Embed Size (px)
Citation preview

BIOSYNTHESIS OF DICARBOXYLIC ACIDS BY CARBON DIOXIDE FIXATION
IV. ISOLATION AND PROPERTIES OF AN ADAPTIVE “MALIC” ENZYME FROM LACTOBACILLUS ARABINOSUS*
BY SEYMOUR KORKES, ALICE DEL CAMPILLO, AND SEVER0 OCHOA
(From the Department of Pharmacology, New York University College of Medicine, New York)
(Received for publication, April 5, 1959)
Lactobadus arabinosus (strain 17-5) may be adapted to ferment l- malic acid (1). As previously reported (2), the enzyme system can be readily extracted from the organism and has been partially purified. The partially purified enzyme catalyzes Reactions 1 and 2.’
(l) (DPN, Mn++)
COOH-CHz---CHOH-COOH ~ ____________________ 1
I-Malic acid
CHI-CHOH-COOH + CO
Lactic acid
(2) COOH-CHs--CO-COOH (Mn++)
-+ CHB-CO-COOH + CO2
Oxalacetic acid Pyruvic acid
Reaction 1 requires DPN and manganous ions. It is specific for I- malic acid; d-malic acid is inactive. TPN cannot replace DPN. Reaction 2 requires the presence of manganous ions. The evidence presented below supports the view that Reaction 1 is a DPN-linked dismutation between Z-malate and pyruvate, resulting from the interaction of lactic dehydrogenase with a DPN-specific “malic” enzyme as shown by Re- actions 3 and 4.
(Mn++) (3) I-Malate + DPN -------- ox, p-------------- pyruvate + COZ + DPN,,d. (“malic” enzyme)
* Aided by grants from the United States Public Health Service, the American Cancer Society (recommended by the Committee on Growth of the National Re- search Council), the Office of Naval Research, and the Lederle Laboratories Di- vision, American Cyanamid Company.
1 Solid line arrows indicate the direction of faster reaction. The usual abbrevi- ations are used: DPN for diphosphopyridine nucleotide, TPN for triphosphopyridine nucleotide, and ATP for adenosinetriphosphate.
891
by guest on September 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from
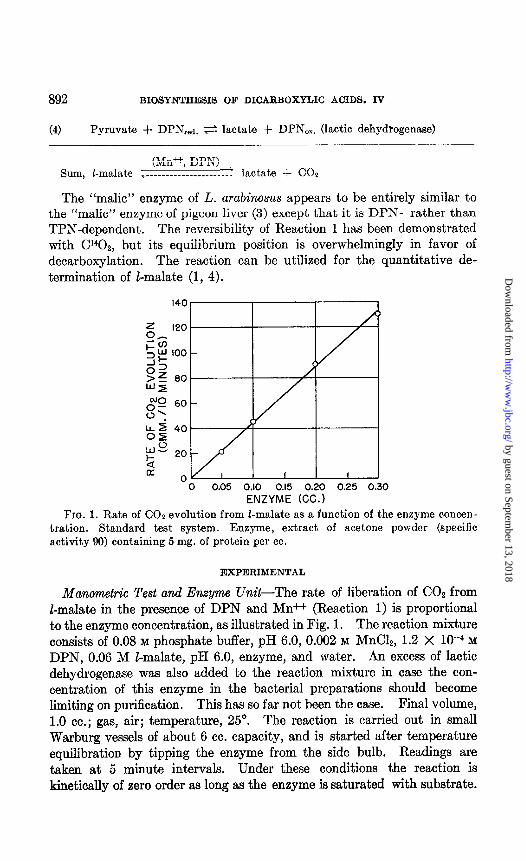
892 BIOSYNTHESIS OF DICARBOXYLIC ACIDS. IV
(4) Pyruvate + DPN,,d. Z$ lactate + DPN,,. (lactic dehydrogenaae)
(Mn++, DPN) Sum, l-malate ’ lactate + CO2 ~---------------------
The “malic” enzyme of L. urabinosus appears to be entirely similar to the “malie” enzyme of pigeon liver (3) except that it is DPN- rather than TPN-dependent. The reversibility of Reaction 1 has been demonstrated with Cr402, but its equilibrium position is overwhelmingly in favor of decarboxylation. The reaction can be utilized for the quantitative de- termination of Lmalate (1, 4).
140 140
120 120
100 100
60 60
60 60
40 40
20 20
0 n -0 0 0.05 0.05 0.10 0.15 0.20 0.25 0.30 0.10 0.15 0.20 0.25 0.30
ENZYME (CC.) ENZYME (CC.) FIG. 1. Rate of CO2 evolution from Z-malate as a function of the enzyme
tration. Standard test system. Enzyme, extract of acetone powder activity 90) containing 5 mg. of protein per cc.
concen- (specific
EXPERIMENTAL
Manometric Test and Enzyme Unit-The rate of liberation of COZ from I-malate in the presence of DPN and Mn++ (Reaction 1) is proportional to the enzyme concentration, as illustrated in Fig. 1. The reaction mixture consists of 0.08 M phosphate buffer, pH 6.0, 0.002 M MnC&, 1.2 X 10V4 M
DPN, 0.06 M I-malate, pH 6.0, enzyme, and water. An excess of lactic dehydrogenase was also added to the reaction mixture in case the con- centration of this enzyme in the bacterial preparations should become limiting on purification. This has so far not been the case. Final volume, 1.0 cc.; gas, air; temperature, 25”. The reaction is carried out in small Warburg vessels of about 6 cc. capacity, and is started after temperature equilibration by tipping the enzyme from the side bulb. Readings are taken at 5 minute intervals. Under these conditions the reaction is kinetically of zero order as long as the enzyme is saturated with substrate.
by guest on September 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

S. KORKES, A. DEL CAMPILLO, AND S. OCHOA 893
The volume of COZ produced is calculated by correcting the vessel constant for the small COZ retention at pH 6.0.
The test for oxalacetic carboxylase activity (Reaction 2) is carried out in the same way. The reaction mixture contains 0.08 M acetate buffer, pH 4.5, 0.08 M phosphate adjusted to the same pH, 0.002 M MnC&, 0.023 M oxalacetate (acid dissolved just before use and adjusted to pH 4.5), enzyme, and water. In this case a blank without enzyme is run simul- taneously to correct for the spontaneous decarboxylation of oxalacetate. The evolution of COZ due to the enzyme is of zero order over a good part of its course and is proportional to the enzyme concentration. 1 enzyme unit is arbit,rarily defined in both tests as the amount of enzyme catalyzing the liberation of 1.0 c.mm. of COZ in 10 minutes, calculated for the second 5 minutes following tipping in the enzyme. The amount of enzyme is adjusted so that the rate of CO1 liberation is between 30 and 90 c.mm. per 10 minutes.
The protein content of the enzyme solution is determined by either of two methods. In the initial stages, when the nucleic acid content is high, protein is estimated by the method of Heidelberger and MacPherson (5) as modified by Lo~ry.~ In the later stages, protein is determined spectrophotometrically by measurement of the light absorption at wave- lengths 280 and 260 mp. A correction for the nucleic acid content, from the data given by Warburg and Christian (6), is used to calculate the protein concentration. The specific activity of the enzyme is expressed as units per mg. of protein.
Isolation of Enzyme
L. urabinosus 17-5 is the starting material. The organisms are grown on a non-synthetic medium containing glucose and I-malate (1). After 24 hours, the cells are harvested by centrifugation and suspended in distilled iced water to make a thin paste.
Preparation of Acetone Powder-The chilled bacterial paste is poured all at once into 10 volumes of acetone chilled to - 10” in a Waring blendor; the mixture is homogenized for 2 minutes and then filtered by suctioq in the cold room. The cake is washed twice with small portions of cold acetone and the suction is continued until the cake is almost dry. The residue is then crumbled by hand, spread on filter paper, and allowed to dry in the air at room temperature. The dry material is ground to a fine powder in a mechanical mortar and stored in stoppered bottles in the ice box. The yield of dry powder is about 3 gm. per liter of growth medium. When stored in the cold, the powder is stable for at least 1 year.
* Personal communication. We are indebted to Dr. 0. H. Lowry for this in. formation.
by guest on September 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

894 BIOSYNTHESIS OF DICARBOXYLIC ACIDS. IV
The following steps are carried out in the cold. Sfep 1. Extracticzn-3 gm. of powder are extracted with 150 cc. of 0.02
M phosphate buffer, pH 7, by grinding the mixture in a mechanical mortar for 3 hours at 3-4’. The insoluble residue is removed by centrifugation at 0” and discarded. The viscous extract (about 130 cc.) contains 4 to 5 mg. of protein per cc. Specific activity 80 to 90.
Step 2. First Ammonium Sulfate Fractionation-To 130 cc. of extract are added 45 gm. of solid ammonium sulfate (degree of saturation 0.5) with mechanical stirring. After 20 minutes, the precipitate is centrifuged at 15,000 r.p.m. and discarded. To the supernatant is added sufficient ammonium sulfate to saturate the solution, and after 30 minutes the mixture is centrifuged and the supernatant discarded. The precipitate is dissolved in 0.02 M phosphate buffer, pH 7.0, and dialyzed against the same buffer overnight. The resultant solution of enzyme (about 30 cc.) contains 7 to 8 mg. of protein per cc. Specific activity about 150.
Step S. Precipitation of Nucleic Acid and Inactive Protein with Pro- tamine-30 cc. of enzyme solution from Step 2 are brought to pH 6.0 with approximately 0.2 cc. of 1.0 N acetic acid. To this solution is slowly added with mechanical stirring 0.85 cc. of a solution of protamine sulfate (20 mg. per cc.; pH 5.0) per 100 mg. of protein. This quantity is sufficient to bring the ratio of light absorption at 280 rnp to light absorption at 260mp of the enzyme solution from values of about 0.5 to 1.0 or slightly higher, indicating a reduction of the nucleic acid content to about 3 per cent, with a minimal precipitation of enzyme protein. The precipitate is centri- fuged and discarded. The supernatant (32 cc.) contains approximately 3 mg. of protein per cc. Specific activity about 280.
Step 4. Removal of Inactive Proteins with Calcium Phosphate Gel-The enzyme solution from Step 3 is adjusted to pH 5.5 with 2 drops of 1.0 N
acetic acid, and calcium phosphate gel (36 mg. of Ca3P04 per cc.) is added slowly with mechanical stirring in the proportion of 4 cc. per 100 mg. of protein. After 20 minutes the mixture is centrifuged and the precipitate discarded. The supernatant (32 cc.) contains the enzyme. The protein concentration is about 1.0 mg. per cc. Specific activity about 600.
Step 6. Second Ammonium Sulfate Fractionation-The procedure of Step 2 is repeated; the solution is first brought to 0.5 saturation with ammonium sulfate, the precipitate discarded, more ammonium sulfate added to satura- tion, and the second precipitate suspended in 0.02 M phosphate buffer, pH 7.0. The solution is dialyzed overnight against the same buffer. Volume, 9 cc.; protein about 2.5 mg. per cc.; specific activity about 950.
Step 6. Adsorption on Calcium Phosphate Gel-2.0 cc. of the enzyme solution from Step 5 were mixed with 0.5 cc. of calcium phosphate gel
by guest on September 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

S. KORKES, A. DEL CAMPILLO, AND S. OCHOA 895
after being adjusted to pH 5.5 with acetic acid. After stirring for 20 minutes, the mixture was centrifuged and the supernatant discarded. The precipitate was washed with 2 cc. of 0.1 M phosphate buffer, pH 5.5, for 10 minutes and centrifuged. The supernatant contained about one-fourth of the enzyme units at specific activity 850. The precipitate was eluted with 2 cc. of 0.1 M phosphate buffer, pH 6.0, under stirring for 20 minutes. The eluate contained about 0.7 mg. of protein per cc. Specific activity about 1800.
Table I summarizes the results of two typical runs with the procedure described above. It will be seen that the purification is fairly reproduci-
TABLE I Purification of L. arabinosus Enzyme
3 gm. of acetone powder (a and 6 separate runs).
step
Phosphate extract
1st ammonium sulfate frac- tionation
Protamine supernatant
Calcium phosphate superna- tant
2nd ammonium sulfate frac- tionation
Elution from calcium phos- phate gel
RIUI 1
_-
-
Iolllme of solution Units Protein
cc. m?.
129 52,000 580 134 54,500 627 31 34,100 217 31 34,000 258 32 25,100 91 31 28,000 100 33 25,000 38 32 26,400 41
8.5 24,300 25 10.4 25,000 26 2.0 2,570 1.4
-
1 m
-
Specific activity
wits per rg. protei
89 87
157 132 275 280 660 643 988 964
1840
_-
n
-
Yield
per cent
100 100 66 62 48 51 48 48 47 46 21
* Aliquot of the previous step.
ble. At various steps in the purification the oxalacetic carboxylase (Re- action 2) and lactic dehydrogenase (Reaction 4) activities of the prepara- tions were measured, and the results expressed as specific activities in Table II. The lactic dehydrogenase test and units were those described in a previous publication (7). It will be noted that over a range of 30- fold purification the ratio of activities with malate and oxalacetate as substrates remains constant, while the ratio of lactic dehydrogenase to “malic” activity falls from about 5 to 1, although lactic dehydrogenase is also partially purified. Although some fumarase is present in the initial extract, as determined by substituting fumarate for malate in the test system, it disappears after the first calcium phosphate treatment.
by guest on September 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

896 BIOSYNTHESIS OF DICARBOXYLJC ACIDS. IV
Properties of Enzyme System
Formation of Lactate-The balance illustrated in Table III indicates that for each mole of malate disappearing 1 mole each of lactate and carbon dioxide is formed, in accordance with Reaction 1. The enzyme is
TABLE II Ratios of Activities at Various Purijication Stages
Specific activity
“Mslic” enzyme Oxalacetic carboxylase Lactic dehydrogenase 0 L
CL) T -E
03 0)
52 238 87 80 310
314 335 943 456 492 1190 515 506 1220 956 915
1840 1755 2620* 2700 3370
238t 223 1675
0.92
1.07 1.08 0.98 0.96
5.5 3.6 3.0 2.6 2.4
1.03 0.94
______
0.95 1.3 7.0
-~ * Results obtained by Dr. S. E. Kaufman in the course of further purification of
the enzyme. t After heating a sample of specific activity 414 to 49’ for 8 minutes.
TABLE III Chemical Balance of Reaction 1
Enzyme (1.0 cc. of extract of acetone powder containing 4.7 mg. of protein per cc.) incubated with 150 PM of l-malate, pH 6.0, 0.08 M acetate buffer, pH 6.0, 0.08 M
phosphate buffer, pH 6.0, 0.33 PM of DPN, and 5 PM of MnC12. Final volume, 2.7 cc.; gas, air; temperature, 25’; incubation time, 100 minutes. Analyses performed on aliquots of trichloroacetic acid filtrates as described under “Methods.” The values are expressed in micromoles.
1 I-Make 1 Lactate ) CO2 1 Pyruvate
Initial. ........................ Final .......................... A .............................. .
quite specific for I-malate; d-malate is not attacked, nor does it inhibit the reaction. It should be noticed that a small amount of pyruvate is formed.
pH Optimum-Fig. 2 represents the pH-activity curves for the reaction with both malate and oxalacetate; i.e., for Reactions 1 and 2. Activity with malate shows a sharp optimum at pH 6.0, whereas activity with oxalacetate exhibits an equally sharp optimum at pH about 4.5. It should
by guest on September 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

l OXALACETI l OXALACETI
-4 4 5 5 6 6 7 7
PH PH FIG. 2. pH-activity curves. 0, oxalacetic carboxylase activity (Reaction 2) ;
0, “malic” activity (Reaction 1). Standard manometric tests as described in text. Enzyme of specific activity 150. All samples were buffered with acetate and phosphate adjusted to the desired pH; the substrates were also adjusted to the pH of the measurement. The pH at the end of each run was at the most 0.1 to 0.2
FIG. 2. pH-activity curves. 0, oxalacetic carboxylase activity (Reaction 2) ;
0, “malic” activity (Reaction 1). Standard manometric tests as described in text. Enzyme of specific activity 150. All samples were buffered with acetate and phosphate adjusted to the desired pH; the substrates were also adjusted to the pH of the measurement. The pH at the end of each run was at the most 0.1 to 0.2 unit higher than the initial value.
MINUTES MINUTES FIG. 3. Components of enzyme system. Enzyme of specific activity 900, di-
alyzed overnight at 2” against 0.05 M potassium chloride; 0.11 mg. of protein per sample. Standard manometric tests. A, system of Reaction 1, Curve 1, complete system; Curve 2, no phosphate (buffered with succinate); Curve 3, no Mn++; Curve 4, either no DPN or TPN in place of DPN; Curve 5, no malate. II, system of Re- action 2, Curve 1, complete system; Curve 2, no phosphate (buffered with acetate only); Curve 3, complete plus 1.2 X 1O-4 M DPN; Curve 4, complete plus 0.05 M Z-malate of pH 4.5; Curve 5, complete plus 0.05 M malonate of pH 4.5; Curve 6, no Mn++. The first 5 minutes after the start of the reaction have been disregarded. All values in B are corrected by subtracting the CO2 liberated from oxalacetate in the absence of enzyme.
by guest on September 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

898 BIOSYNTHESIS OF DICARBOXYLIC ACIDS. IV
be noted that at pH 6.0 the rate of decarboxylation of oxalacetate is only about one-eighth of the maximal rate, a point which will be discussed below.
Components-In addition to E-malate, the system of Reaction 1 requires catalytic amounts of DPN and Mn++; this is illustrated in Fig. 3, A. It is of interest that, in contrast to the pigeon liver “malic” enzyme, TPN is completely without effect. Co++ may replace Mn++ completely, but Mg++ is considerably less effective. The dissociation constants of the complex between the protein and each of the other components of Reaction 1 are as follows: I-malate, 9.8 X 10M3; DPN, 1.96 X 10”; Mn++, 2.9 X 1O-4 (all expressed in moles X liter-‘). In determining the above con- stants for each of the components all others were present in substantial excess. As illustrated in Fig. 3, B the system of Reaction 2 requires only Mn++ in addition to oxalacetate. The dissociation constant of the oxal- acetate-enzyme complex is about 1O-2 (moles of oxalacetate per liter); i.e., essentially the same as that of the I-malate-enzyme complex. The half saturation concentration of Mn++ in Reaction 2 is about 3 X low4 M and is thus identical to the corresponding value for Reaction 1. It is of interest to compare these data with those obtained with pigeon liver ‘Lmalic” enzyme (8). Included in Fig. 3, B are curves showing the in- hibitory action of Z-malate and malonate on Reaction 2. 0.05 M malate, i.e. about the same concentration at which it is used as the substrate of Reaction 1, caused 60 per cent inhibition of Reaction 2. The same con- centration of malonate produced a stronger inhibition (8). Reaction 2 was completely inhibited by 0.12 M malonate, while Reaction 1 was in- hibited only to the extent of 30 per cent.
Whereas, with dialyzed acetone powder extracts, Reaction 1 had been found to proceed about equally well in the absence or presence of ortho- phosphate (2), an activation by phosphate is shown with the purer frac- tions (Fig. 3, A). This is so in spite of adequate buffering with succinate buffer. In the absence of orthophosphate, activity drops to about one- fourth of that in its presence. The effect, of phosphate does not seem to be due to preserving the enzyme from inactivation because activity can be restored to maximal levels on adding phosphate as late as 20 minutes after the reaction is started. Activation by phosphate is only observed in Reaction 1. As shown in Fig. 3, B, removal of phosphate results in no appreciable change in the activity of Reaction 2. Arsenate in low concentra- tions does not replace the phosphate requirement and in higher concen- trations is inhibitory. The mode of action of phosphate remains so far unexplained” It does not seem to be involved in phosphorylation reactions;
8 Note added at proof correction. The activation appears to be due to the cation potassium and not to the phosphate ion (potassium phosphate was the buffer used). When potassium succinate is substituted for the sodium succinate, formerly used as
by guest on September 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

S. KORKES, A. DEL CAMPILLO, AND S. OCHOA 899
no coupled phosphorylation was observed when Reaction 1 was allowed to proceed in the presence of phosphate, glucose, hexokinase, and catalytic amounts of ATP. ATP is without effect on either Reaction 1 or 2.
Reversibilzty of Reaction l--With air in the gas phase, Reaction 1 goes essentially to completion. However, Fig. 4 indicates that the presence of pyruvate with increasing concentrations of CO2 in the gas phase exerts an increasingly inhibitory effect on the rate of the reaction. No inhibition was observed with lactate instead of pyruvate. This indicated reversibil-
I - NO PYRUVATE, 100% N2
2- PYRUVATE, 100% N,
3- PYRUVATE, 95% Np, 5%COz
4- PYRUVATE, 75% Ne,25%C02
5- PYRUVATE, 50% N, , 50%C02
6-PYRUVATE, 100%G02
0 5 IO 15 20
MINUTES FIG. 4. Inhibition of malate dissimilation by pyruvate and COZ. Standard
manometric test, except that 0.022 M, instead of 0.06 M, I-malate was used. 0.12 M
pyruvate was added when indicated. Sufficient NaHC03 to give pH 6.0 when equili- brated with each of the gas mixtures was added when COz was present. Enzyme of specific activity 150 with 0.4 mg. of protein in each sample. Final volume, 1.5 cc.
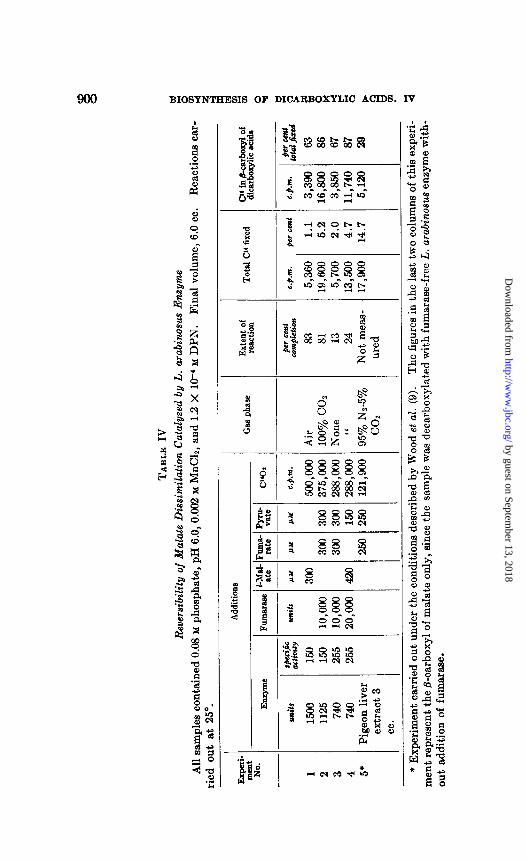
ity of the postulated partial Reaction 3. Evidence for reversibility is pre- sented in Table IV, in which incorporation of isotopic COZ in the dicar- boxylic acids was measured.
In Experiments 2 and 3, fumarate was substituted for malate and, be- sides the bacterial enzyme, purified fumarase was added. Fumarase was
buffer in the absence of phosphate, the system was found to be quite active. The effect of K+ on malate dissimilation by cell suspensions of adapted L. arabinosus was discovered by P. M. Nossal in the laboratory of H. A. Krebs. We are indebted to Dr. Nossal and Dr. Krebs for this information. An activating effect of K+ on a sim- ilar system in cell suspensions of Moraxella Iwo& had been described earlier by Lwoff and Ionesco (19).
by guest on September 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from
TABL
E IV
R
ever
sibi
lity
of M
alate
Di
ssim
ilatio
n Ca
talyz
ed
by L
. ar
abin
osus
En
zym
e m
All
sam
ples
co
ntai
ned
0.03
M p
hosp
hate
, pH
6.
0,0.
002
M M
nCL,
an
d 1.
2 X
lo-”
M D
PN.
Fina
l vo
lum
e,
6.0
cc.
Reac
tions
ca
r- s d
ried
out
at
25O
.
Ewf-
NO.
1 2 3 4 5*
1500
15
0 11
25
150
740
255
740
255
Pige
on
liver
ex
tract
3
cc.
I
Addi
tions
units
10,0
00
10,0
00
20,0
00
CMal.
at
e
PM
300
420
PM
PM
300
300
300
300
150
250
250
00,
c.p.m
.
5(@,oo
o 37
5,00
0 =W
JfJ
238,
000
121,
900
Gas
phas
e
Air
100%
CO
* No
ne
I‘
95%
Nz
-5%
co
2
Exte
nt
of
reac
tion
per
cent
comp
letion
83 81
13
24
Not
m
eas-
ur
ed
- I --
-
Tota
l C”
fix
ed
- I c.
p.m
.
5,36
0 19
,600
5,
700
13,5
00
17,9
00
fier
cent
1.1
5.2
2.0
4.7
14.7
Cl4
in #
-carb
xyl
of
dicsr
boxy
lic
acid
s
C.).
#.
t.%gd
3,
390
63
16,8
00
86
3,85
0 67
11
,740
87
5,
120
29
I
* Ex
perim
ent
carri
ed
out
unde
r th
e co
nditio
ns
desc
ribed
by
Woo
d et
al.
(9).
The
figur
es
in t
he
last
tw
o co
lum
ns
of t
his
expe
ri-
q m
ent
repr
esen
t th
e fl-
carb
oxyl
of
mal
ate
only
, sin
ce t
he
sam
ple
was
deca
rbox
ylate
d wi
th
fum
aras
e-fre
e L.
ara
bino
sus
enzy
me
with
- ou
t ad
ditio
n of
fum
aras
e.
by guest on September 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

S. KORKES, A. DEL CAMPILLO, AND S. OCHOA 901
also added in Experiment 4. This was done with the hope of increasing the total incorporation of isotopic COS. Under these conditions, the fol- lowing reactions occur.
COOH-CH=CH-COOH + Hz0 $ COOH-CHz-CHOH-COOH = CHB-CHOH-COOH + CO,
Establishment of the fumarase-catalyzed equilibrium would result both in an appreciable shift of the equilibrium position of the system in the direction of COz incorporation and in a distribution of the isotopic CO2 primarily fixed in the p-carboxyl group of malate, in both carboxyl groups of the dicarboxylic acids and in the carboxyl groups of lactate and pyruvate.
In one experiment dialyzed pigeon liver extract was substituted for the bacterial enzyme for comparison. This experiment was carried out under conditions similar to those of Wood e2 al. (9). The fixation of COz is greater than with the bacterial enzyme. This may be due to the fact that coupling of Reaction 3 with Reaction 4, which markedly shifts the equilib- rium in the direction of decarboxylation, is less efficient in the liver system because of the TPN specificity of its malic enzyme and of the slow rate at which lactic dehydrogenase reacts with TPN (7).
No fixation of COz in oxalacetate was found when the preparations were allowed to decarboxylate oxalacetate in the presence of CY402 irrespective of the presence or absence of ATP.4 These experiments were carried out by a procedure similar to that described by Utter and Wood (10) and Ven- nesland et al. (11) with pigeon liver enzymes.
Optical Nature of Lactate Fomzed-With intact cells adapted to malate, the product of glycolysis and of malate dissimilation has been shown to be d&lactate. Crude preparations of the enzyme (specific activity about 100) also form d&lactate from E-malate. This is interesting in the light of the strict optical specificity of the enzyme for I-malate as substrate. For iden- tification of the lactate the reaction was allowed to proceed to completion; the lactic acid was extracted with ether and isolated as the zinc salt. This proved to be zinc dl-lactate on the basis of the water of crystallization, as shown by the following data: per cent Hz0 found, 18.09; theory for dl-zinc lactate, 18.17; theory for either d- or Z-zinc lactate, 12.90. Polarimetrically, the substance was non-rotatory, both as the zinc salt in neutral solution and as the molybdate complex. The formation of dl-lactate is of some interest. Christensen et al. (12) have described an enzyme of Clostridium, butylicum which catalyzes the racemization of either d- or Z-lactic acid. The forma- tion of dl-lactate by the L. arabinosus preparations might be due to con- tamination with a racemizing enzyme or to formation of racemic lactate from pyruvate (Reaction 4) by the bacterial lactic dehydrogenase.
4 Harary, I., Veiga Salles, J. B., and Ochoa, S., unpublished results.
by guest on September 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

902 BIOSYNTHESIS OF DICARBOXYLIC ACIDS. IV
DISCUSSION
It is hardly necessary to point out that Reaction 1 cannot be a direct decarboxylation of malate to lactate and COZ. The requirement for DPN indicates the occurrence of an oxidation-reduction. In the light of pre- vious work (3, 7) it is apparent that such an oxidation-reduction must in- volve Reactions 3 and 4. Direct spectrophotometric demonstration of Reaction 3 has proved so far impossible, since the purified preparations still contain lactic dehydrogenase, and hence the reduced DPN formed by Reaction 3 is immediately reoxidized by Reaction 4.
An oxidation-reduction similar to the one proposed above has been shown to occur by interaction of the “malie” enzyme of pigeon liver with lactic dehydrogenase (7, 8). Owing to the TPN specificity of the pigeon liver enzyme, this oxidation-reduction is TPN-dependent and its occurrence is possible because lactic dehydrogenase can react with TPN, though only at l/100 to l/150 of the rate at which it reacts with DPN. In this case too, the net result of the TPN-linked dismutation is the conversion of I-malate to lactate and CO2 (7, 13).
It would thus appear that the preparations of L. arabinosus here de- scribed contain a malic enzyme which, except for its DPN (instead of TPN) specificity, is analogous to that of pigeon liver. In both cases, as far as can be determined, the two activities exhibited by each preparation, i.e. ca- talysis of the reversible oxidative decarboxylation of I-malic acid and of the decarboxylation of oxalacetic acid, seem to be associated with one pro- tein or functional protein unit. This is evidenced by constant ratios at var- ious levels of purity (see Table II). It should also be recalled that in L. arabi- nosus both activities appear simultaneously on adaptation to malate (1).
The DPN specificity of the bacterial malic enzyme, indicated by the DPN specificity of the overall reaction (Reaction l), makes its interac- tion with lactic dehydrogenase extremely rapid and efficient. However, a small amount of pyruvate accumulates, as shown in Table III. This is further evidence that Reaction 1 is the combined result of Reactions 3 and 4.
There is little doubt that in the bacterial preparations lactic dehydro- genase has no relation to the “malic” enzyme other than that of a contam- inant, since, as shown in Table II, the ratio of activity of the former to the latter enzyme can be markedly diminished on purification. Moreover, while it is possible to reduce the malate and oxalacetate activities propor- tionately by progressive heat denaturation, the lactic dehydrogenase ac- tivity is affected to a far lesser degree (Table II). As pointed out in the preceding paper (l), lactic dehydrogenase can be demonstrated as a con- stitutive enzyme in L. cm&noms and the total quantities extracted from unadapted and malate-adapted organisms are about the same.
by guest on September 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

S. KORKES, A. DEL CAMPILLO, AND S. OCHOA 903
It has been pointed out elsewhere (3, 8) that the activity of the “malic” enzyme cannot be attributed to the combined activities of a malic dehy- drogenase and an oxalacetic carboxylase. This is true of both the “malic” enzyme of pigeon liver and that of L. arabinosus (8). In the case of the bacterial enzyme, malic dehydrogenase cannot be detected at any stage of purification. The optical test for malic dehydrogenase, based on the re- action oxalacetate + DPN,,d. ;r‘ I-malate + DPN,,. (7), is highly sensitive and valid under our conditions, since the “malic” enzyme in the preparation will not catalyze Reaction 3, i.e. will not reduce DPN,,., in the absence of Mn++. In addition, it has been shown that, while the rate of disappearance of malate at pH 6.0 is equal to that of oxalacetate at pH 4.5 (Table II), at pH 6.0 the rate of oxalacetate decarboxylation is only about one-eighth of the rate of malate dissimilation (see Fig. 2). If two separate enzy- matic steps were involved with free oxalacetate as an intermediate, the rate of disappearance of malate could never be higher than that of oxal- acetate. Also malonate at concentrations at which it strongly inhibits oxalacetate decarboxylation has little or no effect on malate dissimilation by the bacterial enzyme.
Methods
PreparationsHighly purified amorphous fumarase was prepared ac- cording to Laki and Laki (14). An ammonium sulfate fraction from rabbit muscle, kindly supplied by Dr. E. Racker, was used as a source of lactic dehydrogenase. The following substances were prepared as previously described: DPN (7)) TPN (15)) d-malic acid (3). Calcium phosphate gel was prepared as described by Keilin and Hartree (16). dl- and Z-malic acids were obtained commercially. Protamine sulfate was generously sup- plied by E. R. Squibb and Sons. NazCr408 was obtained on allocation from the United States Atomic Energy Commission.
Analytical-Lactic acid was determined by the method of Barker and Summerson (17). I-Malic and pyruvic acids were determined enzymati- tally, with purified pigeon liver “malic” enzyme and lactic dehydrogenase respectively, as previously described (3).
Isotope Experiments--The reaction mixture was similar to that in the manometric tests. The reaction was carried out in a closed system and, at the end of the experimental period, sulfuric acid was tipped in to give 1.0 N final concentration. The COZ was collected in a solution of sodium hydroxide. The sample was alternately flushed with COZ and air until the rinses were free of radioactivity. The precipitated protein was centri- fuged and the supernatant neutralized. A 0.1 cc. aliquot (Sample 1) was plated and its radioactivity measured in order to determine the total fixa- tion of CY402. Another aliquot (Sample 2) was used for the determination of residual fumaric and malic acids with the L. arabinosus enzyme plus
by guest on September 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

904 BIOSYNTHESIS OF DICARBOXYLIC ACIDS. IV
fumarase (4). An aliquot identical to Sample 2 but larger (Sample 3) was run simultaneously with it in a larger vessel containing 1.0 N NaOH in the center well and 10.0 N HzS04 in the side bulb. Incubation of this sam- ple was continued for 30 minutes after evolution of CO1 in Sample 2 had ceased. The acid was tipped in and the vessel was shaken for another 30 minutes to allow for complete absorption of the COz. This COZ is liber- ated from the fl-carboxyl of malate and one of the fumarate carboxyls. The alkali in the center well was diluted as required and 0.1 cc. aliquots were plated directly, dried under an infra-red lamp, and counted immedi- ately to minimize water absorption. A thin window Geiger-Miiller counter was used. All counts were corrected to zero thickness by the method of Libby (18).
SUMMARY
The isolation, partial purification, and characterization of an adaptive enzyme of Lactobacillus arabinosus, catalyzing the oxidative decarboxyla- tion of I-malic acid, are described. The enzyme also catalyzes the decar- boxylation of oxalacetic acid. Both activities seem to be associated with a single protein or functional protein unit. Owing to the presence of lactic dehydrogenase as a contaminant in the preparations, the over-all reaction with malate as substrate is I-malate ~;t lactate + COZ. This reaction, which has been demonstrated to be reversible by the use of isotopic carbon diox- ide, appears to be the result of interaotion between “malic” enzyme, cat- alyzing the oxidative decarboxylation of I-malate to pyruvate and COZ in the presence of DPN and Mn++, and lactic dehydrogenase, catalyzing the reduction of the pyruvate to lactate.
We wish to thank Mr. Isaac Harary for help with some experiments and Mr. Morton C. Schneider for technical assistance.
BIBLIOGRAPHY
1. Blanchard, M. L., Korkes, S., de1 Campillo, A., and Ochoa, S., J. Biol. Chem., 187,875 (1959).
2. Korkes, S., and Ochoa, S., J. Biol. Chem., 176, 463 (1948). 3. Ochoa, S., Mehler, A. H., and Kornberg, A., J. Biol. Chem., 174, 979 (1948). 4. Ochoa, S., Veiga Salles, J. B., and Ortiz, P. J., J. Biol. Chem., 187, 863 (1956). 5. Heidelberger, M., and MacPherson, C. F. C., Science, 97, 405 (1943). 6. Warburg, O., and Christian, W., Biochem. Z., 310, 384 (194142). 7. Mehler, A. H., Kornberg, A:, Grisolia, S., and Ochoa, S., J. Biol. Chem., 174,
961 (1948). 8. Veiga Salles, J. B., and Ochoa, S., J. Biol. Chem., 187, 849 (1956). 9. Wood, H. G., Vennesland, B., and Evans, E. A., Jr., J. BioZ. Chem., 169, 153
(194.5). 10. Utter, M. F., and Wood, H. G., J. Bio(. Chem., 164, 455 (1946).
by guest on September 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

S. KORKES, A. DEL CAMPILLO, AND S. OCHOA 905
11. Vennesland, B., Evans, E. A., Jr., and Altmen, K. I., J. Biol. Chem., 171, 675 (1947).
12. Christensen, W. B., Johnson, M. J., and Peterson, W. H., J. Biol. Chem., 127, 421 (1939).
13. Moulder, J. W., Vennesland, B., and Evans, E. A., Jr., J. Biol. Chem., 160, 305 (1945).
14. Laki, E., and Laki, K., Enzymologia, 9, 1939 (1940-41). 15. Ochoa, S., J. Biol. ‘Chem., 174, 133 (1948). 16. Keilin, D., and Hartree, E. F., Proc. Roy. Sot. London, Series B, 124, 397 (1938). 17. Barker, S. B., and Summerson, W. H., J. Biol. Chem., 138, 535 (1941). 18. Libby, W. F., Ind. and Eng. Chem., Anal. Ed., 19, 2 (1947). 19. Lwoff, A., and Ionesco, H., Compt. rend. Acad., 224, 1664 (1947).
by guest on September 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Severo OchoaSeymour Korkes, Alice del Campillo andLACTOBACILLUS ARABINOSUS
"MALIC" ENZYME FROMPROPERTIES OF AN ADAPTIVE FIXATION: IV. ISOLATION AND
ACIDS BY CARBON DIOXIDE BIOSYNTHESIS OF DICARBOXYLIC
1950, 187:891-905.J. Biol. Chem.
http://www.jbc.org/content/187/2/891.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
alerts to choose from all of JBC's e-mailClick here
tml#ref-list-1
http://www.jbc.org/content/187/2/891.citation.full.haccessed free atThis article cites 0 references, 0 of which can be
by guest on September 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from





























