Embed Size (px)
Citation preview

Enhanced Survival and MorphologicalFeatures of Basal Forebrain CholinergicNeurons In Vitro: Role of Neurotrophinsand Other Potential Cortically Derived
Cholinergic Trophic Factors
DUN H. HA,1 RICHARD T. ROBERTSON,1 MOHSEN ROSHANAEI,1
AND JOHN H. WEISS1,2,3*1Department of Anatomy and Neurobiology, University of California,
Irvine, Irvine, California 92697–42922Department of Neurology, University of California, Irvine, Irvine, California 92697–4292
3Department of Psychobiology, University of California,Irvine, Irvine, California 92697–4292
ABSTRACTThe present study examined survival- and growth-enhancing effects of cortical cells on
basal forebrain cholinergic neurons (BFCNs) in culture and the degree to which endogenousnerve growth factor (NGF), brain-derived neurotrophic factor (BDNF), and neurotrophin-3(NT-3) contribute to those trophic effects. When fetal (17 days of gestation) basal forebrain(BF) cells were grown for 5 days in coculture with cortical neurons, staining for acetylcholines-terase (AChE) showed a threefold increase in the number of BFCNs relative to BF cultureswithout cortex. Most of these labeled cells also displayed enhanced somatic, dendritic, andaxonal growth. Coculturing cortical neurons with BF cells taken from postnatal animalsproduced similar results but with a somewhat greater degree of morphologic enhancement.Function-neutralizing antibodies to NGF, BDNF, and NT-3 were employed to determinewhether they would block the trophic effects of cortical neurons on postnatal BFCNs.Although no significant changes in numbers or morphological features of AChE(1) neuronswere observed with treatment with individual antibodies, cocultures treated with a combina-tion of all three antibodies displayed fewer morphologically enhanced AChE(1) cells and morenonenhanced cells; the total number of AChE(1) neurons was not significantly changed.Treatment of pure BF cultures with exogenous NGF, BDNF, and NT-3 increased the number ofAChE(1) neurons but did not reproduce the morphologic enhancement of cortical cells onBFCNs. These results suggest that neurotrophins by themselves can increase survival ofpostnatal BFCNs in culture and may work in concert with other unknown cortically derivedfactors to enhance BFCN morphologic differentiation. The unidentified cortical factors mayalso have strong survival-enhancing effects on BFCNs that are independent of the knownneurotrophins. J. Comp. Neurol. 406:156–170, 1999. r 1999 Wiley-Liss, Inc.
Indexing terms: antibody neutralization; cell culture; coculture; acetylcholinesterase; choline
acetyltransferase
Basal forebrain cholinergic neurons (BFCNs) providethe major subcortical source of cholinergic innervation ofneocortex. The innervation is extensive, with evidence ofingrowth of cholinergic axons into all neocortical regionsand layers (Eckenstein et al., 1988; Lysakowski et al.,1989; Calarco and Robertson, 1995). Similar patterns ofbasal forebrain-derived cholinergic axonal ingrowth to
Grant sponsor: NIH; Grant numbers: NS 30884 and NS 30109; Grantsponsor: Pew Scholars Program in the Biomedical Sciences; Grant sponsor:NRSA; Grant number: MH 11483.
*Correspondence to: Dr. John H. Weiss, Department of Neurology,University of California, Irvine, Irvine, CA 92697–4292.E-mail: [email protected]
Received 5 August 1998; Revised 15 October 1998; Accepted 1 November1998
THE JOURNAL OF COMPARATIVE NEUROLOGY 406:156–170 (1999)
r 1999 WILEY-LISS, INC.

neocortex have been demonstrated by using in vitro sys-tems. Organotypic cocultures containing both basal fore-brain (BF) and neocortical tissues have shown prominentgrowth of BF-derived cholinergic axons into the neocorti-cal tissue (Distler and Robertson, 1992, 1993; Baratta etal., 1996). Further, BFCNs in dissociated cocultures withneocortical cells send axons that appear to specificallytarget groups of cortical neurons and to form synapseswith the individual cortical neurons in those groups (Ha etal., 1996). The BFCNs in these cocultures also displayedenhanced somatic, dendritic, and axonal growth, presum-ably induced by the cortical neurons that they innervate.Thus, the differentiation of BFCNs appears to be regu-lated, at least in part, by neocortical cells.
Mechanisms by which neocortical neurons can regulatethe phenotypic expression of BFCNs during developmentare largely unknown, but cortically derived neurotrophins,including nerve growth factor (NGF), brain-derived neuro-trophic factor (BDNF), and neurotrophin-3 (NT-3), mayplay a role. BFCNs express tyrosine-kinase receptors forNGF (Trk-A), BDNF and NT-4/5 (Trk-B), NT-3 (Trk-C), andthe p75 low-affinity neurotrophin receptor, which is recog-nized by all the neurotrophins (Klein et al., 1990; Koh andHiggins, 1991; Barbacid, 1994; Kordower et al., 1994;Lamballe et al., 1994; Sobreviela et al., 1994). Moreover,trophic effects of these factors on BFCNs have beendemonstrated by both in vivo and in vitro studies (Hefti,1986; Hartikka and Hefti, 1988; Alderson et al., 1990;Friedman et al., 1993; Morse et al., 1993; Svendsen et al.,1994; Nonomura et al., 1995; Ha et al., 1996; Kew et al.,1996).
The studies noted above suggest an important role forneurotrophins in BFCN development. However, studies oftransgenic mice lacking the NGF, BDNF, or NT-3 genehave not demonstrated reductions in numbers of choliner-gic neurons in the basal forebrain or significant changes inthe extent of cholinergic innervation of target regions(Crowley et al., 1994; Ernfors et al., 1994; Jones et al.,1994). Organotypic cocultures prepared with brain tissuesderived from homozygote NGF-knockout mice producedsimilar findings, with significant BFCN axonal growth anddense innervation of neocortical tissue (Baratta et al.,1997). These data could be interpreted as indicating thatthe neurotrophins are not essential for the normal develop-ment of BFCNs. Alternatively, there may be redundancy inthe effects of neurotrophins on BFCNs, with the absence ofone compensated for by others. To examine this possibility,one must be able to remove not one but several of theneurotrophins from the system to determine the degree towhich the neurotrophins contribute to the development ofBFCNs.
In a previous study, we showed that the presence ofneocortical neurons enhanced morphological features andincreased survival of BFCNs in dissociated cell cultures(Ha et al., 1996). The present study sought further tocharacterize the survival- and growth-enhancing effects ofneocortical neurons on BFCNs and to use exogenousneurotrophins and neurotrophin-neutralizing antibodiesto examine the role of the neurotrophins in these effects.Also, because axons of BFCNs normally contact corticalcells during the early postnatal period in vivo (Dinopouloset al., 1989; Calarco and Robertson, 1995; De Carlos et al.,1995), the study also examined the degree to which thesecortically derived effects depend on the maturational stateof either the cortical or BF cells.
MATERIALS AND METHODS
Animals
Timed-pregnant Sprague-Dawley rats were purchasedfrom Simonsen Laboratories (Santa Clara, CA) and housedin a vivarium. Pregnant dams were deeply anesthetizedwith sodium pentobarbital before their fetuses were re-moved. Postnatal rat pups were initially cryoanesthetizedand then decapitated. All use of animals was conducted inaccordance with the National Institute of Health Guide forthe Care and Use of Laboratory Animals and was approvedby the University of California Irvine Institutional AnimalCare and Use Committee.
Cell culture preparation
Neuronal tissue from fetal and postnatal animals wasused in the present study. Fetuses were removed fromtimed-pregnant rats at gestational age 17 (G-17) and theirbrains were removed and collected in cold medium. Menin-ges surrounding the brains were removed, and the corticalhemispheres were spread laterally to expose the BF re-gion. By using techniques described previously (Ha et al.,1996; Hartikka and Hefti, 1988), the neocortex and BFregions were then carefully dissected and placed in sepa-rate dishes containing fresh medium. To obtain postnatal(P) BF neurons, young rat pups (P-1 or P-7) were killed andtheir brains removed and sectioned on a Campden Vi-braslicer. Transverse slices containing the BF were se-lected, and the region containing the medial septum anddiagonal band of Broca was dissected under a stereomicro-scope and placed in a dish of fresh medium.
After dissections, the selected brain tissues were mincedand enzymatically dissociated with trypsin (30 minutes at37°C). Additional dissociation was accomplished by tritu-rating with sterile Pasteur pipettes (large and then smallbore). The resulting cell suspensions were diluted in aplating medium consisting of Eagle’s Minimal EssentialMedium (MEM–Earles salts, supplied without glutamine)supplemented with 10% heat-inactivated horse serum,10% fetal bovine serum, glutamine (2 mM), and glucose(total 25 mM). The neurons were plated (1.0–2.0 3 105
cells/cm2) on previously established monolayers of corticalastrocytes (see below) in 24-well tissue culture plates andmaintained in a 37°C/5% CO2 incubator. For experimentsthat extended beyond a week, the cultures were treatedwith 1025 M cytosine arabinoside to reduce nonneuronalcell proliferation.
Pure BF cultures were produced by plating with G-17,P-1, or P-7 BF cells. Cultures receiving postnatal neuronswere also treated with 1025 M cytosine arabinoside on theday of plating to prevent glial proliferation. Pure corticalcultures were produced by plating G-17 cortical cells. Twotypes of cocultures were prepared. For the first type, G-17BF and G-17 cortical cells were plated together and grownfor 5 days. The second type of coculture was prepared byfirst plating and growing G-17 cortical cells alone for 5 or12 days in vitro, followed by the addition of BF cells (G-17,P-1, or P-7) for an additional 5 days. In experiments inwhich BFCNs in different types of cultures were compared,care was taken to ensure that the same amount of BF cellsuspension was added to all cultures. Cocultures receivingpostnatal BF cells were also administered 1025 M cytosinearabinoside.
Monolayer astrocyte cultures were prepared by platingP-1 cortical cells directly on Falcon Primaria culture plates
TROPHIC EFFECTS OF CORTICAL CELLS ON BFCNS 157

in medium supplemented with epidermal growth factor(10 ng/ml). These cultures contained virtually no neurons.When a confluent monolayer of astrocytes was established(about 10 days), the cultures were used to plate neurons,as described above.
Immunocytochemistry
Cultures were fixed for 45 minutes in 4% paraformalde-hyde followed by three washes with phosphate bufferedsaline (PBS; pH 7.4). The cultures were then incubated ina blocking solution consisting of 10% horse serum (forstaining with choline acetyltransferase; ChAT) or 5% goatserum (for p75 staining) in PBS for 1 hour at 25°C.Primary antibodies (Chemicon, Temecula, CA) to ChAT(1:2,000, made in mouse) or to p75 (1:5,000, made inrabbit) were diluted in PBS and added to the cultures forincubation overnight at 4°C. After three PBS washes toremove the primary antibodies, the cultures were incu-bated in secondary antibodies (anti-mouse for ChAT stain-ing or anti-rabbit for p75 staining) conjugated to thefluorescent marker Cy3 (1:200; Jackson ImmunoresearchLaboratories, West Grove, PA) for 1 hour at 25°C. Fordouble labeling, the ChAT- or p75-labeled cells were photo-graphed under fluorescent microscopy (with a Cy3 opticalfilter: excitation 510–560 nm, emission .590 nm) prior toacetylcholinesterase (AChE) histochemistry, as describedbelow.
Acetylcholinesterase histochemistry
This procedure was carried out largely as described byTago et al. (1986), with minor modifications. Cultures werefixed for 45 minutes in 4% paraformaldehyde and thenwashed three times with 0.1 M maleate buffer (pH 6.0). Insome cases, cultures had been processed previously forimmunocytochemistry. The cultures were then incubatedfor 1–2 hours in the dark in a fresh solution consisting of300 µM copper sulfate, 500 µM sodium citrate, 50 µMpotassium ferricyanide, and 30 µM acetylthiocholine io-dide in 0.1 M maleate buffer. After rinsing with PBS (fivetimes), the reaction product was developed with an intensi-fication solution consisting of 0.04% 3,3’diaminobenzidinetetrahydrochloride, 0.3% nickel ammonium sulfate, and0.003% H2O2 in 0.1 M PBS until cells were clearly stained(15–30 minutes).
Neurotrophin neutralization
In an attempt to block enhancing effects of corticalneurons on BFCNs, cocultures were treated with antibod-ies to NGF (50–100 ng/ml), to BDNF (10 µl/1–2 ml), or toNT-3 (10 µl/1–2 ml) on the first, second, and fourth days ofa 5-day coculturing period. Some cocultures were treatedwith a cocktail of all three antibodies at the indicatedconcentrations and times. Control cocultures were un-treated or treated with control turkey serum (10 µl/1–2ml). The monoclonal antibody used to neutralize NGF(clone 72/21; Boehringer Mannheim, Indianapolis, IN)reacts with both the 2.5S and the 7S forms; its efficacy (100ng/ml of antibody completely blocked ChAT activity in-duced by 100 ng/ml of NGF) and specificity for NGF hasbeen demonstrated in vitro and in vivo systems (Rohrer etal., 1988; Nonner and Barrett, 1994; Svendsen et al.,1994). In the present culture system, this antibody hasbeen shown to induce maximal loss of ChAT(1) neurons ata concentration of 50–100 ng/ml (Ha et al., 1996). Theantibodies to BDNF and NT-3 (a gift from Dr. Carnahan of
Amgen, Thousand Oaks, CA) were raised in turkeys andused in the form of turkey antisera to BDNF or NT-3.Specificity of the antisera was demonstrated by proteinimmunoblots with recombinant neurotrophins; antisera toBDNF or NT-3 did not cross react with the other neuro-trophins (Ghosh et al., 1994). Survival assays indicatedthat the antibodies produced effective block of BDNF- orNT-3-induced enhanced survival of dorsal root ganglioncells at 10 µl/ml (Ghosh et al., 1994).
Exogenous neurotrophin exposure
To determine the effects of neurotrophins on BFCNs,cocultures were treated with exogenous NGF (BoehringerMannheim), BDNF (Promega, Madison, WI), and NT-3(Alomone Labs, Jerusalem, Israel), alone or in combina-tion, each at a concentration of 50–100 ng/ml. The factorswere added directly to the cultures on the first, second, andfourth days of a 5-day coculturing period.
Data presentation and analysis
Illustrations were prepared by scanning photographicnegatives. Figures were prepared from imported images(unchanged except for minor adjustment of brightness orcontrast) by using Photoshop software (Adobe) and printedwith a photo-quality inkjet printer.
For cell counts, AChE(1) neurons in 52 consecutive,nonoverlapping microscope fields, covering over 95% ofeach well, were counted by using low power (1003) bright-field optics. Cells were considered as AChE(1) if they wereclearly stained and displayed at least two processes. Ingeneral, the number of AChE(1) neurons in a controlculture well was 50–100. The mean number of AChE(1)neurons in different treated cocultures was normalized ineach experiment to that found in untreated controls (de-fined as 100%). The means of individual experiments (n 53–4) were then averaged to give a final mean.
For morphologic measurements, AChE(1) neurons wererandomly selected, and two-dimensional images of thesecells were imported into a computer for morphometricanalysis by using COMOS software from Bio-Rad (Rich-mond, CA). Cells (n 5 40–100) were imaged from three tofour sister cultures from each of three platings for eachcondition. Three parameters were measured: soma area,total dendritic length, and number of first-, second-, andthird-order dendrites. Also, those AChE(1) cells withdetectable axons were counted, and the total length oftheir axons was measured.
Values are presented as the mean 6 standard error ofthe mean (SEM), normalized to control conditions in eachexperiment, or as relative distributions. Significance of thedata was determined by analysis of variance, with theBonferoni post hoc test by using Instat software (GraphPad Inc., San Diego, CA).
RESULTS
Morphologic enhancement of fetal AChE(1)neurons by cortical cells
Initial experiments examined effects of cortical neuronsat different states of maturation on the development offetal BFCNs in culture. BF cells from fetuses at G-17 werecocultured for 5 days with cortical neurons from the sameplating (5CB cocultures) or with cortical neurons that had
158 D.H. HA ET AL.

been previously grown in culture for 5 (5C-5CB cocultures)or 12 (12C-5CB cocultures) days. Control cultures platedwith only BF cells (5B cultures) were also prepared.Staining for AChE indicated that the labeled neurons inthe 5B cultures appeared relatively small, had shortneurites, and no detectable axons (Fig. 1A). In contrast,many of the AChE(1) neurons found in the cocultures had
larger cell bodies, longer dendrites, and prominent axons(Fig. 1B–D). Further, AChE(1) neurons in the 5C-5CB and12C-5CB cocultures appeared larger and with more exten-sive dendrites than did those found in the 5CB cocultures.Distinct AChE-stained neurons were not present in cul-tures containing only cortical neurons grown for 5, 10, or17 days (Fig. 1E–G).
Fig. 1. Older cortical cells induce greater morphologic enhance-ment of fetal acetylcholinesterase [AChE(1)] neurons. Photomicro-graphs show representative AChE-stained fields from a pure basalforebrain (17 days of gestation; G-17) culture grown for 5 days (A, 5Bcultures), from pure cortical cultures grown for 5 days (E), 10 days(F), or 17 days (G), and from cocultures (B, 5CB; C, 5C-5CB;
D, 12C-5CB) identical to those in E, F, and G, respectively, except thatG-17 BF cells were added for the last 5 days. Note the presence ofmorphologically enhanced AChE(1) neurons only in cocultures; alsonote the larger AChE(1) neurons in cocultures where cortical neuronswere grown previously for a least 5 days. Arrows in A indicate typicalunenhanced AChE(1) neurons. Scale bar 5 200 µm.
TROPHIC EFFECTS OF CORTICAL CELLS ON BFCNS 159

Cell counts indicated an approximately threefold in-crease in the number of AChE(1) neurons in the cocul-tures compared with control 5B cultures; no differenceswere detected in numbers between the different types ofcocultures (Fig. 2A). Consistent with the qualitative obser-vations, measurements of randomly selected AChE(1)neurons indicated a significant enhancement of soma size,
dendritic length, and dendritic branching of most of theAChE(1) neurons found in the cocultures (Fig. 3A–C).Quantitative analysis also showed differences between thedifferent coculture types; enhanced AChE(1) neurons inthe 5C-5CB and 12C-5CB cocultures displayed greatersomatic and dendritic growth than did those in the 5CBcocultures (Fig. 3A–C). The greatest difference was in axonmeasurements, as indicated by the dramatic increase inboth the number of AChE(1) neurons displaying visibleaxons (identified as a neurite at least 1,000 µm in length)and in the length of the axons of AChE(1) neurons in5C-5CB and 12C-5CB co-cultures (Fig. 3D).
Morphologic enhancement of postnatalAChE(1) neurons by cortical cells
Although these experiments indicate morphologic en-hancement of fetal BFCNs grown in the presence ofcortical cells, it is during the early postnatal period thatBFCNs form most connections with cortical cells in vivo,and it is at this time when they are likely to be mostresponsive to cortical trophic signals. Thus, to determinewhether BFCNs from early postnatal animals are moreresponsive than fetal BFCNs to cortical influences, BFneurons derived from postnatal ages were added to corticalcultures that were established 5 days previously; the re-sulting cocultures were then grown for an additional 5 days.
In cocultures prepared with P-1 BF cells (5C-5CB-P1cocultures), AChE staining showed the presence of twotypes of labeled cells that could be distinguished on thecriterion of the presence or absence of AChE-stained axons(axons were identified as neurites at least 1,000 µm long).Those AChE(1) neurons with axons (referred to as type Icells) were more numerous and also displayed enhancedmorphological features, including relatively large cell bod-ies and elaborate dendritic arborization (Fig. 4A,B). Incontrast, the second type of AChE(1) neurons (type II)were fewer in number, appeared much smaller, and hadshorter and less branched dendrites (Fig. 4C). When P-7BF cells were cocultured with cortical neurons underidentical conditions, type I AChE(1) neurons were alsodetected (Fig. 3E,F). Control cultures containing only P-1(5BF-P1) or P-7 BF cells contained AChE(1) neurons thatwere only of the small type (Fig. 4D,G).
Cell counts demonstrated an approximately threefoldincrease in the total number of AChE(1) neurons in5C-5CB-P1 cocultures compared with control 5B-P1 cul-tures; the increase in total number was attributed mainlyto an increase in the number of type I neurons (Fig. 2B).That is, quantitative analysis of AChE(1) neurons in the5C-5CB-P1 cocultures indicated that about 78% of thelabeled cells had detectable axons (type I), averaging about6,000 µm in total length (Fig. 5A); the remaining 22% (typeII) did not show AChE-stained axons. Morphometric analy-sis indicated that most of the type I AChE(1) neuronsdisplayed larger cell size, longer dendrites, and greaterdendritic branching than did the type II cells found in thesame cocultures (Fig. 5B–D). Thus, the presence of aprominent axon appeared to be a single feature that coulddistinguish virtually all morphologically enhanced fromnonenhanced AChE(1) neurons.
Type I AChE(1) neurons also express p75and choline acetyltransferase
Although AChE has been demonstrated to be a goodmarker for BFCNs, some cortical neurons also express this
Fig. 2. Presence of cortical cells increases the number of acetylcho-linesterase [AChE(1)] neurons in cocultures. A: Numbers of AChE(1)neurons in cocultures prepared from gestational day 17 basal fore-brain (BF) neurons and cortical neurons were counted and comparedwith numbers present in control pure BF cultures. The 5CB, 5C-5CB,and 12C-5CB cocultures differ only in the age in vitro of the corticalneurons with which the BF cells were plated. B: Postnatal day 1 BFcells were plated into 5-day-old cortical cultures, and the resultingcocultures (5C-5CB-P1) were grown for 5 additional days. Thesecocultures contained two types of AChE(1) neurons: those withapparent, long (.1,000 µm) axons (type I) and those without detectableaxons (type II). The number of each type was counted and compared withthe total number in pure BF cultures (5B-P1) grown for 5 days. Data arepresented as mean 6 SEM (n 5 3 experiments; total of 10–15 cultures percondition). Asterisk indicates significant differences from all other condi-tions, P , 0.05. Number sign indicates significant differences from 5B-P1cultures, P , 0.001 (analysis of variance with Bonferonni’s post hoc test).
160 D.H. HA ET AL.

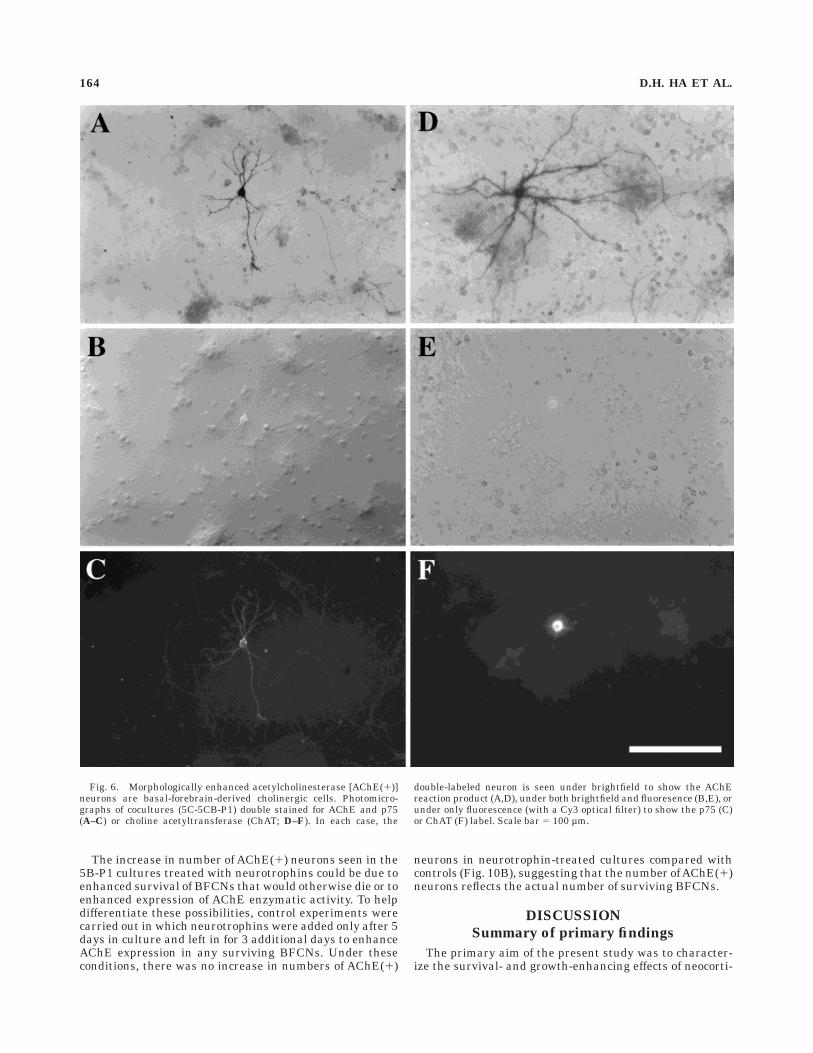
enzyme (Zhu et al., 1984; Hallanger et al., 1986; Mesulamand Geula, 1991; Robertson et al., 1988). To determinewhether the morphologically enhanced AChE(1) neuronsare BFCNs, 5C-5CB-P1 cocultures were double labeled forAChE and the enzyme ChAT or the p75 low-affinityneurotrophin receptor (Hartikka and Hefti, 1988; Koh andHiggins, 1991; Kiss et al., 1993). Of the 80 type I AChE(1)neurons (from three experiments) examined for evidenceof p75 staining, almost all (79 cells) were found to bedouble labeled (Fig. 6A–C). Similarly, all of the 80 type IAChE(1) cells examined also displayed ChAT staining(Fig. 6D–F). Whereas the ChAT labeling was confined tothe cell body and proximal processes, the p75 stainingappeared to be more extensive, with the entire dendritictree and the axon labeled (Fig. 6). Thus, the expression ofall three markers (AChE, ChAT, and p75) by type I cellsindicates that these cells represent a population of BF-derived cholinergic neurons.
Neurotrophin neutralization partly blocksthe morphologic enhancement
of AChE(1) neurons
The studies reported above indicate that cortical cellscan induce trophic effects on BFCNs. The next set ofexperiments sought to investigate possible mechanisms.Function-blocking antibodies to NGF, BDNF, and NT-3were used to determine whether neurotrophin neutraliza-tion would block the ability of cortical cells to enhancemorphological features of AChE(1) neurons. In 5C-5CB-P1 cocultures treated with individual antibodies toNGF, BDNF, or NT-3, the percentage of total AChE(1)neurons that were morphologically enhanced (type I cells;about 75% in each case) was not significantly differentfrom that of untreated controls or from cocultures thatwere treated with control serum (Fig. 7A–E). In coculturestreated with all three antibodies, however, type II AChE(1)
Fig. 3. Older cortical cells induce greater morphologic enhance-ment of fetal acetylcholinesterase [AChE(1)] neurons: quantitativeassessment. AChE(1) neurons from pure basal forebrain cultures (5B)and cocultures (5CB, 5C-5CB, and 12C-5CB) were randomly selected,imaged, and measured for soma area (A), length of dendritic growth(B), and extent of dendritic branching (as indicated by the meannumber of primary, secondary, and tertiary dendrites of all the cellsexamined; C). In cocultures, the fraction of all AChE(1) neurons
examined that had detectable axons was determined and the length oftheir axons measured (D). Data for each parameter measured arepresented both graphically (as histograms) and as means 6 SEM (n 580–100 cells compiled from three experiments). Asterisk indicatessignificant differences from other conditions within each branchingclassification, P , 0.001 for all significant differences (analysis ofvariance with Bonferonni’s post hoc test).
TROPHIC EFFECTS OF CORTICAL CELLS ON BFCNS 161

Fig. 4. Cortical neurons induce morphologic enhancement of post-natal acetylcholinesterase [AChE(1)] neurons in cocultures. Photomi-crographs show representative AChE(1) neurons in cocultures, inwhich postnatal day 1 (P-1) basal forebrain (BF) cells (A–C) or P-7 BFcells (E,F) were plated into 5-day-old cortical cultures and grown for
an additional 5 days. Control cultures, also grown for 5 days, con-tained only P-1 (D) or P-7 (G) BF cells. Both morphologically enhanced(A,B) and nonenhanced (C) AChE(1) neurons are found in the samecoculture prepared with P-1 BF cells. Scale bar 5 100 µm inA,C,D,E,G, 300 µ in B,F.
162 D.H. HA ET AL.

neurons outnumbered type I neurons (Figs. 7F, 8A). Thechange in the relative proportion of type I and II AChE(1)neurons reflected an increase in the number of type II (by204.5 6 54.1%, mean 6 S.E.M., P , 0.01) and decrease inthe number of type I (by 57.5 6 3.9%, P , 0.01) AChE(1)neurons (Fig. 8B), with relatively little change in the totalnumber of AChE(1) neurons (Fig. 8C).
Absence of type I AChE(1) neurons in pureBF cultures treated with exogenous
neurotrophins
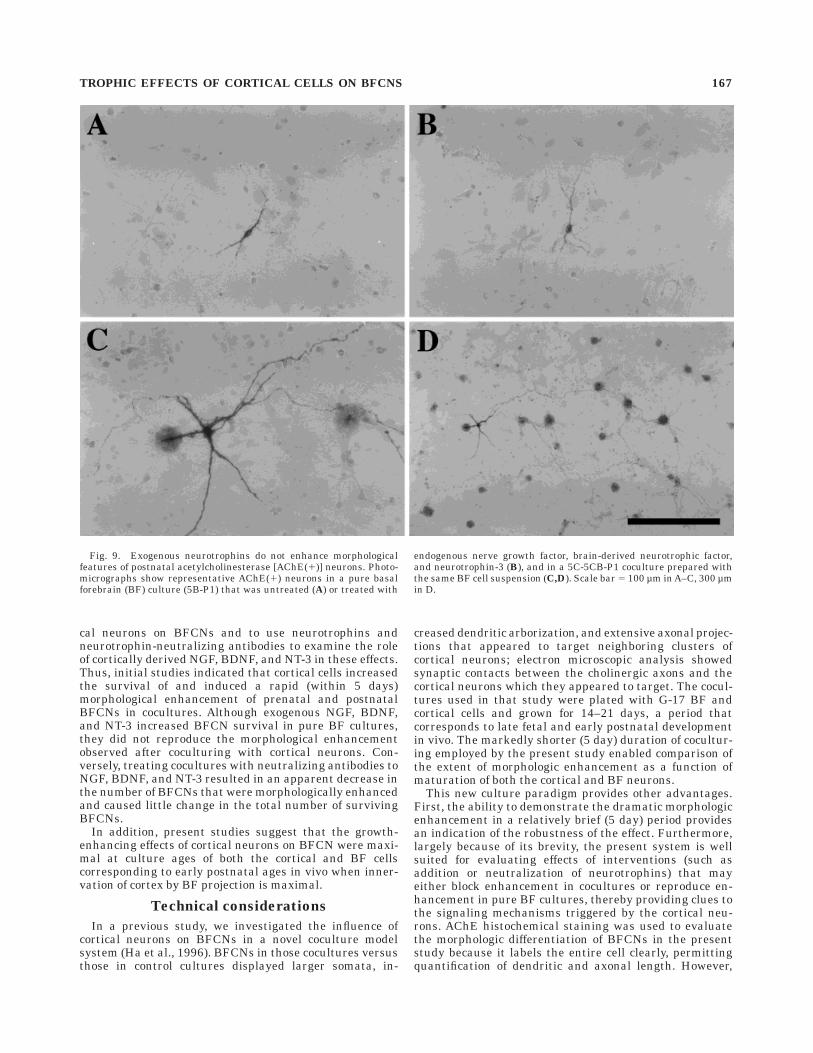
In subsequent experiments, exogenous neurotrophinswere added to pure BF (5B-P1) cultures to test whetherthey could reproduce the trophic effects of cortical neuronson BFCNs. Treating 5B-P1 cultures with a cocktail of NGF,BDNF, and NT-3 for 5 days produced only nonenhanced
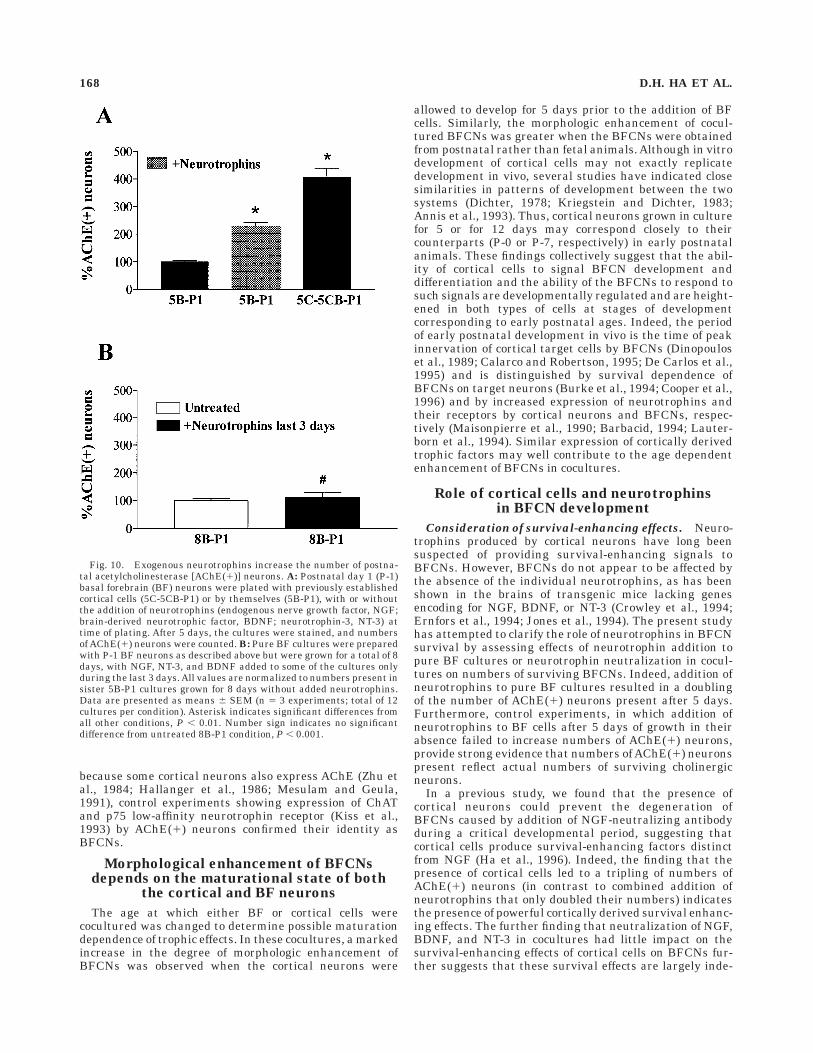
(type II) AChE(1) neurons that appeared morphologicallysimilar to those found in the untreated control cultures(Fig. 9A,B). Similar effects were observed in 5B-P1 cul-tures treated with the individual neurotrophins (data notshown). Although the exogenous neurotrophins did notreproduce the dramatic morphologic enhancement ofBFCNs induced by cortical neurons, cell counts indicatedabout a twofold increase in the total number of AChE(1)neurons compared with untreated controls (Fig. 10A).When BF cells from the same dissection were plated withcortical neurons (as 5C-5CB-P1 cocultures), subsequentstaining showed the presence of numerous type I AChE(1)neurons (Fig. 9C,D) and a threefold increase in the totalnumber of AChE(1) cells (Fig. 10A), indicating that the BFneurons were capable of responding to the cortical influ-ences.
Fig. 5. Cortical neurons induce morphologic enhancement of post-natal acetylcholinesterase [AChE(1)] neurons in cocultures: quantita-tive assessment. AChE(1) neurons from pure basal forebrain (BF)cultures and cocultures prepared with postnatal day 1 BF cells (5B-P1and 5C-5CB-P1 cultures, respectively), were randomly selected, im-aged, and measured for axon length (A), soma area (B), extent ofdendritic branching (as indicated by the mean number of primary,secondary, and tertiary dendrites of all the cells examined; C), andtotal length of dendritic growth (D). Because type I (with distinct
axon . 1,000 µm) and type II (lacking detectable axons) AChE(1)neurons found in the cocultures could be readily distinguished byvisual assessment; values for these two groups of neurons are graphedseparately. Data for each parameter measured are presented bothgraphically (as histograms) and as means 6 SEM (n 5 40–50 cellscompiled from three experiments). Asterisk indicates significant differ-ences from all other conditions within each branching classification inC; P , 0.001 for all significant differences (analysis of variance withBonferonni’s post hoc test).
TROPHIC EFFECTS OF CORTICAL CELLS ON BFCNS 163
The increase in number of AChE(1) neurons seen in the5B-P1 cultures treated with neurotrophins could be due toenhanced survival of BFCNs that would otherwise die or toenhanced expression of AChE enzymatic activity. To helpdifferentiate these possibilities, control experiments werecarried out in which neurotrophins were added only after 5days in culture and left in for 3 additional days to enhanceAChE expression in any surviving BFCNs. Under theseconditions, there was no increase in numbers of AChE(1)
neurons in neurotrophin-treated cultures compared withcontrols (Fig. 10B), suggesting that the number ofAChE(1)neurons reflects the actual number of surviving BFCNs.
DISCUSSION
Summary of primary findings
The primary aim of the present study was to character-ize the survival- and growth-enhancing effects of neocorti-
Fig. 6. Morphologically enhanced acetylcholinesterase [AChE(1)]neurons are basal-forebrain-derived cholinergic cells. Photomicro-graphs of cocultures (5C-5CB-P1) double stained for AChE and p75(A–C) or choline acetyltransferase (ChAT; D–F). In each case, the
double-labeled neuron is seen under brightfield to show the AChEreaction product (A,D), under both brightfield and fluoresence (B,E), orunder only fluorescence (with a Cy3 optical filter) to show the p75 (C)or ChAT (F) label. Scale bar 5 100 µm.
164 D.H. HA ET AL.
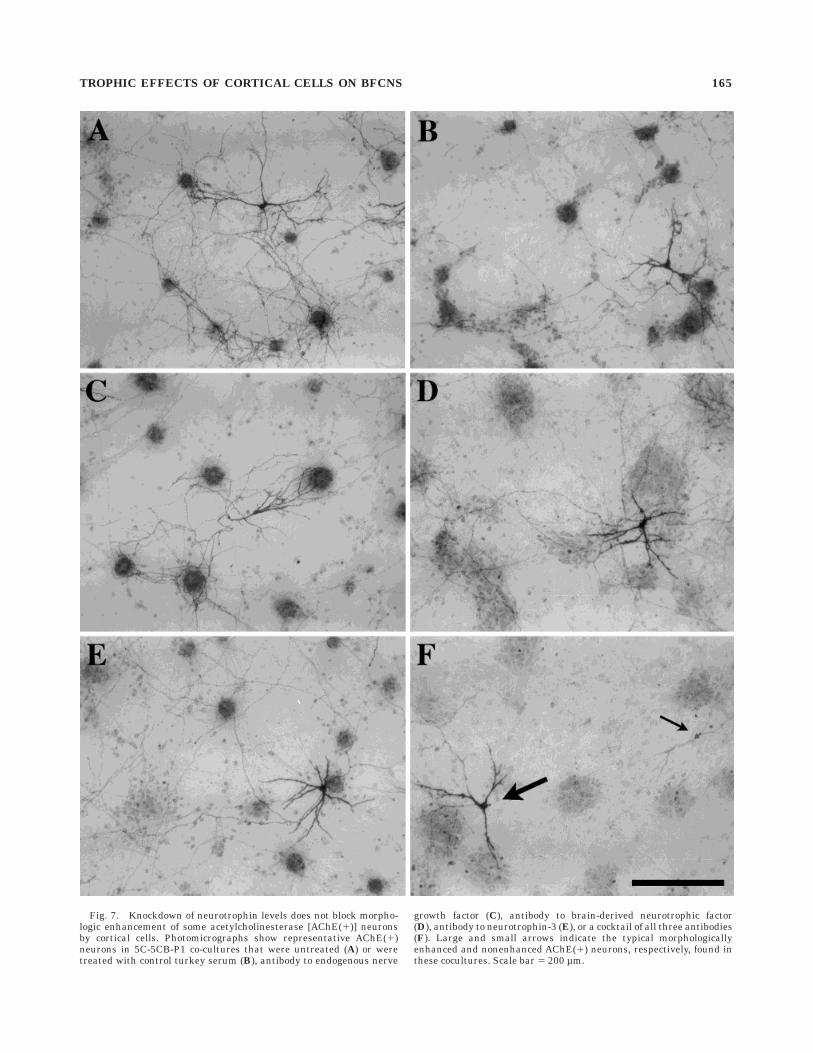
Fig. 7. Knockdown of neurotrophin levels does not block morpho-logic enhancement of some acetylcholinesterase [AChE(1)] neuronsby cortical cells. Photomicrographs show representative AChE(1)neurons in 5C-5CB-P1 co-cultures that were untreated (A) or weretreated with control turkey serum (B), antibody to endogenous nerve
growth factor (C), antibody to brain-derived neurotrophic factor(D), antibody to neurotrophin-3 (E), or a cocktail of all three antibodies(F). Large and small arrows indicate the typical morphologicallyenhanced and nonenhanced AChE(1) neurons, respectively, found inthese cocultures. Scale bar 5 200 µm.
TROPHIC EFFECTS OF CORTICAL CELLS ON BFCNS 165

Fig. 8. Knockdown of neurotrophin levels reduces the number ofmorphologically enhanced acetylcholinesterase [AChE(1)] neuronsand increases the number of nonenhanced cells. The 5C-5CB-P1cocultures were treated with single or combined neurotrophin anti-body exposure for 5 days, stained for AChE, and counted. A: Relativedistribution of type I and type II AChE(1) neurons in each of theindicated conditions. B: Change in number of type I and type IIAChE(1) neurons in each treated condition in comparison withnumbers of each type present in untreated cocultures. C: Change in
number of all AChE(1) neurons in each treated condition comparedwith the number present in untreated cocultures. Data are presentedas the mean 6 SEM (n 5 4 experiments; total of eight cultures for eachcondition). Asterisk and number sign indicate significant differencesin numbers of type I and type II AChE(1) neurons, respectively, fromother conditions in A and from untreated controls in B, P , 0.01.Ampersand indicates no significant differences from other conditions,P . 0.05.
cal neurons on BFCNs and to use neurotrophins andneurotrophin-neutralizing antibodies to examine the roleof cortically derived NGF, BDNF, and NT-3 in these effects.Thus, initial studies indicated that cortical cells increasedthe survival of and induced a rapid (within 5 days)morphological enhancement of prenatal and postnatalBFCNs in cocultures. Although exogenous NGF, BDNF,and NT-3 increased BFCN survival in pure BF cultures,they did not reproduce the morphological enhancementobserved after coculturing with cortical neurons. Con-versely, treating cocultures with neutralizing antibodies toNGF, BDNF, and NT-3 resulted in an apparent decrease inthe number of BFCNs that were morphologically enhancedand caused little change in the total number of survivingBFCNs.
In addition, present studies suggest that the growth-enhancing effects of cortical neurons on BFCN were maxi-mal at culture ages of both the cortical and BF cellscorresponding to early postnatal ages in vivo when inner-vation of cortex by BF projection is maximal.
Technical considerations
In a previous study, we investigated the influence ofcortical neurons on BFCNs in a novel coculture modelsystem (Ha et al., 1996). BFCNs in those cocultures versusthose in control cultures displayed larger somata, in-
creased dendritic arborization, and extensive axonal projec-tions that appeared to target neighboring clusters ofcortical neurons; electron microscopic analysis showedsynaptic contacts between the cholinergic axons and thecortical neurons which they appeared to target. The cocul-tures used in that study were plated with G-17 BF andcortical cells and grown for 14–21 days, a period thatcorresponds to late fetal and early postnatal developmentin vivo. The markedly shorter (5 day) duration of cocultur-ing employed by the present study enabled comparison ofthe extent of morphologic enhancement as a function ofmaturation of both the cortical and BF neurons.
This new culture paradigm provides other advantages.First, the ability to demonstrate the dramatic morphologicenhancement in a relatively brief (5 day) period providesan indication of the robustness of the effect. Furthermore,largely because of its brevity, the present system is wellsuited for evaluating effects of interventions (such asaddition or neutralization of neurotrophins) that mayeither block enhancement in cocultures or reproduce en-hancement in pure BF cultures, thereby providing clues tothe signaling mechanisms triggered by the cortical neu-rons. AChE histochemical staining was used to evaluatethe morphologic differentiation of BFCNs in the presentstudy because it labels the entire cell clearly, permittingquantification of dendritic and axonal length. However,
Fig. 9. Exogenous neurotrophins do not enhance morphologicalfeatures of postnatal acetylcholinesterase [AChE(1)] neurons. Photo-micrographs show representative AChE(1) neurons in a pure basalforebrain (BF) culture (5B-P1) that was untreated (A) or treated with
endogenous nerve growth factor, brain-derived neurotrophic factor,and neurotrophin-3 (B), and in a 5C-5CB-P1 coculture prepared withthe same BF cell suspension (C,D). Scale bar 5 100 µm in A–C, 300 µmin D.
TROPHIC EFFECTS OF CORTICAL CELLS ON BFCNS 167
because some cortical neurons also express AChE (Zhu etal., 1984; Hallanger et al., 1986; Mesulam and Geula,1991), control experiments showing expression of ChATand p75 low-affinity neurotrophin receptor (Kiss et al.,1993) by AChE(1) neurons confirmed their identity asBFCNs.
Morphological enhancement of BFCNsdepends on the maturational state of both
the cortical and BF neurons
The age at which either BF or cortical cells werecocultured was changed to determine possible maturationdependence of trophic effects. In these cocultures, a markedincrease in the degree of morphologic enhancement ofBFCNs was observed when the cortical neurons were
allowed to develop for 5 days prior to the addition of BFcells. Similarly, the morphologic enhancement of cocul-tured BFCNs was greater when the BFCNs were obtainedfrom postnatal rather than fetal animals. Although in vitrodevelopment of cortical cells may not exactly replicatedevelopment in vivo, several studies have indicated closesimilarities in patterns of development between the twosystems (Dichter, 1978; Kriegstein and Dichter, 1983;Annis et al., 1993). Thus, cortical neurons grown in culturefor 5 or for 12 days may correspond closely to theircounterparts (P-0 or P-7, respectively) in early postnatalanimals. These findings collectively suggest that the abil-ity of cortical cells to signal BFCN development anddifferentiation and the ability of the BFCNs to respond tosuch signals are developmentally regulated and are height-ened in both types of cells at stages of developmentcorresponding to early postnatal ages. Indeed, the periodof early postnatal development in vivo is the time of peakinnervation of cortical target cells by BFCNs (Dinopouloset al., 1989; Calarco and Robertson, 1995; De Carlos et al.,1995) and is distinguished by survival dependence ofBFCNs on target neurons (Burke et al., 1994; Cooper et al.,1996) and by increased expression of neurotrophins andtheir receptors by cortical neurons and BFCNs, respec-tively (Maisonpierre et al., 1990; Barbacid, 1994; Lauter-born et al., 1994). Similar expression of cortically derivedtrophic factors may well contribute to the age dependentenhancement of BFCNs in cocultures.
Role of cortical cells and neurotrophinsin BFCN development
Consideration of survival-enhancing effects. Neuro-trophins produced by cortical neurons have long beensuspected of providing survival-enhancing signals toBFCNs. However, BFCNs do not appear to be affected bythe absence of the individual neurotrophins, as has beenshown in the brains of transgenic mice lacking genesencoding for NGF, BDNF, or NT-3 (Crowley et al., 1994;Ernfors et al., 1994; Jones et al., 1994). The present studyhas attempted to clarify the role of neurotrophins in BFCNsurvival by assessing effects of neurotrophin addition topure BF cultures or neurotrophin neutralization in cocul-tures on numbers of surviving BFCNs. Indeed, addition ofneurotrophins to pure BF cultures resulted in a doublingof the number of AChE(1) neurons present after 5 days.Furthermore, control experiments, in which addition ofneurotrophins to BF cells after 5 days of growth in theirabsence failed to increase numbers of AChE(1) neurons,provide strong evidence that numbers of AChE(1) neuronspresent reflect actual numbers of surviving cholinergicneurons.
In a previous study, we found that the presence ofcortical neurons could prevent the degeneration ofBFCNs caused by addition of NGF-neutralizing antibodyduring a critical developmental period, suggesting thatcortical cells produce survival-enhancing factors distinctfrom NGF (Ha et al., 1996). Indeed, the finding that thepresence of cortical cells led to a tripling of numbers ofAChE(1) neurons (in contrast to combined addition ofneurotrophins that only doubled their numbers) indicatesthe presence of powerful cortically derived survival enhanc-ing effects. The further finding that neutralization of NGF,BDNF, and NT-3 in cocultures had little impact on thesurvival-enhancing effects of cortical cells on BFCNs fur-ther suggests that these survival effects are largely inde-
Fig. 10. Exogenous neurotrophins increase the number of postna-tal acetylcholinesterase [AChE(1)] neurons. A: Postnatal day 1 (P-1)basal forebrain (BF) neurons were plated with previously establishedcortical cells (5C-5CB-P1) or by themselves (5B-P1), with or withoutthe addition of neurotrophins (endogenous nerve growth factor, NGF;brain-derived neurotrophic factor, BDNF; neurotrophin-3, NT-3) attime of plating. After 5 days, the cultures were stained, and numbersof AChE(1) neurons were counted. B: Pure BF cultures were preparedwith P-1 BF neurons as described above but were grown for a total of 8days, with NGF, NT-3, and BDNF added to some of the cultures onlyduring the last 3 days. All values are normalized to numbers present insister 5B-P1 cultures grown for 8 days without added neurotrophins.Data are presented as means 6 SEM (n 5 3 experiments; total of 12cultures per condition). Asterisk indicates significant differences fromall other conditions, P , 0.01. Number sign indicates no significantdifference from untreated 8B-P1 condition, P , 0.001.
168 D.H. HA ET AL.

pendent of the neurotrophins. Although we cannot becertain that the antibodies completely eliminated all neu-rotrophin activity from the cultures, we believe they wereeffective because antibodies were used at levels that havebeen shown to block the neuronal survival and ChATexpression induced by exogenous neurotrophins (Rohrer etal., 1988; Ghosh et al., 1994; Nonner and Barrett, 1994;Svendsen et al., 1994; Ha et al., 1996). In addition (asdiscussed below), these same antibody exposures did havean effect on the total numbers of BFCNs undergoingmorphological enhancement.
Consideration of growth-enhancing effects. In addi-tion to increasing the number and thus survival of BFCNs,the presence of cortical cells also enhanced the expressionof morphological features of these neurons, as indicated bythe large somata, long and elaborate dendrites, and exten-sive axonal outgrowth of most AChE(1) cells in cocultures.In contrast, exogenous neurotrophins increased the num-ber but did not enhance morphological features of postna-tal BFCNs in treated pure BF cultures. Interestingly,combined neutralization of NGF, BDNF, and NT-3 incocultures resulted in a substantial decrease in the num-ber of morphologically enhanced BFCNs and an increasein the number of nonenhanced BFCNs. These data indi-cate that, although neurotrophins by themselves cannotinduce morphologic enhancement of postnatal BFCNs,they may play a permissive role by acting in conjunctionwith other cortically derived factors to influence BFCNdifferentiation. Another possible mechanism is that thepresence of cortical neurons allows BFCNs to form synap-tic targets with target cells, and the formation of synapsesleads to healthier cells that in turn may be more respon-sive to neurotrophic factors. Whether the unknown mecha-nisms that increase BFCN survival and those that areinvolved in the morphologic enhancement of BFCNs repre-sent the same set of cortically derived trophic factorsremains to be determined.
CONCLUSIONS
Although recent studies have indicated that neurotroph-ins induce powerful effects on BFCNs, clarification of theirrole in the survival and growth of BFCNs has beenconfounded by the relative paucity of alterations of BFCNsin neurotrophin knockout mouse models. The presentmodel system, by permitting addition or removal of factorsin a highly controlled environment, may be a particularlyuseful one for studying interactions between multiplefactors in modulating BFCN survival or phenotype. In-deed, present findings point strongly to the existence of anas yet unidentified cortically derived cholinergic trophicfactor in addition to the neurotrophins. Such factorsappear to be able to independently increase BFCN sur-vival; they also appear to be involved (perhaps with apermissive role of the neurotrophins) in the enhancementof BFCN growth and differentiation.
Present observations that the effects of cortical neuronson BFCNs occur most dramatically at culture ages corre-sponding to the period of early postnatal development ofBFCNs and cortical cells in vivo support the idea that thiscoculture paradigm is useful for studying developmentaltrophic influences of cortically derived factors on BFCNs.Knowledge of these factors may well prove relevant notonly to early development but also to diseases such as
Alzheimer’s disease, in which degeneration of corticalneurons may induce deleterious effects on the BFCNs.
ACKNOWLEDGMENTS
We thank Kimberly J. Claytor for expert assistance withthe cell cultures. This work was supported by NIH grantsNS 30884 (J.H.W.) and NS 30109 (R.T.R.), the Pew Schol-ars Program in the Biomedical Sciences (J.H.W.), andNRSA postdoctoral fellowship MH 11483 (D.H.H.).
LITERATURE CITED
Alderson RF, Alterman AL, Barde YA, Lindsay RM. 1990. Brain-derivedneurotrophic factor increases survival and differentiated functions ofrat septal cholinergic neurons in culture. Neuron 5:297–306.
Annis CM, Robertson RT, O’Dowd DK. 1993. Aspects of early postnataldevelopment of cortical neurons that proceed independently of normallypresent extrinsic influences. J Neurobiol 24:1460–1480.
Baratta J, Ha DH, Weiss JH, Yu J, Robertson RT. 1996. Cholinergic neuronsfrom different subdivisions of the basal forebrain lack connectionalspecificity for cerebral cortical target sites in vitro. Dev Brain Res97:143–147.
Baratta J, Ha DH, Weiss JH, Yu J, Phillips HS, Robertson RT. 1997. Basalforebrain cholinergic projections to neocortex and hippocampus: normalpatterns of ingrowth in slice cultures from NGF-KO mice. Soc NeurosciAbstr 23:333.
Barbacid M. 1994. The Trk family of neurotrophin receptors. J Neurobiol25:1386–1403.
Burke MA, Mobley WC, Cho J, Wiegand SJ, Lindsay RM, Mufson EJ,Kordowers JH. 1994. Age related vulnerability of developing cholinergicbasal forebrain neurons following excitotoxic lesions of the hippocam-pus. Exp Neurol 128:159–171.
Calarco CA, Robertson RT. 1995. Development of basal forebrain projec-tions to visual cortex: DiI studies in rat. J Comp Neurol 354:608–626.
Cooper JD, Skepper JN, Berzaghi MDP, Lindholm D, Sofroniew MV. 1996.Delayed death of septal cholinergic neurons after excitotoxic ablation ofhippocampal neurons during early postnatal development in the rat.Exp Neurol 139:143–155.
Crowley C, Spencer SD, Nishimura MC, Chen KS, Pitts-Meek S, ArmaniniMP, Ling LH, MacMahon SB, Shelton DL, Levinson AD, Phillips HS.1994. Mice lacking nerve growth factor display perinatal loss of sensoryand sympathetic neurons yet develop basal forebrain cholinergic neu-rons. Cell 76:1001–1011.
De Carlos JA, Schlaggar BL, O’Leary DDM. 1995. Development of acetyl-cholinesterase-positive thalamic and basal forebrain afferents to embry-onic rat neocortex. Exp Brain Res 104:385–401.
Dichter MA. 1978. Rat cortical neurons in cell culture: culture methods, cellmorphology, electrophysiology, and synapse formation. Brain Res 49:279–293.
Dinopoulos A, Eadie LA, Dori I, Parnavelas JG. 1989. The development ofbasal forebrain projections to the rat visual cortex. Exp Brain Res76:563–571.
Distler PG, Robertson RT. 1992. Development of AChE-positive neuronalprojections from basal forebrain to cerebral cortex in organotypic tissueslice cultures. Dev Brain Res 67:181–196.
Distler PG, Robertson RT. 1993. Formation of synapses between basalforebrain afferents and cerebral cortex neurons: an electron microscopicstudy in organotypic slice cultures. J Neurocytol 22:627–643.
Eckenstein FP, Baughman RW, Quinn J. 1988. An anatomical study ofcholinergic innervation in rat cerebral cortex. Neuroscience 25:457–474.
Ernfors P, Lee K-F, Kucera J, Jaenisch R. 1994. Lack of neurotrophin-3leads to deficiencies in the peripheral nervous system and loss of limbproprioceptive afferents. Cell 77:503–512.
Friedman WJ, Ibanez CF, Hallbook F, Persson H, Cain LD, Dreyfus CF,Black IB. 1993. Differential actions of neurotrophins in the locuscoeruleus and basal forebrain. Exp Neurol 119:72–78.
Ghosh A, Carnahan J, Greenberg ME. 1994. Requirement for BDNF inactivity-dependent survival of cortical neurons. Science 263:1618–1622.
Ha DH, Robertson RT, Ribak CE, Weiss JH. 1996. Cultured basal forebraincholinergic neurons in contact with cortical cells display synapses,enhanced morphological features, and decreased dependence on nervegrowth factor. J Comp Neurol 373:451–465.
TROPHIC EFFECTS OF CORTICAL CELLS ON BFCNS 169

Hallanger AE, Wainer BH, Rye DB. 1986. Co-localization of gamma-aminobutyric acid and acetylcholinesterase in rodent cortical neurons.Neuroscience 19:763–769.
Hartikka J, Hefti F. 1988. Development of septal cholinergic neurons inculture: plating density and glial cells modulate effects of NGF onsurvival, fiber growth, and expression of transmitter-specific enzymes.J Neurosci 8:2967–2985.
Hefti F. 1986. Nerve growth factor promotes survival of septal cholinergicneurons after fimbrial transections. J Neurosci 6:2155–2162.
Jones KR, Farinas I, Backus C, Reichard LF. 1994. Targeted disruption ofthe brain-derived neurotrophic factor gene perturbs brain and sensoryneuron but not motor neuron development. Cell 76:989–1000.
Kew JNC, Smith DW, Sofroniew MV. 1996. Nerve growth factor withdrawalinduces the apoptotic death of developing septal cholinergic neurons invitro—protection by cyclic AMP analogue and high potassium. Neurosci-ence 70:329–339.
Kiss J, Shooter EM, Patel AJ. 1993. A low-affinity nerve growth factorreceptor antibody is internalized and retrogradely transported selec-tively into cholinergic neurons of the rat basal forebrain. Neuroscience57:297–305.
Klein R, Martin-Zanca D, Barbacid M, Parada LF. 1990. Expression of thetyrosine kinase receptor gene trkB is confined to the murine embryonicand adult nervous system. Development 109:845–850.
Koh S, Higgins GA. 1991. Differential regulation of the low-affinity nervegrowth factor receptor during postnatal development of the rat brain. JComp Neurol 313:494–508.
Kordower JH, Chen EY, Sladek JR, Mufson EJ. 1994. Trk-immunoreactiv-ity in the monkey central nervous system: forebrain. J Comp Neurol349:20–35.
Kriegstein AR, Dichter MA. 1983. Morphological classification of ratcortical neurons in cell culture. J Neurosci 3:1634–1647.
Lamballe F, Smeyne RJ, Barbacid M. 1994. Developmental expression oftrkC, the neurotrophin-3 receptor, in the mammalian nervous system. JNeurosci 14:14–28.
Lauterborn JC, PJ Isackson, CM Gall. 1994. Cellular localization of NGFand NT-3 mRNAs in postnatal rat forebrain. Mol Cell Neurosci 5:46–62.
Lysakowski A, Wainer BH, Bruce G, Hersh LB. 1989. An atlas of theregional and laminar distribution of choline acetycholinesterase immu-noreactivity in rat cerebral cortex. Neuroscience 28:291–336.
Maisonpierre PC, Belluscio L, Friedman B, Alderson RF, Wiegand SJ,
Furth ME, Lindsay RM, Yancopoulos GD. 1990. NT-3, BDNF, and NGFin the developing rat nervous system: parallel as well as reciprocalpatterns of expression. Neuron 5:501–509.
Mesulam M-M, Geula C. 1991. Differential distribution of a neurofilamentprotein epitope in acetylcholinesterase-rich neurons of human cerebralcortex. Brain Res 544:169–173.
Morse JK, Wiegand SJ, Anderson K, You Y, Cai N, Carnahan J, Miller J,DiStefano PS, Altar CA, Lindsay RM, Alderson RF. 1993. Brain-derivedneurotrophic factor (BDNF) prevents the degeneration of medial septalcholinergic neurons following fimbria transection. J Neurosci 13:4146–4156.
Nonner D, Barrett JN. 1994. Changes in the response of cultured septalcholinergic neurons to nerve growth factor exposure and deprivationduring the first postnatal month. Dev Brain Res 79:219–228.
Nonomura T, Nishio C, Lindsay RM, Hatanaka H. 1995. Cultured basalforebrain cholinergic neurons from postnatal rats show both overlap-ping and non-overlapping responses to the neurotrophins. Brain Res683:129–139.
Robertson RT, Hanes MA, Yu J. 1988. Investigations of the origin oftransient acetylcholinesterase activity in developing rat visual cortex.Dev Brain Res 41:1–23.
Rohrer H, Hofer M, Hellweg R, Korsching S, Stehle AD, Saadat S, ThoenenH. 1988. Antibodies against mouse nerve growth factor interfere in vivowith the development of avian sensory and sympathetic neurones.Development 103:545–552.
Sobreviela T, Clary DO, Reichardt LF, Brandabur MM, Kordower JH,Mufson EJ. 1994. Trk-A immunoreactive profiles in the central nervoussystem: colocalization with neurons containing p75 nerve growth factorreceptor, choline acetylcholinesterase, and serotonin. J Comp Neurol350:587–611.
Svendsen CN, Kew JN, Staley K, Sofroniew MV. 1994. Death of developingseptal cholinergic neurons following NGF withdrawal in vitro: protec-tion by protein synthesis inhibition. J Neurosci 14:75–87.
Tago H, Kimura H, Maeda T. 1986. Visualization of detailed acetylcholines-terase fiber and neuron staining in rat brain by a sensitive histochemi-cal procedure. J Histochem Cytochem 34:1431–1438.
Zhu CH, Delfs JR, Mufson EJ, Dichter MA, Mesulam M-M. 1984. Acetyl-cholinesterase and somatostatin immunoreactivity coexist in neuronsin rat cerebral cortex and hippocampus, but not in Ch4 cholinergicneurons of the basal forebrain. Soc Neurosci Abstr 10:696.2
170 D.H. HA ET AL.