Embed Size (px)
Citation preview

SNP – Epilepsies. Physiopathologie, bases thérapeutiques des épilepsies
1/20
22/12/2017 BREULEUX Claire D1 CR : AMMAR Mélanie SNP Dr Lagarde 20 pages
Epilepsies. Physiopathologie, bases thérapeutiques des épilepsies
Plan
A- Généralités I- Définition de l’épilepsie II- Notions physiopathologiques
B- Mécanismes de l’épilepsie : excitabilité neuronale
I- Excitabilité intrinsèque a. Canaux ioniques potentiel-dépendants en physiologie b. Canaux ioniques potentiel-dépendants dans l’épilepsie
II- Excitabilité extrinsèque a. Récepteurs au glutamate b. Récepteurs au GABA c. Sommation des synapses
C- Conséquences de l’épilepsie et mécanismes d’aggravation
Dans ce cours nous verrons la physiologie et la physiopathologie de l’épilepsie. Nous procéderons en faisant des rappels sur la physiologie de base du système nerveux central puis en regardant ce qui dysfonctionne dans le cas de l’épilepsie.
CR : Le prof a mis son diaporama en ligne sur AmeTice pour les personnes souhaitant voir plus d’images d’EEG car on n’a pas pu tout mettre, le cours faisant déjà 20 pages… ou si vous voulez tout simplement voir les diapos en couleur.
A- Généralités I- Définition de l’épilepsie
Il s’agit d’une pathologie neurologique fréquente qui touche environ 50 000 000 de personnes qui ont une épilepsie active dans le monde, et environ 700 000 personnes en France. A peu près 1% de la population pourra faire une crise d’épilepsie dans sa vie sans forcément être épileptique après.
Si on simplifie, l’épilepsie est une pathologie des rythmes cérébraux.
Un peu comme en cardiologie on peut avoir des troubles du rythme cardiaque, en neurologie on peut avoir des troubles des rythmes cérébraux. Si on enregistre par exemple chez l’animal ce qu’il se passe quand il fait une crise on voit une décharge rythmique et soutenue de certains neurones. Il s’agit de la définition physiologique de la crise d’épilepsie.
SNP – Epilepsies. Physiopathologie, bases thérapeutiques des épilepsies
2/20
La définition clinique se base sur les manifestations qui sont :
- Transitoires (contrairement à d’autres maladies neurologiques qui donneront des déficits persistants) - Une activité neuronale excessive - Une activité neuronale trop synchronisée (synchronie anormale)
II- Notions physiopathologiques
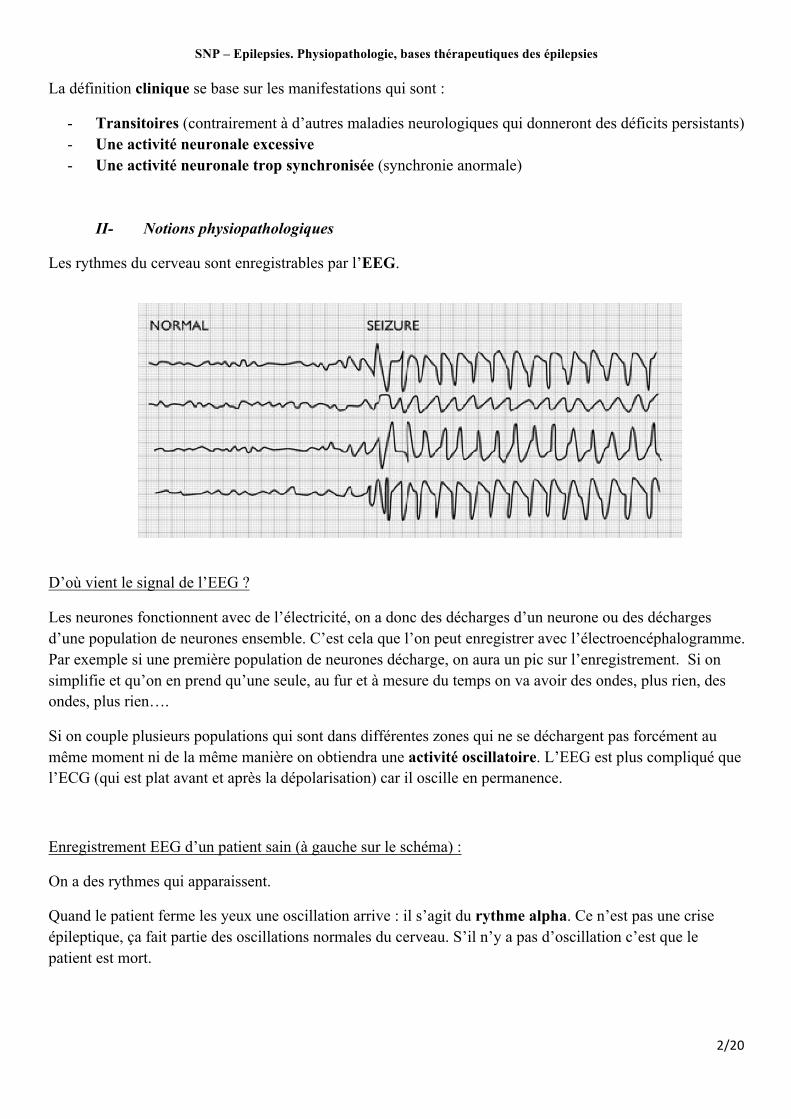
Les rythmes du cerveau sont enregistrables par l’EEG.
D’où vient le signal de l’EEG ?
Les neurones fonctionnent avec de l’électricité, on a donc des décharges d’un neurone ou des décharges d’une population de neurones ensemble. C’est cela que l’on peut enregistrer avec l’électroencéphalogramme. Par exemple si une première population de neurones décharge, on aura un pic sur l’enregistrement. Si on simplifie et qu’on en prend qu’une seule, au fur et à mesure du temps on va avoir des ondes, plus rien, des ondes, plus rien….
Si on couple plusieurs populations qui sont dans différentes zones qui ne se déchargent pas forcément au même moment ni de la même manière on obtiendra une activité oscillatoire. L’EEG est plus compliqué que l’ECG (qui est plat avant et après la dépolarisation) car il oscille en permanence.
Enregistrement EEG d’un patient sain (à gauche sur le schéma) :
On a des rythmes qui apparaissent.
Quand le patient ferme les yeux une oscillation arrive : il s’agit du rythme alpha. Ce n’est pas une crise épileptique, ça fait partie des oscillations normales du cerveau. S’il n’y a pas d’oscillation c’est que le patient est mort.
SNP – Epilepsies. Physiopathologie, bases thérapeutiques des épilepsies
3/20
Pendant une crise (à droite sur le schéma) :
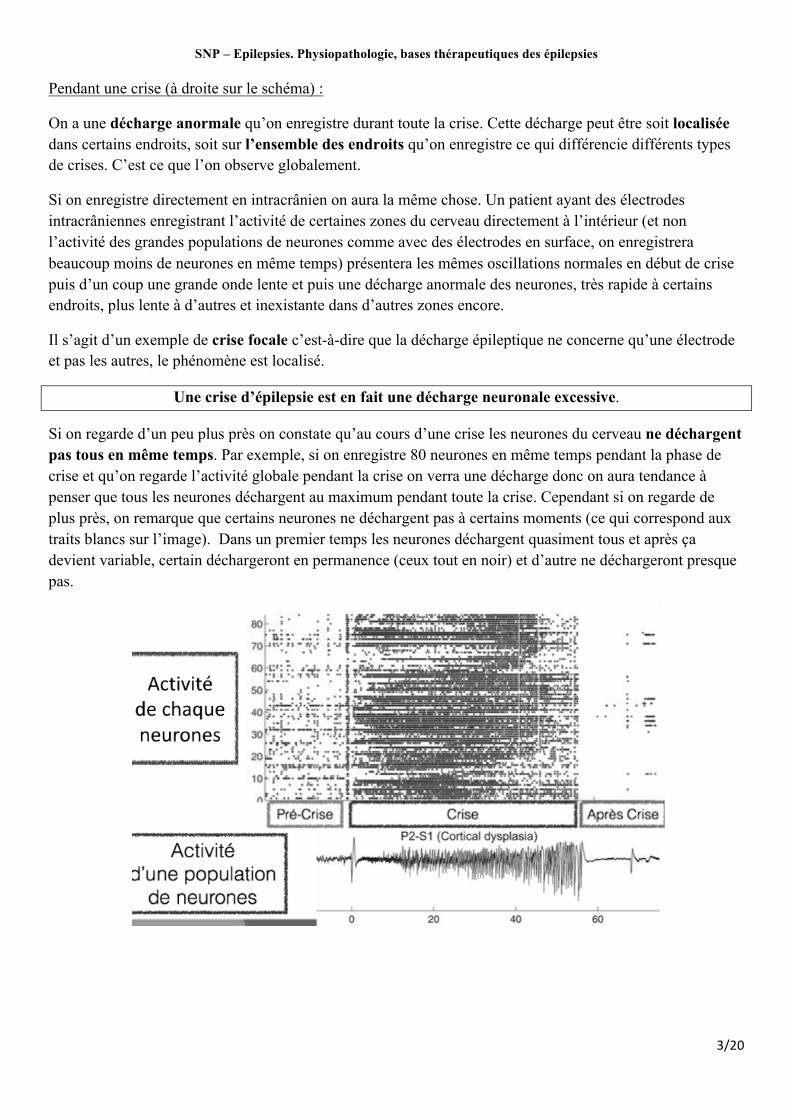
On a une décharge anormale qu’on enregistre durant toute la crise. Cette décharge peut être soit localisée dans certains endroits, soit sur l’ensemble des endroits qu’on enregistre ce qui différencie différents types de crises. C’est ce que l’on observe globalement.
Si on enregistre directement en intracrânien on aura la même chose. Un patient ayant des électrodes intracrâniennes enregistrant l’activité de certaines zones du cerveau directement à l’intérieur (et non l’activité des grandes populations de neurones comme avec des électrodes en surface, on enregistrera beaucoup moins de neurones en même temps) présentera les mêmes oscillations normales en début de crise puis d’un coup une grande onde lente et puis une décharge anormale des neurones, très rapide à certains endroits, plus lente à d’autres et inexistante dans d’autres zones encore.
Il s’agit d’un exemple de crise focale c’est-à-dire que la décharge épileptique ne concerne qu’une électrode et pas les autres, le phénomène est localisé.
Une crise d’épilepsie est en fait une décharge neuronale excessive.
Si on regarde d’un peu plus près on constate qu’au cours d’une crise les neurones du cerveau ne déchargent pas tous en même temps. Par exemple, si on enregistre 80 neurones en même temps pendant la phase de crise et qu’on regarde l’activité globale pendant la crise on verra une décharge donc on aura tendance à penser que tous les neurones déchargent au maximum pendant toute la crise. Cependant si on regarde de plus près, on remarque que certains neurones ne déchargent pas à certains moments (ce qui correspond aux traits blancs sur l’image). Dans un premier temps les neurones déchargent quasiment tous et après ça devient variable, certain déchargeront en permanence (ceux tout en noir) et d’autre ne déchargeront presque pas.

SNP – Epilepsies. Physiopathologie, bases thérapeutiques des épilepsies
4/20
L’épilepsie, c’est-à-dire les crises ou la répétition des crises, a beaucoup de causes très variables :
- Une agression. On peut considérer que tout cerveau peut faire une crise s’il a subi une agression suffisante (par exemple un Stroke c’est-à-dire un AVC) car il y aura une agression tissulaire des neurones qui deviennent alors hyperexcitables. - Des mutations génétiques sur certains gènes qui vont permettre de contrôler l’excitabilité du neurone. - Des anomalies génétiques avec plusieurs variations (et non pas sur un seul gène) donnant une susceptibilité qui, suite à un facteur déclenchant, entraineront une crise (l’épilepsie est retrouvée dans beaucoup de maladies neurologiques qui vont déclencher la crise. Une crise peut être déclenchée par beaucoup de choses).
Les symptômes d’une crise d’épilepsie sont aussi très variables ce qui complique le diagnostic. Il y a deux grands types de crises d’épilepsie :
- Les types généralisés où la décharge va toucher un grand nombre de zones ou d’aires cérébrales en même temps (grosso modo tout le cerveau va décharger plus ou moins en même temps). Il en existe plusieurs types : les tonico-cloniques (celle qu’on a tous en tête quand on pense à l’épilepsie) et les absences (arrêt du contact, le patient perd un peu connaissance transitoirement sans qu’il y ait de mouvements pendant une fraction de seconde puis il récupère) - Les crises focales (ou partielles) où la décharge part d’une zone avant de se propager à d’autres zones. Elles sont plus compliquées car les symptômes sont multiples en fonction des aires concernées où la crise débute et se propage. Par exemple, une crise dans le cortex visuel entrainera des manifestations visuelles, une crise dans le cortex moteur entrainera des secousses motrices controlatérales, une crise d’autres zones pourra entrainer des composantes émotionnelles etc (pas la peine de retenir les symptômes, on les reverra plus tard dans nos études). Autre exemple, un patient ayant une décharge dans des structures impliquées dans les émotions (l’amygdale) aura une peur, puis ça se propagera dans des zones gérant l’audition ce qui entrainera une sensation d’hallucination auditive et progressivement ça se propagera aux aires du langage avec un trouble du langage.

SNP – Epilepsies. Physiopathologie, bases thérapeutiques des épilepsies
5/20
B- Mécanismes de l’épilepsie : excitabilité neuronale
Le système nerveux est composé de milliards de neurones et de cellules de soutien. Les neurones sont d’abord organisés en micro-réseau c’est-à-dire qu’il y a plusieurs neurones qui vont communiquer ensemble à l’échelle de quelques centaines et à l’échelle du millimètre, puis en macro-réseau c’est-à-dire que les différentes aires du cerveau vont aussi communiquer entre elles à l’échelle du centimètre pour faire certaines fonctions. La crise va donc s’organiser à travers toutes ces échelles.
Au niveau du neurone, l’épilepsie représente une hyperexcitabilité c’est-à-dire que les neurones sont trop excitables. Cette hyperexcitabilité vient de plusieurs facteurs que nous allons détailler.
On rappelle que les neurones fonctionnent avec de l’activité électrique : si on enregistre l’activité d’un neurone et qu’on lui fait une stimulation on aura un potentiel d’action qui est une réponse électrique à la stimulation. L’épilepsie va toucher cela aussi. Par exemple si on enregistre un neurone pyramidal et qu’on le dépolarise on pourra enregistrer les bouffées de potentiel d’action.
La membrane plasmatique des neurones est imperméable aux ions métaux ce qui signifie qu’il ne peut pas y avoir d’échanges à travers la membrane d’ions chargés positivement ou négativement.
En théorie il ne serait donc pas possible de faire une décharge de la membrane. Ceci est responsable d’un gradient ionique car on a des concentrations de certains ions différentes à l’intérieur et à l’extérieur du neurone (beaucoup plus de Cl- et de Na+ à l’extérieur, beaucoup plus de K+ à l’intérieur et un peu plus de Ca2+ à l’extérieur). On a donc une différence de potentiel très négative dans le neurone à -80mV : le neurone de base est hyperpolarisé, chargé très négativement. Les ions auraient tendance à passer à travers la membrane s’ils le pouvaient : c’est la force motrice du gradient ionique.

SNP – Epilepsies. Physiopathologie, bases thérapeutiques des épilepsies
6/20
Cette force motrice est importante car si les concentrations étaient toutes égales, le potentiel du neurone serait à 0 et il n’y aurait aucune force motrice pour que les ions entrent ou sortent et il n’y aurait donc aucune chance de dépolariser ou d’hyperpolariser le neurone.
Ce qui permet au neurone de fonctionner ce sont les canaux ioniques. Sans eux il ne pourrait pas y avoir d’échanges et donc de changement de potentiel. Ils permettent le passage des ions dans un sens ou dans un autre.
/ !\ Ici on parle bien des canaux et pas des pompes ! Les canaux permettent un passage uniquement passif des ions dans le sens du gradient c’est-à-dire que les ions n’iront jamais à contresens de la concentration. Si on a plus de Na+ à l’extérieur qu’à l’intérieur de la cellule, il va toujours rentrer dans la cellule par les canaux et ne pourra pas sortir de la cellule par ces canaux. Les pompes consomment de l’énergie pour permettre de faire sortir, même contre le gradient, certains ions. On ne les détaillera pas ici mais elles sont importantes pour revenir à l’état basal (remettre les concentrations à l’état basal).
Les canaux sont nombreux (canaux-sodium, canaux chlore, canaux potassiques et canaux calciques). Il y a deux types de canaux :
- Dépendants du potentiel - Dépendants de certaines substances qui vont se fixer dessus (le plus souvent des
neurotransmetteurs) ou ionotropiques

SNP – Epilepsies. Physiopathologie, bases thérapeutiques des épilepsies
7/20
I- Excitabilité intrinsèque
a. Canaux ioniques potentiel-dépendants en physiologie
Ils sont importants dans l’excitabilité intrinsèque du neurone c’est-à-dire la capacité du neurone à créer un potentiel d’action ou pas, et à en créer « trop » s’il est hyperexcitable. Ils vont s’ouvrir quand il y a un changement du gradient membranaire des neurones. Le gradient va remonter de -80mV à -40mV par exemple ce qui va entrainer leur ouverture.
Il y en a plusieurs : au potassium, au sodium, au calcium et au chlore. Les ions suivent toujours le gradient ionique puisqu’il ne s’agit pas de pompes.
On a le canal qui est fermé au potentiel de base de la membrane (par exemple -80mV), il ne laisse passer aucun ion. On a donc une différence de potentiel de membrane qui apparait (par exemple il passe ici de -80 à 0mV) et le canal va donc s’ouvrir, permettant l’entrée des ions. Si par exemple il s’agit d’ions sodium cela va dépolariser le neurone et le potentiel de membrane va monter en positif. Une fois ouvert, le canal va, à un moment donné, s’inactiver.
Exemple des canaux-sodium : Quand le potentiel de membrane est à -70mV les canaux-sodium sont fermés. Il y a une possibilité d’ouverture s’il y a une dépolarisation c’est-à-dire qu’on passe de -70 à -50mV ou de -70 à +20mV. Le « pont » qui fermait le canal va s’ouvrir et permettre l’entrée de sodium. Automatiquement, quand le potentiel passe de +30 à -70mV il y a fermeture du canal par un autre mécanisme. On a une période réfractaire où le canal n’est plus capable de s’ouvrir. Il s’agit d’un mécanisme de protection pour que le neurone ne décharge pas en permanence de manière anormale.
Ce n’est pas la peine de retenir ce qu’il se passe pour chacun des canaux, c’est le concept qui est important
Ce mécanisme est important pour générer le potentiel d’action et le propager le long de l’axone. Il est également mis en jeu quand le neurotransmetteur est libéré au bout de la terminaison de l’axone : des canaux vont s’ouvrir ce qui va permettre la libération du neurotransmetteur dans la synapse.
SNP – Epilepsies. Physiopathologie, bases thérapeutiques des épilepsies
8/20
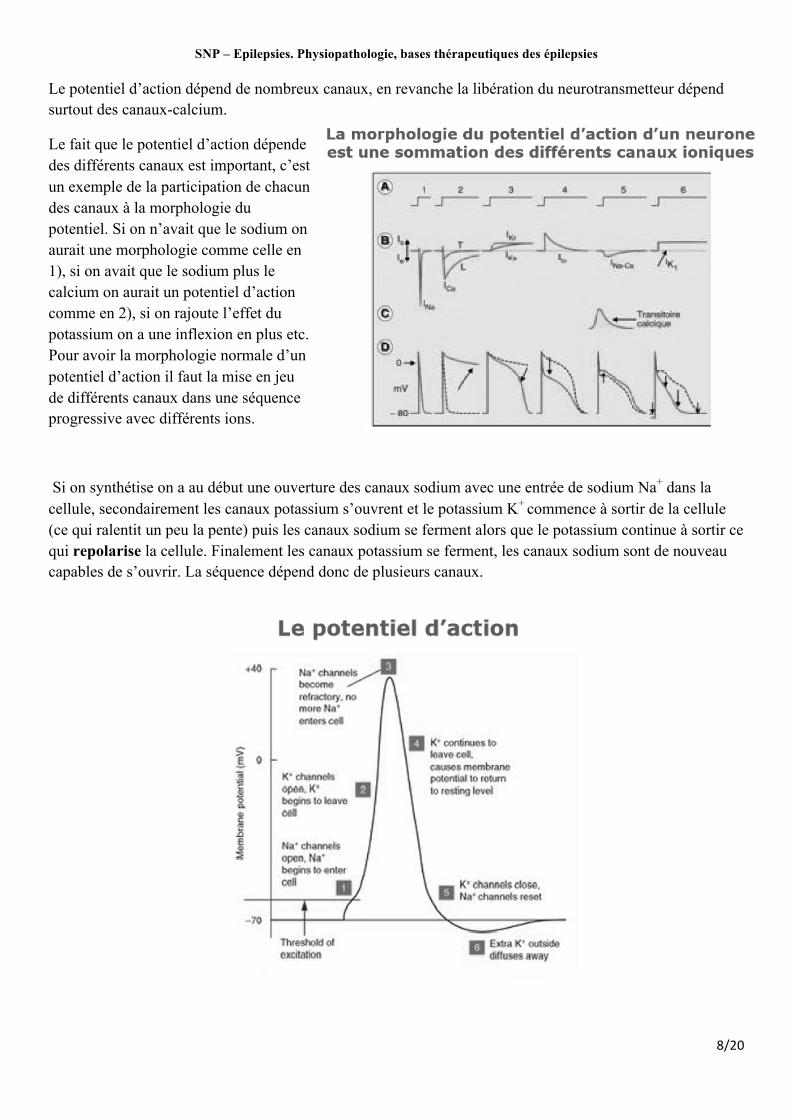
Le potentiel d’action dépend de nombreux canaux, en revanche la libération du neurotransmetteur dépend surtout des canaux-calcium.
Le fait que le potentiel d’action dépende des différents canaux est important, c’est un exemple de la participation de chacun des canaux à la morphologie du potentiel. Si on n’avait que le sodium on aurait une morphologie comme celle en 1), si on avait que le sodium plus le calcium on aurait un potentiel d’action comme en 2), si on rajoute l’effet du potassium on a une inflexion en plus etc. Pour avoir la morphologie normale d’un potentiel d’action il faut la mise en jeu de différents canaux dans une séquence progressive avec différents ions.
Si on synthétise on a au début une ouverture des canaux sodium avec une entrée de sodium Na+ dans la cellule, secondairement les canaux potassium s’ouvrent et le potassium K+ commence à sortir de la cellule (ce qui ralentit un peu la pente) puis les canaux sodium se ferment alors que le potassium continue à sortir ce qui repolarise la cellule. Finalement les canaux potassium se ferment, les canaux sodium sont de nouveau capables de s’ouvrir. La séquence dépend donc de plusieurs canaux.

SNP – Epilepsies. Physiopathologie, bases thérapeutiques des épilepsies
9/20
a. Canaux ioniques potentiel-dépendants dans l’épilepsie
Dans l’épilepsie, tous ces canaux-là peuvent être anormaux. L’exemple le plus simple est certaines épilepsies génétiques. On a une mutation sur un ou plusieurs canaux qui sont dépendants du potentiel qui va par exemple créer un canal qui va s’ouvrir trop tôt ou pas se refermer suffisamment tôt.
Exemple : Un canal sodium (responsable de la dépolarisation) qui s’ouvre trop tôt, trop longtemps ou n’ayant pas d’état réfractaire, le neurone va se dépolariser beaucoup plus facilement et au lieu de faire un potentiel d’action en réponse à une stimulation, il en fera dix. Le neurone décharge trop et déclenche ainsi une crise.
Ceci est possible pour tous les canaux en théorie. Il y a certaines maladies génétiques qui sont liées à ces mutations-là sur les canaux sodiques, d’autres liées à des mutations sur les canaux calciques (permettant de libérer les neurotransmetteurs dans la fente synaptique) ce qui va entrainer trop de libération de neurotransmetteurs excitateurs et donc trop de décharge et d’autres encore liées à des mutations sur les canaux potassiques (ne pas retenir les noms).
Cependant on peut avoir le même phénomène sans mutation génétique c’est-à-dire quand il y a eu une agression avec des crises qui se mettent en place. Le processus épileptique va pérenniser cela car il entraine une répartition différente des canaux sur la membrane qui vont agir différemment.
En laboratoire on peut prendre des cellules de culture de neurones et leur appliquer un médicament ou une substance, ici la Vératridine, qui va activer les canaux sodiques. Au lieu d’avoir un seul potentiel d’action suite à une stimulation, on a une réponse de plusieurs potentiels d’action répétés et si on enregistre toute la population on obtient quelque chose qui ressemble fortement à une crise d’épilepsie. C’est la démonstration expérimentale : si on active trop les canaux sodiques on crée au niveau des neurones quelque chose qui ressemble à une crise c’est-à-dire une décharge excessive des neurones.
Cet exemple pathologique sert également en thérapeutique.

SNP – Epilepsies. Physiopathologie, bases thérapeutiques des épilepsies
10/20
On a ici une image de neurone épileptique (A) et à coté une image de neurone épileptique traité par la Carbamazépine (B) qui est un bloqueur des canaux sodiques. C’est un médicament contre l’épilepsie. Suite à l’application du médicament, au lieu d’avoir une dizaine de potentiels d’action pour une seule stimulation on en aura uniquement cinq. Si on fait un graphique où on représente l’excitabilité, chez les épileptiques c’est 100%, chez les épileptiques traités c’est clairement diminué. Ceci est la base des modes d’action de beaucoup de médicaments antiépileptiques qui vont agir sur les canaux sodiques, calciques, potassiques pour diminuer ou augmenter (selon les canaux) leur activité.
Cela concerne bien les canaux dépendants du potentiel et donc l’excitabilité intrinsèque du neurone.
II- Excitabilité extrinsèque
Il existe aussi une excitabilité extrinsèque : le neurone ne fonctionne pas tout seul, il est également contrôlé par ce qu’il reçoit des autres neurones et par son environnement, ce qui va passer notamment par les récepteurs ionotropiques.
SNP – Epilepsies. Physiopathologie, bases thérapeutiques des épilepsies
11/20
Les récepteurs ionotropiques sont les canaux qui ne sont pas dépendants du potentiel : on peut dépolariser le neurone, ils ne s’ouvriront pas quel que soit le niveau du potentiel de membrane. Ils attendent que des substances viennent se fixer pour ouvrir ou fermer le canal : ils sont dépendants de substances. Par exemple ici on a un canal qui va laisser entrer les ions positifs dans la cellule quand on lui applique une certaine substance ce qui va dépolariser le neurone. Au contraire, d’autres laissent entrer les charges négatives et vont inactiver le neurone c’est-à-dire l’hyperpolariser.
Il y a deux grands systèmes : (à retenir)
- Le système excitateur du glutamate (qui est le neurotransmetteur excitateur majoritaire dans le cerveau). Il fait entrer des ions sodium (et sortir quelques ions potassium) et donc excite le neurone - Le système inhibiteur du GABA qui fait entrer des ions chlore (chargés négativement) ce qui hyperpolarise le neurone et donc l’inhibe.
Il y a plusieurs types de récepteurs selon ce qu’ils reçoivent. Ici nous détaillerons uniquement le glutamate et le GABA mais il faut savoir qu’il y a d’autres substances qui existent (la sérotonine, l’acétylcholine, la nicotine, la glycine, l’ATP…) et qu’il y a des récepteurs spécifiques à ces substances-là qui existent.
Ne pas retenir tout le tableau ci-contre
Le système excitateur du glutamate :
Quand il se fixe il y a une entrée des ions sodium ce qui diminue l’hyperpolarisation (le potentiel remonte vers le 0 voire devient positif). On rapproche donc le neurone du seuil du potentiel d’action.
Si ce seuil est dépassé le canal sodium va s’ouvrir, on aura encore plus de sodium qui entrera et cela va finir par faire un potentiel d’action. Le glutamate rend donc le neurone plus excitable.
Le système inhibiteur du GABA :
Quand il se fixe il y a une entrée des ions chlore ce qui hyperpolarise le neurone. Si on est à -80mV et qu’on applique le GABA on passera à -100mV. On a donc encore moins de chances d’ouvrir les canaux sodium pour faire un potentiel d’action. Le GABA rend donc le neurone encore moins excitable, il est encore plus loin du seuil nécessaire à l’ouverture des canaux.
Cela va permettre aux neurones de communiquer les uns avec les autres au niveau des synapses c’est-à-dire qu’ils peuvent se faire passer une information excitatrice ou inhibitrice. Il s’agit de l’excitabilité extrinsèque : le neurone ne le contrôle pas complètement car cela dépend de son environnement neurochimique qui est à l’extérieur.

SNP – Epilepsies. Physiopathologie, bases thérapeutiques des épilepsies
12/20
a. Récepteurs au glutamate
Il y a trois grands types de récepteurs au glutamate :
- NMDA - AMPA - Kainate
Ils reçoivent tous le glutamate mais fonctionnent un peu différemment. Ils sont essentiellement situés au niveau post-synaptique : ils reçoivent de l’information du neurone pré-synaptique.
De base ils sont fermés à cause de liens chimiques. Lorsqu’on a la mise en contact avec du glutamate, on a une ouverture du canal, un changement de conformité 3D qui permet l’entrée des ions. Si on enregistre le neurone on verra une sorte de décharge électrique au moment où on applique le glutamate.
Finalement le récepteur au glutamate va spontanément se refermer et s’inactiver après coup.
Les AMPA ont le mode de fonctionnement le plus simple. Ils vont spontanément s’ouvrir et faire rentrer des choses dès qu’il y a contact avec du glutamate.
Les NMDA vont être un peu plus lents car ils ont du magnésium qui bloque le canal et qui empêche les ions de rentrer. Il faudra que le neurone soit déjà un peu dépolarisé pour qu’ils s’ouvrent.
Ainsi si on a un neurone à l’état de base et qu’on applique du glutamate, les seuls récepteurs qui pourront s’ouvrir seront les AMPA car ce sont ceux qui se mettent en route en premier et indépendamment du potentiel de membrane. Tant que le neurone sera hyperpolarisé les NMDA ne s’ouvriront pas. Une fois que le neurone a commencé à se dépolariser le magnésium va sortir et débloquer le canal ce qui permet de faire entrer les ions excitateurs. Ils s’activeront donc secondairement.
On a donc une excitation rapide avec les AMPA et une excitation plus lente avec les NDMA.
Si on applique à un neurone normal un agoniste des récepteurs glutamatergique kainate (donc de l’acide kainique qui permet d’activer les récepteurs au kainate) le neurone se met à décharger et on obtient quelque chose qui ressemble clairement à une crise d’épilepsie.
Ce principe existe dans la vraie vie dans les épilepsies liées aux tumeurs cérébrales : si on mesure la concentration du glutamate dans le milieu extracellulaire, il est trop élevé car les cellules tumorales vont augmenter cette concentration. Ceci est responsable des crises d’épilepsies et de l’hyperexcitabilité.
Il y a des médicaments qui agissent sur ces mécanismes-là, il existe des bloqueurs des canaux AMPA.

SNP – Epilepsies. Physiopathologie, bases thérapeutiques des épilepsies
13/20
b. Récepteurs au GABA
On retrouve le même principe que pour les autres : il faut que quelque chose se fixe dessus pour qu’il s’ouvre. Il y a un site de fixation du neurotransmetteur GABA, des sites de fixation des médicaments à d’autres endroits comme les barbituriques qui sont des médicaments antiépileptiques, les benzodiazépines qui sont entre autres antiépileptiques et des sites de fixation des stéroïdes qui sont plutôt anti-inflammatoires. Ces sites de fixation sont différents, on peut donc avoir sur un même canal à la fois l’influence du GABA et du médicament. Ils ne sont pas en compétition.
Un autre exemple de décharge que l’on peut induire : on applique un antagoniste aux récepteurs GABA donc quelque chose qui le bloque (de la même façon que lorsqu’on applique un agoniste des kainates). Par exemple avec de la bicuculline qui est un antagoniste des récepteurs GABA-A on obtient une décharge.
En pathologie on peut imaginer que si on manque de GABA on aura tendance à faire trop de décharges.
Un autre exemple d’application des médicaments antiépileptiques : on a un neurone normal, légèrement hyperpolarisé. Si on applique du GABA le neurone va commencer à s’hyperpolariser encore plus, pas en permanence mais de manière un peu répétitive.
Si on applique du diazépam (c’est-à-dire du Valium ®) qui est une benzodiazépine et un médicament antiépileptique on obtient encore plus de dépolarisation, l’effet du GABA est potentialisé.
Si on applique du Phénobarbital qui est un autre médicament antiépileptique on obtient le même effet. Il y a donc des médicaments antiépileptiques qui ont pour action de potentialiser l’effet du GABA sur les récepteurs et donc d’hyperpolariser les neurones. Les neurones épileptiques étant trop excitables, ces médicaments les ramènent à une excitabilité qui est normale.

SNP – Epilepsies. Physiopathologie, bases thérapeutiques des épilepsies
14/20
Il faut que le GABA se fixe sur le récepteur pour avoir l’action des médicaments. Appliqués seuls, ces médicaments ne vont pas ouvrir le canal car ils potentialisent l’effet du GABA. S’il n’y a pas de GABA, il n’y aura donc pas d’effet. Ceci est différent du cas des glutamates qui ouvrent le canal spontanément pour le kainate par exemple.
c. Sommation des synapses
Dans la vraie vie toute terminaison neuronale a plusieurs canaux, on a donc la combinaison de plusieurs mécanismes ensembles. Ceci participe beaucoup à la spécialisation des synapses. Les neurones communiquent grâce à des synapses. Il en existe deux grands types :
- Les synapses inhibitrices relarguant essentiellement du GABA et ayant essentiellement des récepteurs au GABA. Un premier neurone va inhiber l’action d’un deuxième neurone et éviter qu’il ne décharge. On a un potentiel post synaptique inhibiteur donc plutôt antiépileptique. Par exemple, si on stimule un neurone qui a une synapse avec un deuxième neurone, on obtient un potentiel d’action et le deuxième neurone va passer de -55 à -90Mv (s’hyperpolariser). En activant cette synapse là on inhibe le deuxième neurone. - Les synapses excitatrices qui donnent un potentiel post synaptique excitateur. On a activé un premier neurone et au lieu d’avoir une augmentation de la polarisation on a une dépolarisation du deuxième neurone. Si ce deuxième neurone est suffisamment dépolarisé on peut avoir un potentiel d’action. Si la dépolarisation n’est pas suffisante il n’y aura pas de potentiel d’action et le neurone va revenir au même temps. Ceci est médié plutôt par les récepteurs AMPA et NMDA. La réponse du deuxième neurone dépend du nombre de synapses excitatrices et du nombre de synapses inhibitrices qui vont faire contact avec lui. Si on simplifie avec une excitatrice et une inhibitrice on va peut-être rester à 0 finalement.
Il y a donc différents types de neurones qui vont être plutôt excitateurs ou inhibiteurs dans le système nerveux. Si on simplifie il y en a deux grands types :
- Les cellules pyramidales qui sont des neurones qui vont aller exciter les neurones avec lesquels ils sont en contact. Ils vont avoir des synapses excitatrices et des synapses glutamatergiques pour activer les choses. Ils sont majoritaires et constituent environ 80% des neurones. Ils ont une morphologie très étendue, c’est une cellule classique du cortex. Ces neurones-là font plutôt des connexions à longue distance (jusqu’à 1cm). - Il y a deux types de neurones qui sont inhibiteurs : les interneurones qui ont des connexions locales et constituent 20% des neurones. La réponse d’un neurone va dépendre d’à quel point il est en connexion avec des excitateurs (donc des synapses glutamatergiques) et d’à quel point il est en connexion avec des inhibiteurs c’est-à-dire des synapses GABAergiques. Un neurone reçoit l’information de mille neurones environ. Sur le schéma on peut voir toutes les terminaisons synaptiques qu’il peut y avoir sur le corps cellulaire d’un neurone.
Dans la vraie vie c’est plus compliqué, on a une sommation des informations que le neurone reçoit c’est-à-dire que les informations vont se combiner.
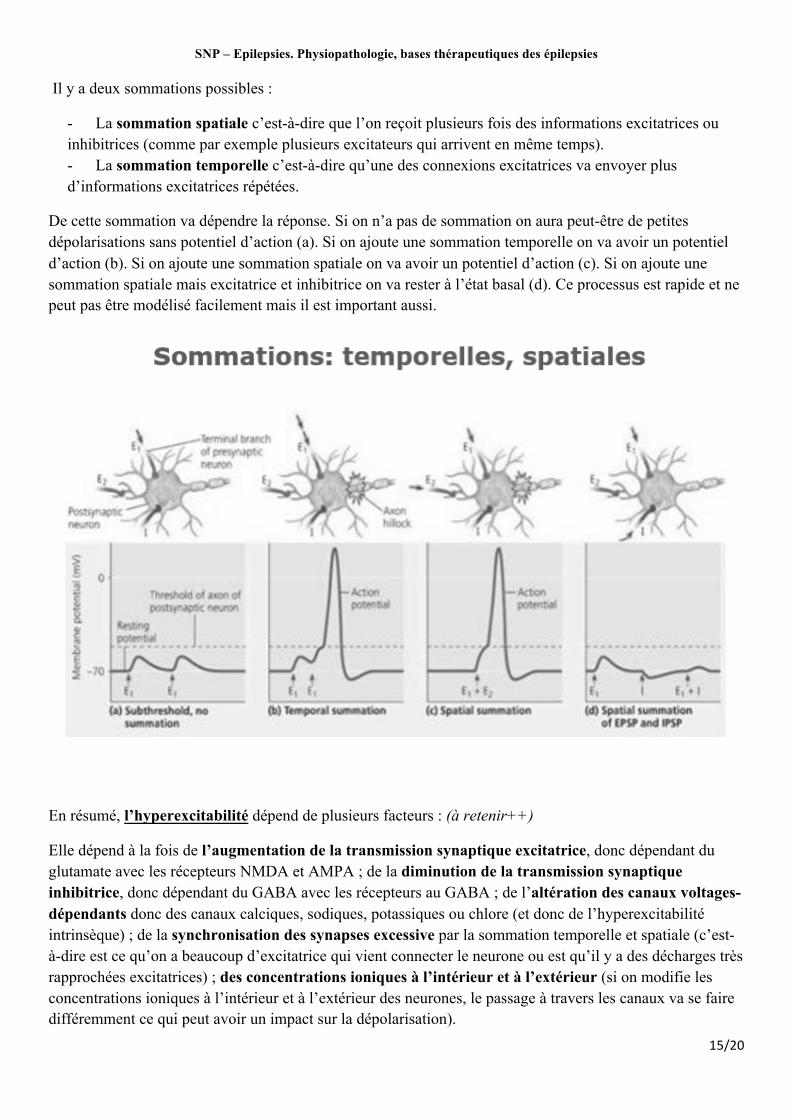
SNP – Epilepsies. Physiopathologie, bases thérapeutiques des épilepsies
15/20
Il y a deux sommations possibles :
- La sommation spatiale c’est-à-dire que l’on reçoit plusieurs fois des informations excitatrices ou inhibitrices (comme par exemple plusieurs excitateurs qui arrivent en même temps). - La sommation temporelle c’est-à-dire qu’une des connexions excitatrices va envoyer plus d’informations excitatrices répétées.
De cette sommation va dépendre la réponse. Si on n’a pas de sommation on aura peut-être de petites dépolarisations sans potentiel d’action (a). Si on ajoute une sommation temporelle on va avoir un potentiel d’action (b). Si on ajoute une sommation spatiale on va avoir un potentiel d’action (c). Si on ajoute une sommation spatiale mais excitatrice et inhibitrice on va rester à l’état basal (d). Ce processus est rapide et ne peut pas être modélisé facilement mais il est important aussi.
En résumé, l’hyperexcitabilité dépend de plusieurs facteurs : (à retenir++)
Elle dépend à la fois de l’augmentation de la transmission synaptique excitatrice, donc dépendant du glutamate avec les récepteurs NMDA et AMPA ; de la diminution de la transmission synaptique inhibitrice, donc dépendant du GABA avec les récepteurs au GABA ; de l’altération des canaux voltages-dépendants donc des canaux calciques, sodiques, potassiques ou chlore (et donc de l’hyperexcitabilité intrinsèque) ; de la synchronisation des synapses excessive par la sommation temporelle et spatiale (c’est-à-dire est ce qu’on a beaucoup d’excitatrice qui vient connecter le neurone ou est qu’il y a des décharges très rapprochées excitatrices) ; des concentrations ioniques à l’intérieur et à l’extérieur (si on modifie les concentrations ioniques à l’intérieur et à l’extérieur des neurones, le passage à travers les canaux va se faire différemment ce qui peut avoir un impact sur la dépolarisation).

SNP – Epilepsies. Physiopathologie, bases thérapeutiques des épilepsies
16/20
C- Conséquences de l’épilepsie et mécanismes d’aggravation
Dans l’épilepsie tous les éléments vus juste avant peuvent être touchés. Les conséquences de cela sont que si on a une hyperexcitabilité pour une quelconque raison, on va avoir la base des anomalies épileptiques au niveau neuronal qui va être le shift de la dépolarisation paroxystique.
Normalement, un neurone doit faire un potentiel d’action puis s’arrêter. Dans l’épilepsie la base est que les neurones sont hyperexcitables donc au lieu de faire un potentiel d’action et de revenir à leur état basal, ils vont rester dépolarisés et faire plusieurs potentiels d’action d’affilée. Si par exemple on prend un neurone normal en noir qui est le contrôle et un neurone où on applique de la pénicilline qui est connue pour être un antibiotique mais active certains canaux et rend les neurones hyperexcitables (c’est d’ailleurs pour ça qu’on peut faire des crises d’épilepsie avec certains antibiotiques), cela excite le neurone qui va se mettre à avoir des dépolarisations. C’est la base de ce qu’on enregistre sur l’EEG : on a une pointe qui correspond à une paroxysmal depolarisation shift qui fait que le neurone s’est mis à décharger, ce qui est du au fait qu’on ait une hyperexcitabilité.
Sur l’EEG normal il n’y a pas de PDS (Paroxysmal Depolarisation Shit). Lorsqu’on en a un, on enregistre une pointe. Si en plus les différents neurones sont bien synchronisés on en enregistre plusieurs qui se mettent en route ensemble. Le PDS est la conséquence de l’hyperexcitabilité et donc l’expression des neurones épileptiques qui vont faire ce comportement là qui n’est pas habituel.
SNP – Epilepsies. Physiopathologie, bases thérapeutiques des épilepsies
17/20
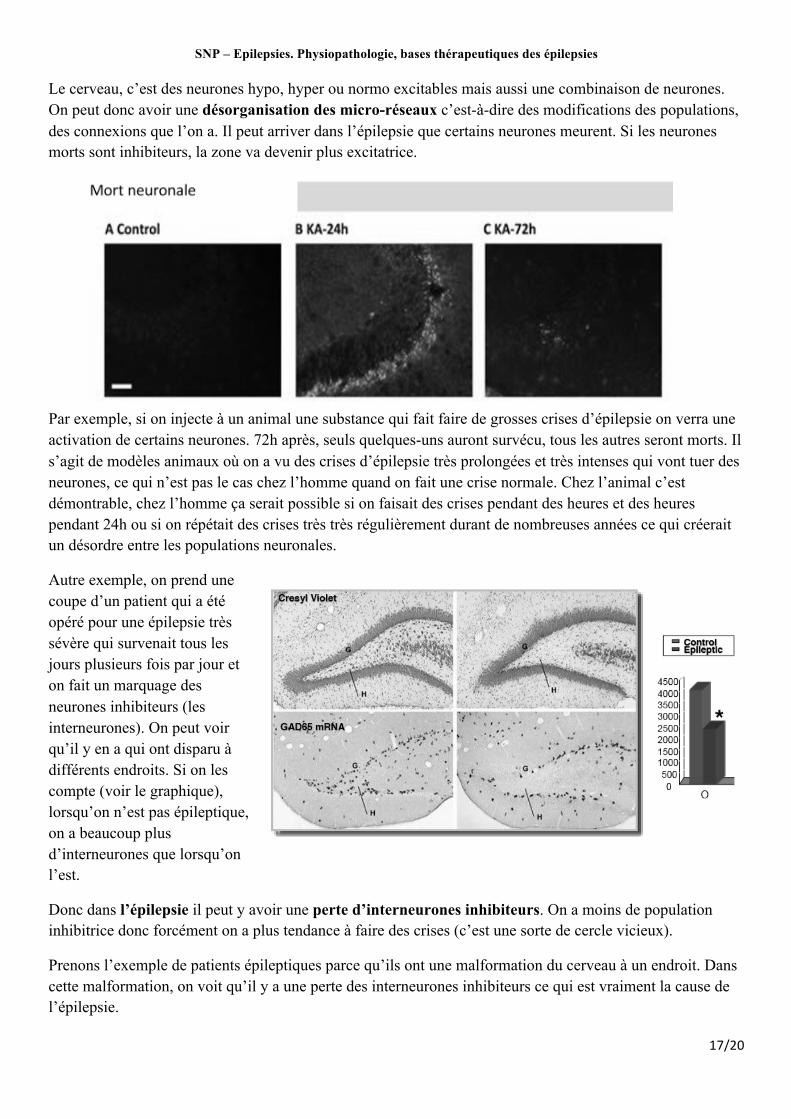
Le cerveau, c’est des neurones hypo, hyper ou normo excitables mais aussi une combinaison de neurones. On peut donc avoir une désorganisation des micro-réseaux c’est-à-dire des modifications des populations, des connexions que l’on a. Il peut arriver dans l’épilepsie que certains neurones meurent. Si les neurones morts sont inhibiteurs, la zone va devenir plus excitatrice.
Par exemple, si on injecte à un animal une substance qui fait faire de grosses crises d’épilepsie on verra une activation de certains neurones. 72h après, seuls quelques-uns auront survécu, tous les autres seront morts. Il s’agit de modèles animaux où on a vu des crises d’épilepsie très prolongées et très intenses qui vont tuer des neurones, ce qui n’est pas le cas chez l’homme quand on fait une crise normale. Chez l’animal c’est démontrable, chez l’homme ça serait possible si on faisait des crises pendant des heures et des heures pendant 24h ou si on répétait des crises très très régulièrement durant de nombreuses années ce qui créerait un désordre entre les populations neuronales.
Autre exemple, on prend une coupe d’un patient qui a été opéré pour une épilepsie très sévère qui survenait tous les jours plusieurs fois par jour et on fait un marquage des neurones inhibiteurs (les interneurones). On peut voir qu’il y en a qui ont disparu à différents endroits. Si on les compte (voir le graphique), lorsqu’on n’est pas épileptique, on a beaucoup plus d’interneurones que lorsqu’on l’est.
Donc dans l’épilepsie il peut y avoir une perte d’interneurones inhibiteurs. On a moins de population inhibitrice donc forcément on a plus tendance à faire des crises (c’est une sorte de cercle vicieux).
Prenons l’exemple de patients épileptiques parce qu’ils ont une malformation du cerveau à un endroit. Dans cette malformation, on voit qu’il y a une perte des interneurones inhibiteurs ce qui est vraiment la cause de l’épilepsie.

SNP – Epilepsies. Physiopathologie, bases thérapeutiques des épilepsies
18/20
Même en dehors du nombre de neurones, les fibres (c’est-à-dire les axones qui se connectent les uns aux autres) peuvent être modifiés. C’est ce qu’il va se passer dans certains signes d’épilepsie. On a des cellules granulaires qui sont des cellules excitatrices : en situation contrôle il y en a peu, elles font peu de synapses. Dans l’épilepsie, il y en a qui auront plus d’axones et il y en a qui seront à des endroits anormaux. Les cellules excitatrices vont donc être plus nombreuses et vont avoir plus de ramifications et donc plus de contact synaptique excitateur avec plus de neurones : c’est ce qu’on appelle le Sprouting, au niveau des hippocampes épileptiques. Il s’agit de connexions excitatrices récurrentes c’est-à-dire qu’il y a plusieurs connexions excitatrices sur un neurone (ce qui normalement ne sert à rien) et qu’elles sont anormales.
On peut donc avoir une désorganisation du nombre de neurones mais aussi une désorganisation des fibres neuronales avec plus de fibres excitatrices dans certaines zones.
Il y a également d’autres mécanismes qui sont en jeu :
- Des altérations de la barrière hémato-méningée ce qui laisse passer plus facilement certaines substances ou non - De l’inflammation, qui sont des phénomènes accessoires aggravant le phénomène d’hyperexcitabilité de base - De l’angiogenèse (sur la diapo mais pas dit à l’oral) - La vascularisation du cerveau est compliquée : elle concerne les neurones mais aussi les cellules gliales (cellules de soutien) qui ont un rôle important dans les synapses (et pas juste le rôle de maintenir la forme du cerveau) avec des canaux qui font des entrées et des sorties ce qui régule aussi la concentration de certaines molécules dans la synapse. Dans l’épilepsie les cellules gliales sont aussi altérées. Par exemple si on induit une crise à un moment donné, on pourra voir 48h après l’activation des cellules gliales parce qu’il va aussi y avoir une atteinte des cellules gliales dans l’épilepsie. Cette altération des cellules gliales va aussi créer un désordre des concentrations ioniques et des concentrations de neurotransmetteurs dans la synapse. Il s’agit d’un autre mode d’action. - Il y a également une altération des cellules immunitaires c’est-à-dire que dans un cerveau normal il n’y a pas de macrophages mais lorsqu’on une crise on va avoir une entrée de macrophages qui vont sécréter des substances inflammatoires qui vont rendre hyperexcitable le neurone.
Egalement, au niveau de la connexion entre les aires cérébrales (milliers voire millions de neurones connectés entre différentes aires), la connexion est également altérée. Dans l’épilepsie on a plusieurs types de réseaux différents : réseau de début des crises (réseau épileptogène), zone de propagation des crises et la zone non-impliquée.
On peut les juger par différentes méthodes. Cette connectivité est altérée c’est-à-dire que les zones épileptiques sont trop connectées les unes avec les autres quand on les mesure, elles sont déconnectées du reste du cerveau et il y a des réseaux non épileptiques qui servent pour le langage, la connexion etc qui sont eux aussi altérés, c’est pour cela qu’il y a des patients épileptiques qui sont aggravés.

SNP – Epilepsies. Physiopathologie, bases thérapeutiques des épilepsies
19/20
Si on résume, dans l’épilepsie on a :
- L’hyperexcitabilité neuronale - La désorganisation des micro-réseaux de neurones
- La désorganisation des macro-réseaux de neurones - L’altération de la barrière hémato encéphalique et l’inflammation
La physiopathologie dépend donc vraiment de plusieurs paramètres différents.
Question : Les zones épileptiques sont trop connectées entre elles, c’est un signe de renforcement particulier ? Oui, on pense que les décharges anormales au niveau micro renforcent les connexions excitatrices et au niveau macro aussi vont renforcer les connexions à longue distance. C’est probablement un phénomène de pérennisation des décharges épileptiques partielles/focales parce que ces zones-là sont tout le temps bombardées d’anomalies épileptiques donc elles vont renforcer ces connexions-là. Cela participe à l’hypersynchronisation que l’on voit dans les crises d’épilepsie et c’est un cercle vicieux.
Merci à Mélanie qui a sauvé ce qu’il me reste d’audition et mon week-end de Noël, bonnes fêtes à tous et bon courage pour ce cours certes un peu épais mais très redondant… :)
CR : si vous lisez la dédicace avant le cours, je sais que 20 pages en 1h c’est assez affolant, le prof parlait très vite, il a expliqué plein de choses et la ronéotypeuse a merveilleusement bien fait son travail en retranscrivant toutes les explications données en cours (Merci Claire ton ronéo est juste un bonheur !) c’est pour cela que c’est un cours très conséquent. Courage…

SNP – Epilepsies. Physiopathologie, bases thérapeutiques des épilepsies
20/20





























