Embed Size (px)
Citation preview

발생에서 선택적 유전자발현
유전체 등가성(genomic equivalence) : 개별 체세포 핵은 다른 모든 체세포핵과 동일한 염색체(유전자 조합)를 가짐
선택적 유전자 발현(differential gene expression)
1.모든 세포의 핵은 수정란에서 확립된동일한 유전체를 가짐(분화된 세포의 DNA는 동일)
2. 분화된 세포에서 이용되지 않은 유전자는파괴 및 돌연변이 되지 않으며 발현될잠재성 내재
3. 유전체의 적은 부분만 개별 세포에서발현되고 각 세포에서 합성되는 RNA 종류는 세포 종류에 따라 특이적

유전자 발현은 서로 다른 종류의 세포가서로 다른 조합의 단백질을 합성하도록
여러 단계에서 조절
1. 유전자의 선택적 전사 : 유전자를핵RNA로 전사되는 것을 조절
2. 핵RNA의 선택적 가공 : 전사된RNA를 세포질로 이동할 mRNA로되는 것을 조절
3. 전령RNA의 선택적 번역 : 세포질에서mRNA가 단백질로 번역되는 것을 조절
4. 단백질의 선택적 변형 : 단백질을 세포에잔류하거나 또는 기능하도록 조절

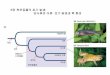
유전체 등가성의 증거✓초파리 염색체 분석에서 처음으로 몸의 모든 세포는 동일한 유전체를 가짐을 밝힘✓포유류 염색체를 김자염로(Giemsa dye)로 염색한 후 관찰 결과 모든 세포의
염색체는 어떤 부위도 없어 지지 않았음✓1952년 브리그스(Briggs)와 킹(King)은 개구리를 이용하여 포배기 세포의 핵을 활
성화된 난자에 이식하여 완벽한 올챙이로 발생 유도(체세포 핵이식(somaticnuclear transfer) 또는 복제(cloning)).
✓1997년 이안 월머트(Ian Wilmut) : 6년생 암양의 젖샘세포(체세포)를 G1로 배양한후 핵을 성숙한 난자(제2감수 분열 중기 핵 제거)와 전기충격으로 세포 흉합 시킨
후 7일간 배양하여 대리모의 암양의 자궁에 이식하여 Dolly를 탄생시킴(그림 2.1)


✓성체 포유류 체세포를 이용하며 현재까지 기니피그, 생쥐, 흰쥐, 고양이, 개, 말, 노새 등을 복제함
✓성체 체세포 핵은 성체로 발생하는데 필요한 모든 유전자를 갖고 있음이 증명됨
✓복제된 동물의 모든 장기(기관)는 성장할 수록 질병에 약함(염색질 사이의메틸화 차이로 기인)
✓복제된 동물이 간혹 동일하지 않을 경우가 발생(염색체 이상이나 환경적 요인으로 인한 변이성 때문)

선택적 유전자 발현발생의 이해
✓유전자의 구조: 활성화 및 불활성화 염색질(chromatin)
✓히스톤(histone)은 유전자 발현의 억제 상태를 담당
✓히스톤 아세틸화(histone actylation)는 전사를 활성화시킴
✓히스톤 메틸화(histone methylation)는 전사를 억제함
✓ 유전자의 구조 : 엑손과 인트론
✓엑손(exon) ⇒ 단백질을 암호화하는 부위
✓인트론(intron) ⇒ 단백질의 아미노산 서열과 무관
✓적혈구 헤모글로빈 단백질을 암호화하는 유전자 구성 요소1. 프로모터 부위(promoter region) : RNA 중합효소의 결합과 그에 따른
전사의 개시 담당2. 전사개시점(transcription initiation site) : RNA 5′말단으로 ACATTTG이며
모자서열이라 함3. 번역개시점(translation initiation site) : ATG이며 mRNA에서 AUG 개시코든

4. 첫 번째 엑손 : 1-30번 아미노산을 암호화하는 90개 염기쌍
5. 첫 번째 인트론 : 130 bp. 아미노산을 암호화하지 않고 mRNA로 가공하고 핵을 빠져 나오는데중요함
6. 2번째 엑손 : 31-104bp7. 2번째 인트론 : 850bp. 단백질의
구조와 아무련 관련이 없음8. 3번째 엑손 : 126bp9. 번역 종료코든(translation stop
codon) : TAA로 mRNA에서UAA 종결 코든
10. 3′비번역부위(3′untranslated region; UTR) : 전사는 되지만
단백질로 번역되지 않음. 아데닐산 중합반응(polyadenylation)에 필요한AATAAA 서열을 가짐. 폴리A꼬리(mRNA 안전성을 부여, mRNA가 핵에서 빠져나 오게 함, mRNA가 단백질로 번역되게 함)
11. 전사종결서열(transcription temination sequence) : 전사는 AATAAA 지점을지나 1000 뉴클레오티드 떨어진 지점에서 종결

핵RNA(nuclear RNA; nRNA) : 1. 최초의 전사산물 2.초기 mRNA라 함 3.모자서열, 5′UTR, 엑손, 인트론, 3′UTR 등을 가짐 4. nRNA가 핵을 빠져 나오기 전에 인트론은 제거되고 남아 있는 엑손은 다 함께 짜깁기 됨

유전자의 구조 : 프로모터와 인핸서✓특정 유전자의 전사되는 시기와 장소의 조절에 필수✓프로모터(promoter) : RNA 중합효소가 전사 개시를 위해 DNA에 결합하는 장소.
✓ mRNA를 합성하는 유전자의 프로모터는 RNA 중합효소가 전사를 개시
하는 장소의 상단부에 위치함.
✓ 이러한 프로모터는 부위를 CpG 섬이라 하며 RNA 중합효소를 불러오고 전사를
개시함
✓ RNA 중합효소 Ⅱ는
모든 프로모터에
동시에 결합하지 못하며
인핸서(enhancer)에서
RNA 중합효소 Ⅱ를 불러와
안정화 시킴.
✓인핸서(enhancer) : 특정 프로모터로
부터 전사의 효율과 속도를 조절하는
DNA 서열. 프로모터가 언제 어디서
사용될지와 얼마나 많은 유전자
산물을 만들지를 알려 줌. 특정 전사인
자 단백질과 결합하여 해당 유전자를
활성화 시킴

✓결합된 전사인자는 보조인자와 결합하여 유전체 부위에 RNA 중합효소의 접근이 가능하도록 뉴클레오솜(nucleosome) 변형효소를 불러와 염색질을 느슨하게 만들어 전사가 일어나게 함✓전사인자는 고리 모양의 다리를 형성하고 프로모터 근처에 위치함. 다리를 형성하는 단백질은nucleosome 변형효소와 TAF(전사결합인자 단백질)를 불러와 RNA 중합효소 Ⅱ를 활성화 시킴(그림 2.6)✓매개자 복합체는 enhancer와 프로모터의 연결enhancer와 프로모터 사이의 다리는 매개자를 다중복합체를 형성하여 발생신호를 enhancer에서RNA 중합효소 Ⅱ로 연결함. 포로모터에 개시준비복합체가 형성되며 코헤신(cohesin)은 전사인자와 결합된 매개인자와 결합함(그림 2.7) ✓매개인자는 프로모터에 RNA 중합효소를 불러오는 것을 도우나 전사를 위해서는 매개인자와RNA 중합효소 Ⅱ 사이의 연결이 끊어 져야 함. RNA 중합효소 Ⅱ의 탈출은 몇몇 전사인자로 이루어진 전사 신장복합체(TEC)에 의해 성취됨. 어떤 경우에는 RNA 중합효소 Ⅱ가 매개자와분리되지 않거나 전사신장억제가 결합하여전사가 중지됨

전사인자의 기능
✓각 집단 내의 전사 인자들은 DNA 결합장소에서 공통된 틀을 가지고 있으며결합장소에 위치한 아미노산의 적은 차이가 서로 다른 DNA 서열을 인식
✓인핸서는 전사인자와 결합함으로 기능
✓각 인핸서는 몇몇 전사인자를 위한 결합장소를 가짐

DNA 메틸화와 전사 조절
✓불활성 유전자의 프로모터는 특정 시토신 염기에서 메틸화되고 메틸화된 시토신은 뉴클레오솜을 안정화시켜 전사 인자의 결합을 차단함
✓시토신의 메틸화 정도는 유전자의 전사 수준을 조절
✓척추동물에서 유전자 프로모터에 메틸화된 시토신의 존재는 해당 유전자의전사 억제와 밀접한 관련이 있음 (그림 2.17)

DNA 메틸화가 전사를 막는 기작
✓인핸서에 전사인자의 결합을 막음(시토신 하나가 메틸화 됨)
✓메틸시토신은 히스톤의 메틸화 또는 탈아세틸화를 촉진하는 단백질을 불러 모아 뉴클레오솜을 안전화 시킴(그림 2.18)

선택적 RNA 가공✓RNA는 활성 단백질을 합성하기 위해
1. 인트론 제거에 따른 전령RNA 가공 2. 핵에서 세포질로 이동3. 단백질 합성 기구에 의해 번역 되어야 함4. 활성을 갖기 위해 번역 후 변형을 거쳐야 함
✓발생과정에서의 조절은 이 모든 단계에서 일어남
✓분화의 본질은 세포의 종류마다 서로 다른 조합의 단백질을 만드는 것
✓선택적 RNA 가공이발생을 조절하는 방식: 1. 핵 전사체가세포질 전령으로가공되는 선택(검열) 2. 서로 다른 잠재적엑손을 사용하여다른 종류의 단백질을
지정하는 전령 RNA전구체의 짜깁기(splicing) - 그림 2.24

선택적 핵RNA 짜깁기를 통한 단백질 집단의 생성
✓nRNA 대체짜깁기(alternative nRNA splicing) : 같은 유전자에서 다양한 종류의단백질을 생성하는 수단. 대부분의 포유류 nRNA는 수많은 엑손을 갖고 있으며 세
포는 종류에 따라 서로 다른 조합의 엑손을 짜깁기 함으로써 여러 종류의 mRNA와그에 따른 서로 다른 단백질을 만들 수 있음
✓사람의 유전자는 대략 95%가 대체짜깁기를 하고 대체 짜깁기는 상대적으로제한된 수의 유전자에서 보다 많은 종류의 단백질 배열을 생성하는 주된 방식
✓초파리 유전체는 14,000개 유전자를 가지고 있으며 단백질의 종류는 이보다 3배많음
✓사람의 유전체는 20,000∼30,000개의 유전자를 가지고 있으나 단백질의 종류는이 보 휠 씬 다양함
✓유전자 수는 사람과 비슷하나 969개 세포로 이루어진 꼬마선충(Caenorhabditis elegans)은 개별 유전자로 1종류의 단백질을 만들지만 사람은 같은 수의 유전자로 대체짜깁기를 통해 다양한 종류의 단백질을 생성함

번역수준에서 유전자 발현의 조절
✓차등적 mRNA 수명 :mRNA의 수명이 길수록 단백질이 더 많이 번역됨mRNA는 특정 세포의 특정 시기에 선택적으로 안정화 됨
✓mRNA 번역의 선택적 억제 :◦ 난자는 종종 수정 이후에 사용할 mRNA를 만들어 저장◦ mRNA는 배란 또는 수정 과정에서 전파되는 이온 신호에 의해 활성화될 때까지
휴면 상태◦mRNA의 일부는 난할과정에서 염색질, 세포막, 세포골격 등의 성분이 될 단백질
암호◦ 일부는 초기 세포분열의 시기를 조절하는 사이클린 단백질 암호화◦ 세포의 운명을 결정하는 단백질을 암호화. 난자에서 대부분의 번역 조절은
억제성

마이크로 RNA : mRNA의 전사와 번역의 특이적 조절인자✓특정 RNA의 번역을 조절하는 가장 효율적인 수단의 하나로 특정 mRNA 부위
에 상보적인 작은 RNA 만드는 것
✓마이크로 RNA가 포유류 심장과 혈구분화에 관여하는 것으로 알려짐
✓마이크로 RNA는 유전자 산물의 양을미세조정하고 제거하는 데 이용
✓3′UTR에 마이크로 RNA의 결합은몇 가지 방식으로 번역 조절 : 1. 결합은 개시인자 또는 리보솜
결합을 막아 번역 개시 차단2. 폴리A꼬리에서 시작하는
핵산 내 가수분해효소를 불러 모음3. 번역 개시는 허용되나 합성된
단백질을 분해하는 단백질분해효소를 불러옴(그림 2.37)

세포질 분포에 따른 RNA 발현의 조절
✓mRNA의 번역 시기뿐만 아니라 RNA 발현 장소도 조절됨
✓mRNA의 분포는 3가지 기작으로 발생 :
1. 확산과 국부적 고정 ⇒ Nanos와 같은 mRNA는 세포질에서 자유롭게 확산되나 초파리 난자의 뒤쪽 극으로 확산될 경우 단백질에 고정되며 이 단백질은 mRNA 번역을 유도함
2. 국부적인 보호 ⇒ Hsp83을 암호화하는 초파리열충격단백질(고열로부터 배아 보호)은 세포질에서 자유롭게이동하나 배아 전 부위에서는 분해되지만 뒤쪽 극에
축적되었을 경우 Hsp83 mRNA가 분해되는 것을 방지함
3. 세포 골격을 따라 능동수송 ⇒ mRNA 분포에 가장 널리이용. mRNA는 3′UTR을 인식하는 특정 단백질의 도움으로 모터단백질과 결합하여 최종 목적지까지 세포골격을 따라 이동. 이들 모터단백질은 디네인(dynein)과키네신(kinesin)임(그림 2.38)

유전자 발현의 번역 후 조절
개별 단백질은 수 많은 다른 단백질과의 관계 속에 통합(복합 생태계)
1. 일부 새로 형성된 단백질은 특정 억제 부위를 제거하지 않으면 불활성
2. 일부 단백질은 기능을 위해 세포 내 특정 목적지로 보내져야 함
3. 일부 단백질은 종종 막, 리보솜, 핵, 미토콘드리아 등 특정 부위에 격리되어야 함
4. 일부 단백질은 이온(ex; Ca2+)과 결합할 때까지 또는 인산기와 초산기의공유 첨가에 의해 변형될 때 까지 불활성









![]YZ ^ `` 89 ! de @09A BC i=contents.kocw.net/KOCW/document/2014/Seowon/Leehongwoo/... · 2016. 9. 9. · ""ÖC](https://img.pdfslide.net/doc/110x75/5fe8d5f7575e0221aa0ddce9/yz-89-de-09a-bc-i-2016-9-9-c-.jpg)

![345])contents.kocw.net/KOCW/document/2014/Seowon/KimJinho/02.pdf1차대전의기원 홉슨의자유주의 2/48 • 제국주의는국내에서의잉여(국내에서는소비할수없는것들)를유출하기위해](https://img.pdfslide.net/doc/110x75/5e51d0cbc0abc67a0d123fa2/345-1eoee-oe-248-a-oeeeeeoeeeoeeoeeeefeeoeoeeoe.jpg)





![10장 기둥 해석 및 설계 - KOCWcontents.kocw.net/KOCW/document/2014/Seowon/Leehongwoo/... · 2016-09-09 · XvY#pAB,!0Q,Ê,,J nËÌ1 v " ° ±,z6RSX Y XmY]AB%#pAB,¶ 89 7¾](https://img.pdfslide.net/doc/110x75/5e830588e1eb6f56d7659bb6/10-ee-e-e-2016-09-09-xvypab0qj-noe1-v-.jpg)