Embed Size (px)
Citation preview

Virus Research 137 (2008) 157–162
Contents lists available at ScienceDirect
Virus Research
journa l homepage: www.e lsev ier .com/ locate /v i rusres
Short communication
Emerging G9 rotavirus strains in the northwest of China
Mustafizur Rahmana,c,∗, Xue-lei Yangb, He Sunb, Khandaker Mahzebind,Natasha Willemina Verstappena, Luís Novoa, Jelle Matthijnssensa, Marc Van Ransta
a Laboratory of Clinical and Epidemiological Virology, Rega Institute, University of Leuven, B-3000, Leuven, Belgiumb Xinjiang Pediatric Institute, People’s Hospital of Xinjiang, 91 Tianchi Road, Urumqi, People’s Republic of Chinac International Centre for Diarrhoeal Disease Research, Bangladesh (ICDDR,B), Mohakhali, Dhaka-1212, Bangladeshd Stamford University Bangladesh, 51, Siddeswari Road, Dhaka-1217, Bangladesh
a r t i c l e i n f o
Article history:Received 21 March 2008Received in revised form 24 June 2008Accepted 3 July 2008
Keywords:Group A rotavirusG9Reassortment
a b s t r a c t
Although G9 rotaviruses have become one of the important rotavirus genotypes worldwide, they havebeen uncommon in China. Recently, we reported G9 rotaviruses as a highly prevalent genotype in Xinjiang,the northwest part of China [Yang, X., Matthijnssens, J., Sun, H., Muhamaiti, J., Zhang, B., Nahar, S., VanRanst, M., Rahman, M., 2008. Temporal changes of rotavirus strain distribution in a northwest city ofChina, 1996–2005. Int. J. Infect. Dis., June (Epub ahead of print)]. Here we report the genetic variationsof the Xinjiang-G9 rotaviruses isolated between 1999 and 2005. Sequence analysis of the VP7 genes ofXinjiang-G9 strains indicated that they were more closely related to the contemporary global G9 strainsthan to the prototype Chinese G9 strains. However, their VP4 genes were most similar to those from thelocally circulating G1P[8], G2P[4], G3P[6] and G3P[8] strains. This indicates that reassortment rather than
SequencingVP7VP4XC
antigenic drift might be the preferred evolutionary mechanism for the emergence of the G9 rotaviruses inXinjiang. These findings will be of major significance for understanding the emergence of newly introduced
aGbHm2hieat2aC(ctec
C2e(VaAXXsfVm
ne
0d
injianghina
rotavirus strains.
Rotaviruses remain one of the most common diarrhea causinggents in children. Among the 19 G and 27 P genotypes, G1P[8],2P[4], G3P[8] and G4P[8], were the most common in humansefore 1995 (Estes and Kapikian, 2007; Matthijnssens et al., 2008a).owever, genotype G9 and G12 have emerged respectively from theid-1990s and 2000s (Matthijnssens et al., 2008; Rahman et al.,
005, 2007a,b). In addition, several rotavirus genotypes (G5 and G8)ave shown their ability as potential candidates for the next emerg-
ng rotavirus strains (Duan et al., 2007; Esona et al., 2004; Gentscht al., 2005; Hong et al., 2007; Matthijnssens et al., 2006; Santosnd Hoshino, 2005). Although G9 rotaviruses have become one ofhe important rotavirus genotypes in many countries (Bresee et al.,005; Santos et al., 2005), surprisingly, they have not been reporteds a common strain in China. The first G9 strain was detected inhina in 1994 and was isolated sporadically in the following yearsFang et al., 2002; Qian et al., 1994; Zhang et al., 2006). Studies
onducted during 2000–2006 in different cities of China indicatedhat G9 strain was an uncommon strain (Fang et al., 2005; Wangt al., 2007; Yang et al., 2007; Zhang et al., 2006). Nevertheless,onsistent detection of G9 strains was reported outside mainland∗ Corresponding author. Tel.: +32 16 332184; fax: +32 16 332131.E-mail address: [email protected] (M. Rahman).
ucMsnMda
168-1702/$ – see front matter © 2008 Elsevier B.V. All rights reserved.oi:10.1016/j.virusres.2008.07.004
© 2008 Elsevier B.V. All rights reserved.
hina, such as in Taiwan and Hong Kong (Chen et al., 2005; Lai et al.,005; Lin et al., 2006; Lo et al., 2005). We recently reported consid-rably high numbers of G9 rotavirus strains from Xinjiang, ChinaYang et al., 2008). In the present study, we analyzed the VP7 andP4 gene segments of the Xinjiang-G9 strains to reveal their originnd their evolutionary relationships with other rotavirus strains.total of 29 G9 rotaviruses were identified during 1999–2005 in
injiang, China. In addition, several P[4], P[6], and P[8] strains frominjiang were also analyzed to establish their genomic relation-hips with the G9 strains. The stool specimens were transportedrom Xinjiang to the Laboratory of Clinical and Epidemiologicalirology, Rega Institute, University of Leuven in Belgium for furtherolecular characterization.Rotavirus RNA extraction, reverse transcription (RT)-PCR and
ucleotide sequencing were carried out as was described by Yangt al. (2008). The chromatogram sequencing files were inspectedsing Chromas 2.23 (Technelysium, Queensland, Australia), andonsensus sequences were prepared using SeqMan II (DNASTAR,adison, WI). Nucleotide and amino acid sequence similarity
earches were performed using the National Centre for Biotech-ology Information (NCBI, National Institutes of Health, Bethesda,D) BLAST (Basic Local Alignment Search Tool) server on GenBank
atabase, release 161.0 (Altschul et al., 1990). Multiple sequencelignments were calculated using CLUSTALX 1.81 (Thompson et al.,

158 M. Rahman et al. / Virus Research 137 (2008) 157–162
Fig. 1. Phylogenetic dendrogram based on partial VP7 nucleotide sequences (nt 49–761) of the G9 rotavirus strains. Xinjiang-G9 strains are shaded; other Chinese G9 strainsare in bold. The numbers adjacent to the nodes represent the percentage of bootstrap support (of 1000 replicates) for the clusters to the right of the node. Bootstrap valueslower than 75% are not shown. Hu, human; Po, porcine. Accession numbers are available upon request. BEL, Belgium; BRA, Brazil; CAM, Cameroon; CHA, China; GHA, Ghana;IND, India; IRE, Ireland; JAP, Japan; MAU, Mauritius; NIG, Nigeria; SLO, Slovenia; THA, Thailand; USA, the United States.

M. Rahman et al. / Virus Research 137 (2008) 157–162 159
F –845a tic ReT
12y(turN
ME
ig. 2. Phylogenetic dendrogram based on partial VP4 nucleotide sequences (nt 85nother Chinese strain are in bold. AUS, Australia; BAN, Bangladesh; DRC, Democraaiwan; VIE, Vietnam.
997). Sequences were manually edited in the GeneDoc Version.6.002 alignment editor (Nicholas et al., 1997). Phylogenetic anal-ses were conducted using the MEGA Version 4 software package
Tamura et al., 2007). The dendrograms were constructed usinghe neighbor-joining method. Genetic distances were calculatedsing the Kimura-2 parameter model. The nucleotide sequenceseported in this paper were submitted to GenBank using theational Center for Biotechnology Information (NCBI, Bethesda,XGgis
) of the P[6] strains. Xinjiang-P[6] strains are shaded; Xinjiang-G9P[6] strains andpublic of Congo; ITA, Italy; KOR, South Korea, MAL, Malawi; SA, South Africa; TAI,
D) Sequin, Version 7.30 under accession numbers EU144080–U144118.
The partial VP7 deduced amino acid sequences (aa 1-237) of all
injiang-G9 strains were compared to other G9 strains available inenBank. Similarity searches indicated that the Xinjiang-G9 VP7enes were very similar to the cotemporary G9 strains circulat-ng worldwide. A dendrogram was constructed which included G9trains from different countries available in GenBank (Fig. 1). The
1 Research 137 (2008) 157–162
pssiXss9IIimCGscbGRhssh
PtstwritastVwatXcK
F7o
nXXp
60 M. Rahman et al. / Virus
hylogenetic tree confirmed the three established lineages withome sub-lineages of G9 strains described earlier. All Xinjiang-G9trains, regardless of P genotype specificities, clustered togethern lineage III with the recently emerging G9 strains. Although theinjiang-G9 strains were closely related to a recent Chinese G9train L169, they were distantly related to another Chinese G9train 97′SZ37 isolated in 1997. It can be noted that the strain7′SZ37 had only 89% identity with Xinjiang-G9 strains (lineageII) and 91% identity with the prototype strains of lineage I andI, and most likely, belonged to a different lineage. The cluster-ng of Xinjiang-G9P[8] strains was quite interesting. They were
ore closely related to the US and African G9 strains (IN826,C149 and MR4723/00) rather than to the Xinjiang-G9P[6] or -9P[4] strains. This indicates that multiple introductions of G9trains might have taken place in this region. G9 strains in mostountries first appeared as G9P[6] and converted to G9P[8] toecome more prevalent strain, which was also seen for Xinjiang-9 strains (Iturriza-Gomara et al., 2000; Matthijnssens et al., 2008;ahman et al., 2005). Since P[8] genotype is the most commonuman rotavirus strains covering around 80% of the total rotavirustrains all over the world, it is possible that acquirement of P[8]pecificity is necessary for rotavirus strains to better fit in humanosts.
The Xinjiang-G9 strains were found in combination with threegenotypes: P[4], P[6] and P[8]. The initial Xinjiang-G9 strain con-
ained a P[6] specificity. A dendrogram based on the partial VP4equences of P[6] strains from different countries revealed thathe Xinjiang-G9P[6] strains clustered in a monophyletic branchith local G3P[6] strains (Fig. 2). In contrast, they were distantly
elated to other Chinese P[6] strains LL4260 and LL36755, isolatedn 2001 and 2003 respectively in Hebei province. These genetic rela-ionships indicate that the VP4 donors of Xinjiang-G9P[6] strainsre most likely the G3P[6] strains circulating in Xinjiang. Six G9trains with P[4] specificity were identified in 2005. Among themhree were found to be mixed with P[6]. Nucleotide sequence ofP4 gene of Xinjiang-G9P[4] strain XJ05-71 was 100% identicalith the locally circulating G2P[4] strains. Moreover, phylogenetic
nalysis of VP4 gene segment of strain XJ05-71 showed that it clus-ered closely with the local G2P[4] strains XJ05-20, XJ05-101, andJ04-647 (Fig. 3). Although the Xinjiang-G9P[4] strain shared theommon VP4 lineage with the G2P[4] strains from Taiwan, Southorea, and Japan, the strain was distantly related to another Chi-
sm(tJ
ig. 3. Phylogenetic dendrogram based on partial VP4 nucleotide sequences (nt9–828) of the P[4] strains. Xinjiang-P[4] strains are shaded; Xinjiang-G9P[4] andther Chinese strain are in bold. PHI, The Philippines.
ese G2P[4] strain TB-Chen. Therefore, it can be speculated that theinjiang-G9P[4] originated through a reassortment event betweeninjiang-G2P[4] and -G9P[6] strain. The G9P[8] became highlyrevalent in Xinjiang in 2005. Sequence analysis of the VP4 gene
egments of the Xinjiang-G9P[8] strains showed that they wereost closely related to the VP4 genes of the Xinjiang-G3P[8] strainsFig. 4). Nonetheless, a rather close relationship was also found withhe P[8] strains from neighbouring countries, such as South Korea,apan, Taiwan, Thailand, Bangladesh and India.
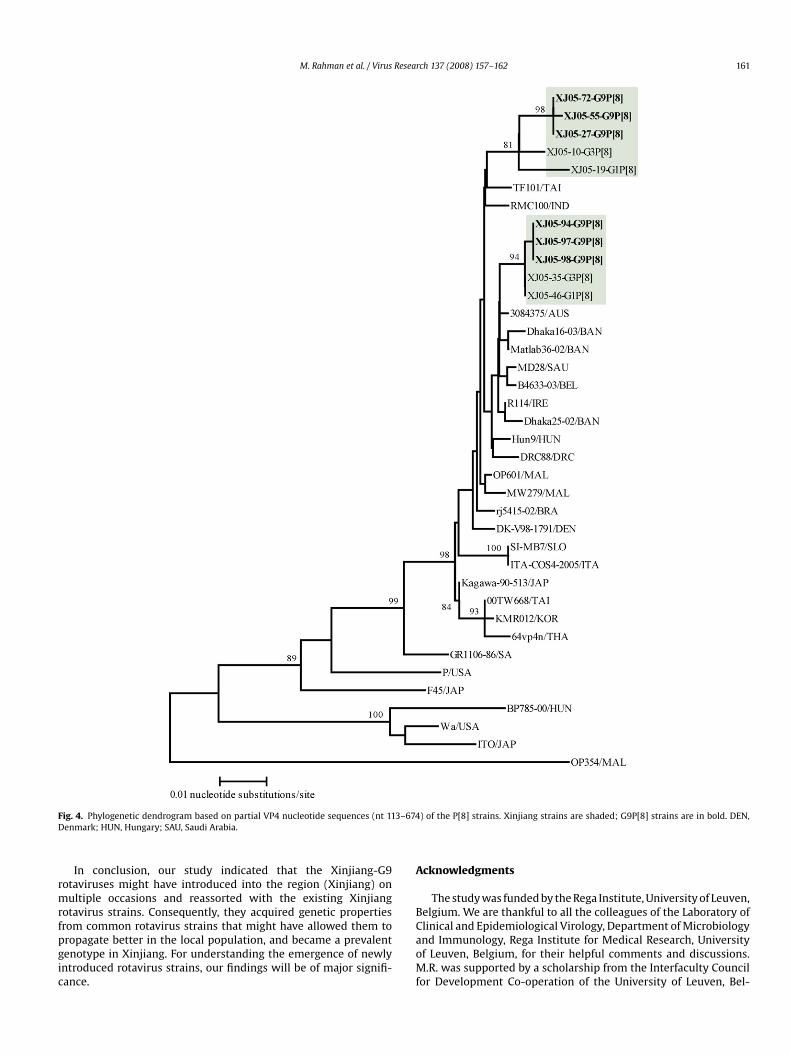
M. Rahman et al. / Virus Research 137 (2008) 157–162 161
F 3–674D
rmrfpgic
A
B
ig. 4. Phylogenetic dendrogram based on partial VP4 nucleotide sequences (nt 11enmark; HUN, Hungary; SAU, Saudi Arabia.
In conclusion, our study indicated that the Xinjiang-G9otaviruses might have introduced into the region (Xinjiang) onultiple occasions and reassorted with the existing Xinjiang
otavirus strains. Consequently, they acquired genetic properties
rom common rotavirus strains that might have allowed them toropagate better in the local population, and became a prevalentenotype in Xinjiang. For understanding the emergence of newlyntroduced rotavirus strains, our findings will be of major signifi-ance.CaoMf
) of the P[8] strains. Xinjiang strains are shaded; G9P[8] strains are in bold. DEN,
cknowledgments
The study was funded by the Rega Institute, University of Leuven,elgium. We are thankful to all the colleagues of the Laboratory of
linical and Epidemiological Virology, Department of Microbiologynd Immunology, Rega Institute for Medical Research, Universityf Leuven, Belgium, for their helpful comments and discussions..R. was supported by a scholarship from the Interfaculty Councilor Development Co-operation of the University of Leuven, Bel-

1 Resea
gooV
R
A
B
C
D
E
E
F
F
G
H
I
L
L
L
M
M
M
N
Q
R
R
R
S
S
T
T
W
Y
Y
62 M. Rahman et al. / Virus
ium. XL.Y. was funded by the Government of the People’s Republicf China. J.M. was supported by the Institute for the Promotionf Innovation through Science and Technology in Flanders (IWTlaanderen).
eferences
ltschul, S.F., Gish, W., Miller, W., Myers, E.W., Lipman, D.J., 1990. Basic local align-ment search tool. J. Mol. Biol. 215 (3), 403–410.
resee, J.S., Hummelman, E., Nelson, E.A., Glass, R.I., 2005. Rotavirus in Asia: the valueof surveillance for informing decisions about the introduction of new vaccines.J. Infect. Dis. 192 (Suppl. 1), S1–S5.
hen, K.T., Chen, P.Y., Tang, R.B., Huang, Y.F., Lee, P.I., Yang, J.Y., Chen, H.Y., Bresee,J., Hummelman, E., Glass, R., 2005. Sentinel hospital surveillance for rotavirusdiarrhea in Taiwan, 2001–2003. J. Infect. Dis. 192 (Suppl. 1), S44–S48.
uan, Z.J., Li, D.D., Zhang, Q., Liu, N., Huang, C.P., Jiang, X., Jiang, B., Glass, R., Steele,D., Tang, J.Y., Wang, Z.S., Fang, Z.Y., 2007. Novel human rotavirus of genotypeG5P[6] identified in a stool specimen from a Chinese girl with diarrhea. J. Clin.Microbiol. 45 (5), 1614–1617.
sona, M.D., Armah, G.E., Geyer, A., Steele, A.D., 2004. Detection of an unusual humanrotavirus strain with G5P[8] specificity in a Cameroonian child with diarrhea. J.Clin. Microbiol. 42 (1), 441–444.
stes, M.K., Kapikian, A.Z., 2007. Rotaviruses. In: Knipe, D.M., Howley, P.M., Griffin,D.E., Lamb, R.A., Martin, M.A., Roizman, B., Straus, S.E. (Eds.), Fields Virology, vol.2, Fifth ed. Lippincott Williams & Wilkins, Philadelphia, pp. 1917–1974.
ang, Z.Y., Wang, B., Kilgore, P.E., Bresee, J.S., Zhang, L.J., Sun, L.W., Du, Z.Q., Tang,J.Y., Hou, A.C., Shen, H., Song, X.B., Nyambat, B., Hummelman, E., Xu, Z.Y., Glass,R.I., 2005. Sentinel hospital surveillance for rotavirus diarrhea in the People’sRepublic of China, August 2001–July 2003. J. Infect. Dis. 192 (Suppl. 1), S94–S99.
ang, Z.Y., Yang, H., Qi, J., Zhang, J., Sun, L.W., Tang, J.Y., Ma, L., Du, Z.Q., He, A.H., Xie,J.P., Lu, Y.Y., Ji, Z.Z., Zhu, B.Q., Wu, H.Y., Lin, S.E., Xie, H.P., Griffin, D.D., Ivanoff, B.,Glass, R.I., Gentsch, J.R., 2002. Diversity of rotavirus strains among children withacute diarrhea in China: 1998–2000 surveillance study. J. Clin. Microbiol. 40 (5),1875–1878.
entsch, J.R., Laird, A.R., Bielfelt, B., Griffin, D.D., Banyai, K., Ramachandran, M., Jain,V., Cunliffe, N.A., Nakagomi, O., Kirkwood, C.D., Fischer, T.K., Parashar, U.D., Bre-see, J.S., Jiang, B., Glass, R.I., 2005. Serotype diversity and reassortment betweenhuman and animal rotavirus strains: implications for rotavirus vaccine pro-grams. J. Infect. Dis. 192 (Suppl. 1), S146–S159.
ong, S.K., Lee, S.G., Lee, S.A., Kang, J.H., Lee, J.H., Kim, J.H., Kim, D.S., Kim, H.M., Jang,Y.T., Ma, S.H., Kim, S.Y., Paik, S.Y., 2007. Characterization of a G11,P[4] strain ofhuman rotavirus isolated in South Korea. J. Clin. Microbiol. 45 (11), 3759–3761.
turriza-Gomara, M., Cubitt, D., Steele, D., Green, J., Brown, D., Kang, G., Desselberger,U., Gray, J., 2000. Characterisation of rotavirus G9 strains isolated in the UKbetween 1995 and 1998. J. Med. Virol. 61 (4), 510–517.
ai, H.C., Lin, S.J., Lin, H.R., Ku, C.S., Wang, L., Yang, C.C., 2005. Phylogenetic analysesof human rotavirus in central Taiwan in 1996, 2001 and 2002. J. Clin. Virol. 32(3), 199–217.
in, Y.P., Chang, S.Y., Kao, C.L., Huang, L.M., Chung, M.Y., Yang, J.Y., Chen, H.Y.,Taniguchi, K., Tsai, K.S., Lee, C.N., 2006. Molecular epidemiology of G9 rotavirusesin Taiwan between 2000 and 2002. J. Clin. Microbiol. 44 (10), 3686–3694.
o, J.Y., Szeto, K.C., Tsang, D.N., Leung, K.H., Lim, W.W., 2005. Changing epidemiol-ogy of rotavirus G-types circulating in Hong Kong, China. J. Med. Virol. 75 (1),170–173.
Z
rch 137 (2008) 157–162
atthijnssens, J., Ciarlet, M., Heiman, E., Arijs, I., Delbeke, T., McDonald, S.M.,Palombo, E.A., Iturriza-Gomara, M., Maes, P., Patton, J.T., Rahman, M., Van Ranst,M., 2008a. Full genome-based classification of rotaviruses reveals common ori-gin between human Wa-like and porcine rotavirus strains and human DS-1-likeand bovine rotavirus strains. J. Virol. 82 (7), 3204–3219.
atthijnssens, J., Rahman, M., Ciarlet, M., Van Ranst, M. 2008, Emerging humanrotavirus genotypes. In: A.E. Palombo (Ed.), Viruses in the Environment. ResearchSignpost, Trivandrum, India.
atthijnssens, J., Rahman, M., Yang, X., Delbeke, T., Arijs, I., Kabue, J.P., Muyembe, J.J.,Van Ranst, M., 2006. G8 rotavirus strains isolated in the Democratic Republic ofCongo belong to the DS-1-like genogroup. J. Clin. Microbiol. 44 (5), 1801–1809.
icholas, K.B., Nicholas, H.B., Deerfield, D.W., 1997. GeneDoc: analysis and visualiza-tion of genetic variation. Embnet News 4 (14).
ian, Y., Yuan, L.J., Xiong, C.H., Zhang, Y., Liu, J., Guan, D.H., Wang, Z.L., 1994. Identi-fication of rotavirus G9 type from a stool specimen collected from a child withdiarrhea in Beijing. Chin. J. Virol. 10 (3), 263–267.
ahman, M., Matthijnssens, J., Goegebuer, T., De Leener, K., Vanderwegen, L., van derDonck, I., Van Hoovels, L., De Vos, S., Azim, T., Van Ranst, M., 2005. Predominanceof rotavirus G9 genotype in children hospitalized for rotavirus gastroenteritis inBelgium during 1999–2003. J. Clin. Virol. 33 (1), 1–6.
ahman, M., Matthijnssens, J., Yang, X., Delbeke, T., Arijs, I., Taniguchi, K., Iturriza-Gomara, M., Iftekharuddin, N., Azim, T., Van Ranst, M., 2007a. Evolutionaryhistory and global spread of the emerging G12 human rotaviruses. J. Virol. 81(5), 2382–2390.
ahman, M., Sultana, R., Ahmed, G., Nahar, S., Hassan, Z.M., Saiada, F., Podder, G.,Faruque, A.S., Siddique, A.K., Sack, D.A., Matthijnssens, J., Van Ranst, M., Azim, T.,2007b. Prevalence of G2P[4] and G12P[6] rotavirus, Bangladesh. Emerg. Infect.Dis. 13 (1), 18–24.
antos, N., Hoshino, Y., 2005. Global distribution of rotavirus serotypes/genotypesand its implication for the development and implementation of an effectiverotavirus vaccine. Rev. Med. Virol. 15 (1), 29–56.
antos, N., Volotao, E.M., Soares, C.C., Campos, G.S., Sardi, S.I., Hoshino, Y., 2005.Predominance of rotavirus genotype G9 during the 1999, 2000 and 2002 seasonsamong hospitalized children in the city of Salvador, Bahia, Brazil: implicationsfor future vaccine strategies. J. Clin. Microbiol. 43 (8), 4064–4069.
amura, K., Dudley, J., Nei, M., Kumar, S., 2007. MEGA4: Molecular Evolution-ary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 24,1596–1599.
hompson, J.D., Gibson, T.J., Plewniak, F., Jeanmougin, F., Higgins, D.G., 1997.The CLUSTAL X windows interface: flexible strategies for multiple sequencealignment aided by quality analysis tools. Nucleic Acids Res. 25 (24),4876–4882.
ang, Y.H., Kobayashi, N., Zhou, D.J., Yang, Z.Q., Zhou, X., Peng, J.S., Zhu, Z.R., Zhao, D.F.,Liu, M.Q., Gong, J., 2007. Molecular epidemiologic analysis of group A rotavirusesin adults and children with diarrhea in Wuhan city, China, 2000–2006. Arch.Virol. 152 (4), 669–685.
ang, J., Wang, T., Wang, Y., Lu, B., Bai, X., Zhang, L.M.W., Wang, H., 2007. Emergenceof human rotavirus group A genotype G9 strains, Wuhan, China. Emerg. Infect.Dis. 13 (10), 1587–1589.
ang, X., Matthijnssens, J., Sun, H., Muhamaiti, J., Zhang, B., Nahar, S., Van Ranst,
M., Rahman, M., 2008. Temporal changes of rotavirus strain distribution in anorthwest city of China, 1996–2005. Int. J. Infect. Dis. (June) (Epub ahead ofprint).hang, Q., Wang, D., Ye, X.H., Duan, Z.J., Du, Z.Q., Jin, Y., Tang, J., Sun, L., Xi, J., Steele,D., Glass, R., Fang, Z.Y., 2006. Molecular epidemiology of rotavirus serotype G9isolated from 9 regions in China. Chin. J. Vaccine Immun. 12 (6), 476–479.






![Rotavirus Vaccines: an Overview - myCMEmedia.mycme.com/documents/23/rotavirus_expert_reviews...rotavirus strains bearing VP7 G serotypes G1 to G4 and G9 and VP4 P genotypes P1B[4],](https://img.pdfslide.net/doc/110x75/5f8e21d6883e9848ab510511/rotavirus-vaccines-an-overview-rotavirus-strains-bearing-vp7-g-serotypes.jpg)


![Molecular characterisation of rotavirus strains detected ... · Rotavirus is the most common cause of severe gastroenteritis in children under five years of age [1]. The currently](https://img.pdfslide.net/doc/110x75/5fff0da4b0f8833e00500254/molecular-characterisation-of-rotavirus-strains-detected-rotavirus-is-the-most.jpg)

![Clinical and Molecular Characteristics of Human Rotavirus ...The VP7 gene of rotavirus strain To14-0 shared the highest nucleotide identity with the VP7 genes of human G8P[8] strains](https://img.pdfslide.net/doc/110x75/5f4c0f217df3225bdf392cfa/clinical-and-molecular-characteristics-of-human-rotavirus-the-vp7-gene-of-rotavirus.jpg)