Embed Size (px)
Citation preview

Plant Physiol. (1 995) 108: 1647-1 656
End-Product Control of Carbon Metabolism in Culture-Grown Sugar Beet Plants'
Molecular and Physiological Evidence on Accelerated Leaf Development and Enhanced Gene Expression
Yelena Kovtun and jaleh Daie*
329 Birge Hall, 430 Lincoln Drive, University of Wisconsin, Madison, Wisconsin 53706-1 381
Sugar beet (Beta vulgaris L.) seedlings were grown on media containing 90 to 300 mM sucrose or glucose. Compared to controls, sugar-grown plants had higher growth rate, photosynthesis, and leaf sugar levels. The steady-state level of transcripts increased signifi- cantly for the small subunit of ribulose-1 ,s-bisphosphate carboxy- lase/oxygenase (Rubisco) (rbcS) and the cytosolic fructose-l,6- bisphosphatase and moderately for the Rubisco large subunit (rbcl). l h e transcript level of sucrose phosphate synthase remained un- changed. Fructose-1,6-bisphosphatase and Rubisco activities did not change in the presence of sugars, but that of sucrose phosphate synthase increased (44 and 90% under selective and nonselective assay conditions, respectively). Accelerated leaf development was indicated by (a) autoradiograms of leaves that showed that sucrose loading occurred earlier, (b) export capacity that also occurred earlier but, after about 2 weeks, differences were not detectable, and (c) sucrose synthase activity that declined significantly. Several conclusions emerged: (a) response was nonosmotic and gene and sugar specific, (b) sugars caused accelerated leaf development and sink-to-source transition, ( c ) enhanced gene expression was due to advanced leaf development, and (d) whereas Rubisco and cytosolic fructose-l,6-bisphosphatase genes were sugar repressed in mature leaves of greenhouse-grown plants, they were unaffected in mature, culture-grown leaves. To our knowledge, these data provide the first evidence in higher plants that, depending on the physiological/ developmental context of leaves, sugars lead to differential regula- t ion of the same gene.
Excessive sugar accumulation in leaves (e.g. when sink demand is lagging behind carbon supply) results in alter- ations in photosynthesis and carbohydrate partitioning in leaves (Stitt and Quick, 1989; Stitt, 1990; Krapp et al., 1991; Goldschmidt and Huber, 1992; Daie, 1993). This end-prod- uct control is similar to the role sugars (and other end products for that matter) play in a wide range of microor- ganisms and mammalian systems in altering, both at the biochemical and molecular levels, major metabolic path- ways (Entain, 1980; Lenz and Holzer, 1980; Lee, 1987; El- Maghrabi et al., 1988). Although a relatively large body of
' This work was supported partially by funds from a U.S. De- partment of Agriculture-National Research Initiative Competitive Grants Program grant (92-37306-8331) to J.D.
* Corresponding author; fax 1-608-265-5482.
literature provides evidence concerning the physiological and biochemical bases of the phenomenon in plants, only recently has molecular evidence become available about sugar-mediated expression of carbon fixation and/or me- tabolism genes (Sheen, 1990, 1994).
Depending on the experimental system, the tissue, and the gene of interest, sugars have been shown to repress (down-regulation) or induce (up-regulation) the expression of different genes that encode key enzymes of carbon fix- ation and metabolism. For example, the Suc synthase gene in the maize endosperm and the ADP-Glc pyrophosphory- lase gene in potato are up-regulated by Suc (Chourey et al., 1986; Salanoubat and Belliard, 1989; Takeda et al., 1994), and severa1 photosynthetic genes as well as carbon metab- olism genes encoding leaf enzymes are repressed in the presence of Suc or Glc (Sheen, 1990; Krapp et al., 1991,1993; Kossmann et al., 1992; J.S. Lee and J. Daie, unpublished results). Presumably, sugar-responsive regulatory se- quences (cis elements) are present at the 5' region of these genes, as has been shown in the case of a number of photosynthetic and carbon metabolism genes (Maas et al., 1990; Sheen, 1990; Huang et al., 1993), as well as for genes that are not involved in carbon metabolism (patatin gene, Wenzler et al., 1989; chalcone-A gene, Tsukaya et al., 1991; nitrate reductase gene, Cheng et al., 1992).
Accumulation of photosynthetic products in leaves was suggested by Wang et al. (1993) to influence the pattern of photosynthetic gene expression in C, amaranth plants. It is therefore conceivable that the developmental and/or phys- iological stage of the leaf could play a role in how leaf carbon balance might influence patterns of gene expression in other plants. Several lines of evidence, in fact, support this proposition. For example, although Glc repressed ex- pression of the rbcS in mature leaves of transgenic tobacco, it did not repress rbcS in immature leaves of those plants that accumulated large quantities of Glc in the leaf result- ing from the overexpression of a yeast-derived invertase (Von Schaewen et al., 1990; Dickinson et al., 1991; Krapp et al., 1993). Likewise, Brusslan and Tobin (1992) reported
Abbreviations: chl-FBPase, chloroplastic Fru-1,6-bisphos- phatase; cyt-FBPase, cytosolic Fru-1,6-bisphosphatase; vbcS and rbcL, small and large subunits of Rubisco, respectively; SPS, Suc phosphate synthase.
1647 www.plantphysiol.orgon June 24, 2018 - Published by Downloaded from Copyright © 1995 American Society of Plant Biologists. All rights reserved.

1648 Kovtun and Daie Plant Physiol. Vol. 108, 1995
that the cab-1 gene was not repressed in etiolated Arabi- dopsis seedlings (grown on Suc-containing media) despite reports of its sugar repression in mature plants. Evidence from other (nonplant) systems, however, is more definitive and shows that, depending on the physiological and/or metabolic context of the cell and developmental stage of the system, molecular signals may serve to either up- or down-regulate the same gene (Miesfeld et al., 1987; Sakai et al., 1988; Diamond et al., 1990).
As part of our efforts to investigate the role of sugars in metabolic repression of transcription in sugar beet leaves, we developed a culture-grown sugar beet seedling system that would allow better control of the variables. Surpris- ingly, but consistent with the conclusions of Diamond et al. (1990), rather than the expected repression by sugars, which we had observed in mature leaves of greenhouse- grown sugar beets, enhanced expression of the same genes occurred (in a highly gene-specific manner) in the culture- grown seedlings. In this paper we report molecular and physiological evidence for sugar-induced accelerated leaf development, sink-to-source transition, and enhanced ex- pression of some carbon metabolism genes in the leaves of culture-grown sugar beet plants. Moreover, we provide the first evidence, to our knowledge, that genes that are sugar repressed in mature leaves of greenhouse/soil-grown plants are either enhanced (in developing leaves) or unaf- fected (in mature leaves) in culture-grown plants.
MATERIALS AND METHODS
Plant Materials
Sugar beet (Beta vulgaris L. cv mono-HyE4) seeds were sterilized in 70% ethanol for 1 min and then were placed in a 20% household bleach (Clorox) solution for 20 min. After the seeds were washed three to four times in water, they were placed in Petri dishes on filter paper that was wetted with a basal culture medium (Sigma), which was the stan- dard salts and additives of Murashige and Skoog (1962). One week later, the seedlings were transferred to clear plastic boxes (GA-7-Magenta box) to which was added 40 mL of the basal medium solidified with 0.8% agar alone (control) or agar plus various concentrations of (90-300 mM) Glc or SUC. A set of plants was placed on osmotically adjusted medium (with ethylene glycol to 300 mM) or 3-O-methyl Glc and monitored as osmotic controls. The growth conditions were 14 h of light (high-pressure so- dium lamps, 250 /LE m-' s-' at the plant level) and day/ night temperatures of 26/22"C. To determine age, emerg- ing leaves (not longer than 5 mm) were dated. Unless otherwise stated in a figure or table caption, leaf sets of the same age (ranging from 10-14 d old within each treatment) were sampled from 4-week-old plants. Leaves from each treatment were pooled before they were frozen in liquid nitrogen and stored at -80°C for further analysis. AI1 experiments were done as triplicates and repeated at least twice. Each triplicate consisted of six to eight leaves (pooled) obtained from three to four different plants.
Vein Accumulation of SUC and Development of Loading Capacity
At least three plants were used for each treatment. Uni- formly labeled ['4ClSuc was delivered through the transpi- ration stream under continuous light conditions (same as growing conditions). A plant stem (with at least two leaf pairs on it) was placed in a solution containing 10 mM unlabeled SUC, 5 mM Mes (pH 5.5), and 60 kBq mL-' of ['4C]Suc (specific activity, 671 mCi/mM). Uptake via the transpiration stream was allowed to continue for 6 h. After exposure to labeled SUC, leaves were cut and washed three times in chilled water (removing most of the label in the apoplastic space). For dark experiments, plants were placed in a chamber in a dark room for 3 h before they were placed in labeled Suc solution for another 6 h of uptake in the dark. Autoradiograms were obtained by placing leaves against x-ray films (Kodak Industrex M) for 10 to 14 d at -80°C. To demonstrate the leaf ability to export SUC, a solution containing 10 mM Suc spiked with 20 kBq ['4C]Suc was applied to the leaf surface, which had been abraded with Carborundum. Translocation of label out of the leaf was allowed to occur in the light for 4 to 10 h, after which excess radioactivity on the leaf surface was removed (by washing the leaf three times, 5 min each, in chilled water). Plants were dissected into various parts before they were digested and bleached overnight in a 4:6 (v/v) perchloric acid:hydrogen peroxide solution at 50°C (as described by Pitcher and Daie, 1991). The label present in various plant segments was counted as disintegrations per minute in a scintillation counter.
Sugar Feeding of Leaves
These experiments were done to determine whether ma- ture leaves from the culture-grown plants respond to sug- ars in a way similar to mature leaves from greenhouse/ soil-grown plants. Fully expanded mature leaves from 4-week-old culture-grown plants were harvested in the middle of the light period (11 AM, maximum mRNA levels) and were immediately placed in water, where the petioles were cut again under water to avoid xylem embolism. The petioles were then placed in sugar solutions (100 mM Glc or mannitol as control) for 24 h under constant light. At least five leaves (each from a different plant) were used for each treatment. In the case of greenhouse-grown plants, discs from fully expanded, mature leaves of 4-month-old plants were placed for 24 h in 40 mM solutions (aerated) of man- nitol (control), SUC, Glc, or Fru (data presented here are from a separate set of experiments done entirely with greenhouse-grown plants).
RNA lsolation and Blot Analysis
Total RNA was isolated as described by Chomczynski and Sacchi (1987) and modified for sugar beet as described by Harn et al. (1993). Briefly, 500 mg of frozen tissue were ground in 5 mL of a solution containing 4 M guanidium thiocyanate, 25 mM sodium citrate (pH 7.0), 0.5% sarcosyl, and 0.1 M P-mercaptoethanol. Five milliliters of water- saturated phenol, 2 mL of 2 M sodium acetate, and 1 mL of
www.plantphysiol.orgon June 24, 2018 - Published by Downloaded from Copyright © 1995 American Society of Plant Biologists. All rights reserved.

Accelerated Leaf Development 1649
ch1oroform:isoamyl alcohol (49:l) were sequentially added to the homogenate, and the final suspension was centri- fuged at l0,OOOg for 20 min. An equal volume of isopropa- no1 was added to the aqueous phase and RNA was precip- itated at 10,OOOg for 20 min. Total RNA (15 p g ) was then loaded onto each lane of a 1% agarose gel and fractionated at 15 V overnight. The gel was then blotted onto a nylon membrane (Hybond-N) soaked in 1OX SSC (3 M NaCl and 0.3 M sodium citrate, pH 7.0) and placed in a Vacugene pump (Pharmacia). The membrane was baked at 80°C for 2 h and hybridized in a solution containing 0.25 mM Na2HPO,*7H,O, pH 7.4, 1 mM EDTA, 1% casein, and 7% SDS with a 32P-labeled cDNA for 20 h at 63°C. The cDNA probes were sugar beet cyt-FBPase (Harn and Daie, 1992a), a 0.52-kb insert from maize SPS (most conserved region located at the middle of the sequence). The full-length SPS cDNA was a gift from Calgene, Inc. (Davis, CA), the se- quence of which was reported by Worrell et al. (1991); the amaranth rbcL and rbcS by Berry et al. (1985); and the radish rRNA by Delseny et al. (1983). Signal intensities were quantified by densitometric scanning with a Laser Scanning Densitometer (LKB, Uppsala, Sweden).
Leaf Photosynthesis and Chl Determination
A portable Li-Cor (Lincoln, NE) LI-6200 was used to measure leaf photosynthesis rates. Chl was determined after the tissue was homogenized in chilled 80% acetone. The homogenate was centrifuged, and Chl content was determined as described by Arnon (1949).
Enzyme Activity
Frozen tissue was homogenized in cold extraction buffer (50 mM Mops-NaOH [pH 7.5],1 mM EDTA, 15 mM MgCl,, 2.5 mM DTT, 0.1% Triton X-100, 2% polyvinylpolypyroli- done, 1 mM PMSF). After the sample was centrifuged at l0,OOOg for 5 min, the supernatant was used to determine enzyme activity and sugar content. Protein content was determined by the method of Bradford (1976) using the Bio-Rad protein assay kit and BSA as the standard.
Activities of the cyt-FBPase and chl-FBPase were assayed according to the method of Kelly et al. (1982) as modified by Harn and Daie (1992b). The production of Fru-6-P was coupled to the reduction of NADP using phosphoglucose isomerase and Glc-6-P dehydrogenase. For the cyt-FBPase, the assay mixture contained 100 mM imidazole (pH 7.0), 5 mM MgCl,, 0.3 mM NADP, and 0.3 mM Fru-1,6-bisphos- phate. The assay mixture for the chl-FBPase contained 100 mM Tris-HC1 (pH 8.8), 10 mM MgCl,, 0.6 mM Fru-1,6- bisphosphate, 0.3 mM NADP, and 0.5 mM EDTA.
To determine SPS activity, crude extracts were desalted on a Sephadex G-25 column equilibrated with extraction buffer minus Triton X-100, polyvinylpolypyrolidone, and PMSF. Activity was then determined as described by Hu- ber et al. (1991). The "selective" (substrate-limiting) assay mixture contained 6 mM Fru-6-P, 24 mM Glc-6-P, 20 mM uridine diphosphoglucose, and 20 mM Pi. The "nonselec- tive" (substrate-saturating) assay contained 20, 80, and 20 mM of the first three ingredients and no Pi. Aliquots of the
desalted extracts were incubated for 10 min at 27°C. The reaction was stopped by adding an equal volume of 30% KOH, followed by boiling for 10 min. Suc was quantified by the anthrone reaction method (Ashwell, 1957).
Suc synthase activity was assayed as described by More11 and Copeland (1985) with some modifications. The assay mixture added to aliquots of the desalted extract contained 50 mM Mops-NaOH (pH 7.5), 15 mM Fru, 15 mM MgCl,, and 15 mM uridine diphosphoglucose. The mixture was incubated at 30°C for 30 min, after which the reaction was stopped by adding an equal volume of 30% KOH. Rubisco activity was measured by incorporation of I4C from so- dium bicarbonate into acid-stable material as described by Ghosh et al. (1989) and modified by Dreesmann et al. (1994). Invertase activity was determined by the method of Claussen et al. (1986).
Carbohydrate Analysis
Starch content was determined using the method de- scribed by Galtier et al. (1993). Frozen tissue was ground in 1 M HCIO,. After the sample was centrifuged, the pellet was extracted in 80% acetone and resuspended in water. The suspension was boiled for 60 min and incubated in 50 mM sodium acetate buffer (pH 4.6) containing 3 units of amylase and 60 units of amyloglucosidae for another 60 min at 50°C. Glc was measured using a kit (Sigma). Suc was quantitated by the anthron method (Ashwell, 1957).
RESULTS
Expression of Some Genes Was Enhanced in Sugar-Grown Plants
RNA blot analysis of rbcL, rbcS, cyt-FBPase, SPS, and 18s rRNA are presented in Figure 1. The transcript level of 18s rRNA remained unchanged (less than 10% variations) un- der a11 treatments, ruling out nonspecific effects on gene expression. Steady-state transcript levels of rbcL, rbcS, and cyt-FBPase increased in the presence of 90 mM Glc (20,40, and 90% increase, respectively), 300 mM Glc (30, 62, and 118% increase, respectively), 90 mM (21, 48, and 92% in- crease, respectively) SUC, and 150 mM (37, 71, and 129% increase, respectively). The response of the rbcL gene was less dramatic than that of rbcS and cyt-FBPase genes. Sug- ars did not have any effect on SPS transcript level, suggest- ing the gene-specific nature of the response. A lack of transcriptional control of SPS is consistent with strong evidence indicating that SPS regulation and its coordina- tion with photosynthesis occur mainly at the biochemical level, i.e. fine control (Stitt and Quick, 1989) and coarse control (posttranslational modification) of SPS protein (Hu- ber and Huber, 1992).
For the genes that did respond, 300 mM Suc was not optimal. Transcript levels were not as high as those in plants grown at lower Suc concentrations (90 or 150 mM). Concentration-dependent gene expression has been ob- served for other systems, such as potato SUC synthase, ADP-Glc pyrophosphorylase, and granule-bound starch synthase genes (Maas et al., 1990; Muller-Rober et al., 1990; Van der Steege et al., 1992).
www.plantphysiol.orgon June 24, 2018 - Published by Downloaded from Copyright © 1995 American Society of Plant Biologists. All rights reserved.

1650 Kovtun and Dale Plant Physiol. Vol. 108, 1995
c0
SucroseGlucose90 300 90 150 300
rbcl
rbcS
cyt-FBPase
SPS 4H H»
ISSrRNA
Figure 1. Total RNA blot analysis of genes encoding four carbonmetabolism enzymes (cyt-FBPase, rbcS, rbcl, and SPS). Sampleswere taken from 4-week-old, culture-grown sugar beet seedlings.Numbers at the top are sugar concentrations (in mM) in the agarmedia. 18S rRNA was used as an internal control. C, Control (mediawithout sugar and osmoticum).
Furthermore, 3-O-methyl Glc, a nonmetabolized analogof Glc, was ineffective in terms of altering the patterns ofgene expression (Fig. 2). Since 3-O-methyl Glc had littleeffect on gene expression in our hands and by others(Krapp et al., 1993; Jang and Sheen, 1994), it appeared thatsome sugar metabolite (or only sugars that can be alteredby metabolism) would have served as the direct molecularsignal in the signal's transduction pathway. Jang and Sheen(1994) recently showed that sugars that serve as substratefor a hexose kinase are direct signals that mediate controlof transcription of photosynthetic genes (in that case re-pression). Based on their observations, they proposed arole for hexose kinase as both a sensor and the transmitterof the signal.
Ineffectiveness of 300 mM 3-O-methyl Glc (which can beconsidered as an osmoticum) suggested the lack of anosmotic effect on gene expression in the system. In addi-tion, we know from our previous work (Harn and Daie,1992b; Dreesmann et al., 1994) that the transcript levels ofcyt-FBPase, Rubisco (both genes), and SPS genes in sugarbeet remain unchanged under water-stress conditions(moderate but long term, 4-5 d). Lack of osmotic effects onsugar-responsive genes have also been concluded by sev-eral other research groups (Krapp et al., 1993; Sheen, 1994).Nonetheless, initial experiments were done with osmoti-cum control (300 mM), which confirmed negligible effectsof the osmotic potential of the media.
Plants Grew Faster on Sugar-Containing Media
Table I presents data on growth parameters of seedlingsgrown for 4 weeks on media containing various levels of
sugar. Leaves of sugar-grown seedlings were darker greenand thicker than control plants. In general, a positive cor-relation existed between biomass production and sugarconcentration in the media; maximal shoot and root growth(2- to 3-fold) occurred with either 300 mM Glc or 150 mMSue (optimal Sue concentration). Growth parameters ofplants grown on osmoticum only (osmotic controls) werenot significantly different from those in controls (withoutosmoticum and sugar), again ruling out nonspecific os-motic effects. In addition, sugar-grown plants had 2- to3-fold higher total protein and total RNA, Chl content, andleaf photosynthesis rates, whereas osmotic control plantsdid not (Table II).
Whereas internal Sue and Glc levels increased 1.5- to3-fold, leaf starch did not change in response to sugar in themedia (Table III). Moreover, concentrations of both Sue andGlc increased in the leaves irrespective of the carbon sourcein the media. Increased leaf Glc content in the presence ofSue is assumed to have been due to invertase activity(which remained high in sugar-grown plants; see below).Also, Glc-grown plants had higher Sue levels compared tocontrols, suggesting either an increased capacity for denovo Sue synthesis (confirmed by higher cyt-FBPase andSPS activities; Table IV) or the possibility for conversion ofexcess Glc to Sue. Collectively, these observations, alongwith that of enhanced gene expression, prompted us tofurther study and characterize the culture-grown system inthe context of leaf carbon metabolism, leaf development,and sink-to-source transition.
Activity of Some Enzymes Was Higher inSugar-Grown Plants
The activity of Rubisco, cyt-FBPase, and chl-FBPase re-mained unchanged in the presence of sugars, but SPSactivity under both nonselective and selective assay condi-tions increased 44 and 90%, respectively (Table IV). On aleaf area basis, the activities of all measured enzymes weresignificantly greater in sugar-grown plants than in con-trols, which is a reflection of the overall increase in leafprotein content (except SPS, which showed an increase inspecific activity as well). Noteworthy is the increase in the
MG
rbcS
Cyt-FBPase
Figure 2. Total RNA blot analysis of rfacS and cyt-FBPase genes.Samples were taken from 4-week-old, culture-grown sugar beet seed-lings. C, Control; S, 150 mM Sue; C, 300 mM Glc; and MG, 300 mM3-O-methyl Glc. www.plantphysiol.orgon June 24, 2018 - Published by Downloaded from
Copyright © 1995 American Society of Plant Biologists. All rights reserved.
Accelerated Leaf Development 1651
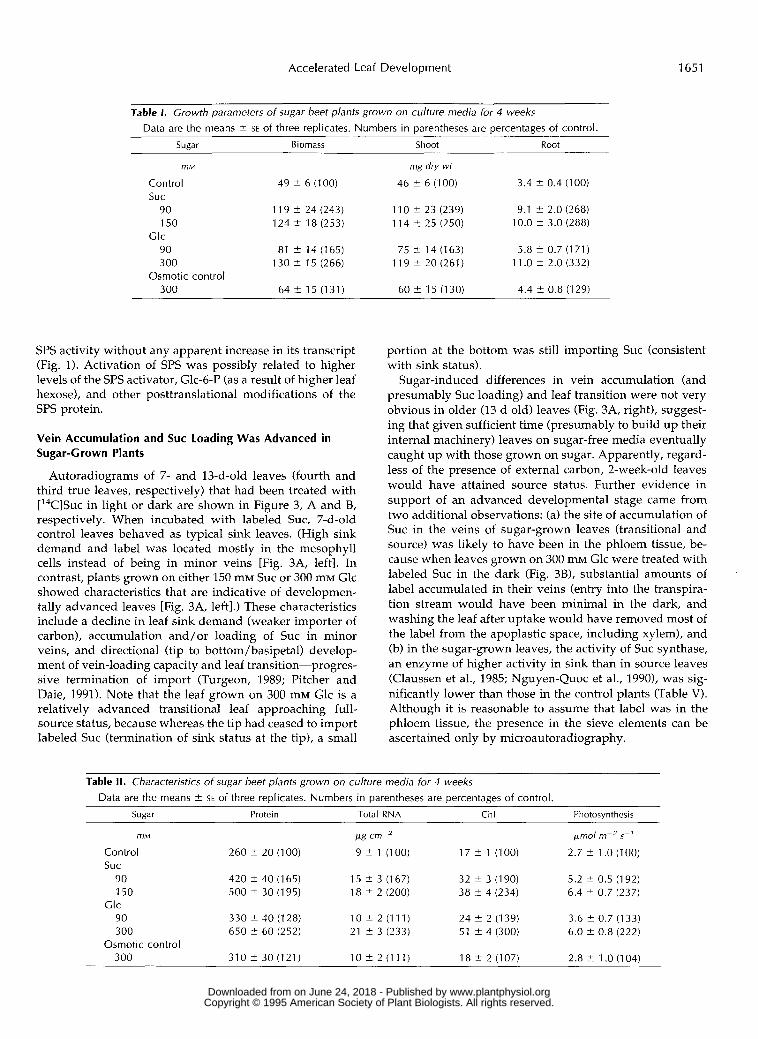
Table 1. Crowth parameters o f sugar beet plants grown on culture media for 4 weeks
Data are the means 2 SE of three replicates. Numbers in parentheses are percentages of control. Sugar Biomass Shoot Root
mM mg dry wt
Control 49 t- 6 (100) 46 i 6 (100) 3.4 2 0.4 (100) SUC
90 119 2 24 (243) 110 2 23 (239) 9.1 t 2.0 (268) 150 124 t- 18 (253) 114 t- 25 (250) 10.0 2 3.0 (288)
90 81 t 14(165) 75 t 14 (163) 5.8 i 0.7 (171) 300 130 t 15 (266) 119?20(261) 11 .O i 2.0 (332)
300 64 i 15 (131) 60 t- 15 (130) 4.4 2 0.8 (1 29)
Clc
Osmotic control
SPS activity without any apparent increase in its transcript (Fig. 1). Activation of SPS was possibly related to higher levels of the SPS activator, Glc-6-P (as a result of higher leaf hexose), and other posttranslational modifications of the SPS protein.
Vein Accumulation and SUC Loading Was Advanced in Sugar-Crown Plants
Autoradiograms of 7- and 13-d-old leaves (fourth and third true leaves, respectively) that had been treated with ['4C]Suc in light or dark are shown in Figure 3, A and B, respectively. When incubated with labeled SUC, 7-d-old control leaves behaved as typical sink leaves. (High sink demand and label was located mostly in the mesophyll cells instead of being in minor veins [Fig. 3A, left]. In contrast, plants grown on either 150 mM Suc or 300 mM Glc showed characteristics that are indicative of developmen- tally advanced leaves [Fig. 3A, left].) These characteristics include a decline in leaf sink demand (weaker importer of carbon), accumulation and/or loading of Suc in minor veins, and directional (tip to bottom/basipetal) develop- ment of vein-loading capacity and leaf transition-progres- sive termination of import (Turgeon, 1989; Pitcher and Daie, 1991). Note that the leaf grown on 300 mM Glc is a relatively advanced transitional leaf approaching full- source status, because whereas the tip had ceased to import labeled Suc (termination of sink status at the tip), a small
portion at the bottom was still importing Suc (consistent with sink status).
Sugar-induced differences in vein accumulation (and presumably Suc loading) and leaf transition were not very obvious in older (13 d old) leaves (Fig. 3A, right), suggest- ing that given sufficient time (presumably to build up their interna1 machinery) leaves on sugar-free media eventually caught up with those grown on sugar. Apparently, regard- less of the presence of externa1 carbon, 2-week-old leaves would have attained source status. Further evidence in support of an advanced developmental stage came from two additional observations: (a) the site of accumulation of Suc in the veins of sugar-grown leaves (transitional and source) was likely to have been in the phloem tissue, be- cause when leaves grown on 300 mM Glc were treated with labeled Suc in the dark (Fig. 38), substantial amounts of label accumulated in their veins (entry into the transpira- tion stream would have been minimal in the dark, and washing the leaf after uptake would have removed most of the label from the apoplastic space, including xylem), and (b) in the sugar-grown leaves, the activity of Suc synthase, an enzyme of higher activity in sink than in source leaves (Claussen et al., 1985; Nguyen-Quoc et al., 1990), was sig- nificantly lower than those in the control plants (Table V). Although it is reasonable to assume that label was in the phloem tissue, the presence in the sieve elements can be ascertained only by microautoradiography.
Table II. Characteristics o f sugar beet plants grown on culture media for 4 weeks Data are the means t- SE of three replicates. Numbers in parentheses are percentages of control.
Sugar Protein Total RNA Chl Photosvnthesis
mM
Control 260 2 20 (100) SUC
90 420 t- 40 (165) 150 500 2 30 (195)
90 330 t- 40 (128) 300 650 i 60 (252)
300 310 2 30 (121)
Clc
Osmotic control
pg cm-'
9 2 1 (100)
15 t 3 (167) 18 2 2 (200)
10 2 2 (111) 21 i 3 (233)
10 i 2 (111)
ymol m-z 5~~
2.7 2 1 .O (100)
5.2 t- 0.5 (1 92) 6.4 i 0.7 (237)
3.6 i 0.7 (1 33) 6.0 i 0.8 (222)
17 2 l ( lO0)
32 t- 3 (190) 38 2 4 (234)
24 t- 2 (139) 51 t- 4 (300)
2.8 2 1 .O (1 04) 18 t- 2 (107)
www.plantphysiol.orgon June 24, 2018 - Published by Downloaded from Copyright © 1995 American Society of Plant Biologists. All rights reserved.
1652 Kovtun and Daie Plant Physiol. Vol. 108, 1995
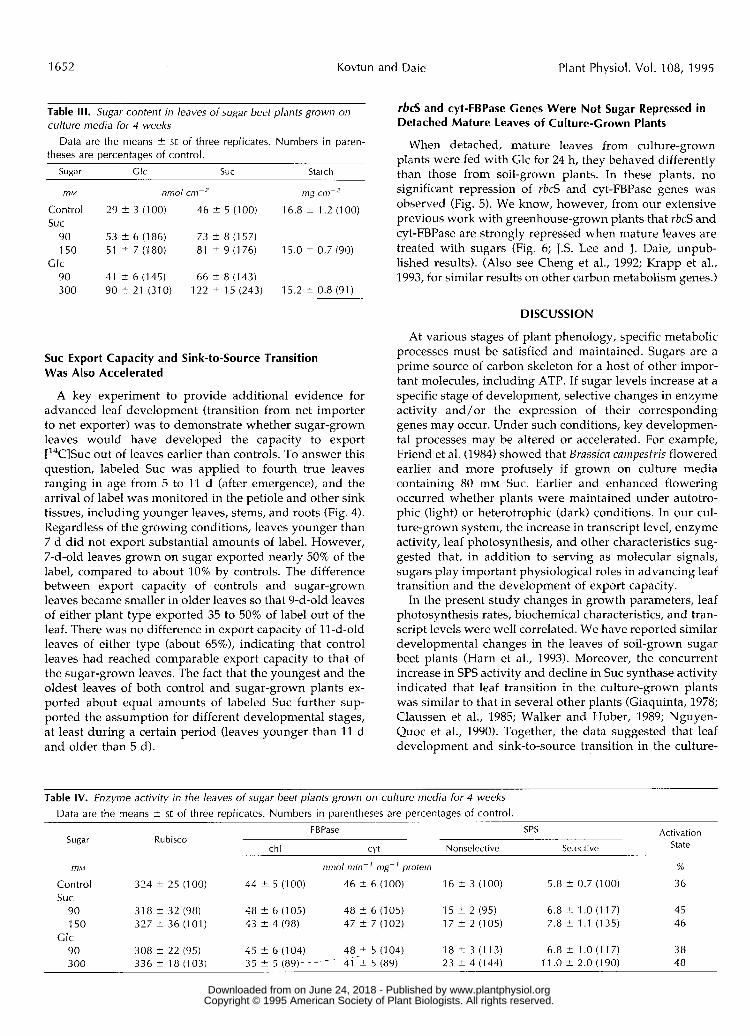
Table 111. Sugar content in leaves o f sugar beet plants grown on culture media for 4 weeks
theses are percentages of control. Data are the means 2 SE of three replicates. Numbers in paren-
Sunar Glc SUC Starch
mM nmol cm-' mg cm-'
Control suc
90 150 51 i 7 (180) 81 i 9 (176) 15.0 i 0.7 (90)
90 41 ? 6 (145) 66 2 8 (143) 300 90 2 21 (310) 122 2 15 (243) 15.2 ? 0.8 (91)
29 t 3 (1 00)
53 i 6 (1 86)
46 i 5 (1 00)
73 2 8 (1 57)
16.8 t 1.2 (1 00)
Glc
rbcS and cyt-FBPase Genes Were Not Sugar Repressed in Detached Mature Leaves of Culture-Grown Plants
When detached, mature leaves from culture-grown plants were fed with Glc for 24 h, they behaved differently than those from soil-grown plants. In these plants, no significant repression of rbcS and cyt-FBPase genes was observed (Fig. 5). We know, however, from our extensive previous work with greenhouse-grown plants that rbcS and cyt-FBPase are strongly repressed when mature leaves are treated with sugars (Fig. 6; J.S. Lee and J. Daie, unpub- lished results). (Also see Cheng et al., 1992; Krapp et al., 1993, for similar results on other carbon metabolism genes.)
SUC Export Capacity and Sink-to-Source Transition Was Also Accelerated
A key experiment to provide additional evidence for advanced leaf development (transition from net importer to net exporter) was to demonstrate whether sugar-grown leaves would have developed the capacity to export ['4C]Suc out of leaves earlier than controls. To answer this question, labeled Suc was applied to fourth true leaves ranging in age from 5 to 11 d (after emergence), and the arrival of label was monitored in the petiole and other sink tissues, including younger leaves, stems, and roots (Fig. 4). Regardless of the growing conditions, leaves younger than 7 d did not export substantial amounts of label. However, 7-d-old leaves grown on sugar exported nearly 50% of the label, compared to about 10% by controls. The difference between export capacity of controls and sugar-grown leaves became smaller in older leaves so that 9-d-old leaves of either plant type exported 35 to 50% of label out of the leaf. There was no difference in export capacity of 11-d-old leaves of either type (about 65%), indicating that control leaves had reached comparable export capacity to that of the sugar-grown leaves. The fact that the youngest and the oldest leaves of both control and sugar-grown plants ex- ported about equal amounts of labeled Suc further sup- ported the assumption for different developmental stages, at least during a certain period (leaves younger than 11 d and older than 5 d).
DISCUSSION
At various stages of plant phenology, specific metabolic processes must be satisfied and maintained. Sugars are a prime source of carbon skeleton for a host of other impor- tant molecules, including ATP. If sugar levels increase at a specific stage of development, selective changes in enzyme activity and/or the expression of their corresponding genes may occur. Under such conditions, key developmen- tal processes may be altered or accelerated. For example, Friend et al. (1984) showed that Brassica campestris flowered earlier and more profusely if grown on culture media containing 80 mM SUC. Earlier and enhanced flowering occurred whether plants were maintained under autotro- phic (light) or heterotrophic (dark) conditions. In our cul- ture-grown system, the increase in transcript level, enzyme activity, leaf photosynthesis, and other characteristics sug- gested that, in addition to serving as molecular signals, sugars play important physiological roles in advancing leaf transition and the development of export capacity.
In the present study changes in growth parameters, leaf photosynthesis rates, biochemical characteristics, and tran- script levels were well correlated. We have reported similar developmental changes in the leaves of soil-grown sugar beet plants (Harn et al., 1993). Moreover, the concurrent increase in SPS activity and decline in Suc synthase activity indicated that leaf transition in the culture-grown plants was similar to that in severa1 other plants (Giaquinta, 1978; Claussen et al., 1985; Walker and Huber, 1989; Nguyen- Quoc et al., 1990). Together, the data suggested that leaf development and sink-to-source transition in the culture-
Table IV. Enzyme activity in the leaves of sugar beet plants grown on culture media for 4 weeks Data are the means i SE of three replicates. Numbers in parentheses are percentages of control.
FBPase SPS Activation State Sugar Rubisco
chl C\'t Nonselective Selective
f7lM nmol min- ' mg-' protein %
Control 324 2 25 (100) 44 t 5 (100) 46 ? 6 (100) 16 t 3 (100) 5.8 2 0.7 (100) 36 suc
90 318 i 32 (98) 48 i 6 (105) 48 i 6 (105) 15 f 2 (95) 6.8 t 1 .O (1 17) 45 150 327 t 36 (101) 43 2 4 (98) 47 ? 7 (102) 17 2 2 (105) 7.8 i 1.1 (1 35) 46
90 308 2 22 (95) 45 i 6 (104) 48-? 5 (104) 18 t 3 (113) 6.8 ? 1 .O (1 17) 38 3 00 3 3 6 k 18 (103) 35 2 5 (89)- -I-- 41 ? 5 (89) 23 2 4 (144) 1 1 .o ? 2.0 (1 90) 48
Glc
www.plantphysiol.orgon June 24, 2018 - Published by Downloaded from Copyright © 1995 American Society of Plant Biologists. All rights reserved.

Accelerated Leaf Development 1653
Sucrose (ISO mM)
lit
t*
Glucose (300 mM)
Glucose (300 mM)
Figure 3. Autoradiograms of leaves from culture-grown sugar beetseedlings grown for 4 weeks in the presence of 0 to 300 mM Sue orGlc. Left and right panels are 7- and 13-d-old leaves, respectively.Seedling stems were placed in 10 mM labeled Sue solutions for 6 hunder light conditions (A) or in the dark (B). Same-age leaves wereused as described in "Materials and Methods."
grown plants were fundamentally similar to processes thatoccur in developing leaves of soil-grown plants.
Based on work with a culture-grown, albino tobaccosystem, Turgeon (1989) concluded that preprogrammed,developmental processes (i.e. leaf transition) override met-abolic conditions in the leaves. However, in our culture-
Table V. Enzyme activity in the leaves of sugar beet plants grownon culture media for 4 weeks
Data are the means ± SE of three replicates. Numbers in paren-theses are percentages of control.
Sugar
mM
ControlSue
90150
Glc90300
Sue Synthase
nmol mlrT '
7.6 ± 1.1 (100)
6.2 ± 1.0(82)4.6 ± 0.6 (60)
8.6 ± 1.3 (113)2.5 ± 0.1 (33)
Invertase
mg~ ' protein
40.8 ± 3.8(100)
22.4 ± 0.3 (55)24.6 ± 0.9 (60)
35.2 ± 2.2 (86)29.4 ± 1.3 (72)
grown plants, higher leaf carbon balance was correlatedwith an accelerated pace of sink-to-source transition, sug-gesting that metabolic conditions (elevated leaf sugar con-tent) may override some aspects of leaf development. Inaddition to the present data, other evidence suggests thatmetabolic conditions can override some types of regulation(e.g. light-dependent expression of rbcS [Sheen, 1990] andchl-FBPase [Kossmann et al., 1992]). Chourey and Taliercio(1994) recently concluded that metabolic regulatory con-trols override the normal controls of tissue and cell speci-ficity for the expression of two Sue synthase genes inmaize.
Evidence obtained in our laboratory with 4-month-old,greenhouse-grown sugar beet plants (Fig. 6; J.S. Lee and J.Daie, unpublished results) indicates that when matureleaves are exposed to sugar solutions some genes encodingcarbon metabolism enzymes are repressed significantlyand rapidly (as early as 4 h). In such sugar-feeding exper-iments, repression was most pronounced for rbcS and cyt-FBPase and moderate for rbcL, and SPS expression was notaffected. Those results were consistent with other work inwhich sugar repression of various photosynthetic geneswas reported (Sheen, 1990; Cheng et al., 1992; Criqui et al.,1992; Harter et al., 1993; Krapp et al., 1993). To our surprise,results of this study with culture-grown plants revealedthat the expression of rbcS and cyt-FBPase genes wasstrongly enhanced in sugar-grown plants. Furthermore,rbcL expression was moderately enhanced and expressionof SPS gene was unaltered. Therefore, the sugar responsewas gene specific, whether it was in the direction of repres-sion or enhancement. It is interesting that there was alsoconsistency in the magnitude of the response in the twosystems. It is likely that the actual in situ (compartmental)
100J IGlucose >,40
70 -
~ni§ 60 -S?
8 50-oo
C/3 40
P| 30-
o& 20 -
LJJ
10-
0
CDControl '
4J
-28
-24__^
-20 b^s
-16 §
.,.?-8
-4
0
5 7 9 11Leaf age (days)
Figure 4. Export of labeled Sue out of leaves by 4-week-old, culture-grown sugar beet seedlings grown with or without 300 mM Glc in theagar media. Labeled Sue (10 mM) was applied to same-age, abradedattached leaves. Translocation out of the leaf was allowed to con-tinue for up to 10 h under light conditions. After the plant waswashed, label was measured in all plant segments. Export was cal-culated from label in the petiole, stem, and roots as a percentage ofthe total label taken up by the leaf. www.plantphysiol.orgon June 24, 2018 - Published by Downloaded from
Copyright © 1995 American Society of Plant Biologists. All rights reserved.
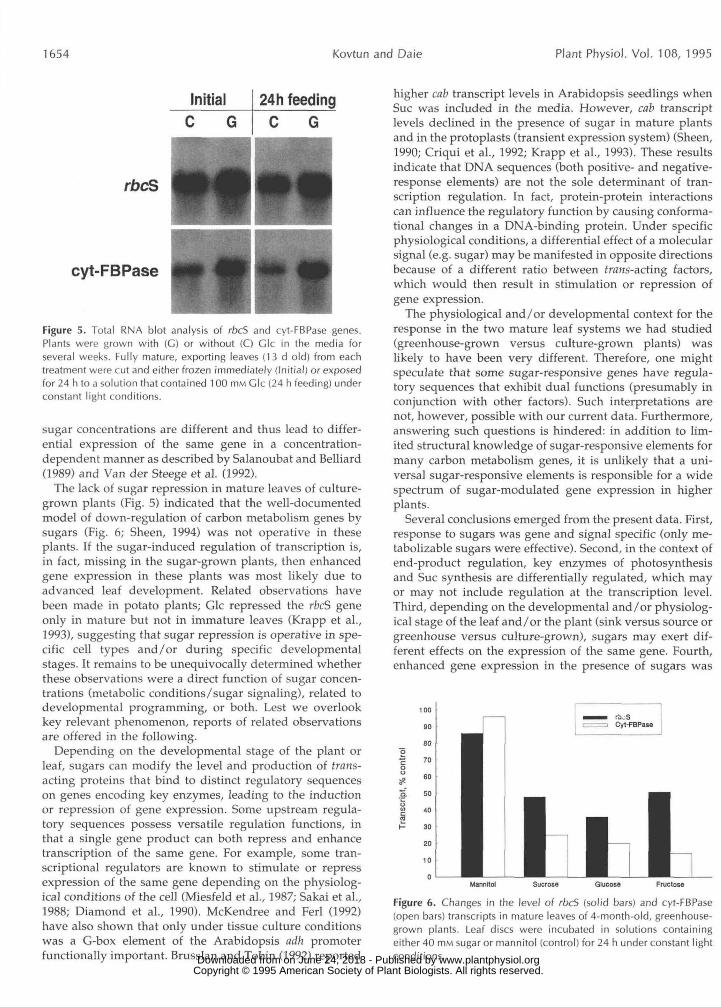
1654 Kovtun and Dale Plant Physiol. Vol. 108, 1995
Initial 24h feeding
rbcS
cyt-FBPase
Figure 5. Total RNA blot analysis of rbcS and cyt-FBPase genes.Plants were grown with (C) or without (C) Glc in the media forseveral weeks. Fully mature, exporting leaves ( 1 3 d old) from eachtreatment were cut and either frozen immediately (Initial) or exposedfor 24 h to a solution that contained 100 HIM Glc (24 h feeding) underconstant light conditions.
sugar concentrations are different and thus lead to differ-ential expression of the same gene in a concentration-dependent manner as described by Salanoubat and Belliard(1989) and Van der Steege et al. (1992).
The lack of sugar repression in mature leaves of culture-grown plants (Fig. 5) indicated that the well-documentedmodel of down-regulation of carbon metabolism genes bysugars (Fig. 6; Sheen, 1994) was not operative in theseplants. If the sugar-induced regulation of transcription is,in fact, missing in the sugar-grown plants, then enhancedgene expression in these plants was most likely due toadvanced leaf development. Related observations havebeen made in potato plants; Glc repressed the rbcS geneonly in mature but not in immature leaves (Krapp et al.,1993), suggesting that sugar repression is operative in spe-cific cell types and/or during specific developmentalstages. It remains to be unequivocally determined whetherthese observations were a direct function of sugar concen-trations (metabolic conditions/sugar signaling), related todevelopmental programming, or both. Lest we overlookkey relevant phenomenon, reports of related observationsare offered in the following.
Depending on the developmental stage of the plant orleaf, sugars can modify the level and production of trans-acting proteins that bind to distinct regulatory sequenceson genes encoding key enzymes, leading to the inductionor repression of gene expression. Some upstream regula-tory sequences possess versatile regulation functions, inthat a single gene product can both repress and enhancetranscription of the same gene. For example, some tran-scriptional regulators are known to stimulate or repressexpression of the same gene depending on the physiolog-ical conditions of the cell (Miesfeld et al., 1987; Sakai et al.,1988; Diamond et al., 1990). McKendree and Fed (1992)have also shown that only under tissue culture conditionswas a G-box element of the Arabidopsis adh promoterfunctionally important. Brusslan and Tobin (1992) reported
higher cab transcript levels in Arabidopsis seedlings whenSue was included in the media. However, cab transcriptlevels declined in the presence of sugar in mature plantsand in the protoplasts (transient expression system) (Sheen,1990; Criqui et al., 1992; Krapp et al., 1993). These resultsindicate that DNA sequences (both positive- and negative-response elements) are not the sole determinant of tran-scription regulation. In fact, protein-protein interactionscan influence the regulatory function by causing conforma-tional changes in a DNA-binding protein. Under specificphysiological conditions, a differential effect of a molecularsignal (e.g. sugar) may be manifested in opposite directionsbecause of a different ratio between frans-acting factors,which would then result in stimulation or repression ofgene expression.
The physiological and/or developmental context for theresponse in the two mature leaf systems we had studied(greenhouse-grown versus culture-grown plants) waslikely to have been very different. Therefore, one mightspeculate that some sugar-responsive genes have regula-tory sequences that exhibit dual functions (presumably inconjunction with other factors). Such interpretations arenot, however, possible with our current data. Furthermore,answering such questions is hindered: in addition to lim-ited structural knowledge of sugar-responsive elements formany carbon metabolism genes, it is unlikely that a uni-versal sugar-responsive elements is responsible for a widespectrum of sugar-modulated gene expression in higherplants.
Several conclusions emerged from the present data. First,response to sugars was gene and signal specific (only me-tabolizable sugars were effective). Second, in the context ofend-product regulation, key enzymes of photosynthesisand Sue synthesis are differentially regulated, which mayor may not include regulation at the transcription level.Third, depending on the developmental and/or physiolog-ical stage of the leaf and/or the plant (sink versus source orgreenhouse versus culture-grown), sugars may exert dif-ferent effects on the expression of the same gene. Fourth,enhanced gene expression in the presence of sugars was
100
90
_ 80o~ 70O
* 60•f£ 50
40
H 30
20
10
0
Figure 6. Changes in the level of rbcS (solid bars) and cyt-FBPase(open bars) transcripts in mature leaves of 4-month-old, greenhouse-grown plants. Leaf discs were incubated in solutions containingeither 40 mM sugar or mannitol (control) for 24 h under constant lightconditions. www.plantphysiol.orgon June 24, 2018 - Published by Downloaded from
Copyright © 1995 American Society of Plant Biologists. All rights reserved.

Accelerated Leaf Development 1655
likely to have been due to accelerated leaf development. Collectively, the data suggested that addition of a sugar to the culture media caused accelerated leaf growth, develop- ment, and transition from sink to source. The observed accelerated pace of growth and development was mani- fested a t physiological, biochemical, and molecular levels. Together with other data obtained in our laboratory and by other investigators (mostly f rom nonplant systems), it is suggested that, depending on the physiological and /or developmental status of the plant, sugars may regulate expression of the same gene in different directions.
ACKNOWLEDCMENTS
We wish to thank Alexander Kovtun for technical assistance. We also thank Professor Ray Evert (Botany Department, Univer- sity of Wisconsin-Madison) and Professor Robert Turgeon (Section of Plant Biology, Cornell University) for stimulating discussions and helpful suggestions in the preparation of the manuscript.
Received March 17, 1995; accepted April 27, 1995. Copyright Clearance Center: 0032-0889/95/108/1647/10.
LITERATURE ClTED
Arnon DI (1949) Copper enzymes in isolated chloroplasts, poly- phenol oxidase in Beta vulgaris. Plant Physiol 24: 1-15
Ashwell G (1957) Calorimetric analysis of sugars. Methods Enzy- mo1 3: 73-105
Berry OJ, Nikolau BJ, Carr JP, Klessig DF (1985) Transcriptional and post-transcriptional regulation of ribulose 1:5-bisphosphate carboxylase gene expression in light- and dark-grown Amaranth cotyledons. Mo1 Cell Biol 5 2238-2246
Bradford MM (1976) A rapid and sensitive method for the quan- titation of microgram quantities of protein vitalizing the princi- ple of protein-dye binding. Anal Biochem 72: 248-254
Brusslan JA, Tobin E (1992) Light-independent developmental regulation of cab gene expression in Arabidopsis tkaliana seed- lings. Proc Natl Acad Sci USA 89: 7791-7795
Cheng C-L, Acedo GN, Cristinsin M, Conkling MA (1992) Su- crose mimics the light induction of Arabidopsis nitrate reduc- tase gene transcription. Proc Natl Acad Sci USA 89: 1861-1864
Chomczynski N, Sacchi N (1987) Single step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem 161: 156-159
Chourey PS, Latham MD, Still PE (1986) Expression of two su- crose synthase gene in endosperm and seedlings of maize: evi- dente of tissue-specific polymerization of promoters. Mo1 Gen Genet 203: 251-255
Chourey PS, Taliercio EW (1994) Epistatic interaction and func- tional compensation between the two tissue- and cell-specific sucrose synthase genes in maize. Proc Natl Acad Sci USA 91:
Claussen W, Hawker JS, Loveys BR (1985) Comparative investi- gations on the distribution of sucrose synthase activity and invertase activity within growing mature and old leaves of some 3-carbon photosynthetic pathway and 4-carbon photosynthetic pathway plant species. Physiol Plant 65: 275-280
Claussen W, Loveys BR, Hawker JS (1986) Influence of sucrose and hormones on the activity of sucrose synthase and invertase in detached leaves and leaf sections of eggplants (Solanum mel- ongena). J Plant Physiol 124: 345-357
Criqui M-C, Durr A, Parmentier Y, Marbach J, Fleck J, Jamet E (1992) How are photosynthetic genes repressed in freshly-iso- lated mesophyll protoplasts of Nicotiana syluestris? Plant Physiol Biochem 3 0 597-601
791 7-7921
Daie J (1993) Cytosolic fructose-l,6-bisphosphatase: a key enzyme in the sucrose biosynthetic pathway. Photosynth Res 38: 5-14
Delseny M, Cooke R, Penon P (1983) Sequence heterogeneity in radish nuclear ribosomal RNA genes. Plant Sei Lett 3 0 107-119
Diamond MI, Miner JN, Yoshinaga SK, Yamamoto KR (1990) Transcription factor interactions: selectors of positive or nega- tive regulation from a single DNA element. Science 249 1266- 1272
Dickinson C, Altabella T, Chrispeels MJ (1991) Slow-growth phenotype of transgenic tomato expressing apoplastic invertase. Plant Physiol 95: 420425
Dreesmann DC, Harn C, Daie J (1994) Expression of genes encod- ing Rubisco in sugarbeet (Beta vulgaris L) plants subjected to gradual desiccation. Plant Cell Physiol 3 5 645-653
El-Maghrabi MR, Pilkis J, Marker AJ, Colosia AD, D’angelo G, Fraser BA, Pilkis SJ (1988) cDNA sequence of rat liver fructose- 1,6-bisphosphatase and evidence for down regulation of its RNA by insulin. Proc Natl Acad Sci USA 85: 8430-8434
Entain K-D (1980) Genetic and biochemical evidence for hexoki- nase PII as a key enzyme involved in carbon catabolite repres- sion in yeast. Mo1 Gen Genet 178: 633-637
Friend DJC, Bodson M, Bernier G (1984) Promotion of flowering in Brussica campestris L. cv Ceres by sucrose. Plant Physiol 75:
Galtier N, Foyer CH, Huber J, Voelker TA, Huber SC (1993) Effects of elevated sucrose-phosphate synthase activity on pho- tosynthesis, assimilate partitioning and growth in tomato (Lyco- persicon esculentum var UC82B). Plant Physiol 101: 535-543
Ghosh S, Gepstein S , Glick BR, Heikkila JJ, Dumbroff EB (1989) Thermal regulation of phosphoenol pyruvate carboxylase and ribulose-l,5-bisphosphate carboxylase in C3 and C4 plants na- tive to hot and temperate climates. Plant Physiol 9 0 1298-1304
Giaquinta R (1978) Source and sink leaf metabolism in relation to phloem translocation. Plant Physiol 61: 380-385
Goldschmidt EE, Huber SC (1992) Regulation of photosynthesis by end-product accumulation in leaves of plants storing starch, sucrose and hexose sugars. Plant Physiol 99: 1443-1448
Harn C, Daie J (19924 Cloning and nucleotide sequence of cDNA encoding the cytosolic fructose-1,6-bisphosphatase of sugarbeet (Beta vulgaris L.). Plant Physiol 9 8 790-791
Harn C, Daie J (1992b) Regulation of the cytosolic fructose-1,6- bisphosphatase by post-translational modification and protein leve1 in drought-stressed leaves of sugarbeet. Plant Cell Physiol
Harn C, Khayat E, Daie J (1993) Expression dynamics of genes encoding key carbon metabolism enzymes during sink to source transition of developing leaves. Plant Cell Physiol34 1045-1053
Harter K, Talke-Messerer C, Barz W, Schafer E (1993) Light and sucrose-dependent gene expression in photomixotrophic cell suspension cultures and protoplasts of rape. Plant J 4: 507-516
Huang N, Chandler J, Thomas BR, Koizumi N, Rodriguez RL (1993) Metabolic regulation of a-amylase gene expression in transgenic cell culture of rice (Oriza sutiva L.). Plant Mo1 Biol23: 737-747
Huber JL, Hite DRC, Outlaw WH, Huber SC (1991) Inactivation of highly activated spinach leaf sucrose-phosphate synthase by dephosphorylation. Plant Physiol 9 5 291-297
Huber SC, Huber JL (1992) Role of sucrose-phosphate synthase in sucrose metabolism in leaves. Plant Physiol 99: 1275-1278
Jang J-C, Sheen J (1994) Sugar sensing in higher plants. Plant Cell
Kelly GJ, Zimmermann G, Latzko E (1982) Fructose-bisphos- phatase from spinach leaf chloroplast and cytoplasm. Methods Enzymol90: 271-279
Kossmann J, Muller-Rober B, Dyer TA, Raines CA, Sonnewald U, Willmitzer L (1992) Cloning and expression analysis of plas- tidic fructose-1,6-bisphosphatase coding sequence from potato: circumstantial evidence for import of hexoses into chloroplasts. Planta 188: 7-12
Krapp A, Hofmann B, Schaefer C, Stitt M (1993) Regulation of the expression of rbcS and other photosynthetic genes by carbohy- drates: a mechanism for the ”sink regulation” of photosynthesis? Plant J 3: 817-828
1085-1089
33: 763-770
6 1665-1679
www.plantphysiol.orgon June 24, 2018 - Published by Downloaded from Copyright © 1995 American Society of Plant Biologists. All rights reserved.

Krapp A, Quick WP, Stitt M (1991) Rubulose-1,5-bisphosphate carboxylase-oxygenase, other photosynthetic enzymes and chlo- rophyll decrease when glucose is supplied to mature spinach leaves via transpiration stream. Planta 186: 58-69
Lee AS (1987) Coordinated regulation of a set of genes by glucose and calcium ionophores in mammalian cells. Trends Biol Sci 12
Lenz AG, Holzer H (1980) Rapid reversible inactivation of fruc- tose-1,6-bisphosphatase in Saccharomyces cerevisiae by glucose. FEBS Lett 1 0 9 271-274
Maas C, Schaal S, Werr W (1990) A feedback control element near the transcription start site of the maize Shrunken gene deter- mines promoter activity. EMBO J 9: 3447-3452
McKendree WL, Ferl RJ (1992) Functional elements of Arubidopsis Adh promoter include the G-box. Plant Mo1 Biol 1 9 859-862
Miesfeld R, Godowski PJ, Maler BA, Yamamoto KR (1987) Glu- cocorticoid receptor mutants that define a small region sufficient for enhancer activation. Science 326: 423428
More11 M, Copeland L (1985) Sucrose synthase of soybean nod- ules. Plant Physiol 78: 149-154
Muller-Rober BT, Kossmann J, Hannah LC, Willmitzer L, Son- newald U (1990) One of two different ADP-glucose pyrophos- phorylase genes from potato responds strongly to elevated lev- els of sucrose. Mo1 Gen Genet 224 136-146
Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco tissue cultures. Plant Physiol 15: 473-497
Nguyen-Quoc B, Krivitzky M, Huber SC, Lecharny A (1990) Sucrose synthase in developing maize leaves. Plant Physiol 94:
Pitcher LH, Daie J (1991) Growth and sink to source transition in developing leaves of sugarbeet. Plant Cell Physiol 3 2 335-342
Sakai DD, Helms S, Carlstedt-Duke J, Gustafsson J-A, Rottman FM, Yamamoto KR (1988) Hormone-mediated repression: a negative glucocorticoid response element from the bovine pro- lactin gene. Genes Dev 2 1144-1154
Salanoubat M, Belliard G (1989) The steady-state level of potato sucrose synthase mRNA is dependent on wounding, anaerobi- osis and sucrose concentration. Gene 84: 181-185
Sheen J (1990) Metabolic repression of transcription in higher plants. Plant Cell 2: 1027-1038
Sheen J (1994) Feedback control of gene expression. Photosynth Res 39: 428438
Stitt M (1990) The flux of carbon between the chloroplast and
20-23
516-523
1656 Kovtun and Daie Plant Physiol. Vol. 108, 1995
cytoplasm. In DT Dennis, DH Turpin, eds, Plant Physiology, Biochemistry and Molecular Biology. Longman Scientific, New York, pp 308-326
Stitt M, Quick WP (1989) Photosynthetic carbon partitioning: its regulation and possibility for manipulation. Physiol Plant 77:
Stitt M, Von Schaewen A, Willmitzer L (1991) “Sink regulation of photosynthetic metabolism in transgenic tobacco plants ex- pressing yeast invertase in their cell wall involves a decrease of the Calvin cycle enzymes and an increase of glycolytic enzymes. Planta 183: 40-50
Takeda S, Mano S , Ohto M, Nakamura K (1994) Inhibitors of protein phosphatases 1 and 2A block the sugar-inducible gene expression in plants. Plant Physiol 106: 567-574
Tsukaya H, Ohshima T, Naito S, Chino M, Komeda Y (1991) Sugar-dependent expression of the CHS-A gene for chalcone synthase from petunia in transgenic Arabidopsis. Plant Physiol
Turgeon R (1989) The sink-source transition in leaves. Annu Rev Plant Physiol Plant Mo1 Biol 40: 119-138
Van der Steege G, Nieboer M, Swaving J, Tempelaar MJ (1992) Potato granule-bound starch synthase promoter-controlled GUS expression: regulation of expression after transient and stable transformation. Plant Mo1 Biol 20: 19-30
Von Schaewen A, Stitt M, Schmidt R, Sonnewald U, Willmitzer L (1990) Expression of a yeast-derived invertase in the cell wall of tobacco and Arabidopsis plants leads to accumulation of carbohydrate and inhibition of photosynthesis and strongly in- fluentes growth and phenotype of transgenic tobacco plants.
Walker JL, Huber SC (1989) Regulation of sucrose-phosphate synthase activity in spinach leaves by protein level and covalent modification. Planta 177: 116-120
Wang J-L, Turgeon R, Carr JP, Berry JO (1993) Carbon sink-to- source transition is coordinated with establishment of cell-spe- cific gene expression in a C4 plant. Plant Cell 5 289-296
Wenzler HC, Mignery GA, Fisher LM, Park WD (1989) Analysis of a chimeric class-1 patatin-GUS gene in transgenic potato plants:
in cultured leaf and stem explants. Plant Mo1 Biol 12: 41-50 Worrell AC, Bruneau JM, Summerfelt K, Boesig M, Voelker TA
(1991) Expression of a maize sucrose phosphate synthase in toma- to alters leaf carbohydrate partitioning. Plant Cell 3: 1121-1131
633-641
97: 1414-1421
EMBO J 9: 3033-3044
high-leve1 expression in tubers and sucrose-inducible expression
www.plantphysiol.orgon June 24, 2018 - Published by Downloaded from Copyright © 1995 American Society of Plant Biologists. All rights reserved.





![Phytochrome, Carbon Sensing, Metabolism, and …...Phytochrome, Carbon Sensing, Metabolism, and Plant Growth Plasticity1[CC-BY] Johanna Krahmer, Ashwin Ganpudi, Ammad Abbas, Andrés](https://img.pdfslide.net/doc/110x75/5e52046a46d25750920125b7/phytochrome-carbon-sensing-metabolism-and-phytochrome-carbon-sensing-metabolism.jpg)























