Embed Size (px)
Citation preview

JOURNALOF
www.elsevier.com/locate/jpsychires
Journal of Psychiatric Research 40 (2006) 37–46
PSYCHIATRIC
RESEARCH
Error processing in major depressive disorder: Evidence fromevent-related potentials
Martin Ruchsow *, Barbel Herrnberger, Petra Beschoner, Georg Gron,Manfred Spitzer, Markus Kiefer
Department of Psychiatry at the University of Ulm, Leimgrubenweg, D-89075 Ulm, Germany
Received 16 June 2004; revised 1 February 2005; accepted 25 February 2005
Abstract
In a previous study we showed that errors following errors activate a strategic (prefrontal) mechanism. In an error trial (trial n)following an erroneous previous trial (trial n � 1) healthy control subjects were found to have enlarged (more negative) amplitudesof the error related negativity (ERN)/error negativity (Ne), an electrophysiological correlate of response monitoring, in response to anegative feedback signal. Contrary to that, patients with major depressive disorder showed smaller (less negative) ERN/Ne ampli-tudes. It has been discussed controversially whether errors of choice (e.g., pressing an incorrect response button in an Eriksen flankertask) and errors of commission (e.g., pressing a button when one is not supposed to in a Go/Nogo task) are related to differentERN/Ne mechanisms. In the present study, we examined whether our previous result only holds for errors of choice in an Eriksenflankers task or extends to errors of commission in a Go/Nogo task, as well. Ten patients with DSM-IV major depressive disorderand 10 matched controls participated in a Go/Nogo task with performance feedback which signaled monetary reward. Patients withmajor depressive disorder again showed a less negative ERN/Ne amplitude in error trials following error trials. This result mightreflect impaired response monitoring processes in major depressive disorder resulting from an impaired activation of a centralreward pathway and/or a deficit in strategic reasoning.� 2005 Elsevier Ltd. All rights reserved.
Keywords: Error related negativity/error negativity; Performance feedback; Major depressive disorder
1. Introduction
The anterior cingulate cortex (ACC) and the prefron-tal cortex (PFC) have been the subject of increasingattention in neuroimaging research on major depressivedisorder (MDD). Two subdivisions of the ACC can bediscerned. The first, referred to as the ventral (affective)subdivision, encompasses the rostral and ventral areasof the ACC (Brodmann�s areas 25, 32, 33 and rostralarea 24), whereas the second, referred to as the dorsal
0022-3956/$ - see front matter � 2005 Elsevier Ltd. All rights reserved.
doi:10.1016/j.jpsychires.2005.02.002
* Corresponding author. Tel.: +49 731 50021468; fax: +49 73150021549.
E-mail address: [email protected] (M. Ruch-sow).
(cognitive) subdivision, involves dorsal regions of theACC (caudal area 24 0 and 32 0 and cingulate motor area)(Davidson et al., 2002). From a functional perspective,activation of the cognitive subdivision of the ACC hasbeen reported during monitoring of cognitive (Carteret al., 2000) and reward-related (Rogers et al., 1999)conflicts among other experimental manipulations. Theaffective subdivision is thought to be involved in regulat-ing visceral and autonomic responses to stressful behav-ioral and emotional events (Davidson et al., 2002).
In patients with MDD several PET and fMRI studieshave shown a hypoactivity in the dorsal subdivision ofthe ACC (Beauregard et al., 1998; Bench et al., 1992;Drevets et al., 1997). In addition, Mayberg et al.(1997) demonstrated that increased pretreatment

38 M. Ruchsow et al. / Journal of Psychiatric Research 40 (2006) 37–46
activity in the ventral (rostral) ACC is associated withbetter treatment response. Recently, Pizzagalli et al.(2001) replicated this finding with EEG source localiza-tion techniques and showed, that the magnitude of clin-ical improvement was predicted by baseline levels ofactivation in the same region of the ACC as identifiedby Mayberg et al. (1997).
Moreover, several studies have documented de-creased activation in both dorsolateral and dorsomedialPFC in patients with MDD (Davidson et al., 2002). Thereduction of activation in the PFC region seems to be atleast partially due to a reduction in the volume of graymatter as revealed by magnetic resonance imaging-de-rived morphometric measures (Drevets et al., 1997).
Patients with MDD often are biased towards evaluat-ing their performance in a more negative way resultingin a situation of learned helplessness and leading to det-rimental effects on performance (Elliott et al., 1996).Abnormal response to negative feedback has beenshown to be specifically related to clinical severity ofdepression (Elliott et al., 1997). Elliott and coworkershypothesized an impaired activation of a central rewardpathway as core feature of MDD involving limbic andprefrontal brain structures (Elliott et al., 1998; Schultzet al., 1992).
Event-related potentials (ERPs) are an effective tech-nique to track the time course of cognitive processeswith a time resolution in the range of milliseconds. Dur-ing the last years the error-related negativity (ERN;Gehring et al., 1990) or error negativity (Ne; Falkensteinet al., 1990), which refers to a negative-going ERP com-ponent following an incorrect response in choice reac-tion time tasks, has been intensively investigated andhas been interpreted as index of response monitoringprocesses. The ERN/Ne is a negative component withmaximum amplitude over fronto-central recording sitespeaking between 100 and 150 ms after the onset of anerroneous response (response-related electromyographic(EMG) activity) in forced choice reaction time para-digms like the Eriksen flanker task (Gehring et al.,1993) or Go/Nogo tasks (Falkenstein et al., 1999).Using source localization techniques, several studies(e.g., Dehaene et al., 1994; Miltner et al., 1997; Ruchsowet al., 2002) have localized the source of the ERN/Ne inthe area of the ACC, which belongs to the brain 0s limbicsystem. Several neuroimaging studies confirmed the acti-vation of the ACC in association with errors (Carteret al., 1998; Kiehl et al., 2000). In addition to ACC,an involvement of left prefrontal regions has beendescribed in several studies (Carter et al., 1998; Ruchsowet al., 2002) suggesting an activation of strategic reason-ing, generation of new rules (Wolford et al., 2000) orexertion of cognitive control (Carter et al., 1998).
Miltner et al. (1997) and Ruchsow et al. (2002) founda negative component that peaked around 200–300 msafter the onset of a negative feedback stimulus, the so-
called feedback-related ERN/Ne (henceforth fERN/Ne). Its phenomenology corresponded closely to thatof the response-related ERN/Ne (henceforth rERN/Ne), and source localization analyses suggested thatthe scalp distribution of the negative potential was con-sistent with a source in or very near the ACC – the samelocus as that reported for the rERN/Ne. Miltner et al.(1997) and Ruchsow et al. (2002) concluded thatrERN/Ne and fERN/Ne are manifestations of the sameresponse evaluation system.
There are several approaches to the functional signif-icance of the ERN/Ne components. The mismatch the-ory of error processing holds that rERN/Ne andfERN/Ne reflect a process that compares a representa-tion of a correct response with a representation of theactual response (e.g., Coles et al., 2001). An alternativeview states that the ACC monitors response conflictand is closely connected to a dopamine modulated re-ward system (Braver and Cohen, 2000). More recently,Holroyd and Coles (2002) proposed a theory combiningelements of mismatch theory and reinforcement learningprinciples. According to their suggestion the ERN/Ne
components are generated when a negative reinforce-ment learning signal is conveyed to the ACC via themesencephalic dopamine system. This signal is used bythe ACC to modify performance on the task at hand.
In a previous study (Ruchsow et al., 2004) we foundreduced fERN/Ne amplitudes to negative feedback in er-ror trials (trial n) following erroneous previous trials(trial n � 1) in patients with MDD. We used an Eriksenflanker task where subjects received immediate feedbackafter their response (button press). In contrast to theEriksen task which only induces errors of choice (i.e.,pressing an incorrect response button) the present Go/Nogo paradigm only elicits errors of commission (i.e.,responding when one is not supposed to). There is somecontroversy whether the ERN/Ne amplitude differs withregard to error type (errors of choice vs. errors of com-mission). A study by Gehring et al. (1993) suggested dif-ferent ERN/Ne mechanisms for the two error typeswhereas Scheffers et al. (1996) observed comparableERN/Ne amplitudes for errors of choice and errors ofcommission.
Using a Go/Nogo task, we examined in the presentstudy whether our previous result of smaller fERN/Ne
amplitudes in patients with MDD also holds for errorsof commission. First of all, we examined the rERN/Ne
and the fERN/Ne component irrespective of the previ-ous trial. In a second step, we investigated the effect ofthe correctness of the response in the previous trial (trialn � 1) on the ERPs elicited by the current trial (trial n)by calculating ERP waveforms of four different condi-tions: Error trials after error trials (EE), correct trialsafter error trials (EC), error trials after correct trials(CE), and correct trials after correct trials (CC). As pa-tients with MDD often show a hypoactivity in the dorsal

M. Ruchsow et al. / Journal of Psychiatric Research 40 (2006) 37–46 39
subdivision of the ACC (Beauregard et al., 1998; Benchet al., 1992; Drevets et al., 1997) which is crucial for cog-nitive (Carter et al., 2000) and reward-related (Rogerset al., 1999) conflicts we reasoned that patients withMDD should have smaller amplitudes in condition EEpossibly reflecting underactivity in a central rewardpathway (Elliott et al., 1998).
2. Methods
2.1. Subjects
Ten patients (nine female) with MDD and 10 controls(nine female) participated in the study. The patientgroup consisted of eight subjects with recurrent majordepressive disorder (296.3·) and two subjects with ma-jor depressive disorder, single episode (296.2·) accord-ing to DSM-IV (APA, 1994). Severity of depressionwas assessed with the Hamilton Depression Scale (Ham-ilton, 1960). Sum scores of the Hamilton Anxiety Scale(Hamilton, 1976), the Yale-Brown Obsessive Compul-sive Scale (Goodman et al., 1989), and the Brief Psychi-atric Rating Scale (Overall and Gorham, 1962) wereobtained in order to exclude patients with anxiety disor-der, obsessive compulsive disorder, and schizoaffectivedisorder from the study. Patients with concurrent diag-nosis of psychoactive substance misuse or neurologicalor general medical disorders (e.g., dementia, stroke, Par-kinson�s disease, head injury) were excluded from thestudy.
Groups were matched for gender, age and education.All subjects were right-handed (Oldfield, 1971) Germannative speakers and gave informed consent before par-ticipation. The experiment has been approved by the lo-cal Ethical Committee. All patients were examinedduring remission. Mean duration of major depressiveepisode was 31.4 days (SD = 18.5). Medication was sta-ble for at least 10 days (see Table 1).
2.2. Stimuli and procedures
A modified Go/Nogo task was used during whicheight different letter strings (congruent: BBBBB,DDDDD, VVVVV and UUUUU; incongruent:BBDBB, DDBDD, UUVUU and VVUVV) were pre-sented in the center of a computer screen in randomizedorder. Subjects had to focus on the target letter in themiddle of the letter string and had to press a responsekey upon appearance of letters B and U (Go condition)and to withhold the key press upon appearance of D andV (Nogo condition). From previous studies it is knownthat errors are more frequent for incongruent letterstrings than for congruent letter strings (Pailing et al.,2002). In order to enhance error rates in the Nogo con-dition (erroneous key presses) without reducing the
amount of correct Go trials (correct key press) BBDBBand UUVUU (incongruent Nogo condition) andBBBBB and UUUUU (congruent Go condition) werepresented with a probability of 0.2, whereas all other let-ter strings were presented with a probability of 0.05. Theentire experiment consisted of 600 trials. Thus, letterstrings BBDBB, UUVUU, BBBBB and UUUUU werepresented 120 times and letter strings DDBDD,VVUVV, DDDDD and VVVVV were presented 30times with a presentation time of 400 ms, each. Subjectsreceived feedback according to their performance750 ms after key press. As feedback stimuli we usedthe German expressions for ‘‘correct’’, ‘‘false’’ and ‘‘fas-ter’’. Feedback stimuli were presented for 500 ms. Theinter-trial-interval was 2600 ms. Participants got amonetary reward, winning or loosing a small amountof money each trial (5 Euro cents). Participants had torespond as fast and as accurately as possible. Beforerecording the EEG subjects had a practice period of120–240 trials. The time window for correct responseswas set individually after the training period rangingbetween 250 and 400 ms in order to obtain at least 5%of erroneous trials (i.e., 30 erroneous trials) and 10%of correct trials (i.e., 60 correct trials). The total set oftrials was divided into 5 blocks with 120 trials each(300 Go trials; 300 Nogo trials). Participants were seatedin a comfortable chair in a sound-attenuating, electri-cally shielded booth. The experiment lasted about2.5 h, including pauses, electrode placement, andremoval.
2.3. EEG recording and analysis
EEG was recorded using 64 channels mounted in anelastic cap (Easy-cap� system). Electrodes werepositioned with equal distances. All electrodes werereferenced to an electrode between Cz and FCz andre-referenced to average reference off-line. Eye move-ments were registered by vertical and horizontal EOG.Electrode impedances were kept below 5 kX. The EEGwas amplified by Neuroscan amplifiers (bandwidthDC-50 Hz; 50 Hz notch filter) and A/D converted with12-bit resolution at a rate of 250 Hz and digitally low-pass filtered with 16 Hz and digitally high-pass filteredwith 0.10 Hz.
For the analysis of the rERN/Ne component EEGwas baseline corrected to an interval between �150and 0 ms before the onset of the subject�s response. Arti-fact-free EEG-segments of a length of 650 ms were usedto compute response locked ERPs.
For the analysis of the fERN/Ne EEG was baselinecorrected to an interval between �150 and 0 ms beforethe onset of feedback stimulus. We used artifact-freeEEG-segments of a length of 650 ms to compute feed-back locked ERPs. Integrated power scores were usedfor data analysis.

Table 1Sociodemographic parameters, clinical evaluation and medication in patients with major depressive disorder and healthy participants
Major depressive disorder Healthy controls
N 10 10Gender (f/m) 9/1 9/1Age 37.5 ± 11.6 (20–60) 33.3 ± 12.8 (24–61)Years of education 9.6 ± 0.5 (9–10) 11.0 ± 2.2 (9–13)Tricyclic antidepressants (N = 10) 281.6 ± 158.2Chlorpromazine (N = 3) 28.1 ± 49.3Benzodiazepines (N = 2) 1.0 ± 2.1HAMD-21 12.8 ± 6.5 (10–28)HAMA 11.9 ± 6.2 (0–15)Y-BOCS 2.4 ± 1.6 (1–7)BPRS 35.1 ± 8.5 (23–49)
Values are means ± standard deviation (SD), ranges in brackets.Tricyclic antidepressants: reference substance was imipramine; N denotes number of patients with respective medication.Benzodiazepines: reference substance was diazepam.HAMD-21: Hamilton Depression Scale, 21 items (Hamilton, 1960).HAMA: Hamilton Anxiety Scale (Hamilton, 1976).Y-BOCS: Yale-Brown Obsessive Compulsive Scale (Goodman et al., 1989).BPRS: Brief Psychiatric Rating Scale (Overall and Gorham, 1962).
40 M. Ruchsow et al. / Journal of Psychiatric Research 40 (2006) 37–46
In the patient group 15.5% of the artifact-free seg-ments belonged to condition EE, 22.2% to EC, 25.9%to CE, and 36.3% to CC, whereas in the control group11.8% belonged to condition EE, 24.9% to EC, 27.7%to CE, and 35.6% to CC.
For statistical analysis, electrodes were selected fromtwo different scalp regions: a frontal electrode groupincluded midline electrodes FPz, AFz and Fz and thelateral pairs of electrodes AF3/AF4 and F1/F2. A cen-tro-parietal electrode group included midline electrodesFCz, Cz and CPz and the lateral pairs of electrodesC1/C2 and CP1/CP2. Voltages at midline and lateralelectrodes were submitted to repeated measures analysesof variance (ANOVAs). When applicable degrees offreedom were adjusted according to the method ofGreenhouse–Geisser, and the adjusted p-values as wellas the Greenhouse–Geisser � are reported.
The ANOVA for lateral electrodes included the fac-tors group, hemisphere, electrode position and condi-tion (EE/EC/CE/CC), the ANOVA for midlineelectrodes included the factors group, electrode positionand condition (EE/EC/CE/CC). When appropriate,mean differences between conditions were further evalu-ated with Fisher LSD post-hoc tests.
3. Results
The mean reaction time (RT) to the letter strings forall conditions was 237 ms (SD 26). An ANOVA withgroup as between-subject factor and condition (EE/EC/CE/CC) as within-subject factor revealed a signifi-cant main effect of condition [F(3.54) = 4.70; � = 0.44;p = 0.03]. Post-hoc tests revealed that RTs in conditionsEE and CE were significantly shorter than RTs in con-ditions CC and EC (all p-values < 0.05). Groups did
not differ in RTs [F(3.54) = 0.03; p = 0.99]. Mean RTin both groups for condition EE was 225 ms (SD 28),mean RT for EC was 245 ms (SD 28), mean RT forCE was 232 ms (SD 18), and mean RT for CC was245 ms (SD 23). Error rates did not differ betweengroups ([F(1,18) = 0.01; p = 0.94]).
Based on peak analyses we determined two differenttime windows of interest for the rERN/Ne and fERN/Ne components, respectively. In the difference waves(condition E minus C) the rERN/Ne peaked at electrodeFPz at an average latency of 56 ms (SD 33.3) after but-ton press. ERPs were analyzed response locked startingwith button press in a time window between �20 and140 ms (rERN/Ne) because the rERN/Ne is reportedto peak 100–150 ms after response (Gehring et al.,1993).
The fERN/Ne component peaked at electrode FPz atan average latency of 250 ms (SD 28.1) after the onset ofthe feedback stimulus in the difference waves (conditionE minus C). ERPs were analyzed feedback locked start-ing with the presentation of the feedback stimuli in atime window between 0 and 300 ms because thefERN/Neusually peaks 200–300 ms after negative feed-back (Miltner et al., 1997).
In order to reduce complexity of the results sectiononly effects involving experimental condition (error vs.correct trials) are reported. Main effects and interactionsof purely spatial factors (hemisphere, electrode site),which are not of theoretical interest, are omitted.
3.1. The rERN/Ne and the fERN/Ne component
irrespective of previous trial
For the rERN/Ne component (time window: �20 to140 ms) we found a significant condition effect([F(1,18) = 23.14; p < 0.01]) at midline frontal elec-
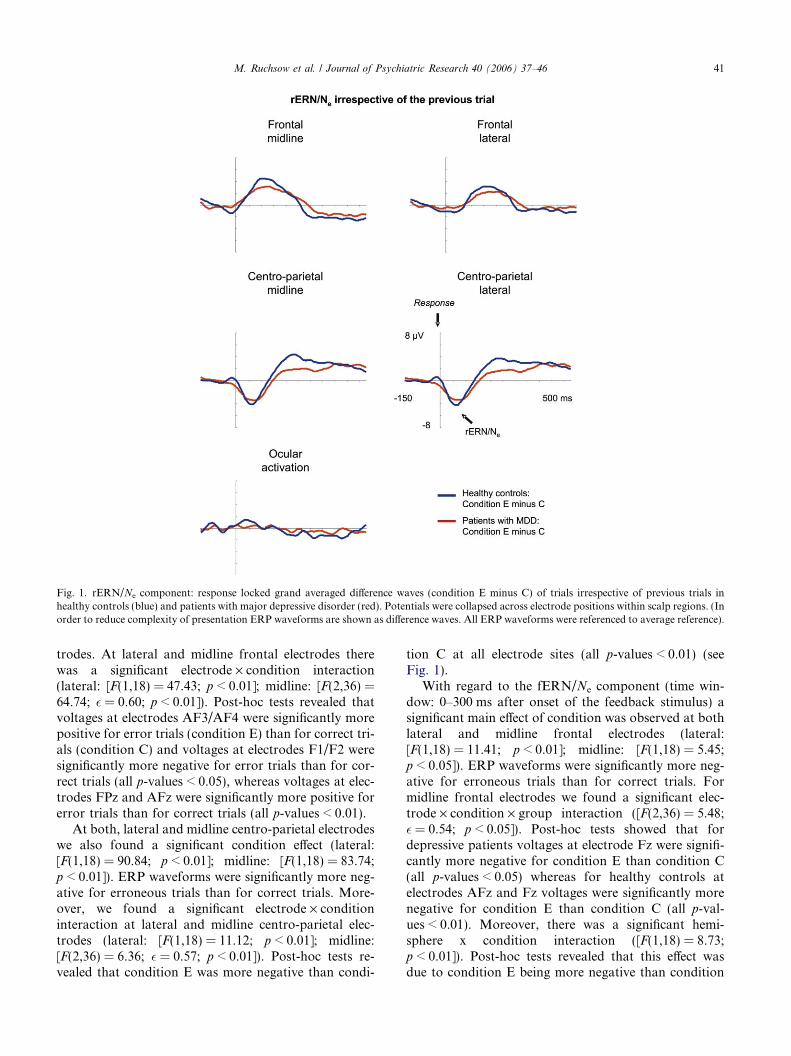
Fig. 1. rERN/Ne component: response locked grand averaged difference waves (condition E minus C) of trials irrespective of previous trials inhealthy controls (blue) and patients with major depressive disorder (red). Potentials were collapsed across electrode positions within scalp regions. (Inorder to reduce complexity of presentation ERP waveforms are shown as difference waves. All ERP waveforms were referenced to average reference).
M. Ruchsow et al. / Journal of Psychiatric Research 40 (2006) 37–46 41
trodes. At lateral and midline frontal electrodes therewas a significant electrode · condition interaction(lateral: [F(1,18) = 47.43; p < 0.01]; midline: [F(2,36) =64.74; � = 0.60; p < 0.01]). Post-hoc tests revealed thatvoltages at electrodes AF3/AF4 were significantly morepositive for error trials (condition E) than for correct tri-als (condition C) and voltages at electrodes F1/F2 weresignificantly more negative for error trials than for cor-rect trials (all p-values < 0.05), whereas voltages at elec-trodes FPz and AFz were significantly more positive forerror trials than for correct trials (all p-values < 0.01).
At both, lateral and midline centro-parietal electrodeswe also found a significant condition effect (lateral:[F(1,18) = 90.84; p < 0.01]; midline: [F(1,18) = 83.74;p < 0.01]). ERP waveforms were significantly more neg-ative for erroneous trials than for correct trials. More-over, we found a significant electrode · conditioninteraction at lateral and midline centro-parietal elec-trodes (lateral: [F(1,18) = 11.12; p < 0.01]; midline:[F(2,36) = 6.36; � = 0.57; p < 0.01]). Post-hoc tests re-vealed that condition E was more negative than condi-
tion C at all electrode sites (all p-values < 0.01) (seeFig. 1).
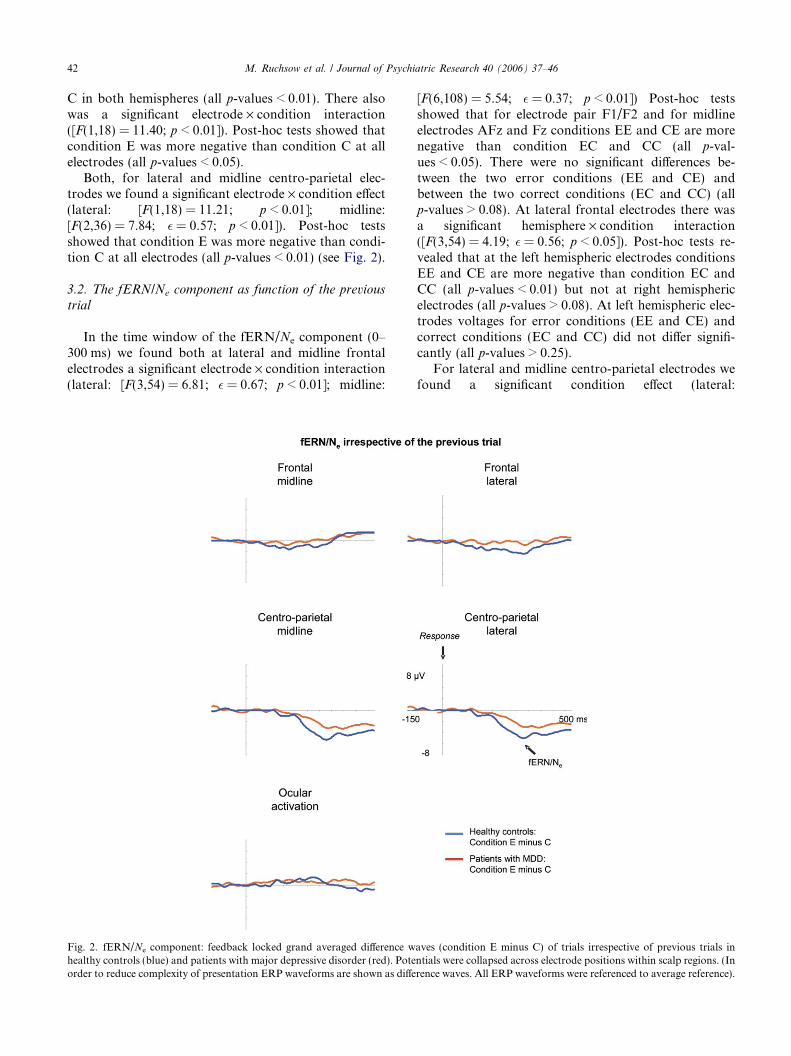
With regard to the fERN/Ne component (time win-dow: 0–300 ms after onset of the feedback stimulus) asignificant main effect of condition was observed at bothlateral and midline frontal electrodes (lateral:[F(1,18) = 11.41; p < 0.01]; midline: [F(1,18) = 5.45;p < 0.05]). ERP waveforms were significantly more neg-ative for erroneous trials than for correct trials. Formidline frontal electrodes we found a significant elec-trode · condition · group interaction ([F(2,36) = 5.48;� = 0.54; p < 0.05]). Post-hoc tests showed that fordepressive patients voltages at electrode Fz were signifi-cantly more negative for condition E than condition C(all p-values < 0.05) whereas for healthy controls atelectrodes AFz and Fz voltages were significantly morenegative for condition E than condition C (all p-val-ues < 0.01). Moreover, there was a significant hemi-sphere x condition interaction ([F(1,18) = 8.73;p < 0.01]). Post-hoc tests revealed that this effect wasdue to condition E being more negative than condition
42 M. Ruchsow et al. / Journal of Psychiatric Research 40 (2006) 37–46
C in both hemispheres (all p-values < 0.01). There alsowas a significant electrode · condition interaction([F(1,18) = 11.40; p < 0.01]). Post-hoc tests showed thatcondition E was more negative than condition C at allelectrodes (all p-values < 0.05).
Both, for lateral and midline centro-parietal elec-trodes we found a significant electrode · condition effect(lateral: [F(1,18) = 11.21; p < 0.01]; midline:[F(2,36) = 7.84; � = 0.57; p < 0.01]). Post-hoc testsshowed that condition E was more negative than condi-tion C at all electrodes (all p-values < 0.01) (see Fig. 2).
3.2. The fERN/Ne component as function of the previous
trial
In the time window of the fERN/Ne component (0–300 ms) we found both at lateral and midline frontalelectrodes a significant electrode · condition interaction(lateral: [F(3,54) = 6.81; � = 0.67; p < 0.01]; midline:
Fig. 2. fERN/Ne component: feedback locked grand averaged difference whealthy controls (blue) and patients with major depressive disorder (red). Poteorder to reduce complexity of presentation ERP waveforms are shown as diffe
[F(6,108) = 5.54; � = 0.37; p < 0.01]) Post-hoc testsshowed that for electrode pair F1/F2 and for midlineelectrodes AFz and Fz conditions EE and CE are morenegative than condition EC and CC (all p-val-ues < 0.05). There were no significant differences be-tween the two error conditions (EE and CE) andbetween the two correct conditions (EC and CC) (allp-values > 0.08). At lateral frontal electrodes there wasa significant hemisphere · condition interaction([F(3,54) = 4.19; � = 0.56; p < 0.05]). Post-hoc tests re-vealed that at the left hemispheric electrodes conditionsEE and CE are more negative than condition EC andCC (all p-values < 0.01) but not at right hemisphericelectrodes (all p-values > 0.08). At left hemispheric elec-trodes voltages for error conditions (EE and CE) andcorrect conditions (EC and CC) did not differ signifi-cantly (all p-values > 0.25).
For lateral and midline centro-parietal electrodes wefound a significant condition effect (lateral:
aves (condition E minus C) of trials irrespective of previous trials inntials were collapsed across electrode positions within scalp regions. (Inrence waves. All ERP waveforms were referenced to average reference).
M. Ruchsow et al. / Journal of Psychiatric Research 40 (2006) 37–46 43
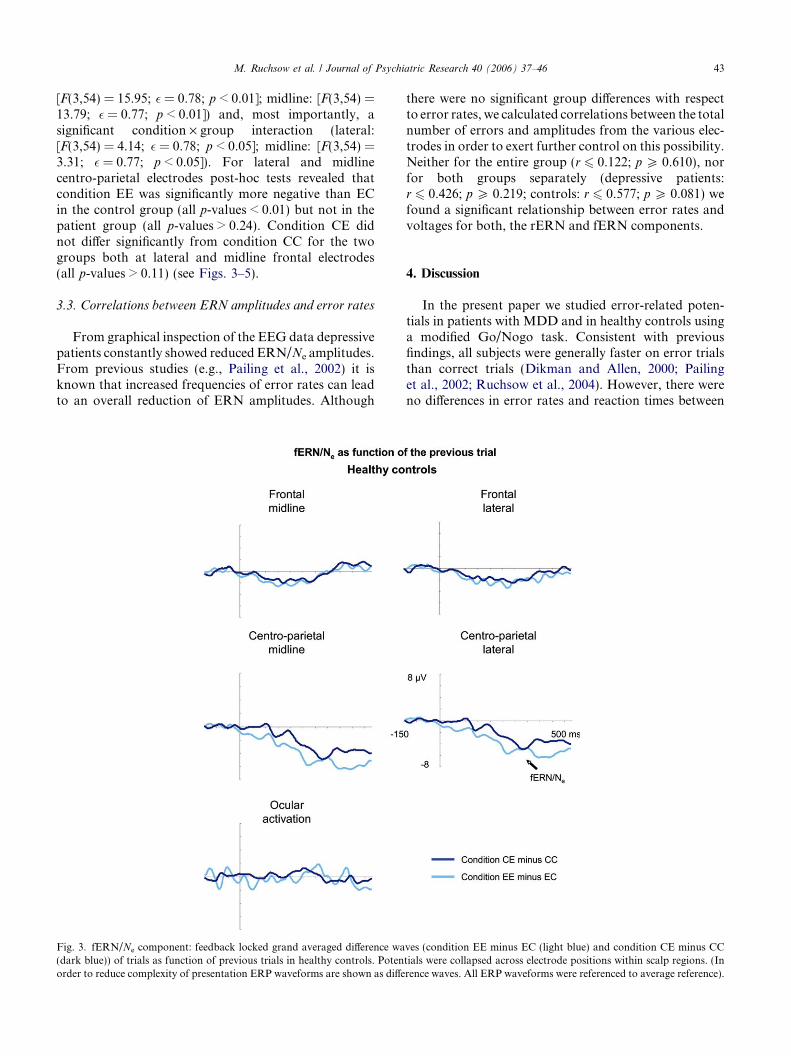
[F(3,54) = 15.95; � = 0.78; p < 0.01]; midline: [F(3,54) =13.79; � = 0.77; p < 0.01]) and, most importantly, asignificant condition · group interaction (lateral:[F(3,54) = 4.14; � = 0.78; p < 0.05]; midline: [F(3,54) =3.31; � = 0.77; p < 0.05]). For lateral and midlinecentro-parietal electrodes post-hoc tests revealed thatcondition EE was significantly more negative than ECin the control group (all p-values < 0.01) but not in thepatient group (all p-values > 0.24). Condition CE didnot differ significantly from condition CC for the twogroups both at lateral and midline frontal electrodes(all p-values > 0.11) (see Figs. 3–5).
3.3. Correlations between ERN amplitudes and error rates
From graphical inspection of the EEG data depressivepatients constantly showed reduced ERN/Ne amplitudes.From previous studies (e.g., Pailing et al., 2002) it isknown that increased frequencies of error rates can leadto an overall reduction of ERN amplitudes. Although
Fig. 3. fERN/Ne component: feedback locked grand averaged difference wa(dark blue)) of trials as function of previous trials in healthy controls. Potenorder to reduce complexity of presentation ERP waveforms are shown as diffe
there were no significant group differences with respectto error rates, we calculated correlations between the totalnumber of errors and amplitudes from the various elec-trodes in order to exert further control on this possibility.Neither for the entire group (r 6 0.122; p P 0.610), norfor both groups separately (depressive patients:r 6 0.426; p P 0.219; controls: r 6 0.577; p P 0.081) wefound a significant relationship between error rates andvoltages for both, the rERN and fERN components.
4. Discussion
In the present paper we studied error-related poten-tials in patients with MDD and in healthy controls usinga modified Go/Nogo task. Consistent with previousfindings, all subjects were generally faster on error trialsthan correct trials (Dikman and Allen, 2000; Pailinget al., 2002; Ruchsow et al., 2004). However, there wereno differences in error rates and reaction times between
ves (condition EE minus EC (light blue) and condition CE minus CCtials were collapsed across electrode positions within scalp regions. (Inrence waves. All ERP waveforms were referenced to average reference).
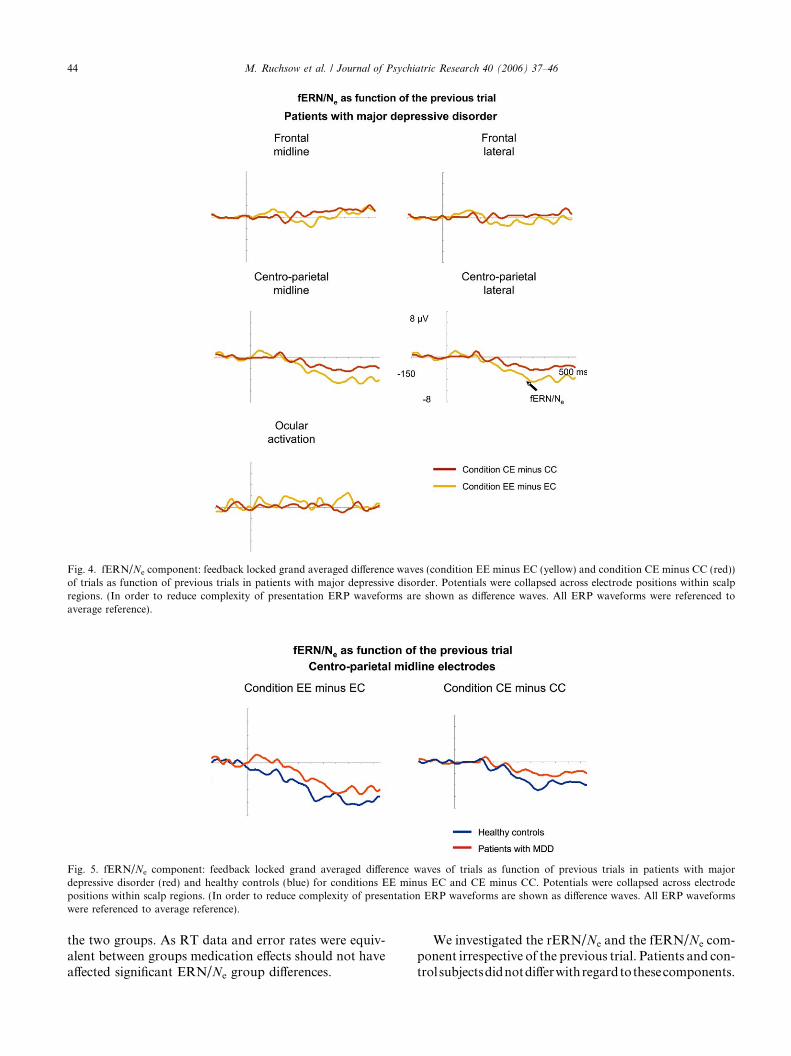
Fig. 4. fERN/Ne component: feedback locked grand averaged difference waves (condition EE minus EC (yellow) and condition CE minus CC (red))of trials as function of previous trials in patients with major depressive disorder. Potentials were collapsed across electrode positions within scalpregions. (In order to reduce complexity of presentation ERP waveforms are shown as difference waves. All ERP waveforms were referenced toaverage reference).
Fig. 5. fERN/Ne component: feedback locked grand averaged difference waves of trials as function of previous trials in patients with majordepressive disorder (red) and healthy controls (blue) for conditions EE minus EC and CE minus CC. Potentials were collapsed across electrodepositions within scalp regions. (In order to reduce complexity of presentation ERP waveforms are shown as difference waves. All ERP waveformswere referenced to average reference).
44 M. Ruchsow et al. / Journal of Psychiatric Research 40 (2006) 37–46
the two groups. As RT data and error rates were equiv-alent between groups medication effects should not haveaffected significant ERN/Ne group differences.
We investigated the rERN/Ne and the fERN/Ne com-ponent irrespective of the previous trial. Patients and con-trolsubjectsdidnotdifferwithregardtothesecomponents.

M. Ruchsow et al. / Journal of Psychiatric Research 40 (2006) 37–46 45
As in our previous study (Ruchsow et al., 2004)we theninvestigated the effect of the performance of the previoustrial (trial n � 1) on the fERN/Ne component elicited bythe current trial (trial n). This approach was guided bystudies of Elliott et al. (1996) who reported a detrimentaleffect on the performance of the subsequent trial due tonegative feedback in the previous trial. ERP waveformsfor four different conditions were calculated: Error trialsafter error trials (EE), correct trials after error trials(EC), error trials after correct trials (CE), and correct tri-als after correct trials (CC). For error trials following er-ror trials (condition EE) we again found a smaller fERN/Ne component in patients with MDD compared tohealthy controls suggesting difficulties in the evaluationof negative feedback.With regard to error trials followingcorrect trials (condition CE) there were no differences ofthe fERN/Ne component between groups.
In line with the previous study, the present data sug-gest that increasing frequencies of error trials might acti-vate a strategic (prefrontal) mechanism which exceedsthe duration of a single trial resulting in higher (morenegative) amplitudes of the fERN/Ne in healthy con-trols. Most importantly, significant group differenceswere found only for erroneous trials following erroneoustrials and not for erroneous trials following correct ones.This might be indicative of deficits in prefrontal functionin MDD leading to impairments in goal-directed behav-ior, strategic reasoning, generation of new rules, andexertion of cognitive control (Carter et al., 1998; David-son et al., 2002; Wolford et al., 2000).
Moreover, the present study shows that our previousfinding of smaller ERN/Ne amplitudes for condition EEin patients with MDD can be replicated in a Go/Nogotask, as well. Both, for errors of choice and errors ofcommission a smaller (less negative) fERN/Ne in pa-tients with MDD but not in healthy controls can be ob-served. This is in good harmony with several studies(e.g., Scheffers et al., 1996) which could not find differ-ences of the ERN/Ne amplitude between errors of choiceand errors of commission. However, besides the rela-tively small sample size, there are certainly some limita-tions of the present study that caution against apremature generalization of the results. Future studieson the same subject should control for variables, likecognitive impairment, drive, or emotional state.
Our results are at variance with those from Holroydand Coles (2002) and Nieuwenhuis et al. (2002) whofound large fERN/Ne amplitudes in trial-and-errorlearning tasks where subjects must rely on the feedbackinformation and small fERN/Ne amplitudes in speededresponse tasks when subjects already know the stimu-lus-response mappings and need not rely on feedbackinformation. We did not vary stimulus-response map-pings in the present study, therefore the impact of thisvariable on the rERN/Ne and fERN/Ne component isbeyond the scope of our paper.
Our data are in accordance with several functionalneuroimaging studies which revealed decreased activa-tion both in the dorsal subdivision of the ACC (Beaure-gard et al., 1998; Bench et al., 1992; Drevets et al., 1997)and the dorsolateral and dorsomedial PFC (Drevets,1998) in patients with MDD. Using source localizationtechniques like BESA (Brain Electric Source Analysis;Scherg and Berg, 1990) especially the source of thefERN/Ne component has been localized in the area ofACC and PFC. This result was confirmed by severalfMRI studies (Carter et al., 1998; Kiehl et al., 2000). Gi-ven the combination of prefrontal and cingulate deficitsin MDD, it seems reasonable to us to expect smaller(less negative) fERN/Ne amplitudes in patients withMDD compared to healthy controls.
Our present investigation was guided by studies ofBeats et al. (1996) and Elliott et al. (1997, 1998, 1996).Beats et al. (1996) described a ‘‘catastrophic responseto executive failure’’ in depressed patients which is mostlikely due to a greater sensitivity to negative feedbackrelative to controls (Elliott et al., 1996). From thesestudies one would expect poorer performance of depres-sive patients following negative feedback. However, inthe present study this prediction could not be observed.Several reasons might account for the absence of this ef-fect among which severity of depression and/or levels oftask difficulty appear to be the most probable factors.Assuming that the detrimental effects after negativefeedback are the cognitive part of learned helplessnessone might speculate that severity of depression and/ortask difficulty were not high enough such that partici-pants could have still the feeling of considerable controlduring the course of the experiment.
The present study replicates and extends our previousresults by demonstrating that the impact of inter-trial ef-fects on the fERN/Ne component in a Go/Nogo taskcan also be observed for errors of commission and isnot restricted to errors of choice. Whether our findingsare specific for patients with MDD or generalize to otherpsychiatric disorders as well remains an open questionand appears to be a reasonable target for future imagingstudies.
References
American Psychiatric Association (APA). Diagnostic and statisticalmanual of mental disorders (DSM-IV). Washington, DC: Amer-ican Psychiatric Association; 1994.
Beats B, Sahakian B, Levy R. Cognitive performance in tests sensitiveto frontal lobe dysfunction in the elderly depressed. PsychologicalMedicine 1996;26:591–603.
Beauregard M, Leroux JM, Bergman S, Arzoumanian Y, Beaudoin G,Bourgouin P, Stip E. The functional neuroanatomy of majordepression: an fMRI study using an emotional activation para-digm. Neuroreport 1998;9:3253–8.
Bench C, Friston K, Brown R, Scott L, Frackowiak R, Dolan R.The anatomy of melancholia-focal abnormalities of cerebral

46 M. Ruchsow et al. / Journal of Psychiatric Research 40 (2006) 37–46
blood flow in major depression. Psychological Medicine 1992;22:607–15.
Braver TS, Cohen JD. On the control of control: the role of dopaminein regulating prefrontal function and working memory. In: MonsellS, Driver J, editors. Control of cognitive processes. Cambridge,Mass: MIT Press; 2000. p. 713–37.
Carter C, Braver T, Barch D, Botvinick M, Noll D, Cohen J. Anteriorcingulate cortex, error detection, and the online monitoring ofperformance. Science 1998;280:747–9.
Carter CS, Macdonald AM, Botvinick M, Ross LL, Stenger VA, NollD, Cohen JD. Parsing executive processes: strategic vs. evaluativefunctions of the anterior cingulate cortex. Proceedings of theNational Academy of Sciences of the United States of America2000;97:1944–8.
Coles MGH, Scheffers MK, Holroyd CB. Why is there an ERN/Ne oncorrect trials? Response representations, stimulus-related compo-nents, and the theory of error-processing. Biological Psychology2001;56:173–89.
Davidson RJ, Pizzagalli D, Nitschke JB, Putnam K. Depression:perspectives from affective neuroscience. Annual Review of Psy-chology 2002;53:545–74.
Dehaene S, Posner M, Tucker D. Commentary: localization of aneural system for error detection and compensation. PsychologicalMedicine 1994;5:303–5.
Dikman Z, Allen J. Error monitoring during reward and avoidancelearning in high- and low-socialized individuals. Psychophysiology2000;37:43–54.
Drevets WC. Functional neuroimaging studies of depression: theanatomy of melancholia. Annual Review of Medicine1998;49:341–61.
Drevets WC, Price JL, Simpson Jr JR, Todd RD, Reich T, Vannier M,Raichle ME. Subgenual prefrontal cortex abnormalities in mooddisorders. Nature 1997;386:824–7.
Elliott R, Baker S, Rogers R, O�Leary D, Paykel E, Frith C, Dolan R,Sahakian B. Prefrontal dysfunction in depressed patients perform-ing a complex planning task: a study using positron emissiontomography. Psychological Medicine 1997;27:931–42.
Elliott R, Sahakian B, Michael A, Paykel E, Dolan R. Abnormalneural response to feedback on planning and guessing tasks inpatients with unipolar depression. Psychological Medicine1998;28:559–71.
Elliott R, Sahakian BJ, McKay AP, Herrod JJ, Robbins TW, PaykelES. Neuropsychological impairments in unipolar depression: theinfluence of perceived failure on subsequent performance. Psycho-logical Medicine 1996;26:975–89.
Falkenstein M, Hohnsbein J, Hoormann J, Blanke L. Effects of errorsin choice reaction tasks on the ERP under focused and dividedattention. In: Brunia C, Gaillard A, Kok A, editors. Psychophys-iological brain research. Tilburg: Tilburg University Press; 1990.p. 192–5.
Falkenstein M, Hoormann J, Hohnsbein J. ERP components in Go/Nogo tasks and their relation to inhibition. Acta Psychologica(Amsterdam) 1999;101:267–91.
Gehring WJ, Coles MGH, Meyer DE, Donchin E. The error-relatednegativity: an event-related brain potential accompanying errors(abstract). Psychophysiology 1990;27:S34.
Gehring W, Goss B, Coles M, Meyer D, Donchin E. A neural systemfor error detection and compensation. Psychological Science1993;4:385–90.
Goodman WK, Price LH, Rasmussen SA, Mazure C, FleischmannRL, Hill CL, Heninger GR, Charney DS. The Yale-Brown
obsessive compulsive scale. I. Development, use, and reliability.Archives of General Psychiatry 1989;46:1006–11.
Hamilton M. A rating scale for depression. Journal of NeurologyNeurosurgery and Psychiatry 1960;23:56–62.
Hamilton M. HAMA. The Hamilton Anxiety Scale. In: Guy W,editor. ECDEU Assessment manual for psychopharmacology,revised edition. Maryland: Rockville; 1976.
Holroyd CB, Coles MGH. The neural basis of human error process-ing: reinforcement learning, dopamine, and the error-relatednegativity. Psychological Review 2002;109:679–709.
Kiehl KA, Liddle PF, Hopfinger JB. Error processing and the rostralanterior cingulate: an event-related fMRI study. Psychophysiology2000;37:216–23.
Mayberg HS, Brannan SK, Mahurin RK, Jerabek PA, Brickman JS,Tekell JL, Silva JA, McGinnis S, Glass TG, Martin CC, Fox PT.Cingulate function in depression: a potential predictor of treatmentresponse. Neuroreport 1997;8:1057–61.
Miltner W, Braun C, Coles MGH. Event-related brain potentialsfollowing incorrect feedback in a time-estimation task: evidence fora generic neural system for error detection. Journal of CognitiveNeuroscience 1997;9:788–98.
Nieuwenhuis S, Ridderinkhof KR, Talsma D, Coles MGH, HolroydCB, Kok A, van der Molen MW. A computational account ofaltered error processing in older age: dopamine and the error-related negativity. Cognitive, Affective and Behavioral Neurosci-ence 2002;2:19–36.
Oldfield RC. The assessment and analysis of handedness: the Edin-burgh inventory. Neuropsychologia 1971;9:97–113.
Overall JE, Gorham DR. The brief psychiatric rating scale. Psycho-logical Reports 1962;10:799–812.
Pailing PE, Segalowitz SJ, Dywan J, Davies PL. Error negativity andresponse control. Psychophysiology 2002;39:198–206.
Pizzagalli D, Pascual-Marqui RD, Nitschke JB, Oakes TR, LarsonCL, Abercrombie HC, Schaefer SM, Koger JV, Benca RM,Davidson RJ. Anterior cingulate activity as a predictor of degree oftreatment response in major depression: evidence from brainelectrical tomography analysis. American Journal of Psychiatry2001;158:405–15.
Rogers RD, Owen AM, Middleton HC, Williams EJ, Pickard JD,Sahakian BJ, Robbins TW. Choosing between small, likely rewardsand large, unlikely rewards activates inferior and orbital prefrontalcortex. Journal of Neuroscience 1999;19:9029–38.
RuchsowM, Grothe J, Spitzer M, Kiefer M. Human anterior cingulatecortex is activated by negative feedback: evidence from event-related potentials in a guessing task. Neuroscience Letters2002;325:203–6.
Ruchsow M, Herrnberger B, Wiesend C, Gron G, Spitzer M, KieferM. The effect of erroneous responses on response monitoring inpatients with major depressive disorder: a study with event relatedpotentials. Psychophysiology 2004;41:833–40.
Scheffers MK, Coles MGH, Bernstein P, Gehring WJ, Donchin E.Event-related brain potentials and error-related processing: ananalysis of incorrect responses to go and no-go stimuli. Psycho-physiology 1996;33:42–53.
Scherg M, Berg P. BESA-Brain electric source analysis hand-book. Munich: Max-Planck Institute for Psychiatry; 1990.
Schultz W, Apicell P, Scarnati E, Ljungberg T. Neuronal activity inmonkey ventral striatum related to the expectation of reward.Journal of Neuroscience 1992;12:4595–610.
Wolford G, Miller MB, Gazzaniga M. The left hemisphere�s role inhypothesis formation. Journal of Neuroscience 2000;20:RC64.





























