Embed Size (px)
Citation preview

AMER. ZOOL., 22:795-805 (1982)
The Renewing Cell Populations of Ascidians1
THOMAS H. ERMAK
Department of Physiology, University of California,San Francisco, California 94143
SYNOPSIS. Renewing cell populations are tissues or groups of cells which rapidly prolif-erate and whose cell division is balanced by cell loss. The rapid proliferation of cells canbe determined in autoradiograms by the uptake of tritiated thymidine into DNA synthe-sizing cells, and the migration and loss of cells can be followed by taking samples of tissuesat increasing time intervals after exposure to the radioisotope. In ascidians, renewingpopulations are the testis, ovary, blood cells, and epithelial lining of the digestive tract.They are made of subpopulations called compartments: 1) germinal compartments ofrelatively undifferentiated, dividing stem cells (these cells are labeled at short time inter-vals), 2) mature compartments of fully differentiated, nondividing cells (these becomelabeled with time by the migration of stem cells), and 3) transitional compartments of cellsin intermediate stages. The stomach groove cell population is a model system for celldifferentiation and renewal in the digestive tract. Germinal compartments can be distin-guished from mature compartments by their morphology (pseudostratified vs. simplecolumnar epithelia), by the uptake of tritiated thymidine in autoradiograms, by histo-chemical staining, by their surface features in the scanning electron microscope, and bytheir intracellular organelles when viewed in the transmission electron microscope. Stemcells differentiate into absorptive cells and zymogen cells. In differentiating, stem cellsincrease in size and lose their abundant free ribosomes. Absorptive cells develop longermicrovilli, a long cilium, smooth apical vesicles, and large supranuclear lysosomes. Zym-ogen cells produce abundant rough endoplasmic reticulum and numerous zymogen gran-ules. The renewing cell populations of blood (lymph nodule) and digestive tract are im-portant model systems for studying cell differentiation, morphogenesis, and the phylogenyof vertebrate hemopoietic, immune, and digestive systems.
INTRODUCTION
From a developmental perspective, a re-newing cell population is an adult tissue orgroup of cells which maintains a subpop-ulation of embryonic-like cells which con-tinually replace aging differentiated cellsduring the lifetime of the animal. Theserenewing populations serve a variety offunctions in the body. For example, theymay form a protective barrier of cellsagainst the external environment, such asin the digestive tract or skin, or they mayprovide a constant supply of a cellularproduct, such as the blood cells or ga-metes.
The presence of these embryonic-likecells, today called stem cells (Cairnie et ai,1976), in rapidly dividing tissues was rec-ognized for years based upon histologicalobservations. However, it was not until theuse of autoradiography in the late 1940s
1 From the Symposium on The Developmental Biologyof the Ascidians presented at the Annual Meeting ofthe American Society of Zoologists, 27-30 December1981, at Dallas, Texas.
that the turnover of these renewing cellpopulations could truly be established andfollowed. The pioneering work was per-formed by Leblond and his associates witha variety of radioisotopes to label dividingcells and follow their fate with time (Le-blond and Stevens, 1948; Leblond andWalker, 1956). They found that tritiumgave the best resolution in autoradiogramsand that thymidine preferentially labeledreplicating DNA. Although thymidine isnot normally a precursor of DNA, it cannonetheless enter the biosynthetic pathwayand label DNA during both pre-mitotic andpre-meiotic DNA synthesis (for review, seeCleaver, 1967). When administered, thetritiated thymidine is available for only ashort period of time, and only those cellsthat are synthesizing DNA at the time ofinjection and immediately afterwards arelabeled. After DNA synthesis, all cells pre-sumably undergo cell division. The fate ofthe DNA synthesizing cells can be followedin autoradiograms by taking samples of thetissues at increasing time intervals after theinitial exposure period.
795
by Thom
as Erm
ak on April 24, 2010
http://icb.oxfordjournals.orgD
ownloaded from
796 THOMAS H. ERMAK
cellextrusion
celldivision
celldifferentiation
B
absorptionand secretion
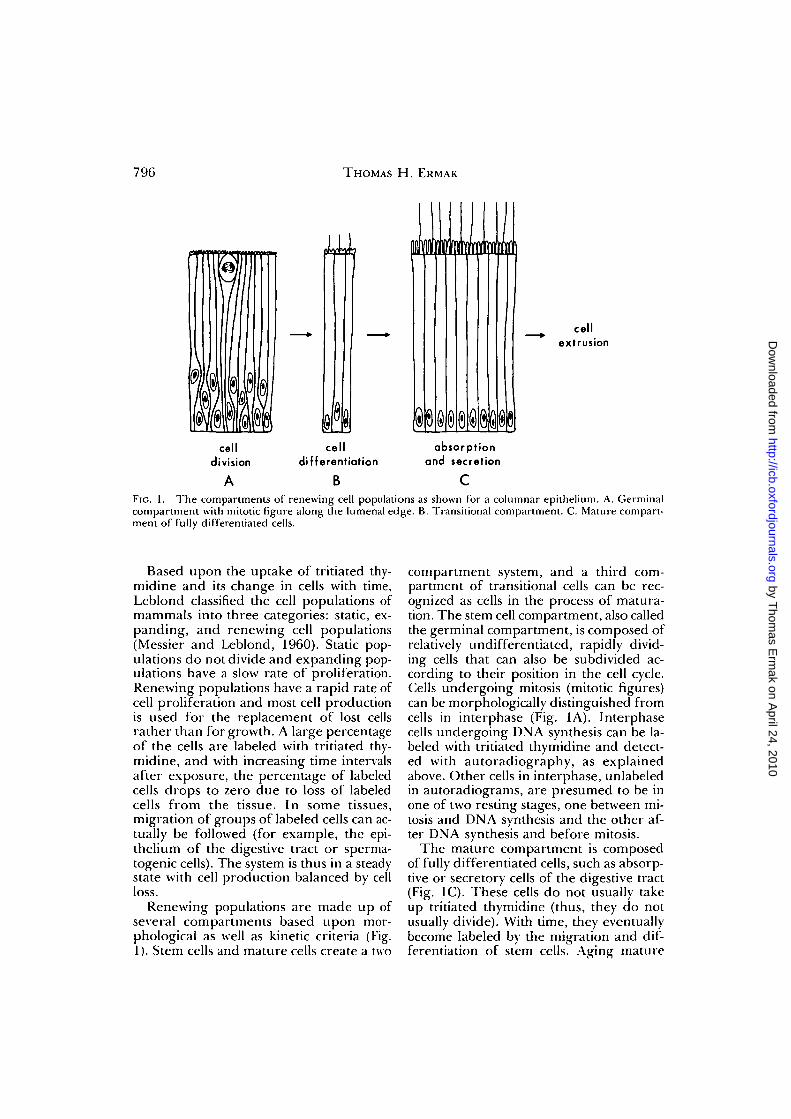
FIG. 1. The compartments of renewing cell populations as shown for a columnar epithelium. A. Germinalcompartment with mitotic figure along the lumenal edge. B. Transitional compartment. C. Mature compart-ment of fully differentiated cells.
Based upon the uptake of tritiated thy-midine and its change in cells with time,Leblond classified the cell populations ofmammals into three categories: static, ex-panding, and renewing cell populations(Messier and Leblond, 1960). Static pop-ulations do not divide and expanding pop-ulations have a slow rate of proliferation.Renewing populations have a rapid rate ofcell proliferation and most cell productionis used for the replacement of lost cellsrather than for growth. A large percentageof the cells are labeled with tritiated thy-midine, and with increasing time intervalsafter exposure, the percentage of labeledcells drops to zero due to loss of labeledcells from the tissue. In some tissues,migration of groups of labeled cells can ac-tually be followed (for example, the epi-thelium of the digestive tract or sperma-togenic cells). The system is thus in a steadystate with cell production balanced by cellloss.
Renewing populations are made up ofseveral compartments based upon mor-phological as well as kinetic criteria (Fig.1). Stem cells and mature cells create a two
compartment system, and a third com-partment of transitional cells can be rec-ognized as cells in the process of matura-tion. The stem cell compartment, also calledthe germinal compartment, is composed ofrelatively undifferentiated, rapidly divid-ing cells that can also be subdivided ac-cording to their position in the cell cycle.Cells undergoing mitosis (mitotic figures)can be morphologically distinguished fromcells in interphase (Fig. 1A). Interphasecells undergoing DNA synthesis can be la-beled with tritiated thymidine and detect-ed with autoradiography, as explainedabove. Other cells in interphase, unlabeledin autoradiograms, are presumed to be inone of two resting stages, one between mi-tosis and DNA synthesis and the other af-ter DNA synthesis and before mitosis.
The mature compartment is composedof fully differentiated cells, such as absorp-tive or secretory cells of the digestive tract(Fig. 1C). These cells do not usually takeup tritiated thymidine (thus, they do notusually divide). With time, they eventuallybecome labeled by the migration and dif-ferentiation of stem cells. Aging mature
by Thom
as Erm
ak on April 24, 2010
http://icb.oxfordjournals.orgD
ownloaded from

RENEWING CELL POPULATIONS OF ASCIDIANS 797
TABLE 1. Compartments of asddian renewing cell populations classified according to morphology and cell kinetics.
Cell population
Testis
Ovary
Blood cells
Digestive tract
Type of cell
SpermalogoniaSpermatocytesSpermatidsSpermatozoaOogoniaPrimary oocytesOocytes I and IIOocytes IIIFollicle cellsTest cellsHemoblastsLeukocytesBasophilic cellsAbsorptive, zymogen and ciliated cells
Type of compartment
germinaldividing transitionaltransitionalmaturegerminaldividing transitionaltransitionalmaturedividing transitionalmaturegerminalmaturegerminalmature
* Each compartment originates from the preceding compartment except in the ovary, where germinal cellsgive rise to both oocytes and follicle cells.
cells are lost from the population by cellextrusion, cell death, or cell excretion.
The time that elapses between a cell en-tering a compartment and that cell or itsprogeny leaving it is its transit time. I usethis term to denote the shortest time for alabeled stem cell to migrate through thetransitional and mature compartments.
Renewing cell populations have been ex-tensively investigated in mammals and in-clude the epidermis, the epithelial liningof the alimentary canal, the hemopoieticcells of bone marrow, the lymphatic cellsof thymus, spleen, and lymph nodes, thespermatogenic cells, and the uterine andvaginal epithelial cells (Messier and Le-blond, 1960). The list of renewing cellpopulations in ascidians is almost as im-pressive: the epithelial lining of the diges-tive tract (Ermak, 1975a, c,' 1976a, 1981),the hemopoietic cells of lymph nodules andcirculating blood (Ermak, 19756, 1977), thespermatogenic cells (Ermak, 19766), andthe ovarian cells (Ermak, 19766). As inmammals, these populations can be sub-divided into germinal, transitional, andmature compartments (Table 1). I have ex-amined the kinetics of cell renewal in a sin-gle, solitary species, Styela clava (Ermak,1975a, b, c, 1976a, b, 1977), and comparedthem to those in a variety of other ascidianspecies (Ermak, 1977, 1981). Renewal of
the gonads occurs in all animal phyla (butnot all adult species) and the morphologi-cal events characterizing cell differentia-tion are familiar to most biologists. I will,therefore, only briefly review the kineticsof renewal in ascidians. On the other hand,the renewal of blood cells and digestivetract are less common, occurring mainly inthe vertebrates and a few invertebrates.These populations provide importantmodels for the differentiation of somaticcells in the adult and are important sys-tems for studying the phylogeny of verte-brate hemopoietic, immune (Wright, 1977),and digestive systems (Ermak, 1981). Forthese populations, the kinetics of cell re-newal will be described as well as the over-all aspects of cell differentiation duringmaturation and migration of cells.
GONADS
In the gonads, germ cells in both pre-meiotic and pre-mitotic DN A synthesis canbe labeled with tritiated thymidine and de-tected with autoradiography (Ermak,19766). In the testis, each compartment isin a direct transit line with the next. Sper-matogonia (stem cells) form the basal layeraround the circumference of the follicle.Spermatocytes and spermatids form con-tinuous layers within the basal layer andspermatozoa fill the lumen. In Styela, the
by Thom
as Erm
ak on April 24, 2010
http://icb.oxfordjournals.orgD
ownloaded from

798 THOMAS H. ERMAK
transit time from DNA synthesis in the pri-mary spermatocyte to the appearance oflabeled spermatozoa is about 10 days (Er-mak, 19766). A one-hour exposure to tri-tiated thymidine labels many spermato-gonia and spermatocytes, and after 10 daysspermatids and spermatozoa become la-beled. With even longer times (20-30 days),labeled spermatozoa appear in the spermducts.
In the ovary, the germinal epitheliumgives rise to oocytes in sequential stages(usually noted as I, II, and III) as well asfollicle cells surrounding the oocytes(Tucker, 1942; Kessel and Kemp, 1962).Test cells are presumably derived fromfollicle cells. DNA synthesizing cells areoogonia and preleptotene primary oo-cytes. The smallest oocytes lie within theovarian wall, and larger oocytes becomeprogressively displaced from the germinalepithelium as they grow larger, become lessbasophilic, and acquire accessory cells. Al-though not followed for more than 60 days,the total time period for oogenesis is prob-ably on the order of several months (Er-mak, 19766). At one hour to 20 days afterexposure, cells in the germinal epitheliumas well as follicle cells around the oocytesare labeled. By 60 days, a few stage I oo-cytes and test cells are labeled. Test cellsare presumed to originate from follicle cellswhereas oocytes are presumed to originatefrom gonial cells in the ovarian wall asoriginally proposed by Tucker (1942).
BLOOD CELLS
The hemopoietic tissue of ascidians isorganized into clusters of stationary cellscalled lymph nodules and circulating cells(George, 1939; Millar, 1953). Lymph nod-ules occur in the pharyngeal wall, aroundthe digestive tract, and in the body wall ofadvanced species (Ermak, 1977). In prim-itive ascidians, hemopoietic tissue appearsdiffuse whereas in advanced species, it isorganized into distinct nodules. In Styela,a nodule is composed of a few clusters ofdividing hemoblasts (the stem cells) sur-rounded by non-dividing, maturing bloodcells (Ermak, 1977). Lymph nodules liewithin connective tissue below the pharyn-
geal or atrial epithelium and along bloodchannels.
In Styela, blood cells are renewed withinseveral weeks (Ermak, 19756). A one-hourexposure to tritiated thymidine labels he-moblasts in the centers of nodules and incirculating blood. By 20 days after expo-sure, cells in the peripheral parts of a nod-ule become labeled whereas cells in the in-terior are no longer labeled. By 60 days, anodule as well as most circulating bloodcells are no longer labeled.
Although some differentiated blood cellscan apparently divide, most labeled cellsare probably hemoblasts. These cells arealso sensitive to X rays (Freeman, 1964,1970), a characteristic shared by stem cellsof vertebrates (Patt and Quastler, 1963).Most fully differentiated blood cells prob-ably do not divide. Vacuolated cells in Stye-la are not labeled at early time intervals(Ermak, 19756). In Perophora annectens,compartment cells and phagocytes but notother fully differentiated cells are report-ed to be labeled with tritiated thymidine(Freeman, 1970).
The lymph nodule of Styela is a modelsystem for the differentiation of blood cells(Ermak, 1977). Hemoblasts are relativelyundifferentiated, spherical cells which havea large nucleus containing one or more nu-cleoli and little chromatin. The cytoplasmis filled with numerous polyribosomes, afew cisternae of rough endoplastic reticu-lum, and an occasional small, dense gran-ule. A pair of centrioles (probably involvedin centromere formation during mitosis) issometimes observed near the Golgi com-plex.
Differentiating leukocytes around thecentral hemoblasts lose their prominentnucleus as the amount of chromatin in-creases (Ermak, 1977). Large electrondense granules usually appear in the cy-toplasm before the nuclear changes arecompleted. As one proceeds away from thecenter of a nodule, maturing blood cellsincrease in size and their dense granulesbecome larger and more numerous. Celldifferentiation is also marked by the lossof polyribosomes and the development ofelongate mitochondria, a larger Golgi
by Thom
as Erm
ak on April 24, 2010
http://icb.oxfordjournals.orgD
ownloaded from

RENEWING CELL POPULATIONS OF ASCIDIANS 799
complex, and long cisternae of rough en-doplasmic reticulum.
DIGESTIVE TRACT
Most of the epithelia lining the digestivetract are renewing cell populations. In Styekithey are the dorsal tubercle, dorsal lamina,branchial bars, zone 1 of the endostyle,stigmata, esophagus, stomach, and intes-tine (Ermak, 1975c). In the postbranchialdigestive tract of most other ascidians, re-newing populations are also a characteris-tic feature of the esophageal and stomachepithelia, but expanding populationssometimes cover all or part of the intestine(Ermak, 1981). The renewing populationsare adapted to different degrees of bodysize, organization, and evolutionary ad-vancement, and with an increase in bodysize, the digestive organs undergo exten-sive folding and multiplication of pairs ofgerminal and mature compartments (cellrenewal units). Colonial ascidians gener-ally have a single cell renewal unit per cellpopulation in their small organs whereassolitary ascidians have highly folded gutepithelia with multiplication of cell renew-al units (Ermak, 1981). The groove pop-ulation in the stomach of Styela provides amodel system for cell renewal and differ-entiation in the digestive tract. This epi-thelium covers the majority of each stom-ach fold as shown in the scanning electronmicrograph2 of the stomach wall (Fig. 2).Germinal zones occur at the base of eachfold and form a pseudostratified columnarepithelium (Ermak, 1975c). A one-hourexposure to tritiated thymidine labels about10% of the groove height (Ermak, 1976a).Stem cell nuclei migrate to the cell apexand undergo mitosis there. Mitotic figuresare labeled within several hours after ex-posure (Ermak, 1975c). With time, stemcells migrate as a band of labeled cells over
the underlying connective tissue throughthe transition zone and up the sides of thefolds into the non-proliferating maturezone of absorptive and zymogen cells. Themature zone forms a simple, columnar ep-ithelium and represents about 80% of theheight of the fold. Migrating labeled cellsreach the top of the fold in about 16 days(Ermak, 1976a). Aging mature cells arepresumably extruded into the gut lumenat so-called "extrusion zones" at the top ofthe fold (at the junction with the crest pop-ulation of ciliated mucous cells). Migrationrates in other regions of the digestive tractrange from 10 days for the stigmata to 5weeks in the intestine (Ermak, 1975c).
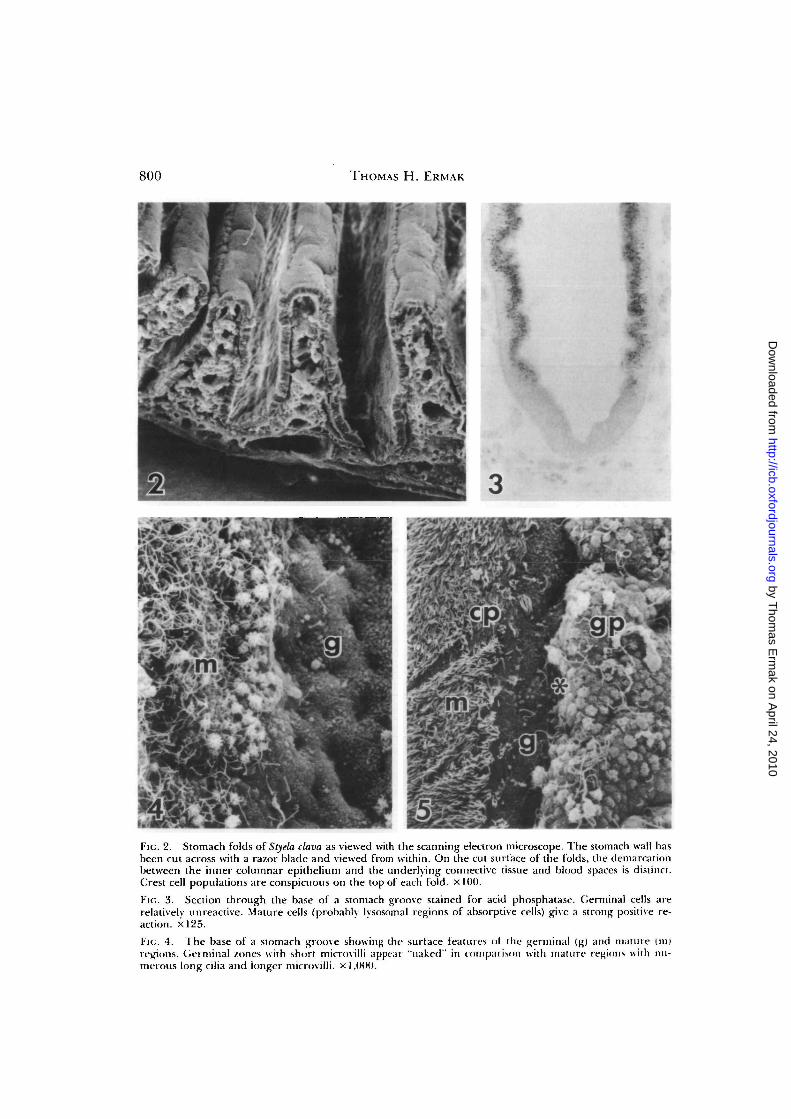
Germinal regions can be distinguishedfrom mature regions at the light micro-scopic level with the use of histochemicalstains. With routine hematoxylin and eo-sin, the stem cell is characterized by baso-philic cytoplasm, and with toluidine blueon epon sections, only the lysosomal re-gions of mature cells are stained. The ma-ture region also gives a strong positive re-action for acid phosphatase3 which is absentfrom the cytoplasm of stem cells (Fig. 3).The brush border at the base of the grooveusually shows only a weak alkaline phos-phatase reaction in comparison to the in-tense reaction on the sides of the folds (Er-mak, 1975a). Finally, the cytoplasm of thegerminal region takes up large amounts oftritiated leucine (Ermak, 1975a), a reflec-tion of active protein synthesis in these cells.
Germinal regions can also be distin-guished from mature regions in surfaceview with the scanning electron micro-scope (Fig. 4). Stem cells appear free ofsurface organelles marking differentia-tion, namely cilia or long microvilli. In-stead the germinal compartment is markedby smooth, short microvilli. At the top ofthe fold, the "extrusion zone" can be iden-
- For scanning electron microscopy, stomach tissuewas fixed in 2% glutaraldehycle in 74% sea water for24 hr (Holland and Jespersen, 1973; Nemanic andPitelka, 1971) and dried in a Freon critical point dryer.Specimens were coated with about 15 nm of gold-palladium alloy and examined with a Stereoscan S4scanning electron microscope.
3 To test for the presence of acid phosphatase, tis-sues were prepared according to the standard cou-pling azo dye technique (Pease, 1968) using frozensections incubated at room temperature with sodiuma-naphthyl phosphate, poly vinyl pyrrolidone, and fastgarnet CBC salt in 0.1 M acetate buffer at pH 5.0.
by Thom
as Erm
ak on April 24, 2010
http://icb.oxfordjournals.orgD
ownloaded from
800 THOMAS H. ERMAK
FIG. 2. Stomach folds of Styela clava as viewed with the scanning electron microscope. The stomach wall hasbeen cut across with a razor blade and viewed from within. On the cut surface of the folds, the demarcationbetween the inner columnar epithelium and the underlying connective tissue and blood spaces is distinct.Crest cell populations are conspicuous on the top of each fold, x 100.
FIG. 3. Section through the base of a stomach groove stained for acid phosphatase. Germinal cells arerelatively unreactive. Mature cells (probably lysosomal regions of absorptive cells) give a strong positive re-action. X125.
Fit;. 4. The base of a stomach groove showing the surface features ot the germinal (g) and mature (in)regions. Germinal /ones with short microvilli appear "naked" in comparison with mature regions with nu-merous long cilia and longer microvilli. x 1,000.
by Thom
as Erm
ak on April 24, 2010
http://icb.oxfordjournals.orgD
ownloaded from

RENEWING CELL POPULATIONS OF ASCIDIANS 801
tified as the junction between the groovepopulation and the ciliated crest cell pop-ulation (Fig. 5).
The differentiation of stem cells into ab-sorptive cells and zymogen cells (Fig. 6) hasbeen followed by transmission electronmicroscopy4 in the stomach groove popu-lation of S. clava (Ermak, 1975a; Thorn-dyke, 1977) and on the stomach folds ofCiona intestinalis (Thomas, 1970). Thesemature cells appear to be characteristic ofthe stomach and digestive diverticulum ofother ascidians as well (Degail and Levi,1964; Burighel and Milanesi, 1973).
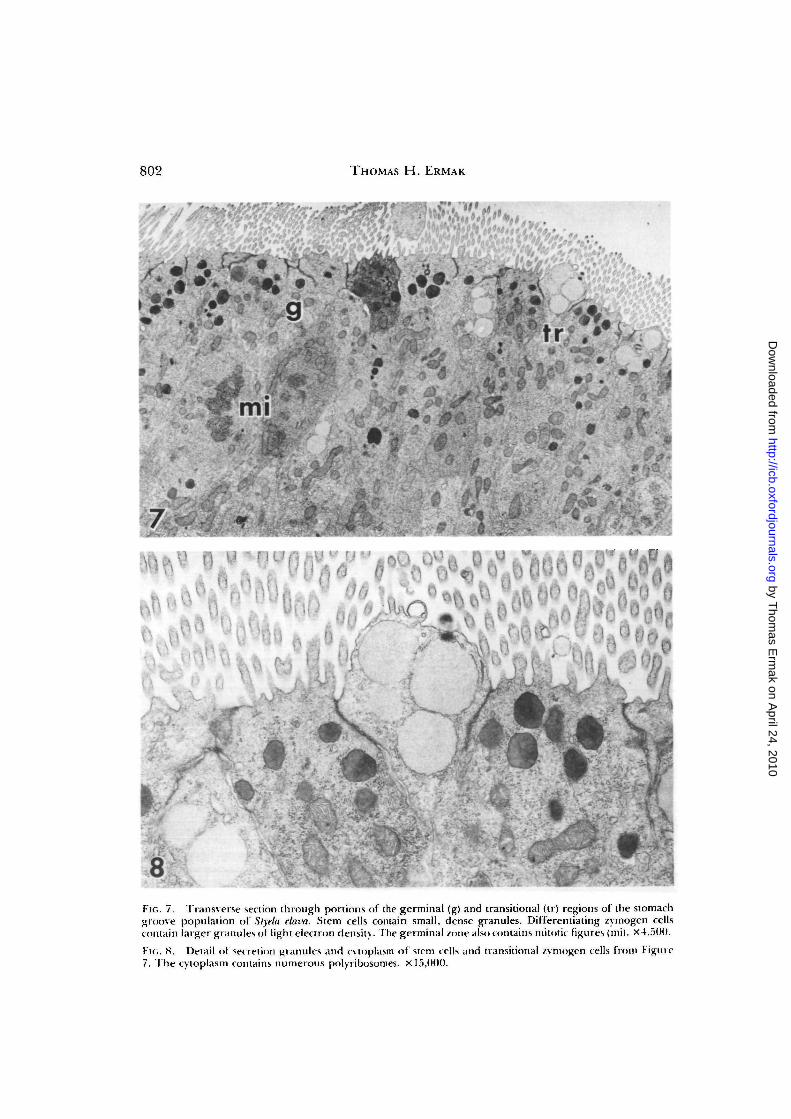
Stem cells (Fig. 6A) are smaller in size,have a higher nucleus-to-cytoplasm ratio,and are relatively undifferentiated instructure (Fig. 7). Their most chracteristicfeatures, like those of the hemoblast in alymph nodule, are a nucleus containing aprominent nucleolus and little chromatin(Thorndyke, 1977) and numerous free ri-bosomes in the cytoplasm (Fig. 8). The api-cal ends of the cells bear several microvilliwhich are shorter (about 3 /xm) and lessnumerous than those on the absorptive cellsand several small, dense granules. Belowthe cell apex, mitotic figures are occasion-ally observed (Fig. 7). There are no ciliaextending into the stomach lumen, but apair of diplosomal centrioles is located justbelow the microvilli. These organelles arepresumably involved in centromere for-mation during mitosis and probably alsogive rise to the cilia of absorptive cells.
During differentiation into absorptivecells (Fig. 6B) and zymogen cells (Fig. 6C),stem cells lose their dense secretory gran-
4 For transmission electron microscopy, stomachtissue was fixed in 3% glutaraldehyde in 0.1 M phos-phate buffer (pH 7.3) with 0.70 M sucrose for 1.5 hrfollowed by postfixation in ice cold 1% OsO4 in thesame sucrose-buffered medium for 1 hr. Samples wereembedded in Epon 812 and examined in a JEM-100Belectron microscope.
A B C
FIG. 6. Schematic diagrams of the stem cell (A), ab-sorptive cell (B), and zymogen cell (C) in the stomachgroove cell population of Styela clava. c, cilium; dc,diplosomal centrioles; go, Golgi apparatus; ly, lyso-some; mb, multivesicular body; rer, rough endoplas-mic reticulum; sg, secretory granule of stem cell; v,apical vesicles; z, zymogen granule.
ules and numerous free robisomes. Thenucleus of the absorptive cell loses itsprominent nucleolus, but it is retained inthe zymogen cell.
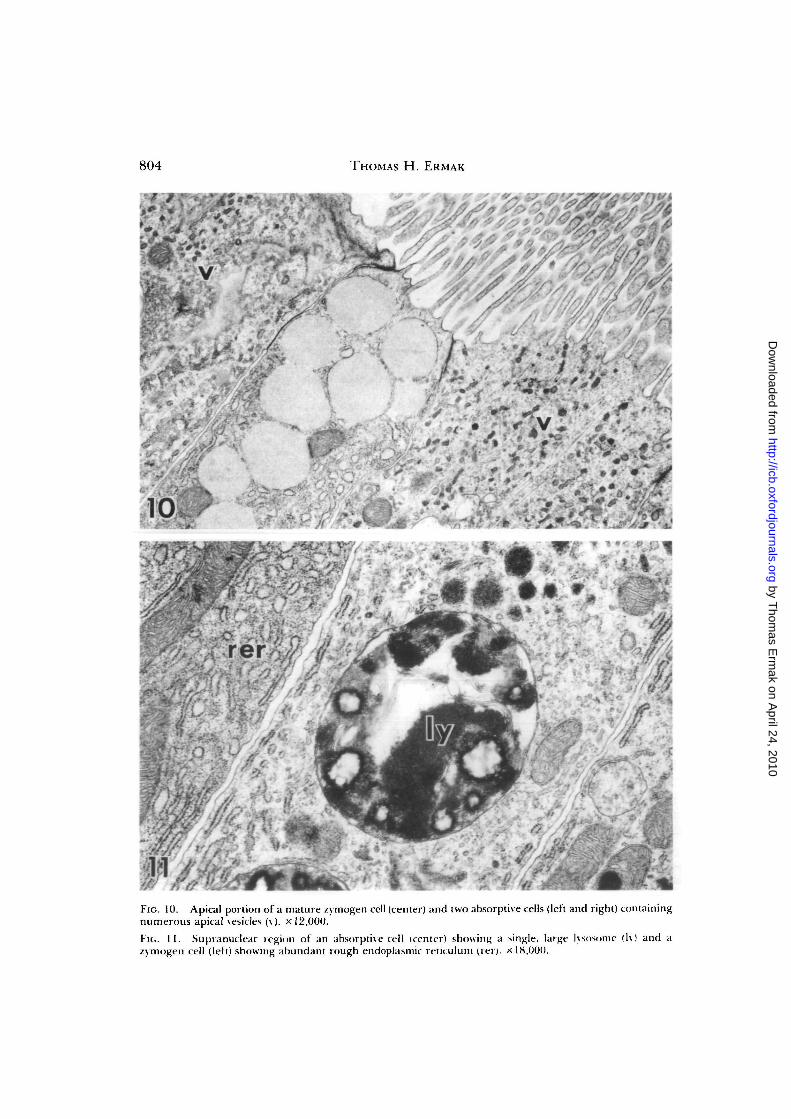
Absorptive cells (Fig. 6B) predominatein the mature region and are characterizedby long microvilli (about 8 /xm long), a longcilium, apical smooth vesicles, elements ofrough endoplasmic reticulum, and a stackof large supranuclear lysosomes (Figs. 9—
FIG. 5. Junction (asterisk) of a groove population (gp) with a crest population (cp) at the top of a stomachfold. Cell extrusion presumably occurs at this point. Crest population also shows differentiation between"naked" germinal (g) regions and ciliated mature (m) regions, x 1,000.
by Thom
as Erm
ak on April 24, 2010
http://icb.oxfordjournals.orgD
ownloaded from
802 THOMAS H. ERMAK
iV/'<
•? /
8• a *
FIG. 7. Transverse section through portions of the germinal (g) and transitional (tr) regions of the stomachgroove population of Styela clava. Stem cells contain small, dense granules. Differentiating zvmogen cellscontain larger granules ot light electron density. The germinal zone also contains mitotic figures (mi). x4,5(>0.
FK.. 8. Detail of secretion granules and cvtoplasm of stem cells and transitional zvmogen cells from Figure7, The cytoplasm contains numerous polyribosomes. x 15,000.
by Thom
as Erm
ak on April 24, 2010
http://icb.oxfordjournals.orgD
ownloaded from

RENEWING CELL POPULATIONS OF ASCIDIANS 803
''•' '•'•xXiyi. . * *'• .^m. ^ B L J ^ . '^ .-.••'/?".- ' ^~ ' x">-:
FIG. 9. Apical portion of transitional absorptive cells showing intermediate stage of apical vesicle (v) accu-mulation and short, blunt cilia (c). The latter appear to be true intermediates rather than tangentially sectionedlong cilia, x 9,000.
11). Transitional cells have smaller lyso-somes, an intermediate number of apicalvesicles and profiles of short, blunt ciliasuggestive of ciliary growth (Fig. 9; see alsoThorndyke, 1977). The lysosomes mea-sure about 2-4.5 /u.m in diameter and arefilled with membrane lamellae and clumpsof light and dark granular material (Fig.11). Although digestion in ascidians hasbeen described as extracellular (van Weel,1940; Morton, 1960), this organelle mightbe an important site of intracellular diges-tion (Ermak, 1975a; Burighel, 1979).
Differentiating zymogen cells are en-countered just outside of the zone of stemcells prior to the development of lysosomesin the absorptive cells (Figs. 7, 8). Maturezymogen cells (Fig. 6C) contain a large, ve-sicular Golgi complex, numerous zymogengranules (1—2 yjm in diameter), both in thecell apex (Fig. 10) and the vicinity of theGolgi complex, and abundant tubular ele-
ments of rough endoplasmic reticulum(Fig. 11).
CONCLUSIONS
The renewing cell populations of ascid-ians are characterized by a rapid rate ofproliferation, with cell production bal-anced by cell loss; thus, the system is in asteady state. These populations are un-common among invertebrates, particularlyrenewing blood cells and digestive epithe-lia. Considering the affinities of ascidiansto vertebrates (Berrill, 1955), it is possiblethat renewing cell populations are char-acteristic of chordates as a whole. Thesepopulations provide excellent model sys-tems for the study of cell differentiationand have been useful in examining theorigins of cell renewal (Ermak, 1981) andthe phylogeny of the vertebrate immunesystem (Ermak, 1977; Wright, 1977). Therole of these populations in morphogene-
by Thom
as Erm
ak on April 24, 2010
http://icb.oxfordjournals.orgD
ownloaded from
804 THOMAS H. ERMAK
.L.f
i ;
.:- /
\ *
,'i m
* » •
"•• * * •*-%'„'
•l~ft l i
.<» v̂*'
FIG. 10. Apical portion of a mature zymogen cell (center) and two absorptive cells (left and right) containingnumerous apical vesicles (v). x 12,000.
Fit.. I I . Supranuclear region of an absorptive cell (center) showing a single, large lysnsome (Iy) and azymogen cell (left) showing abundant rough endoplasmic retitulum (iei). x 18,000.
by Thom
as Erm
ak on April 24, 2010
http://icb.oxfordjournals.orgD
ownloaded from

RENEWING CELL POPULATIONS OF ASCIDIANS 805
sis, regeneration, and regression (of colo-nial ascidians) is virtually unknown and de-serving of further exploration.
ACKNOWLEDGMENTS
Part of this work (scanning electron mi-croscopy, enzyme histochuinistry, and fix-ation of tissues for transmission electronmicroscopy) was conducted at the ScrippsInstitution of Oceanography, La Jolla, Cal-ifornia 92037.
REFERENCES
Berrill, N. J. 1955. The origin of vertebrates. OxfordUniversity Press, London.
Burighel, P. 1979. Peroxidase absorption in the as-cidian gut. J. Exp. Zool. 207:131-142.
Burighel, P. and C. Milanesi. 1973. Fine structureof the gastric epithelium of the ascidian Botryllusschlosseri. Vacuolated and zymogenic cells. Z. Zell-Forsch. 145:541-555.
Cairnie, A. B., P. K. Lala, and D. G. Osmond, (eds.)1976. Stem cells of renewing cell populations. Aca-demic Press, New York.
Cleaver, J. E. 1967. Thymidine metabolism and cell ki-netics. John Wiley and Sons, Inc., New York.
Degail, L. and C. Levi. 1964. Etude au microscopeelectronique de la glande digestive des Pyuridae(Ascidies). Cah. Biol. Mar. 5:41 1-422.
Ermak, T. H. 1975a. Cell proliferation in the asci-dian Styela clava: An autoradiographic and elec-tron microscopic investigation emphasizing cellrenewal in the digestive tract of this and fourteenother species of ascidians. Ph.D. Diss., Univ. Cal-if., San Diego.
Ermak, T. H. 19756. An autoradiographic dem-onstration of blood cell renewal in Styela clava(Urochordata: Ascidiacea). Experientia 31:837-838.
Ermak, T. H. 1975c. Cell proliferation in the diges-tive tract oF Styela clava (Urochordata: Ascidi-acea) as revealed by autoradiography with triti-ated thymidine. J. Exp. Zool. 194:449^166.
Ermak, T. H. 1976a. Cell migration kinetics in thestomach of Styela clava (Urochordata: Ascidi-acea). J. Exp. Zool. 197:339-346.
Ermak, I. H. 19766. Renewal of the gonads in Stye-la clava (Urochordata:Ascidiacea) as revealed byautoradiography with tritiated thymidine. Tiss.Cell 8:471-478.
Ermak, I. H. 1977. The hematogenic tissues of tu-nicates. In R. K. Wright and E. L. Cooper (eds.),The phytogeny of thymus and bone marrow-bursa cells,pp. 45-56. Elsevier/North Holland, Amsterdam.
Ermak, T. H. 1981. A comparison of cell prolifer-
ation patterns of the digestive tract of ascidians.J. Exp. Zool. 217:325-339.
Freeman, G. 1964. The role of blood cells in theprocess oF asexual reproduction in the tunicatePerophora viridis. J. Exp. Zool. 156:157—184.
Freeman, G. 1970. The reticuloendothelial systemof tunicates. J. Reticuloendothelial Soc. 7:183-194.
George, W. C. 1939. A comparative study of theblood of the tunicates. Quart. J. Microscop. Sci.81:391^128.
Holland, N. D. and A. Jespersen. 1973. The finestructure of the fertilization membrane of thefeather star Comanthus japonica (Echinodermata:Crinoidea). Tiss. Cell 5:209-214.
Kessel, R. G. and N. E. Kemp. 1962. An electronmicroscope study on the oocyte, test-cell, and fol-licular envelope of the tunicate, Molgula munhat-tensis. J. Ultrastruct. Res. 6:57-76.
Leblond, C. P. and C. E. Stevens. 1948. The con-stant renewal of the intestinal epithelium in thealbino rat. Anal. Rec. 100:357-378.
Leblond, C. P. and B. E. Walker. 1956. Renewal ofcell populations. Physiol. Rev. 36:255-276.
Messier, B. and C. P. Leblond. 1960. Cell prolif-eration and migration as revealed by radioautog-raphy after injection of thymidine-H3 into malerats and mice. Am. J. Anat. 106:247-265.
Millar, R. H. 1953. Ciona. L.M.B.C. Memoirs XXXV.University Press, Liverpool.
Morton, J. E. 1960. The functions of the gut inciliary feeders. Biol. Rev. 35:92-140.
Nemanic, M. K. and D. R. Pitelka. 1971. A scanningelectron microscope study ot the lactating mam-mary gland. J. Cell Biol. 48:410-415.
Patt, H. M.and H. Quastler. 1963. Radiation effectson cell renewal and related systems. Physiol. Rev.43:357-396.
Pease, A. G. E. 1968. Histochemistry, theoretical andapplied. Little, Brown, and Co., Boston.
Thomas, N. W. 1970. Morphology of cell types fromthe gastric epithelium of Ciona intestinalis. J. Mar.Biol. Ass. U.K. 50:737-746.
Thorndyke, M. C. 1977. Observations on the gas-tric epithelium of ascidians with special referenceto Styela clava. Cell Tiss. Res. 184:539-550.
Tucker, G. H. 1942. The histology of the gonadsand development of the egg envelopes of an as-cidian (Styelaplicata Lesueur). J. Morphol. 70:81-113.
Weel, P. B. van. 1940. Beitrage zur Ernahrungs-biologie der Ascidien. Pub. Sta. Zool. Napoli 18:50-79.
Wright, R. K. 1977. Phylogenetic origin of the ver-tebrate lymphocyte and lymphoid tissue. In R. K.Wright and E. L. Cooper (eds.), The phylogeny ofthymus and bone marrow-bursa cells, pp. 57-70. El-sevier/North Holland, Amsterdam.
by Thom
as Erm
ak on April 24, 2010
http://icb.oxfordjournals.orgD
ownloaded from

by Thom
as Erm
ak on April 24, 2010
http://icb.oxfordjournals.orgD
ownloaded from





























