Embed Size (px)
Citation preview

A novel mycovirus that is related to the human pathogen Hepatitis E virus and 1
rubi-like viruses 2
Huiquan Liu1,2
, Yanping Fu2, Daohong Jiang
1,2﹡, Guoqing Li
1,2, Jun Xie
1,2, Youliang Peng
3, 3
Xianhong Yi2, Said A Ghabrial
4 4
1, National Key Laboratory of Agriculture Microbiology, Huazhong Agricultural University, Wuhan 5
430070, Hubei Province, P R China 6
2, The Provincial Key Lab of Plant Pathology of Hubei Province, College of Plant Science and 7
Technology, Huazhong Agricultural University, Wuhan, 430070, Hubei Province, P R China 8
3, Department of Plant Pathology, China Agricultural University, Yuanmingyuan West Road No. 1, 9
Haidian District, 100093, Beijing, P R China 10
4, Department of Plant Pathology, University of Kentucky, 201F Plant Science Building, 1405 11
Veterans Drive, University of Kentucky, Lexington, KY 40546-0312, USA 12
﹡ Corresponding author 13
Dr Daohong Jiang, Professor 14
Plant Pathology, College of Plant Science and Technology, Huazhong Agricultural University 15
Wuhan, 430070, Hubei Province, P R China 16
Tel: 86-27-87280487; Fax: 86-27-87397735; E-mail: [email protected] 17
﹟Current address: The college of Life Science, Hainan University, Haikou, Hainan Province, P R 18
China 19
Total number of words in text: 4266 20
Tables,2 and figures, 5 21
Summary: 287 words 22
Supplementary materials: Table, 1 and figures, 2 23
Running title: Mycovirus related to hepatitis E virus 24
GenBank Accession Number:EU779934 25
ACCEPTED
Copyright © 2008, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.J. Virol. doi:10.1128/JVI.01897-08 JVI Accepts, published online ahead of print on 10 December 2008
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

Abstract 26
Previously, we reported that three dsRNA segments, designated L, M and S, were detected in 27
Sclerotinia sclerotiorum strain Ep-1PN. Of these, the M dsRNA segment was derived from the 28
genomic RNA of a potexvirus-like positive-strand RNA virus, Sclerotinia sclerotiorum 29
debilitation-associated RNA virus. Here we present the complete nucleotide sequence of the L 30
dsRNA, which is 6043 nucleotides in length, excluding the poly (A) tail. Sequence analysis 31
revealed the presence of a single open reading frame (nt positions 42-5936) that encodes a protein 32
with significant similarity to the replicases of “alphavirus-like” supergroup of positive-strand RNA 33
viruses. Sequence comparison of the L dsRNA-encoded putative replicase protein containing 34
conserved methyltransferase, helicase and RNA-dependent RNA polymerase motifs showed that it 35
has significant sequence similarity to the replicase of Hepatitis E virus (HEV), a virus infecting 36
humans. Furthermore, we presented convincing evidence that the virus-like L-dsRNA could 37
replicate independently with slight impact on growth and virulence of its host. Our results suggest 38
that the L dsRNA from strain Ep-1PN is derived from the genomic RNA of a positive-strand RNA 39
virus, which we named Sclerotinia sclerotiorum RNA virus L (SsRV-L). As far as we know, this is 40
the first report of a positive-strand RNA mycovirus that is related to a human virus. Phylogenetic 41
and sequence analyses of the conserved motifs of the RNA replicase of SsRV-L showed that it 42
clustered with the rubi-like viruses, and that it is related to the plant clostero-, beny- and 43
tobamoviruses and to the insect omegatetraviruses. Considering the fact that these related 44
alphavirus-like positive-strand RNA viruses infect a wide variety of organisms, these findings 45
suggest that the ancestral positive-strand RNA viruses might be of ancient origin and/or they might 46
have radiated horizontally among vertebrates, insects, plants and fungi. 47
KEY WORDS: Mycovirus, Sclerotinia sclerotiorum, Sclerotinia sclerotiorum RNA virus L, 48
Hypovirulence, Hepatitis E Virus, Virus evolution, Strain Ep-1PN, Sclerotinia sclerotiorum 49
debilitation-associated RNA virus 50
51
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

Introduction 52
Sclerotinia sclerotiorum is a destructive soil borne plant pathogenic fungus with a wide host 53
range that includes more than 450 species and subspecies among 64 genera of plants (6). As an 54
important and unique plant pathogenic fungus, the sequence of the whole genomic DNA of S. 55
sclerotiorum has been determined 56
(http://www.ncbi.nlm.nih.gov/sutils/genom_table.cgi?organism=fungi). DsRNA-associated 57
hypovirulence in S. sclerotiorum was first reported for strain 91 (5) and later for strain Ep-1PN (36). 58
In strain Ep-1PN, three dsRNA segments, designated L, M and S-dsRNAs with estimated sizes of 59
7.4, 6.4 and 1.0 kbp, respectively, were associated with hypovirulence of S. sclerotiorum (36). Of 60
these three dsRNA segments, only the M-dsRNA was consistently detected in association with the 61
hypovirulence phenotype (36). Evidence was recently presented that the M-dsRNA was derived 62
from the genomic RNA of a positive-strand RNA virus, Sclerotinia sclerotiorum 63
debilitation-associated RNA virus (SsDRV) (67). Furthermore, sequence analysis of the S-dsRNA 64
segment showed that it is a defective RNA derived from SsDRV. The L-dsRNA segment may 65
represent a novel mycovirus different from SsDRV since it lacks sequence similarity to SsDRV, as 66
determined by northern hybridization analysis (Daohong Jiang, unpublished data). 67
Discovery of novel mycoviruses may expand our knowledge of global ecology and evolution 68
of viruses. Although mycoviruses typically have isometric particles and dsRNA genomes (e.g., 69
members of the families Totiviridae, Chrysoviridae and Partitiviridae) (14), viruses in these 70
families also infect organisms other than fungi. Whereas some members in the family Totiviridae 71
infect protozoa, a number of the viruses in the families Partitiviridae and Chrysoviridae infect 72
plants. The mycoreoviruses from hypovirulent strains of Cryphonectria parasitica and Rosellinia 73
necatrix represent a distinct group of dsRNA mycoviruses with reovirus-like particle morphology, 74
and they are most closely related to the tick-borne animal pathogens belonging to the genus 75
Coltivirus in the family Reoviridae (19, 52). Viruses with dsRNA genomes infect a broad range of 76
hosts (vertebrates, invertebrates, fungi, plants, protozoa, and bacteria) and are grouped in six 77
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

families of dsRNA viruses: Totiviridae, Birnaviridae, Partitiviridae, Cystoviridae, Chrysoviridae, 78
and Reoviridae (10). Comparative analysis of the amino acid sequences of proteins encoded by 79
dsRNA viruses revealed little similarity between viruses of different genera, even those belonging 80
to the same family, e.g. those belonging to the family Reoviridae. Even though the RdRps are the 81
most highly conserved genes among RNA viruses, phylogenetic analysis of the RdRps suggests a 82
polyphyletic origin for dsRNA viruses. The dsRNA viral RdRps tend to group with different 83
supergroups of the positive-strand RNA viruses (29). The concept of multiple origins of dsRNA 84
viruses from diverse lineages of positive-strand RNA viruses is presently well accepted (1, 29). 85
Recently, there is an increasing number of reports of positive-strand RNA mycoviruses whose 86
RdRp and helicase gene lineages are within the lineages of positive-strand RNA plant viruses: e.g., 87
the potexvirus-like mycoviruses FgV-DK21 in Fusarium graminearum (33), Botrytis virus X (22) 88
and Oyster mushroom spherical virus (65). Many of these positive-strand RNA mycoviruses do not 89
encode coat proteins and they occur in their hosts as dsRNA derivatives of their genomic 90
positive-strand RNAs, but are phylogenetically related to plant viruses. The mycoviruses that lack 91
typical virions include: members of the genus hypovirus that infect Cryphonectria parasitica, with 92
lineage to plant potyviruses (44); SsDRV, an unassigned mycovirus from Sclerotinia sclerotiorum 93
(67), which is related to allexiviruses in the family Flexiviridae, and Diaporthe ambigua RNA virus 94
(DaRV), with lineage to tombusviruses (47). Mitoviruses that infect C. parasitica (46), 95
Ophiostoma novo-ulmi (20) and Botrytis cinerea (66) are phylogenetically related to positive-strand 96
RNA bacteriophages in the family Leviviridae. Considering the fact that these related 97
positive-strand RNA viruses infect a wide variety of organisms, the ancestral RNA virus might be of 98
ancient origin and/or might have spread out horizontally among animals, plants, fungi, protozoa and 99
prokaryotes. 100
Some mycoviruses are associated with debilitation/hypovirulence of their hosts, and these 101
mycoviruses are potential bio-control agents to combat plant fungal diseases and to probe the 102
pathogenicity of host on molecular level (44). Among the debilitation/hypovirulence-associated 103
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

mycoviruses, the hypovirus/C. parasitica system has been the most thoroughly studied. Significant 104
insight has been gained on the molecular basis of hypovirulence in this system and its potential 105
implementation for biological control of chestnut blight (27, 38, 44, 55). The depth of knowledge 106
gained from studying the hypovirus/C. parasitica system should now pave the way for 107
investigations on other similar fungal virus systems. 108
In a recent study, Li et al. (37) identified a small number of genes whose expression was down 109
regulated in the virus-infected S. sclerotiorum strain Ep-1PN and discussed the probability that the 110
predicted depleted levels of the corresponding proteins may contribute to the characteristic 111
debilitation and hypovirulence of this strain. In the present study, molecular cloning and 112
sequencing of the L-dsRNA segment from a debilitated fungal strain were carried out and the 113
sequences generated were assembled and subjected to sequence and phylogenetic analyses to 114
determine whether the L-dsRNA is related to previously characterized mycoviruses and to examine 115
its relationships to viruses infecting organisms other than fungi. 116
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

Methods and materials 117
Fungal Strains 118
S. sclerotiorum hypovirulent strain Ep-1PN, which contained L-dsRNA, M-dsRNA (SsDRV), 119
and S-dsRNA, was originally isolated from a sclerotium collected from a diseased eggplant (35). 120
Strain Ep-1PNA367, a virus-free strain, was a single-ascospore isolate derived from Ep-1PN. Strain 121
Ep-1PNSA-8, Ep-1PNSA-23 and Ep-1PNSA-34 were isolated from individual sclerotia of strain 122
Ep-1PN. All fungal strains were grown at 18–22 oC on potato dextrose agar medium (PDA) and 123
stored on PDA slants at 4–8 oC. 124
Extraction of double-stranded RNA 125
The procedure for dsRNA extraction, previously described by De Paulo and Powell (7), was 126
used with minor modifications. For the extraction of dsRNA from mycelium of strains Ep-1PNSA-8, 127
Ep-1PNSA-23, Ep-1PNSA-34 and Ep-1PNA367, small agar mycelial plugs were placed on 128
cellophane membranes placed on top of the PDA medium (CM-PDA) in Petri plates for 2 days, and 129
then mycelium was harvested from the cellophane membranes. To extract the dsRNA from strain 130
Ep-1PN, the mycelium growing on CM-PDA for up to one week was harvested, and then 131
homogenized in a sterilized mortar with a pestle. The homogenate was spread on fresh CM-PDA 132
plates with cellophane membranes for 2 days, and then mycelium was harvested and stored at -80 133
oC. 134
cDNA synthesis, molecular cloning and sequencing 135
To obtain sequence information for the L-dsRNA, dsRNA (1.0 µg) was mixed with 0.1 µg 136
random hexamer primers and 3 µl 100% DMSO, and DEPC-treated ddH2O was added to a final 137
volume of 12 µl. The mixture was heated at 95–98 oC for 14 min and chilled on ice for 3 min. 138
First and second-strand cDNAs were synthesized as described by Sambrook et al. (51). The 139
resulting cDNA was purified by filtration through a Sephadex G-50 column and A-tailed with Taq 140
DNA polymerase and dNTP at 72 oC for 30 min. The A-tailed double stranded cDNA was ligated 141
into the pMD18-T vector according to the manufacturer’s instructions (TaKaRa) and transformed 142
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

into competent cells of E. coli JM109. Sequence-specific primers were used for RT-PCR to amplify 143
parts of the genome which were not cloned by the initial random cDNA synthesis. Denatured 144
dsRNA was reverse transcribed using RevertAidTM
M-MuLV Reverse Transcriptase (Fermentas) 145
and a sequence-specific reverse primer and incubated for 60 min at 45 o
C. After reverse 146
transcription, the mixture was treated with RNase H (1 U at 37 oC for 30 min; TaKaRa) and 2 % of 147
the reaction volume was used for PCR amplification with the pertinent forward and reverse primers, 148
GC Buffer and LA Taq DNA Polymerase (TaKaRa). The resulting PCR product was fractionated by 149
electrophoresis on 1% agarose gel and purified using a gel extraction kit (Axygen). The PCR 150
product was cloned into the pMD18-T vector. 151
Clones for the terminal sequences of the dsRNA were generated by T4 RNA ligase 152
oligonucleotide-mediated amplification as described by Lambden et al. (34). The 3’ terminus of 153
each strand of dsRNA was ligated at 5–15 o
C for 16–18 h with the 5’-end phosphorylated 154
oligonucleotide 5’-GCATTGCATCATGATCGATCGAATTCTTTAGTGAGGGTTAATTGCC- 155
(NH2)-3’ using T4 RNA ligase (Fermentas). The oligo-nucleotide-ligated dsRNA was denatured 156
and used for the reverse transcription reaction with RevertAidTM
M-MuLV Reverse Transcriptase 157
and 10 pmol of a primer with sequence complementary to the oligonucleotide used for the RNA 158
ligation (oligoREV, 5’-GGCAATTAACCCTCACTAAAG-3’). The reaction product was treated 159
with RNase H, as described above, and the cDNA was amplified with another primer 160
complementary to the RNA ligation oligonucleotide (5’-TCACTAAAGAATTCGATCGATC-3’) 161
and the sequence-specific primer corresponding to the 5’- and 3’-terminal sequences of the dsRNA, 162
respectively. The expected PCR products were recovered and purified with a gel extraction kit 163
(Axygen), cloned into the pMD18-T vector (TaKaRa). 164
Sequencing was carried out by the dideoxynucleotide termination method using a Big Dye 165
Terminator Sequencing kit (BigDye terminator v. 2.0; ABI) and an ABI PRISM 377-96 automated 166
sequencer (Beijing Sunbiotech). M13 universal primers or sequence-specific primers were used for 167
sequencing and each base was determined by sequencing at least two independent clones (usually 168
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

three to five clones) from both orientations. 169
Sequence and phylogenetic analyses 170
The DNAMAN 5.2.9 version (Lynnon Biosoft, USA) software package was used for sequence 171
annotations, including nucleotide statistics and open reading frame (ORF) searching. Sequence 172
similarity searches of GenBank, Swissprot and EMBL databases were conducted using the BLAST 173
program (3). Searches for amino acid signatures and protein motifs were conducted using the 174
programs included in the ExPASy proteomics tools (http://www.expasy.org/tools/). Multiple 175
alignments of amino acid were made with the program MUSCLE Version 3.6 (9) and the resulting 176
alignment was manually adjusted according to Koonin’s alignments (28). Two independent methods 177
for the generation of tentative phylogenetic trees were used, namely Neighbor-joining (NJ) 178
algorithm and Maximum likelihood (ML) method. Neighbor-joining (NJ) algorithm was performed 179
using PAUP* 4.0b10 (59), assuming the BLOSUM 62 matrix (18). Bootstrap values were 180
calculated from 1000 bootstrap replicates. Maximum likelihood (ML) method was performed 181
using program TREE-PUZZLE version 5.2 (54). Likelihoods were calculated using the VT model 182
of amino acid substitution (42) and the relevant parameter values estimated from the data (available 183
upon request). 184
Northern blot hybridization 185
Northern hybridization analysis was performed as previously described (25). To verify the 186
authenticity of the cDNA clones generated with the purified dsRNA, the cDNA clones were labeled 187
with [32
P] dCTP using a radio-labeling kit (TaKaRa) and used to probe the RNA blot. 188
Reverse transcription-PCR 189
Total RNA from isolates derived from the debilitated strain Ep-1PN was isolated according to 190
Sambrook et al. (51). First-strand cDNA was synthesized using RevertAidTM
M-MuLV Reverse 191
Transcriptase (Fermentas). To detect M-dsRNA, the reverse primer SsDRV-PCRpREV 192
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

(5’-CAGTCCCTAGTTTCATCTCGTTCC-3’) was used and the first-strand cDNA was then 193
subjected to PCR using the SsDRV-PCRpREV primer and the forward primer SsDRV-PCRpFOR 194
(5’-TGCAGGAAACAGTCATGGCAAC-3’) with a predicted size of 871 bp. To detect L-dsRNA, 195
the reverse primer Ss-7.4RP (5’-GAAGCCACAGGGACAGCAAG-3’) was used and the 196
first-strand cDNA was then subjected to PCR using the Ss-7.4RP primer and the forward primer 197
Ss7.4-FP (5’-CCACCGACGCAGGCAAATAC-3’) with a predicted size of 721 bp. The conditions 198
for cDNA amplification included an initial denaturation step of 4 min at 94℃, followed by 30 199
cycles of 30 sec at 94℃, 30 sec at 61℃ and 1 min at 72℃, with a final elongation step of 10 min at 200
72℃. PCR products were fractionated by gel electrophoresis on 1% agarose gels and stained with 201
ethidium bromide. 202
Mycelial growth and virulence test 203
To evaluate the effect of SsRV-L on colony morphology and virulence on rapeseed of 204
Sclerotinia sclerotiorum, SsRV-L-infected strains were compared with the original strain Ep-1PN 205
and virus-free ascospore offspring of strain Ep-1PN using the procedures of Li et al. (35). 206
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

Results 207
Synthesis and sequencing of cDNA from L-dsRNA 208
The L-dsRNA segment extracted from mycelia of strain Ep-1PN was electrophoretically 209
separated on 1% agarose gel and purified with a gel extraction kit, and then, subjected to cDNA 210
synthesis using random primers (hexamer). The ds-cDNA fragments were cloned and the cloned 211
cDNAs were transformed into E. coli strain JM109. More than forty cDNA clones with inserts of 212
200 to 800 bp were obtained and confirmed to be derived from the L-dsRNA using Reverse 213
Northern dot-blot hybridization analysis (data not shown). Among these, 10 randomly selected 214
cDNA clones were sequenced in both directions (Fig. 1A; clones A to J). RT-PCR was conducted to 215
fill the gaps between clones with specific primers designed based on these cDNA sequences, and 216
used an oligo-ligation strategy to obtain the sequences of the 5' and 3' termini. A total of 26 clones 217
were obtained and sequenced (Fig. 1A). Computer-assisted sequence assembly showed that the 218
full-length L-dsRNA cDNA is 6043 bp in length, excluding the poly(A) tail. The cloning strategy 219
for dsRNA is outlined in Fig. 1A. The sequence was deposited in the GenBank under accession no. 220
EU779934. 221
Sequence analysis of the complete cDNA of the L-dsRNA 222
Analysis of the complete cDNA sequence revealed the presence of a single large open reading 223
frame (ORF; nt 42–5936), potentially encoding a polypeptide of 1964 amino acid residues with a 224
predicted mass of 213.4 kDa (Fig. 1B). Motif Scan searches showed that this protein contains 225
conserved methyltransferase, viral RNA helicase and RNA-dependent RNA polymerase (RdRp) 226
domains characteristic of the replicases of many of the positive-strand RNA viruses in the 227
alphavirus-like supergroup of viruses (Fig. 1B). The 5’ noncoding region consists of 41 nucleotides 228
and could be folded into a hairpin structure, and the 3’ noncoding region consists of 107 nucleotides 229
and could be folded into three independent hairpin structures (see Fig. S1 in the supplemental 230
material). Such hairpin structures are also found in other viruses in the alphavirus-like supergroup 231
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

(15, 17, 41, 43, 58). Homology searches of the methyltransferase, helicase and RdRp conserved 232
motifs of the L-dsRNA-encoded replicase indicated that they are related to several animal viruses 233
belonging to the genus Hepevirus including Hepatitis E virus (HEV), swine hepatitis E virus 234
(HEV-swine) and Avian hepatitis E virus (AHEV) (Fig. 2). BLASTP database searches of the 235
L-dsRNA RdRp conserved domain revealed that it shares significant sequence similarity (E values 236
of 3e-11
or lower) with the RdRp encoded by the hepeviruses and by the closteroviruses. The RdRp 237
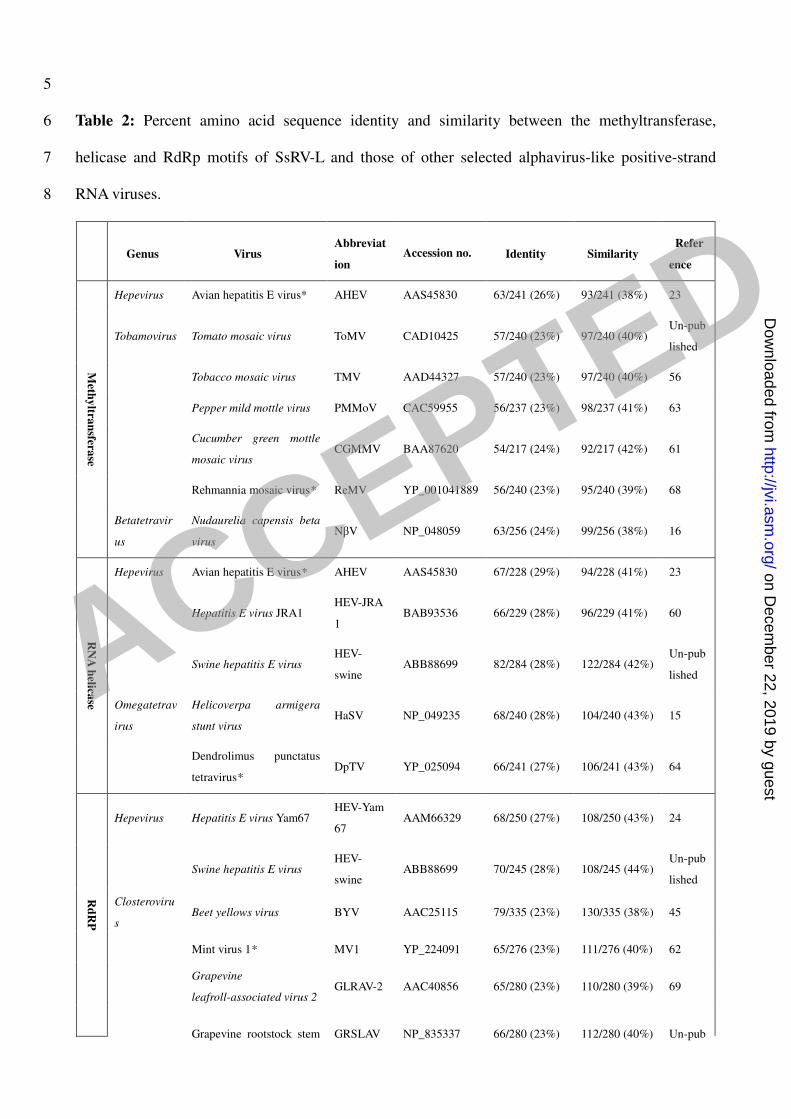
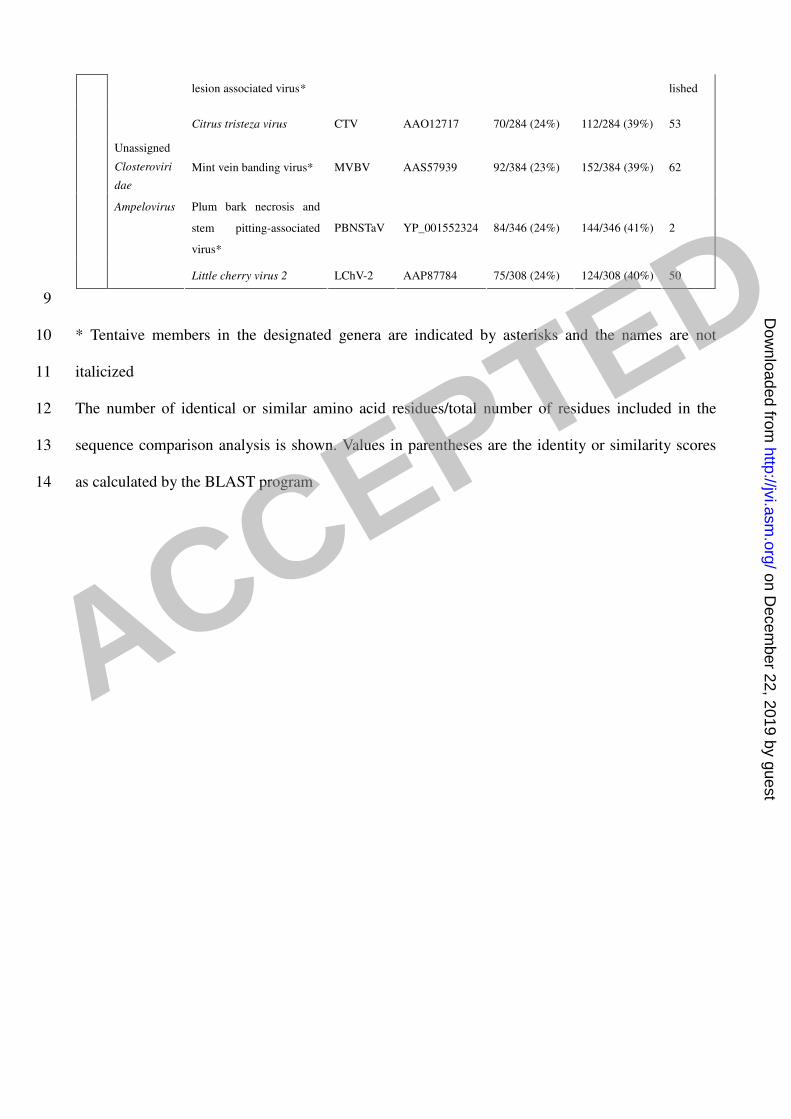
domain of L-dsRNA shares 27% identity and 43% similarity with HEV, and 28% identity and 44% 238
similarity with HEV-swine (Table 2). These L-dsRNA RdRp conserved motifs are also related to 239
those of plant viruses in the genus Closterovirus. The RdRp conserved motifs of SsRV-L share 23 % 240
identity and 40 % similarity with mint virus 1. Likewise, the identity and similarity scores 241
between SsRV-L and plum bark necrosis and stem pitting-associated virus are 24 % and 41 %, 242
respectively. The corresponding scores for mint vein banding virus are 24 % and 40 % (Table 2). 243
Thus, these results suggest that the L-dsRNA probably represents the replicative form or a 244
replicative intermediate of the genomic RNA of a positive-strand mycovirus co-infecting the 245
debilitated S. sclerotiorum strain Ep-1PN along with SsDRV. The newly characterized virus 246
co-infecting strain Ep-1PN was designated Sclerotinia sclerotiorum RNA virus-L (SsRV-L). 247
Sequence comparison of the viral RNA helicase motifs of SsRV-L showed that they share 248
significant sequence similarity with viruses in the genera Hepevirus and Omegatetravirus (E values 249
of 4e-6
or lower). The helicase motifs identity and similarity scores between SsRV-L and AHEV or 250
HEV are 28 % and 41 %, or 29 % and 42 %, respectively. Interestingly, the RNA helicase motifs of 251
SsRV-L also share significant sequence similarities with the insect viruses Helicoverpa armigera 252
stunt virus (HaSV) and Dendrolimus punctatus tetravirus (DpTV), the percent identity and 253
similarity of the helicase domain between SsRV-L and those two insect viruses are 27 % and 41 %, 254
respectively (Table 2). Both HaSV and DpTV belong to the genus Omegatetravirus in the family 255
Tetraviridae (15, 64). 256
Furthermore, the methyltransferase motifs of SsDRV-L and AHEV share sequence similarities 257
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

with the tobamoviruses tomato mosaic virus, tobacco mosaic virus, pepper mild mottle virus, 258
cucumber green mottle mosaic virus and rehmannia mosaic virus, and the insect betatetravirus 259
Nudaurelia capensis beta virus (Table 2). 260
Phylogenetic analysis 261
Maximum likelihood distance comparisons of amino acid sequences of the RdRp domain of 262
SsRV-L and representative viruses of alphaviruses, endornaviruses, tymo-like viruses, rubi-like 263
viruses and tobamo-like viruses showed that SsRV-L is most closely related to Hepatitis E virus 264
belonging to the genus Hepevirus (see Table S1 in the supplemental material). Phylogenetic trees 265
based on multiple alignments of RdRp conserved motifs of SsRV-L and these viruses were 266
independently generated with Neighbor-joining (NJ) algorithm and Maximum likelihood (ML). The 267
resulting NJ and ML trees had similar topology and showed that SsRV-L clusters with several 268
rubi-like viruses including benyviruses, hepeviruses, omegatetravirus and rubivirus. The ML tree 269
is shown in Fig. 3A. 270
A phylogram based on multiple alignments of viral RNA helicase conserved motifs of SsRV-L 271
and representative alphaviruses, endornaviruses, tymo-like viruses, rubi-like viruses and 272
tobamo-like viruses was similar to the tree generated by multiple alignments of RdRp conserved 273
motifs (Fig. 3B). Likewise, a similar tree was generated based on the multiple alignments of 274
methyltransferase conserved motifs of SsRV-L and selected viruses (see Fig. S2 in the supplemental 275
material). 276
Evidence for autonomous replication of SsRV-L 277
The single-sclerotium isolates Ep-1PNSA-8, Ep-1PNSA28 and Ep-1PNSA32, which were 278
derived from strain Ep-1PN, lacked the M-dsRNA and S-dsRNA segments, but contained only the 279
L-dsRNA segment. This was confirmed with RT-PCR detection (Fig. 4). Thus, SsRV-L could 280
replicate independently in S. sclerotiorum. 281
Compared to the virus-free strain Ep-1PNA367, an ascospore isolate derived from strain 282
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

Ep-1PN, the hyphal growth and virulence on detached rapeseed leaves of strains Ep-1PNSA-8, 283
Ep-1PNSA28 and Ep-1PNSA32, were slightly reduced, but their growth and virulence were 284
significantly higher than those of the hypovirulent strain Ep-1PN (Fig. 5). The sclerotial growth was 285
normal on the PDA medium and there was little or no difference in colony morphology between 286
these strains and that of the wild type strain of S. sclerotiorum. Thus, SsRV-L contributes little if 287
any to the debilitation of strain Ep-1PN. 288
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

Discussion 289
This study represents the first report on molecular characterization of a positive strand RNA 290
mycovirus, SsRV-L that is phylogenetically related to the human pathogen, Hepatitis E virus (HEV) 291
and rubi-like viruses. Sequence analysis of the full-length cDNA clone containing the coding 292
sequence of a putative viral replicase revealed the presence of conserved methyltransferase, viral 293
RNA helicase and RdRp-2 superfamily motifs characteristic of the viral RNA replicase proteins of 294
positive-strand RNA viruses and with significant similarity to the human pathogen HEV, a member 295
of the genus Hepevirus (30). Like HEV, the genome of SsRV-L also has a short 5’-untranslated 296
leader sequence and 3’ untranslated region with poly(A) tail. However, unlike the genomes of 297
HEV and HEV-swine, which contain three ORFs, SsRV-L genomic RNA contains only a single 298
ORF (ORF1) with significant sequence similarity to ORF1 of HEV and those of HEV-swine and 299
AHEV. Furthermore, ORF1 of SsRV-L lacks coding sequences for papain peptidase or papain 300
peptidase-like proteins. Interestingly, the viral RNA helicase and methyltransferase motifs of 301
SsRV-L and HEV (including HEV-swine and AHEV) share significant sequence similarity with the 302
insect tetraviruses and the plant tobamo-like viruses (Table 2). 303
The positive strand RNA viruses were classified into three superfamilies: alpha-like, 304
picorna-like and flavi-like. The superfamily of alpha-like viruses comprises three lineages: rubi-like, 305
tobamo-like and tymo-like viruses (31). Whereas tobamo- and tymo-like viruses infect plants, the 306
rubi-like viruses infect plants, vertebrates and insects. Recently, evidence was presented that the 307
benyvirus (rubi-like lineage) Beet necrotic yellow vein virus (BNYVV) replicates in its 308
plasmodiophorid vector Polymyxa betae suggesting that BNYVV may also be considered a virus of 309
plasmodiophorids (39). Our results from phylogenetic analysis support the conclusion that 310
SsRV-L could be classified with the rubi-like viruses, thus, the host range of rubi-like viruses is 311
more diverse than once thought. The reason for the broad host range of rubi-like viruses is not 312
clearly known. 313
Although, SsRV-L shares sequence similarities with HEV and the insect viruses HaSV and 314
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

DpTV, it is more likely that theses positive-strand RNA viruses were derived independently from 315
plant viruses, possibly those related to tobamoviruses. Since vertebrates and insects, are not hosts of 316
S. sclerotiorum, it is unlikely that S. sclerotiorum acquired the ancestral SsRV-L from these 317
organisms. S. sclerotiorum, a plant pathogenic fungus, might have acquired the ancestral 318
SsRV-L-like virus from a virus-infected plant since it shares many hosts with viruses in the genus 319
Tobamovirus. As a matter of fact, the hypovirulent strain Ep-1PN was originally isolated from a 320
diseased plant of eggplant (Solanum melongena) (35). Humans might have obtained HEV from 321
herbivores, e.g., swine was one of the reservoirs of HEV (11), and wild animals, which could be 322
infected by HEV or HEV-like viruses (57). Insects, like Helicoverpa armigera and Dendrolimus 323
punctatus, could have obtained HaSV and DpTV from plants, and it is feasible that birds obtained 324
avian HEV from virus-infected insects. Although these inferences are consistent with the hypothesis 325
that the ancestral alpha-like positive-strand RNA virus might have spread out horizontally among 326
plants, vertebrates and insects, this does not rule out the ancient origin of the progenitor virus in a 327
single cell type prior to the separation of fungi, plants and animals. Recently, Koonin et al. (32) 328
presented evidence that picorna-like virus evolution antedates the radiation of eukaryotic 329
supergroups. 330
The genomic organization for SsRV-L and SsDRV are similar to each other, both of them lack 331
unnecessary genes including genes for coat protein and movement protein. Hypoviruses and DaRV 332
are also examples of mycoviruses that lack coat and movement proteins. These viruses, like SsRV-L 333
and SsDRV, are phylogenetically related to plant viruses. Botrytis virus F (BVF), represents an 334
example of a mycovirus that is phylogenetically related to plant viruses (potexviruses) but which 335
codes for a coat protein in addition to the replicase (21). Unlike potexviruses, however, BVF does 336
not code for a movement protein. It is not known how viruses delete unnecessary genes from their 337
genomes; SsDRV, SsRV-L, DaRV, hypoviruses and similar viruses may represent examples of 338
regressive evolution by viruses in fungi. 339
The occurrence of SsRV-L and SsDRV in the hypovirulent strain Ep-1PN of S. sclerotiorum 340
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

represents a novel type of co-infection involving two positive strand RNA mycoviruses. Mixed 341
infections with two or more dsRNA viruses appear to be of common occurrence among 342
mycoviruses (14). Examples of mixed infections with dsRNA viruses belonging to distinct virus 343
families include the totivirus Hv190SV and the chrysovirus Hv145SV that co-infect C. victoriae (12, 344
13). There are many examples of mixed infections by distinct members of the same virus family 345
including the two totiviruses, SsRV-1 and SsRV-2 that co-infect Sphaeropsis sapinea (48), the two 346
totiviruses ScV-L-A and ScV-L-BC that infect Saccharomyces cerevisae and the partitiviruses 347
Penicillium stoloniferum viruses S and F that infect Penicillium stoloniferum. 348
As previously reported, it is not clear whether SsRV-L contributes to the hypovirulence 349
phenotype of strain Ep-1PN (36) since S. sclerotiorum isolates that carry SsRV-L alone exhibit a 350
normal phenotype. Thus, we assume that S. sclerotiorum might have acquired SsRV-L earlier than 351
SsDRV, and SsDRV could have been acquired through hyphal anastomosis with other 352
SsDRV-infected fungal strains since mycoviruses could be transmitted between vegetatively 353
incompatible strains though at lower frequencies. 354
Unlike C. parasitica that can produce both ascospores and conidial spores, S. sclerotiorum can 355
only produce ascospores that form in apothecia germinating from dormant sclerotia. Thus SsRV-L 356
and SsDRV cannot be dispersed via conidial spores. Furthermore, our previous work showed that 357
both SsRV-L and SsDRV could not be transmitted through ascospores of strain Ep-1PN (26, 67), 358
and hyphae do not present valid dormant material for dispersal of S. sclerotiorum. Therefore, 359
SsRV-L and SsDRV could only be transmitted and dispersed by sclerotia, and the distribution of 360
SsRV-L and SsDRV would then be confined to limited areas. Moreover, the survival ability of 361
fungal strains doubly infected with SsRV-L and SsDRV is predicted to be very low since they grow 362
slowly, virtually lose their virulence and produce only few sclerotia (35). Thus, we reasoned that 363
horizontal transfer of the ancestral viruses of SsRV-L and SsDRV from other fungi or plants to S. 364
sclerotiorum might have occurred relatively recently. Because S. sclerotiorum has a broad host 365
range of plants known to be susceptible to many closteroviruses and potexviruses, the possibility 366
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

that closterovirus-like (SsRV-L) and potexvirus-like (SsDRV) viruses might have transferred from 367
plants to S. sclerotiorum seems feasible (8, 40). 368
SsRV-L appears to be more stable than SsDRV in S. sclerotiorum since subcultures that 369
contain SsRV-L but lacks SsDRV are relatively easy to obtain by hyphal tipping and single 370
sclerotium isolation. Like SsDRV, SsRV-L is also eliminated through sexual reproduction of S. 371
sclerotiorum; this phenomenon is common among mycoviruses of higher ascomycetous hosts, e.g., 372
the hypovirus/C. parasitica system (4), but the underlying mechanism is not known. 373
Currently, it is not understood whether there is an interaction between SsRV-L and SsDRV in 374
doubly infected S. sclerotiorum and whether this interaction has any bearing on the debilitation 375
phenotype since the two viruses can replicate independently. S. sclerotiorum strains singly-infected 376
with SsRV-L show little or no adverse effects, whereas strains that are singly infected with SsDRV 377
exhibit a debilitated phenotype. In a recent study, Li et al. (37) identified several genes whose 378
expression was down regulated in a doubly infected S. sclerotiorum strain. The 379
hypovirulence/debilitation system of Sclerotinia sclerotiorum and its associated mycoviruses 380
presents an attractive system to explore the molecular basis of pathogenicity in this devastating 381
plant pathogen (37). Future construction of infectious full-length cDNA clones for these two viruses 382
and the development of RNA transfection systems would be useful in deciphering the interaction 383
between SsRV-L and SsDRV-1 and their effects on their common host. 384 ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

Acknowledgment 385
This work was supported by grants from the National Basic Research Program (2006CB101901-1), 386
the Program for New Century Excellent Talents in University (NCET-06-0665) and the Fok Ying 387
Tong Education Foundation for Young Teacher of Universities and Colleges (No. 80125), and 388
our publication is in the memory of Mr Fok Ying Tong. We also thank the anonymous reviewers for 389
their constructive and helpful comments. 390
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

References 391
1. Ahlquist, P. 2006. Parallels among positive-strand RNA viruses, reverse- transcribing viruses 392
and double-stranded RNA viruses. Nature Rev. Microbiol. 4:371-382. 393
2. Alrwahnih, M., Uyemoto, J. K., Falk, B. W. & Rowhani, A. 2007. Molecular characterization 394
and detection of plum bark necrosis stem pitting-associated virus. Arch Virol 152:2197-2206. 395
3. Altschul, S. F., Madden, T. L., Scha¨ ffer, A. A., Zhang, J., Zhang, Z., Miller, W. & Lipman, 396
D. J. 1997. Gapped BLAST and PSI-BLAST: a new generation of protein database search 397
programs. Nucleic Acids Res 25:3389-3402. 398
4. Anagnostakis, S. L. 1982. Biological control of chestnut blight. Science 215:466-471. 399
5. Boland, G. J. 1992. Hypovirulence and double-stranded RNA in Sclerotinia sclerotiorum. Can J 400
Plant Pathol 14:10-17. 401
6. Boland, G.F. & Hall, R. 1994. An index of plant hosts susceptible to Sclerotinia sclerotiorum. 402
Can J Plant Pathol 16:93-108. 403
7. De Paulo, J. J. and Powell, C. A. 1995. Extraction of double-stranded RNA from plant tissues 404
without the use of organic solvents. Plant Dis. 79:246-248. 405
8. Dolja VV, Karasev AV and Koonin EV. 1994. Molecular Biology and Evolution of 406
Closteroviruses: Sophisticated Build-up of Large RNA Genomes. Annual Review of 407
Phytopathology 32:261-285. 408
9. Edgar, R. C. 2004, MUSCLE: multiple sequence alignment with high accuracy and high 409
throughput, Nucleic Acids Research 32:1792-97. 410
10. Fauquet, C.M., Mayo, M.A., Maniloff, J., Desselberger, U., and Ball, L.A. 2005. Virus 411
Taxonomy: Eighth Report of the International Committee for theTaxonomy of Viruses (San 412
Diego: Academic Press). 413
11. Feagins A. R., Opriessnig T., Guenette D. K., Halbur P. G., and Meng X.-J. 2007. Detection 414
and characterization of infectious Hepatitis E virus from commercial pig livers sold in local 415
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

grocery stores in the USA. J. Gen. Virol. 88:12-917. 416
12. Ghabrial, S. A. 1986. A transmissible disease of Helminthosporium victoriae evidence for a 417
viral etiology. In: Buck, K.W. (Ed.), Fungal Virology. CRC Press, Boca Raton, pp. 164-176. 418
13. Ghabrial, S. A., Soldevila, A. I. and Havens, W. M. 2002. Molecular genetics of the viruses 419
infecting the plant pathogenic fungus Helminthosporium victoriae. In Molecular biology of 420
double-stranded RNA: Concepts and applications in agriculture, forestry and medicine. pp. 421
213-236. Edited by S. Tavantzis. Boca Raton: CRC Press. 422
14. Ghabrial, S. A. and Suzuki, N. 2008. Fungal viruses. In Encyclopedia of Virology, 3rd edn, vol. 423
2, pp. 284-291. Edited by B. W. J. Mahy & M. H. V. Van Regenmortel. Oxford: Elsevier. 424
15. Gordon, K. H. J., Johnson, K. N., and Hanzlik, T. N. 1995. The larger genomic RNA of 425
Helicoverpa armigera stunt tetravirus encodes the viral RNA polymerase and has a novel 426
39-terminal tRNA-like structure. Virology 208:84-98. 427
16. Gordon, K. H., Williams, M. R., Hendry,D.A. and Hanzlik, T. N. 1999. Sequence of the 428
genomic RNA of nudaurelia beta virus (Tetraviridae) defines a novel virus genome 429
organization. Virology 258:42-53. 430
17. Hanzlik, T. N., Johnson, K. N., and Gordon K. H. J. 1995. Sequence of RNA2 of the 431
Helicoverpa armigera stunt virus (Tetraviridae) and bacterial expression of its genes. J. Gen. 432
Virol. 76:799-811. 433
18. Henikoff, S. and Henikoff, J. G. 1992. Amino acid substitution matrices from protein blocks. 434
Proc. Natl. Acad. Sci. USA, 89:10915–10919. 435
436
19. Hillman, B. I., Supyani, S., Kondo, H. and Suzuki, N. 2004. A reovirus of the fungus 437
Cryphonectria parasitica that is infectious as particles and related to the Coltivirus genus of 438
animal pathogens. Journal of Virology 78:892-898. 439
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

20. Hong, Y., Dover, S. L., Cole, T. E, Brasier, C. M. and Buck, K. W. 1999. Multiple 440
mitochondrial viruses in an isolate of the Dutch Elm disease fungus Ophiostoma novo-ulmi. 441
Virology 258:118-127. 442
21. Howitt, R. L. J., Beever, R. E., Pearson, M. N. and Forster, R. L. S. 2001. Genome 443
characterization of Botrytis virus F, a flexuous rod-shaped mycovirus resembling plant 444
'potex-like' viruses. J. Gen. Virol. 82:67-78. 445
22. Howitt, R. L. J., Beever, R. E., Pearson, M. N. and Forster, R. L. S. 2006 Genome 446
characterization of a flexuous rod-shaped mycovirus, Botrytis virus X, reveals high amino acid 447
identity to genes from plant 'potex-like' viruses. Arch. Virol. 151: 563-579. 448
23. Huang, F. F., Sun, Z. F., Emerson, S. U., Purcell, R. H., Shivaprasad, H. L., Pierson, F. W., 449
Toth, T. E. and Meng, X. J. 2004. Determination and analysis of the complete genomic 450
sequence of avian hepatitis E virus (avian HEV) and attempts to infect rhesus monkeys with 451
avian HEV. J. Gen. Virol. 85:1609-1618. 452
24. Jameel, S., Zafrullah, M., Chawla, Y. K. and Dilawari, J. B. 2002. Reevaluation of a North 453
India isolate of hepatitis E virus based on the full-length genomic sequence obtained following 454
long RT-PCR. Virus Res. 86:53-58. 455
25. Jiang, D. and Ghabrial, S. A. 2004. Molecular characterization of Penicillium chrysogenum 456
virus: reconsideration of the taxonomy of the genus Chrysovirus. J. Gen. Virol. 85:2111-2121. 457
26. Jiang, D., Li, G., Wang, D. and Zhou, Q. 2000. The sexual inheritance of cultural 458
characteristics of Sclerotinia sclerotiorum hypovirulent isolate Ep-1PN. Mycosystema 459
19:236-240. 460
27. Jacob-Wilk, D., Turina, M. and Van Alfen, N.K. 2006. Mycovirus cryphonectria hypovirus 1 461
elements cofractionate with trans-golgi network membranes of the fungal host Cryphonectria 462
parasitica. Journal of Virology 80:6588-6596. 463
28. Koonin, E. V. 1991. The phylogeny of RNA-dependent RNA polymerases of positive-strand 464
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

RNA viruses. J. Gen. Virol., 72:2197 - 2206. 465
29. Koonin, E. V. 1992. Evolution of double-stranded RNAviruses: a case for polyphyletic origin 466
from different groups of positive-stranded RNA viruses, Sem in Virol., 3:327-339. 467
30. Koonin, E. V., Gorbalenya, A. E., Purdy, M. A. and Rozanov, M. N. 1992. Computer-assisted 468
assignment of functional motifs in the nonstructural polyprotein of hepatitis E virus: 469
Delineation of an additional group of positive-strand RNA plant and animal viruses. Proc. Natl. 470
Acad. Sci. USA 89:8259-8263. 471
31. Koonin, E. V. and Dolja, V. V. 1993. Evolution and taxonomy of positive-strand RNA viruses: 472
implications of comparative analysis of amino acid sequences. Crit Rev Biochem Mol Biol 473
28:375-430. 474
32. Koonin, E. V, Wolf, Y. I., Nagasaki, K. and Dolja, V. V. 2008. The big bang of picorna-like 475
virus evolution antedates the radiation of eukaryotic supergroups. Nature Rev. Microbiol. 476
6:926-939. 477
33. Kwon, S. J., Lim, W. S., Park, S. H., Park, M. R. and Kim, K. H. 2007. Molecular 478
characterization of a dsRNA mycovirus, Fusarium graminearum Virus-DK21, which is 479
phylogenetically related to hypoviruses but has a genome organization and gene expression 480
strategy resembling those of plant potex-like viruses. Molecules and Cells 23:304-315. 481
34. Lambden, P. R., Cooke, S. J., Caul, E. O. and Clarke, I. N. 1992. Cloning of noncultivatable 482
human rotavirus by single primer amplification. J. Virol. 66:1817-1822. 483
35. Li, G., Wang, D., Huang, H. C. and Zhou, Q. 1996. Polymorphisms of Sclerotinia 484
sclerotiorum isolated from eggplant in Jiamusi, Heilongjiang Province. ACTA 485
Phytopathologica sinica 26:237-242. 486
36. Li, G., Jiang, D., Wang, D., Zhu, B. and Rimmer, R. 1999. Double-stranded RNAs associated 487
with the hypovirulence of Sclerotinia sclerotiorum strain Ep-1PN. Progress in Natural Science 488
9:836-841. 489
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

37. Li, H., Fu, Y. P., Jiang, D. H., Li, G. Q., Ghabrial, S. A. and Yi, X. H. 2008. Down-regulation 490
of Sclerotinia sclerotiorum gene expression in response to infection with Sclerotinia 491
sclerotiorum debilitation-associated RNA virus. Virus Res. 135:95-106 492
38. Lin, H. Y., Lan, X. W., Liao, H., Parsley, T. B., Nuss, D. L. and Chen, B. S. 2007. Genome 493
sequence, full-length infectious cDNA clone, and mapping of viral double-stranded RNA 494
accumulation determinant of hypovirus CHV1-EP721. Journal of Virology 81:1813-1820. 495
39. Lubicz, J. V., Rush, C. M., Payton, M., Colberg, T. 2007. Beet necrotic yellow vein virus 496
accumulates inside resting spores and zoosporangia of its vector Polymyxa betae BNYVV 497
infects P. betae. Virol J, 4:37. 498
40. Martelli, G. P., Adams, M. J., Kreuze,. J. F. and Dolja, V. V. 2007. Family Flexiviridae: A 499
case study in virion and genome plasticity. Annu. Rev. Phytopathol. 45:73-100. 500
41. Miller, E. D., Plante, C. A., Kim, K. H., Brown, J. W. and C. Hemenway. 1998. Stem-loop 501
structure in the 5' region of potato virus X genome required for plus-strand RNA accumulation. 502
J. Mol. Biol. 284:591-608. 503
42. Müller, T. and Vingron, M. (2000) Modeling amino acid replacement. J. Comput. Biol., 504
7:761–776. 505
43. Nakhasi, H. L., Rouault, T. A., Haile, D. J., Liu, T. Y. and Klausner, R. D. 1990. Specific 506
high affinity binding of host cell proteins to the 3’ region of rubella virus RNA. New Biol. 507
2:255-264. 508
44. Nuss, D. L. 2005. Hypovirulence: mycoviruses at the fungal-plant interface. Nat. Rev. 509
Microbiol. 3:632-642. 510
45. Peremyslov, V.V., Hagiwara, Y. and Dolja, V. V. 1998. Genes required for replication of the 511
15.5-kilobase RNA genome of a plant closterovirus. J. Virol. 72:5870-5876. 512
46. Polashock, J. J. and Hillman, B.I. 1994. A small mitochondrial double-stranded (ds)RNA 513
element associated with a hypovirulenct strain of the chestnut blight fungus and ancestrally 514
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

related to yeast cytoplasmic T and W dsRNAs. Proc. Natl. Aca. Sci. 91:8680-8684. 515
47. Preisig, O., Moleleki, N., Smit, W., Wingfield, B. D. and Wingfield, M. J. 2000. A novel 516
RNA mycovirus in a hypovirulent isolate of the plant pathogen Diaporthe ambigua. J. Gen. 517
Virol. 81:3107-3114. 518
48. Preisig, O., Wingfield, B. D. and Wingfield, M. J. 1998. Coinfection of a fungal pathogen by 519
two distinct double-stranded RNA viruses. Virology 252:399-406. 520
49. Rozanov, M. N., E. V. Koonin, and A. E. Gorbalenya. 1992. Conservation of the putative 521
methyltransferase domain: a hallmark of the ‘Sindbis-like’ supergroup of positive-strand RNA 522
viruses. J. Gen. Virol. 73:2129-2134.[ 523
50. Rott, M. E. and Jelkmann,W. 2005. Little cherry virus-2: Sequence and genomic organization 524
of an unusual member of the Closteroviridae. Arch. Virol. 150:107-123. 525
51. Sambrook, J., Frisch, E. F. and Maniatis, T. 1989. Molecular Cloning: A Laboratory Manual, 526
second ed. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY. 527
52. Sasaki, A., Kanematsu, S., Onoue, M., Oikawa, Y., Nakamura, H. and Yoshida, K. 2007. 528
Artificial infection of Rosellinia necatrix with purified viral particles of a member of the genus 529
Mycoreovirus reveals its uneven distribution in single colonies. Phytopathology 97:278-286. 530
53. Satyanarayana, T., Gowda, S., Boyko, V.P., Albiach-Marti, M.R., Mawassi, M., 531
Navas-Castillo, J., Karasev, A.V., Dolja, V., Hilf, M.E., Lewandowski, D.J., Moreno, P., 532
Bar-Joseph, M., Garnsey, S.M. and Dawson, W.O. 1999. An engineered closterovirus RNA 533
replicon and analysis of heterologous terminal sequences for replication. Proc. Natl. Acad. Sci. 534
U.S.A. 96:7433-7438. 535
54. Schmidt, H.A., K. Strimmer, M. Vingron, and A. von Haeseler. 2002. TREE-PUZZLE: 536
maximum likelihood phylogenetic analysis using quartets and parallel computing. 537
Bioinformatics. 18:502-504. 538
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

55. Segers, G. C., Zhang, X. M., Deng, F. Y., Sun, Q. H. and Nuss, D. L. 2007. Evidence that 539
RNA silencing functions as an antiviral defense mechanism in fungi. PNAS 104:12902-12906. 540
56. Sheng, Q. B., Gong, Y., Xiuhua, Z. and Po, T. 2000. Complete nucleotide sequence of 541
attenuated tomato mosaic virus K genome. Patent: China (00100211.2); CAS, Institute of 542
Microbiology, Beijing, China. 543
57. Sonoda, H., Abe, M., Sugimoto, T., Sato, Y., Bando, M., Fukui, E., Mizuo, H., Takahashi, 544
M., Nishizawa, T., and Okamoto, H. 2004. Prevalence of Hepatitis E Virus (HEV) Infection 545
in Wild Boars and Deer and Genetic Identification of a Genotype 3 HEV from a Boar in Japan. 546
J. Clin. Microbiol. 42:5371-5374. 547
58. Strauss, J. H., Kuhn, R. J., Niesters, H. G. M. and Strauss, E. G. 1990. Functions of the 548
5'-terminal and 3'-terminal sequences of the Sindbis virus genome in replication. New Aspects 549
of Positive-Strand RNA Viruses. MA Brinton and FX Heinz. Washington, D.C., American 550
Society of Microbiology, 61-66. 551
59. Swofford, D. L. 2002. PAUP*: Phylogenetic Analysis Using Parsimony and Other Methods, 552
Version 4.0. Sinauer Associates, Sunderland, Massachusetts. 553
60. Takahashi, K., Kang, J. H., Ohnishi, S., Hino, K. and Mishiro, S. 2002. Genetic 554
heterogeneity of Hepatitis E virus recovered from Japanese patients with acute sporadic 555
hepatitis. J. Infect. Dis. 185:1342-1345. 556
61. Tan, S. H., Nishiguchi, M., Murata, M. and Motoyoshi, F. 2000. The genome structure of 557
kyuri green mottle mosaic tobamovirus and its comparison with that of cucumber green mottle 558
mosaic Tobamovirus. Arch. Virol. 145:1067-1079. 559
62. Tzanetakis, I. E., Postman, J. D. and Martin, R. R. 2005. Characterization of a novel 560
member of the Closteroviridae from Mentha. Phytopathology 95:1043-1048. 561
63. Velasco, L., Janssen, D., Ruiz-Garcia, L., Segundo, E. and Cuadrado, I.M. 2002. The 562
complete nucleotide sequence and development of a differential detection assay for a pepper 563
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

mild mottle virus (PMMoV) isolate that overcomes L3 resistance in pepper. J. Virol. Methods 564
106:135-140. 565
64. Yi, F., Zhang, J., Yu, H., Liu, C., Wang, J. and Hu, Y. 2005. Isolation and identification of a 566
new tetravirus from Dendrolimus punctatus larvae collected from Yunnan Province, China. J. 567
Gen. Virol. 86:789-796. 568
65. Yu,H.J., Lim,D. and Lee,H.S. 2003. Characterization of a novel single-stranded RNA 569
mycovirus in pleurotus ostreatus. Virology 314:9-15. 570
66. Wu, M. D., Zhang, L., Li, G. Q., Jiang, D. H., Hou, A. S. and Huang, H. C. 2007. 571
Hypovirulence and double-stranded RNA in Botrytis cinerea. Phytopathology 97:1590-1599. 572
67. Xie, J., Wei, D., Jiang, D., Fu, Y., Li, G., Ghabrial, S. and Peng, Y. 2006. Characterization of 573
debilitation-associated mycovirus infecting the plant-pathogenic fungus Sclerotinia 574
sclerotiorum. J. Gen. Virol. 87:241-249. 575
68. Zhang, Z. C., Lei, C. Y., Zhang, L. F., Yang, X. X., Chen, R. and Zhang, D. S. 2007. The 576
complete nucleotide sequence of a novel Tobamovirus, Rehmannia mosaic virus. Arch. Virol. 577
153:595-599. 578
69. Zhu, H. Y., Ling, K. S., Goszczynski, D. E., McFerson, J. R. and Gonsalves, D. 1998. 579
Nucleotide sequence and genome organization of grapevine leafroll-associated virus-2 are 580
similar to beet yellows virus, the closterovirus type member. J. Gen. Virol. 79:1289-1298. 581
582
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

Figure Legends 583
Figure 1. Schematic representation of the strategy used in cDNA cloning of SsRV-L dsRNA and 584
predicted genome organization. A: cDNA clones (A to J; red lines) were synthesized using random 585
hexamer primers and denatured dsRNA as a template. Sequences of the regions of the dsRNA that 586
were not covered by these cDNA clones were obtained from cloned RT-PCR products using 587
sequence-specific primers (designed based on the sequences of these cDNA clones), and clones 588
correspondent to 5’ and 3’ termini were amplified using the method of Lambden et al. (34) (Clones 589
1 to 26; black lines). B: Diagrammatic representation of the genomic organization of SsRV-L 590
dsRNA showing the presence of a single ORF. The ORF encodes a putative protein containing a 591
methyltransferase domain, a helicase domain typical of superfamily 1 of viral RNA helicases and 8 592
conserved motifs characteristic of RdRps of positive-strand RNA viruses. 593
594
Figure 2. Amino acid sequence alignment of the putative methyltransferase (A), helicase (B) and 595
RdRp (C) motifs of SsRV-L and those of selected viruses in the genus of Hepevirus. The positions 596
of the conserved motifs in these motifs (shaded areas) correspond to those previously described (28, 597
31, 49) and are indicated with horizontal lines above the shaded areas. Asterisks indicate identical 598
amino acid residues, and colons indicate similar residues. Numbers in brackets refer to the amino 599
acid position in the ORF. See Table 1 for abbreviations of virus names and viral protein accession 600
numbers. 601
602
Figure 3. Phylogenetic analysis of the conserved motifs and flanking sequences of RdRp (A) and 603
viral RNA helicase (B) derived from aligned deduced amino acid sequences of SsRV-L and selected 604
viruses. Neighbor-joining (NJ) algorithm and Maximum likelihood (ML) were used to generate 605
tentative phylogenetic trees independently. NJ algorithm was performed using PAUP* 4.0b10 (59) 606
and ML was performed using program TREE-PUZZLE version 5.2 (54). The resulting ML-tree is 607
shown. The first number indicated at the nodes represents the bootstrap values (%) calculated from 608
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

the NJ tree inferred from 1000 bootstrap replicates and the second number represents the quartet 609
puzzling support values (%) inferred from 10000 puzzling steps; a minus sign (–) indicates that a 610
node is absent in the corresponding NJ method. Bootstrap or quartet puzzling support values of only 611
>50% are indicated. The scale relates branch lengths to the number of substitutions per site. The 612
tree was outgroup-rooted to the viruses in carmo-like group for RdRp analysis and to the arteri-like 613
viruses (IBV) for viral RNA helicase analysis. See Table 1 for abbreviations of virus names, viral 614
protein accession numbers and the amino acid positions in the replicase sequences of the viruses 615
used for phylogenetic analysis. 616
617
Figure 4 Northern hybridization analysis of Sclerotinia sclerotiorum strains singly or doubly 618
infected with Sclerotinia sclerotiorum RNA virus L (SsRV-L). Five strains of Sclerotinia 619
sclerotiorum were used: strain Ep-1PN is doubly infected with SsRV-L and SsDRV; strains 620
Ep-1PNSA-8, Ep-1PNSA-23 and Ep-1PNSA-34 are single-sclerotium-isolates derived from strain 621
Ep-1PN and contain only SsRV-L; and strain Ep-1PNA367, single-ascospore-isolate derived from 622
strain Ep-1PN and is virus-free,. Panel A, electrophoretic analysis of dsRNA samples on agarose 623
gels; Panel B, dsRNA samples were hybridized with an α-32
P labeled cDNA probe of SsRV-L; Panel 624
C, total RNA samples were reversely transcribed with MLV-reverse transcriptase and specific RT 625
primer (5’-CAGTCCCTAGTTT CATCTCGTTCC-3’) designed based on the sequence of SsDRV, 626
and PCR-amplified with SsDRV specific primers (Reverse 627
primer:5’-CAGTCCCTAGTTTCATCTCGTTCC-3’ and Forward Primer 628
5’-TGCAGGAAACAGTCATGGCAAC-3’) . 629
630
Figure 5 Effect of Sclerotinia sclerotiorum RNA virus L (SsRV-L) on hyphal growth and virulence 631
of its host. Five strains of Sclerotinia sclerotiorum were used: strain Ep-1PN is doubly infected with 632
SsRV-L and SsDRV; strains Ep-1PNSA-8, Ep-1PNSA-23 and Ep-1PNSA-34 are 633
single-sclerotium-isolates derived from strain Ep-1PN and contain only SsRV-L; and strain 634
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

Ep-1PNA367, single-ascospore-isolate derived from strain Ep-1PN and is virus-free. Panel A,635
Comparison among five selected strains of S. sclerotiorum for their mycelial growth rate on PDA 636
plate at 20 oC; Panel B, Comparison among the five strains for their virulence on detached leaves 637
of rapeseed (Brassica napus) as determined by induced lesion diameter at 20 oC for 48 h. 638
639
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from
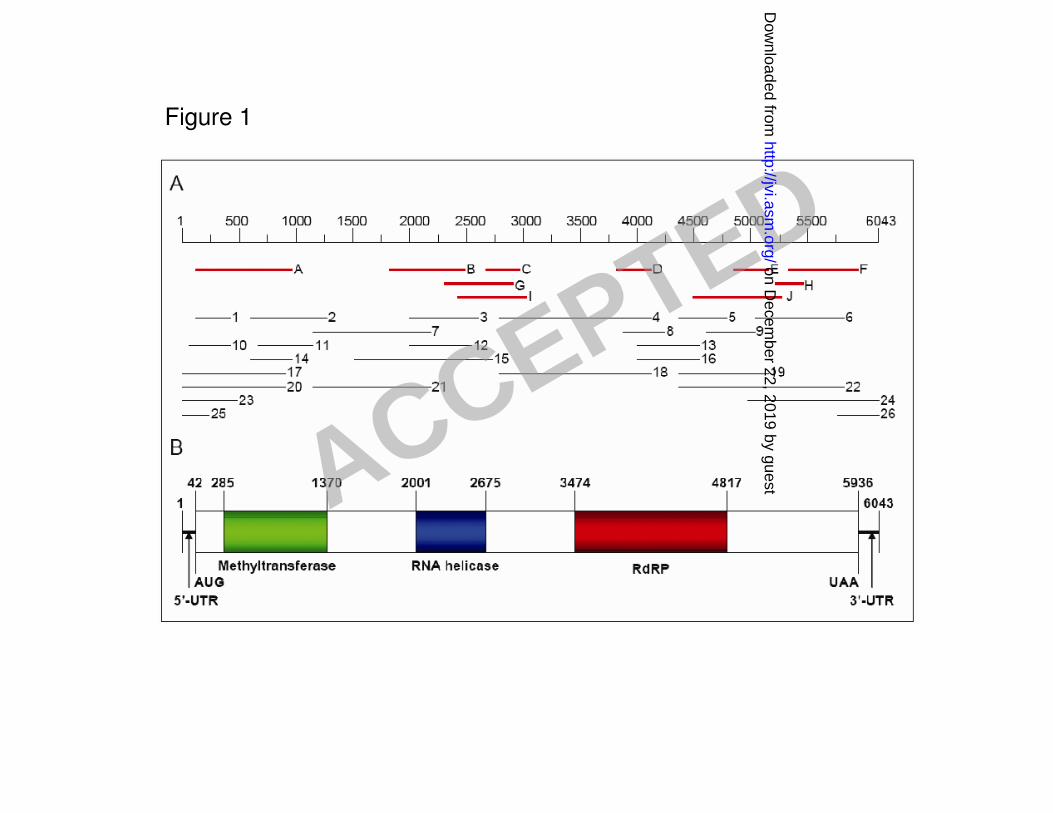
Figure 1
ACCEPTED on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

Figure 2
ACCEPTED on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from
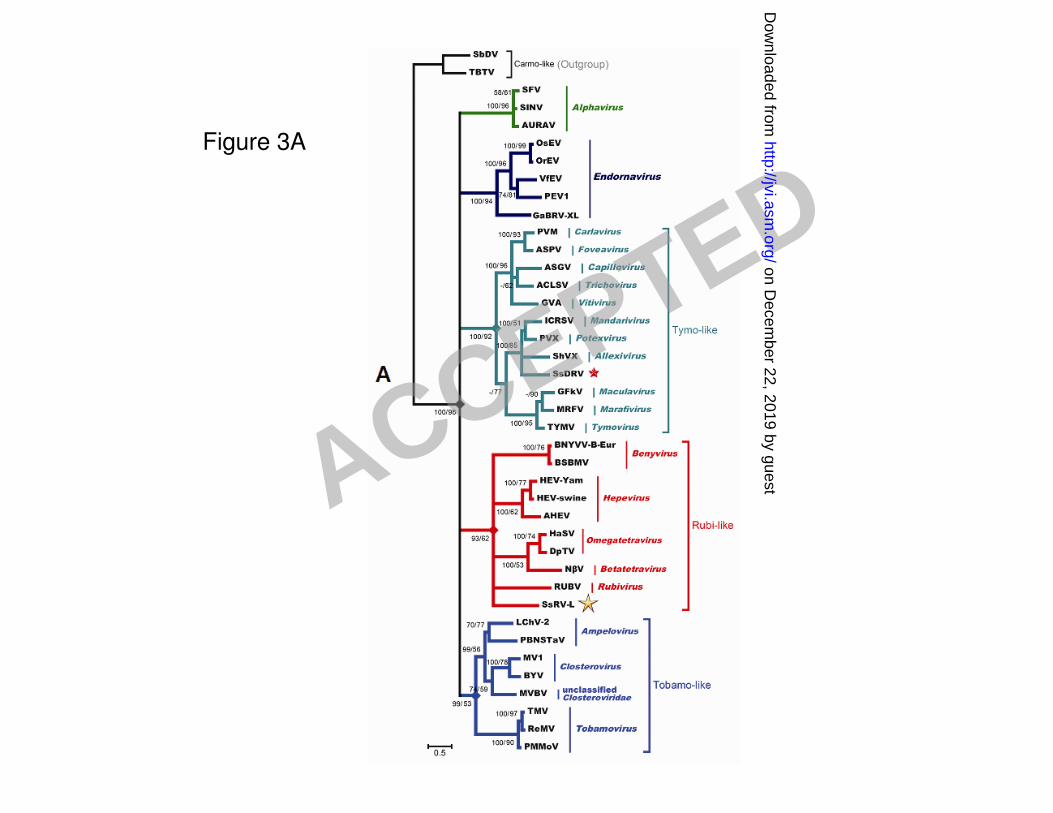
Figure 3A
ACCEPTED on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from
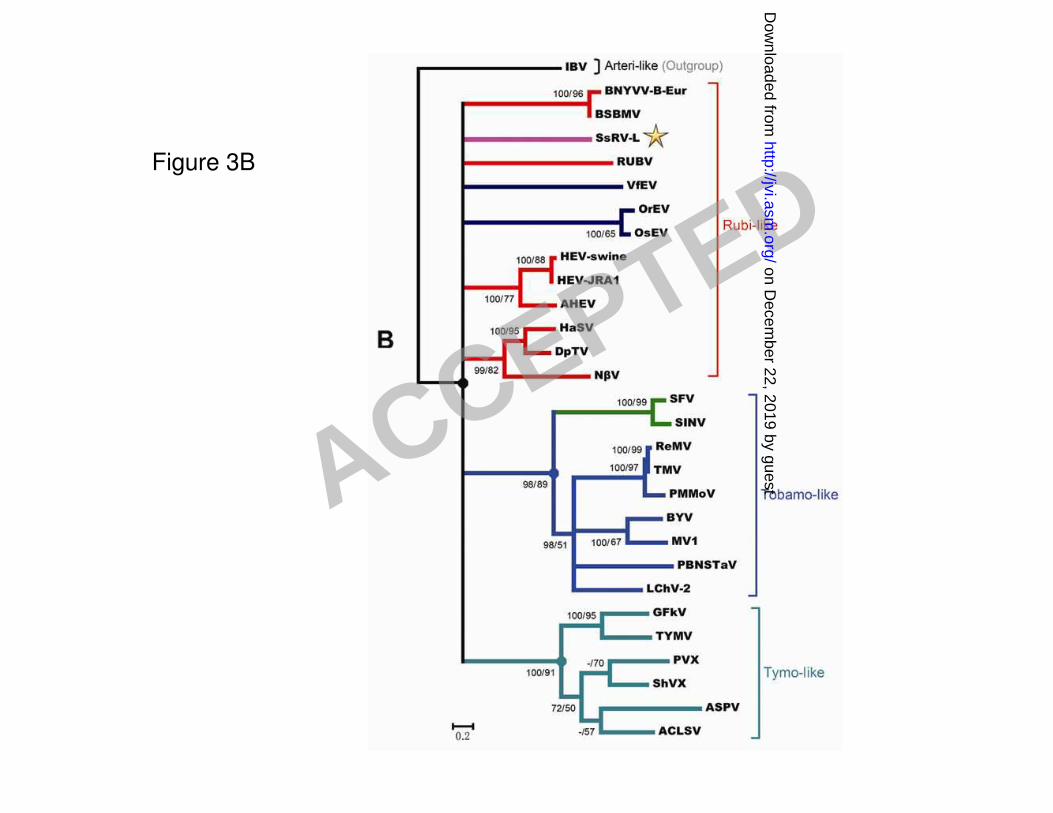
Figure 3B
ACCEPTED on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from
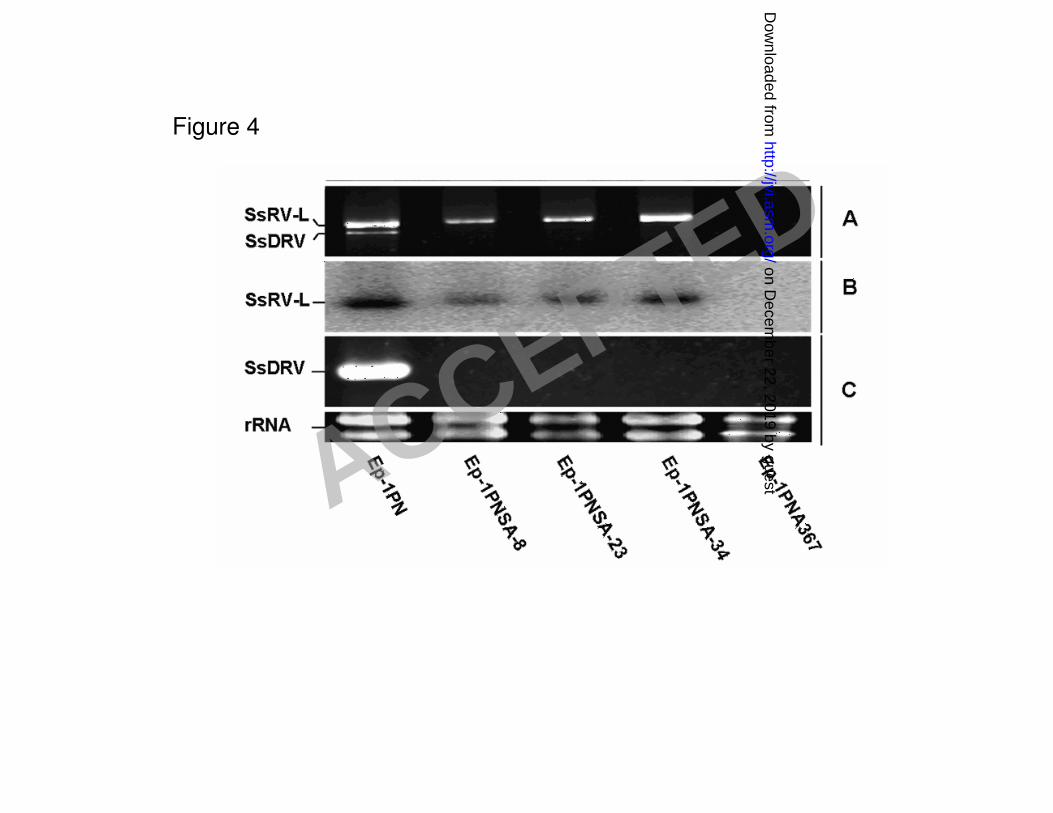
Figure 4
ACCEPTED on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from
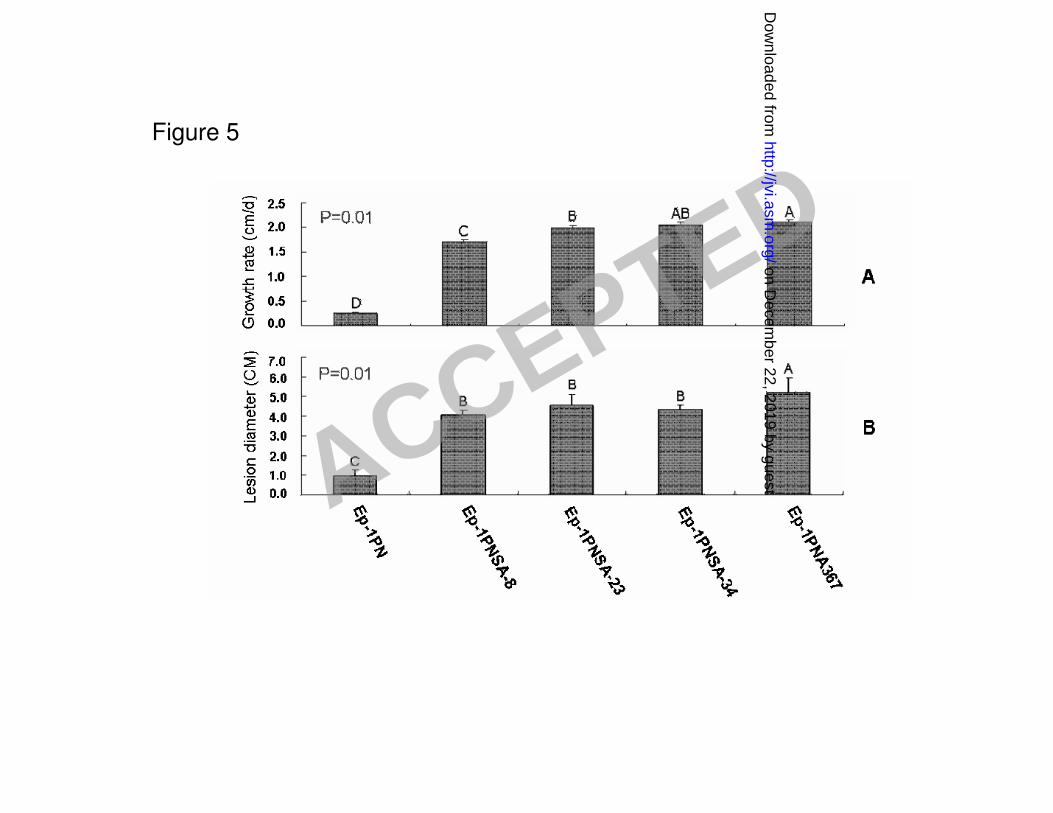
Figure 5
ACCEPTED on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from
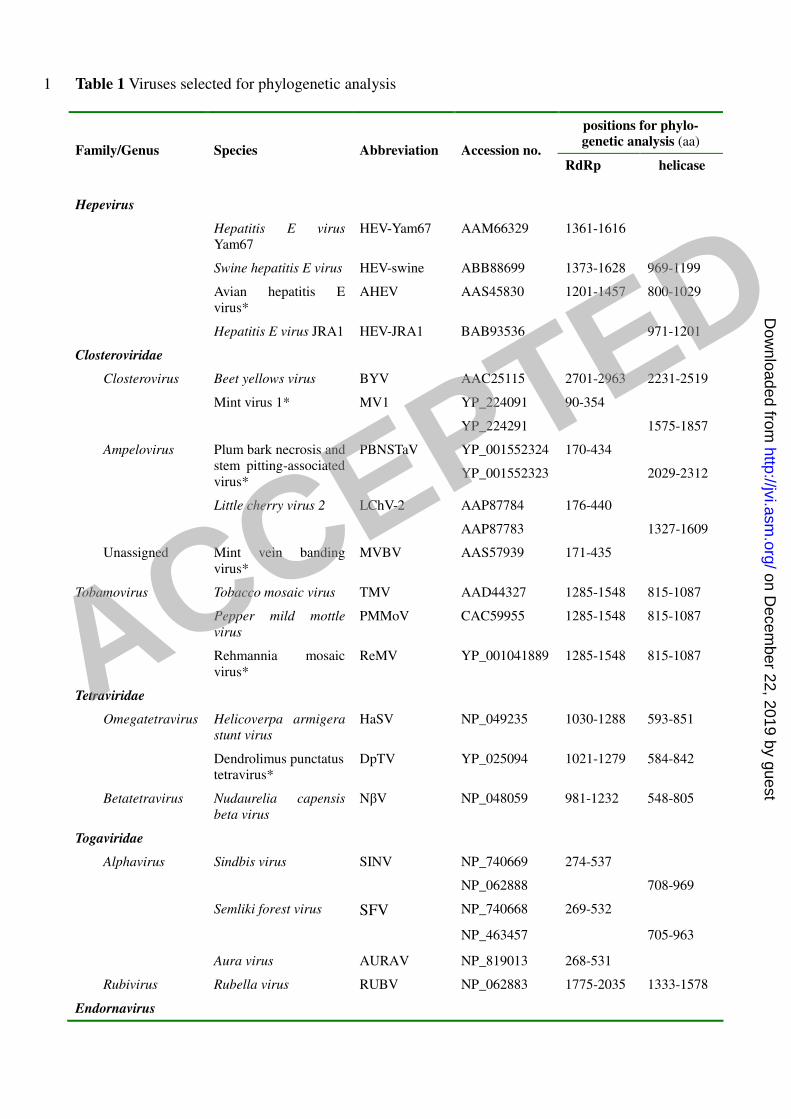
Table 1 Viruses selected for phylogenetic analysis 1
positions for phylo-
genetic analysis (aa) Family/Genus Species Abbreviation Accession no.
RdRp helicase
Hepevirus
Hepatitis E virus
Yam67
HEV-Yam67 AAM66329 1361-1616
Swine hepatitis E virus HEV-swine ABB88699 1373-1628 969-1199
Avian hepatitis E
virus*
AHEV AAS45830 1201-1457 800-1029
Hepatitis E virus JRA1 HEV-JRA1 BAB93536 971-1201
Closteroviridae
Closterovirus Beet yellows virus BYV AAC25115 2701-2963 2231-2519
YP_224091 90-354 Mint virus 1* MV1
YP_224291 1575-1857
YP_001552324 170-434 Ampelovirus Plum bark necrosis and
stem pitting-associated
virus*
PBNSTaV
YP_001552323 2029-2312
AAP87784 176-440 Little cherry virus 2 LChV-2
AAP87783 1327-1609
Unassigned Mint vein banding
virus*
MVBV AAS57939 171-435
Tobamovirus Tobacco mosaic virus TMV AAD44327 1285-1548 815-1087
Pepper mild mottle
virus
PMMoV CAC59955 1285-1548 815-1087
Rehmannia mosaic
virus*
ReMV YP_001041889 1285-1548 815-1087
Tetraviridae
Omegatetravirus Helicoverpa armigera
stunt virus
HaSV NP_049235 1030-1288 593-851
Dendrolimus punctatus
tetravirus*
DpTV YP_025094 1021-1279 584-842
Betatetravirus Nudaurelia capensis
beta virus
NβV NP_048059 981-1232 548-805
Togaviridae
Alphavirus Sindbis virus SINV NP_740669 274-537
NP_062888 708-969
Semliki forest virus SFV NP_740668 269-532
NP_463457 705-963
Aura virus AURAV NP_819013 268-531
Rubivirus Rubella virus RUBV NP_062883 1775-2035 1333-1578
Endornavirus
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from
Oryza sativa
endornavirus
OsEV YP_438200 4219-4482 793-1055
Oryza rufipogon
endornavirus
OrEV YP_438202 4275-4537 1548-1811
Vicia faba
endornavirus
VfEV YP_438201 5375-5637 1947-2212
Phytophthora
endornavirus 1*
PEV1 YP_241110 4259-4522
Gremmeniella abietina
type B RNA virus XL*
GaBRV-XL YP_529670 3052-3308
Tymoviridae
Tymovirus Turnip yellow mosaic
virus
TYMV NP_663297 1478-1729 958-1207
Marafivirus maize rayado fino
virus MRFV NP_734076 218-468
Maculavirus Grapevine fleck virus GFkV NP_542612 1588-1837 1050-1279
Flexiviridae
Potexvirus Potato virus X PVX NP_040882 1140-1397 717-965
Allexivirus Shallot virus X ShVX NP_620648 1319-1569 910-1144
Capillovirus Apple stem grooving
virus
ASGV NP_044335 1262-1517
Carlavirus Potato virus M PVM NP_705707 183-436
Foveavirus Apple stem pitting
virus
ASPV NP_604464 1867-2117 1356-1635
Mandarivirus Indian citrus ringspot
virus
ICRSV NP_203553 1341-1598
Trichovirus Apple chlorotic leaf
spot virus
ACLSV NP_040551 1539-1789 1040-1307
Vitivirus Grapevine virus A GVA NP_619662 1375-1625
Luteoviridae
Luteovirus Soybean dwarf virus SbDV NP_150431 484-757
Umbravirus
Tobacco bushy top
virus TBTV NP_733848 124-401
Benyvirus
Beet necrotic yellow
vein virus-F2, France
BNYVV-B-Eur CAA28795 1737-1997 927-1182
Beet soil-borne mosaic
virus BSBMV NP_612601 1745-2005 929-1184
Coronaviridae
Coronavirus Infectious bronchitis
virus
IBV NP_066134 5132-5447
Unassigned Mycovirus
Sclerotinia
sclerotiorum
SsDRV YP_325662 1273-1530
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

debilitation-associated
RNA virus
Sclerotinia
sclerotiorum RNA
virus-L
SsRV-L ACE88957 1275-1530 640-878
2
* Tentative members in the designated genera are indicated by asterisks and the names are not 3
italicized 4
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from
5
Table 2: Percent amino acid sequence identity and similarity between the methyltransferase, 6
helicase and RdRp motifs of SsRV-L and those of other selected alphavirus-like positive-strand 7
RNA viruses. 8
Genus Virus
Abbreviat
ion Accession no. Identity Similarity
Refer
ence
Hepevirus Avian hepatitis E virus* AHEV AAS45830 63/241 (26%) 93/241 (38%) 23
Tobamovirus Tomato mosaic virus ToMV CAD10425 57/240 (23%) 97/240 (40%) Un-pub
lished
Tobacco mosaic virus TMV AAD44327 57/240 (23%) 97/240 (40%) 56
Pepper mild mottle virus PMMoV CAC59955 56/237 (23%) 98/237 (41%) 63
Cucumber green mottle
mosaic virus CGMMV BAA87620 54/217 (24%) 92/217 (42%) 61
Rehmannia mosaic virus* ReMV YP_001041889 56/240 (23%) 95/240 (39%) 68
Meth
yltra
nsfera
se
Betatetravir
us
Nudaurelia capensis beta
virus NβV NP_048059 63/256 (24%) 99/256 (38%) 16
Hepevirus Avian hepatitis E virus* AHEV AAS45830 67/228 (29%) 94/228 (41%) 23
Hepatitis E virus JRA1 HEV-JRA
1 BAB93536 66/229 (28%) 96/229 (41%) 60
Swine hepatitis E virus HEV-
swine ABB88699 82/284 (28%) 122/284 (42%)
Un-pub
lished
Omegatetrav
irus
Helicoverpa armigera
stunt virus HaSV NP_049235 68/240 (28%) 104/240 (43%) 15
RN
A h
elicase
Dendrolimus punctatus
tetravirus* DpTV YP_025094 66/241 (27%) 106/241 (43%) 64
Hepevirus Hepatitis E virus Yam67 HEV-Yam
67 AAM66329 68/250 (27%) 108/250 (43%) 24
Swine hepatitis E virus HEV-
swine ABB88699 70/245 (28%) 108/245 (44%)
Un-pub
lished
Closteroviru
s Beet yellows virus BYV AAC25115 79/335 (23%) 130/335 (38%) 45
Mint virus 1* MV1 YP_224091 65/276 (23%) 111/276 (40%) 62
Grapevine
leafroll-associated virus 2 GLRAV-2 AAC40856 65/280 (23%) 110/280 (39%) 69
Rd
RP
Grapevine rootstock stem GRSLAV NP_835337 66/280 (23%) 112/280 (40%) Un-pub
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from
lesion associated virus* lished
Citrus tristeza virus CTV AAO12717 70/284 (24%) 112/284 (39%) 53
Unassigned
Closteroviri
dae
Mint vein banding virus* MVBV AAS57939 92/384 (23%) 152/384 (39%) 62
Ampelovirus Plum bark necrosis and
stem pitting-associated
virus*
PBNSTaV YP_001552324 84/346 (24%) 144/346 (41%) 2
Little cherry virus 2 LChV-2 AAP87784 75/308 (24%) 124/308 (40%) 50
9
* Tentaive members in the designated genera are indicated by asterisks and the names are not 10
italicized 11
The number of identical or similar amino acid residues/total number of residues included in the 12
sequence comparison analysis is shown. Values in parentheses are the identity or similarity scores 13
as calculated by the BLAST program 14
ACCEPTED
on Decem
ber 22, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from





























