., Koop, D., Bromham, L., Noonan, E., and Degnan, B. M.2005. Nervous and muscle system development in Phascolion strombus(Sipuncula). Dev. Genes Evol. 215: 509–518.
Younossi-Hartenstein, A., and Hartenstein, V. 2000. The embryonic devel-opment of the polyclad Imogine mcgrathi.Dev. Genes Evol. 210: 383–398.
Embryonic muscle development in marine £atworms 301Bolan� os and Litvaitis












![Arthur Veis1,4, Kevin Tompkins1, Keith Alvares1, Kuiru ... · A rat incisor tooth odontoblast-pulp cDNA library was screened using ... [8,11] by embryonic rat muscle fibroblasts (EMF)](https://img.pdfslide.net/doc/110x75/5e88b9b3a5a6643ec265d245/arthur-veis14-kevin-tompkins1-keith-alvares1-kuiru-a-rat-incisor-tooth-odontoblast-pulp.jpg)
















