-
8/12/2019 Enhancement of Taxane Production in Hairy Root
1/5
Journal of Plant Physiology 166 (2009) 19501954
SHORT COMMUNICATION
Enhancement of taxane production in hairy root
culture of Taxus x media var. Hicksii
Katarzyna Syklowska-Baranek, Agnieszka Pietrosiuk, Anna
Kokoszka,Miroslawa Furmanowa
Department of Biology and Pharmaceutical Botany, Faculty of
Pharmacy, Medical University of Warsaw, ul. Banacha 1,
02-697 Warsaw, Poland
Received 23 March 2009; received in revised form 4 May 2009;
accepted 5 May 2009
KEYWORDS
10-deacetylbaccatinIII;Hairy roots;Paclitaxel;Taxus
SummaryThis study assessed the effect of two precursors
(L-phenylalanine and p-aminobenzoic acid) used alone or in
combination with methyl jasmonate, on the growthand accumulation of
paclitaxel, baccatin III and 10-deacetylbaccatin III in hairy
rootcultures of Taxus x media var. Hicksii. The greatest increase
in dry biomass wasobserved after 4 weeks of culturing hairy roots
in medium supplemented with 1 mM ofL-phenylalanine (6.2 g L1).
Addition of 1 mM of L-phenylalanine to the medium alsoresulted in
the greatest 10-deacetylbaccatin III accumulation (422.7 mg L1),
whichwas not detected in the untreated control culture.
Supplementation with 100 mM ofL-phenylalanine together with 100 mM
of methyl jasmonate resulted in theenhancement of paclitaxel
production from 40.3 mg L1 (control untreated culture)to 568.2 mg
L1, the highest paclitaxel content detected in the study. The
effect of p-amino benzoic acid on taxane production was less
pronounced, and the highest yieldof paclitaxel (221.8 mg L1) was
observed when the medium was supplemented with100 mM of the
precursor in combination with methyl jasmonate.Baccatin III was not
detected under the conditions used in this experiment and
theinvestigated taxanes were not excreted into the medium.&2009
Elsevier GmbH. All rights reserved.
Introduction
Paclitaxel is the active substance of Taxols
(Bristol-Myers Squibb), a very valuable plant-derived drug that
displays activity against differentcancers, including carcinomas of
the ovary, breast,head and neck, bladder and cervix, melanomas,lung
cancers and AIDS-related Kaposis sarcoma. In
ARTICLE IN PRESS
www.elsevier.de/jplph
0176-1617/$ - see front matter&2009 Elsevier GmbH. All
rights reserved.doi:10.1016/j.jplph.2009.05.001
Corresponding author. Tel.: +48 22 5720933;fax: +4822
5720984.
E-mail address:[email protected](K. Syklowska-Baranek).
http://www.elsevier.de/jplphhttp://localhost/var/www/apps/conversion/tmp/scratch_3/dx.doi.org/10.1016/j.jplph.2009.05.001mailto:[email protected]:[email protected]:[email protected]:[email protected]://localhost/var/www/apps/conversion/tmp/scratch_3/dx.doi.org/10.1016/j.jplph.2009.05.001http://www.elsevier.de/jplph
-
8/12/2019 Enhancement of Taxane Production in Hairy Root
2/5
addition, many clinical trials are underway to testthe efficacy
of paclitaxel, used as monotherapy andin combination with other
chemotherapeuticagents, against various other neoplasms (Nims
etal., 2006). The base taxane structure is utilized asthe starting
skeleton for molecules with improvedbiological properties (Kingston
and Newman, 2007).
Plant cell cultures provide a very promisingapproach to the
production of paclitaxel andrelated taxanes, and do not require
forest har-vestation. Although production has been scaled-up,a
serious drawback of cell suspension cultures istheir instability
and the unpredictability of second-ary metabolite formation
(Frense, 2007). Trans-formed or hairy roots, offer opportunities
forproduction of important phytochemicals as theyare characterized
by genetic and biochemicalstability and capacity for
organogenesis-associatedsynthesis of metabolites (Georgiev et al.,
2007).
The precursor feeding or elicitation strategy hasbeen also found
to effectively stimulate taxaneaccumulation in both callus and
suspension culturesofTaxusspecies (Zhong, 2002). However, yew
hairyroot cultures have not been sufficiently explored asa
potential source of taxanes.
In this paper we report the enhancement ofpaclitaxel and
10-deacetylbaccatin III (10-DAB III)production in transgenic root
cultures of Taxus xmedia var. Hicksii Rehd. achieved through
precur-sors and elicitor feeding.
Materials and methods
Transformed root cultures
Hairy root culture of Taxus x media var. Hicksiiwas derived from
a single root developed at thewounded site of the leaf as described
previously(Furmanowa and Syklowska-Baranek, 2000). Thehairy roots
were cultivated in 250mL Erlenmeyerflasks containing 50mL of
hormone-free DCR-Mmedium. This was the medium used by Gupta and
Durzan (1985)with the content of MgSO4increasedto 400mgL1
(Flores et al., 1993), and with theaddition of 500 mg L1 of
L-glutamine. The culturewas maintained at 251C in the dark on a
GioGyrotorys Shaker (New Brunswick Scientific Co.) at122 rpm.
Precursors and elicitor feeding
We investigated the effects of L-phenylalanine(PHE) and p-amino
benzoic acid (PAB), either alone
or in combination with methyl jasmonate, on hairyroot growth and
taxane production.
The precursors were added to the media prior toautoclaving, and
methyl jasmonate was addedafter autoclaving. The 0.6770.12 g of
fresh weightof 28-day-old hairy roots were placed into 250
mLErlenmeyer flasks with 50mL DCR-M medium
supplemented with precursors at concentrationsof 1 mM, 10 mM or
100 mM. The hairy roots from twoflasks were harvested after 48h, 1,
2, 3 and 4weeks, and their dry weight (D.W.) was determinedafter
lyophilization.
In the second part of the experiment, the 4-week-old hairy roots
were placed into mediasupplemented with precursors (1, 10 or
100mM)and 100 mM methyl jasmonate and cultured for 2weeks. Hairy
roots were also cultured for 4 weeksin DCR-M medium without either
precursors orelicitor to serve as controls.
HPLC analysis
The contents of paclitaxel, baccatin III and 10-deacetylbaccatin
III were determined in powdereddry tissue of transformed roots and
in the mediumsamples. Sample cleaning was performed using themethod
described by Theodoridis et al. (1998a).The residue was
re-dissolved in 100% methanol(300 mL). The resulting solution (25
mL) was inves-tigated by HPLC analysis using the DIONEX systemwith
a UVD 340S diode array detector and an
automated sample injector (ASI-100). For com-pound separation, a
NovaPak Phenyl column(150 3.9 mm) was eluted employing the
gradientprogram described by Theodoridis et al. (1998b).The DAD
spectrophotometer was set at a wave-length range from 215 to 275
nm. The taxanes wereidentified and quantified at 227 nm. The peaks
wereassigned by spiking the samples with the standardsand comparing
the retention times and UV spectra.The standard compounds
paclitaxel and baccatin IIIwere purchased at Sigma and
10-deacetylbaccatinIII was donated by Prof. Jaziri from the
Free
University of Brussels.All the experiments were conducted in
triplicate,with the contents of two flasks assayed from
eachtreatment.
Results
The aim of this study was to examine the effectsof the two
precursors L-phenylalanine and p-aminobenzoic acid, alone and in
combination with methyl
ARTICLE IN PRESS
Enhancement of taxane production in hairy root culture ofTaxus x
media var. Hicksii 1951
-
8/12/2019 Enhancement of Taxane Production in Hairy Root
3/5
jasmonate, on the growth and taxane accumulationin transgenic
root cultures (Figure 1).
Under the conditions assayed, the greatest drybiomass increase
was observed after 4 weeks ofculturing hairy roots in the presence
of theprecursor compounds (Table 1). Compared to thecontrol
culture, the maximum dry biomass (6.2 g
L1
) was recorded when 1 mM of PHE was added tothe medium. In
addition, the stimulatory effect of10 mM PAB was noted where the
dry mass of hairyroots rose up to 5.3 g L1 at the end of the
culture.
The media supplementation with the elicitorsignificantly
(Po0.05, t-test) reduced the growthof cultured hairy roots compared
to the controlculture and cultures supplemented with
precursorsalone (Table 1).
PHE influenced taxane production in a dose-dependent manner. The
addition of 1 mM PHE to themedia resulted in an 8-fold increase of
paclitaxel
content after 4 weeks of culture. 10-DAB III was not
detected in the control (untreated) culture, whilemedium
supplementation with 1 mM of PHE pro-duced the greatest 10-DAB III
accumulation underthe conditions of this experiment (Tables 2 and
3).In the presence of 10 mM of PHE, paclitaxel and 10-DAB III were
determined in roots only at 48h ofculture and there were no taxanes
in the hairy root
cultures growing in the medium supplemented with100 mM of PHE
alone. However, the combination of100 mM PHE with 100 mM methyl
jasmonate yieldedthe greatest (almost 28-fold) increase in
paclitaxelproduction (Table 2).
Paclitaxel and 10-DAB III accumulated in hairyroots during the
entire culture period carried out inthe media supplemented with
PAB. In most cases,the amounts of the investigated taxanes
weresignificantly higher (Po0.05,t-test) than in controlcultures
(Table 3). The highest paclitaxel contentwas detected when hairy
roots grew in the media
supplemented with 10 or 100 mM PAB in combinationwith methyl
jasmonate, 85.6 and 130.5 mg g1 D.W.,respectively. No taxanes were
found in the tissue ofhairy roots when 1 mM PAB and methyl
jasmonatewere added to the medium.
In this study, baccatin III was not detected eitherin the root
tissues or in the post-culture media.None of the examined taxanes
was excreted intothe media.
Discussion
The beneficial effect of precursor and elicitorfeeding on
paclitaxel and related taxane accumula-tion in Taxus cell culture
is well documented(Zhong, 2002; Pinol et al., 2007). The strategy
ofcombining of several precursors and/or elicitors
ARTICLE IN PRESS
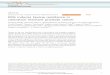
Figure 1. Transgenic roots ofTaxus x media var.
Hicksiicultivated 4 weeks in liquid hormone-free DCR-M medium(Photo
I.Rudnicki).
Table 1. Dry mass (g L1) increase ofTaxus x media var. Hicksii
transformed roots cultivated in DCR-M mediumsupplemented with
precursors or precursors and elicitor.
Time L-phenylalanine p-amino benzoic acid
1 mM 10 mM 100 mM 1 mM 10 mM 100 mM
48 h 1.270.1 1.370.1 1.570.5 1.470.3 1.570.3 1.870.31 week
1.670.3 1.770.2 1.570.3 2.470.4 2.370.4 2.170.12 weeks 2.170.3
2.270.8 2.470.2 3.170.7 3.270.2 2.470.53 weeks 3.270.4 1.970.1
1.870.4 3.670.8 3.470.4 3.2270.54 weeks 6.270.4 3.570.5 3.370.5
3.770.9 5.370.7 3.8470.5
L-phenylalanine+methyl jasmonate 100 mM p-amino benzoic
acid+methyl jasmonate 100 mM
2 weeks 1.770.7 1.570.7 1.770.8 1.570.4 1.570.3 1.570.4
Data are means7SE.Control growth of 4 week-old hairy roots in
DCR-M medium without precursors or elicitor: 3.4 g L170.7.
K. Syklowska-Baranek et al.1952
-
8/12/2019 Enhancement of Taxane Production in Hairy Root
4/5
Table 2. Taxane content in hairy root culture of Taxus x media
var. Hicksii carried out in DCR-M medium supplemented w11.670.9 mg
g1 D.W., 40.376 mg L1; 10-DAB III and baccatin III were not
detected).
L-phenylalanine
Time 1 mM 10 mM 100 mM
Paclitaxel 10-deacetylbaccatin III Paclitaxel
10-deacetylbaccatin III Paclitaxel
mg g1 D.W. mg L1 mg g1D.W. mg L1 mg g1 D.W. mg L1 mg g1D.W. mg
L1 mg g1 D.W.
48 h 15.375 20.875 116.977 158.179 1 week 2 weeks 3 weeks 18.872
55.377 22.373 65.376 4 weeks 52.377 335.679 65.878 422.7718
L-phenylalanine+methyl jasmonate 100 mM2 weeks 41.476 48.074
76.875 89.176 319.7714
Table 3. Taxane content in hairy root culture ofTaxus x media
var. Hicksii carried out in DCR-M medium supplemented with
p11.670.9 mg g1 D.W., 40.376 mg L1; 10-DAB III and baccatin III
were not detected).
p-amino benzoic acid
Time 1 mM 10 mM 100 mM
Paclitaxel 10-deacetylbaccatin III Paclitaxel
10-deacetylbaccatin III Paclitaxel
mg g1 D.W. mg L1 mg g1 D.W. mg L1 mg g1 D.W. mg L1 mg g1 D.W. mg
L1 mg g1D.W.
48 h 33.376 48.176 39.473 47.374 23.474 37.875 24.872 34.774
83.775 1 week 8.272 19.873 20.273 50.576 12.773 28.876 30.174
68.175 27.774
2 weeks 15.274 37.176 33.574 37.173 4.271 13.074 18.174 56.475
6.571 3 weeks 20.576 79.778 16.373 52.475 16.874 56.877 30.176
101.679 12.572 4 weeks 16.574 60.175 30.674 136.1711 18.2 78.478
11.873 42.274 12.172
p-amino benzoic acid+methyl jasmonate 100 mM2 weeks 85.679
145.7712 11.775 22.372 130.579
-
8/12/2019 Enhancement of Taxane Production in Hairy Root
5/5
seems to be promising. However, to date, taxaneproduction in
cultures ofTaxushairy roots has beendescribed in only two studies.
Huang et al. (1997)examined 5 clones ofT. brevifoliatransgenic
roots.The highest paclitaxel content was determinedafter 3 weeks of
culture (480 mg g1 D.W.) and thecompound was also found in the
medium (13 mg
L1
). In our previous study, the greatest paclitaxelyield (420 mg
g1 D.W.) was achieved after a 3-weekelicitation with methyl
jasmonate (Furmanowaand Syklowska-Baranek, 2000), but the
elicitordid not cause any increase in the 10-DAB IIIcontent.
Nims et al. (2006) observed increased transcriptabundance of
genes involved in late steps of thetaxane biosynthetic pathway
(10-DAB III, baccatinIII and paclitaxel formation) after methyl
jasmo-nate addition. The enhanced enzyme activitycorrelated with
the induction of taxane accumula-
tion. However, low activity of enzymes engaged in10-DAB III
biosynthesis was also noted in unelicitedcultures. In our
experiments, neither 10 DAB III norbaccatin III were found in the
unelicited controlculture, but paclitaxel was detected after 4
weeksof culture.
Our results may suggest that the regulation ofgene expression
engaged in the taxane biosyntheticpathway may differ in Taxus cell
suspensioncultures and Taxus hairy root cultures. The studyalso
demonstrates the suitability of the precursorand elicitor feeding
strategy for augmentation ofpaclitaxel and 10-deacetylbaccatin III
production in
Taxus x media var. Hicksii hairy root cultures.
Acknowledgements
The Medical University of Warsaw Research Fundis thanked for
financial assistance. We are gratefulto Prof. Mondher Jaziri from
the Laboratory ofBiotechnology and Plant Morphology, Free
Univer-sity of Brussels for the standard substance of 10-
deacetylbaccatin III.
References
Flores T, Wagner LJ, Flores HE. In vitro Cell
Dev-Pl1993;29:1605.
Frense D. Appl Microbiol Biotechnol 2007;73:123340.Furmanowa M,
Syklowska-Baranek K. Biotechnol Lett
2000;22:6836.Georgiev MI, Pavlov AI, Bley T. Appl Microbiol
Biotechnol
2007;74:117585.Gupta PK, Durzan DJ. Plant Cell Rep
1985;4:1779.
Huang Z, Mu Y, Zhou Y, Chen W, Xu K, Yu Z, et al. Acta
BotYunnanica 1997;19:2926.Kingston DGJ, Newman DJ. Curr Opin Drug
Discovery
2007;10:13044.Nims E, Dubois CP, Roberts SC, Walker EL. Metab
Eng
2006;8:38594.Pinol MT, Cusido RM, Palazon J, Bonfill M. In: Pua
EC,
Davey MR, editors. Biotechnology in Agriculture andForestry.
Berlin Heidelberg: Springer-Verlag; 2007.p. 20523.
Theodoridis G, Jong CF, Laskaris G, Verpoorte R.Chromatographia
1998a;47:2534.
Theodoridis G, Laskaris G, Jong CF, Hofte AJP, VerpoorteR. J
Chromatogr A 1998b;802:297305.
Zhong JJ. J Biosci Bioeng 2002;94:591
9.
ARTICLE IN PRESS
K. Syklowska-Baranek et al.1954