Embed Size (px)
Citation preview

ELSEVIER Mutation Research 317 (1994) 111-132
m
Genetic Toxicology
Ethylene thiourea (ETU). A review of the genetic toxicity studies
Kerry L. Dear f ie ld *
U.S. Environmental Protection Agency, Office of Pesticide Programs, Health Effects Division (7509C), 401 M St. S. W, Washington, DC 20460, USA
(Received 15 June 1993) (Accepted 26 October 1993)
Abstract
Ethylene thiourea (ETU) is a common contaminant, metabolite and degradation product of the fungicide class of ethylene bisdithiocarbamates (EBDCs); as such, they present possible exposure and toxicological concerns to exposed individuals. ETU has been assayed in many different tests to assess genotoxicity activity. While a great number of negative results are found in the data base, there is evidence that demonstrates ETU is capable of inducing genotoxic endpoints. These include responses for gene mutations (e.g. Salmonella), structural chromosomal alterations (e.g. aberrations in cultured mammalian cells as well as a dominant lethal assay) and other genotoxic effects (e.g. bacterial rec assay and several yeast assays).
It is important to consider the magnitude of the positive responses as well as the concentrations/doses used when assessing the genotoxicity of ETU. While ETU induces a variety of genotoxic endpoints, it does not appear to be a potent genotoxic agent. For example, it is a weak bacterial mutagen in the Salmonella assay without activation in strain TA1535 at concentrations generally above 1000/zg/plate. Weak genotoxic activity of this sort is usually observed in most of the assays with positive results. Since ETU does not appear very potent and is not extremely toxic to test cells and organisms, it is not surprising to find that ETU does not produce consistent effects in many of the assays reviewed. Consequently, in many instances, mixed results for the same assay type are reported by different investigators, but as reviewed herein, these results may be dependent upon the test conditions in each individual laboratory. A primary shortcoming with many of the reported negative results is that the concentrations or doses used are not high enough for an adequate test for ETU activity. There are also problems with many of the negative assays generally in protocol or reporting, particularly with the in vivo studies (e.g. inappropriate sample number a n d / o r sampling times; inadequate top dose employed).
Overall, while ETU does not appear to be a potent genotoxic agent, it is capable of producing genotoxic effects (e.g. gene mutations, structural chromosomal aberrations). This provides a basis for weak genotoxic activity by ETU. Furthermore, based on a suggestive dominant lethal positive result, there may be a concern for heritable effects. Due to the many problems with the conduct and assessment of the in vivo assays, it is worth repeating in vivo
* Corresponding author, Tel.703 305 6780; Fax 703 305 5147. This manuscript has been reviewed by the Office of Preven- tion, Pesticides and Toxic Substances, U.S. Environmental Protection Agency and approved for publication. Approval
does not signify that the contents necessarily reflect the views or policies of the Agency, nor does mention of trade names or commercial products constitute endorsement or recommenda- tion for use.
0165-1110/94/$26.00 © 1994 Elsevier Science B.V. All rights reserved SSDI 0165-1 1 10(93)E0018-S

112 K.L. Dearfield /Mutation Research 317 (1994) 111-132
cytogenetic assays and a dominant lethal assay (with acceptable test procedures and data generation) to determine if these results would continue to support a heritable mutagenicity concern.
Key words: Ethylene thiourea; Genetic toxicity; Review
Contents
A b s t r a c t . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 111
1. I n t r o d u c t i o n . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 112
2. A s s a y e v a l u a t i o n s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 119
(A) B a c t e r i a l a ssays . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
(B) Y e a s t assays . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 122
(C) D r o s o p h i l a assays . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 123
(D) M a m m a l i a n cells in c u l t u r e - - g e n e m u t a t i o n assays . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 123
(E) M a m m a l i a n cel ls in c u l t u r e - - a b e r r a t i o n assays . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 124
(F) In vivo c y t o g e n e t i c s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 124
( G ) D o m i n a n t l e tha l assays . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 125
( H ) U n s c h e d u l e d D N A syn thes i s ( U D S ) assays . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 125
(I) S i s t e r - c h r o m a t i d e x c h a n g e ( S C E ) assays . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 126
(J) Cell t r a n s f o r m a t i o n assays . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 126
(K) G e r m cell a ssays . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 126
(L) O t h e r geno tox ic i t y assays . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 127
(M) E T U n i t r o s a t i o n p r o d u c t s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 127
3. O t h e r c o n s i d e r a t i o n s a n d c o n c l u s i o n . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 128
4. A c k n o w l e d g e m e n t . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 129
5. R e f e r e n c e s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 129
I. Introduction
Ethylene thiourea (ETU; CAS Registry No. 96-45-7; chemical name 2-imidazolidinethione) is a heterocyclic thionamide (Fig. 1). ETU introduc- tion into the environment occurs mainly through the use of ethylene bisdithiocarbamates (EBDCs) which are a group of fungicides used to control fungal pathogens (NTP, 1992; USEPA, 1992). E TU is a common contaminant, metabolite and
NH
Fig. 1. S t r u c t u r e o f e t h y l e n e t h i o u r e a ( E T U ) .
degradation product of these fungicides (e.g. see Engst and Schnaak, 1974; Jordan and Neal, 1979; Marshall, 1977; Truhaut et al., 1973). The general population is exposed to ETU primarily from food exposed to EBDCs. It is estimated that 8-12 million pounds of EBDCs are used in the United States per year (USEPA, 1992). Thus the use of and potential exposure to ETU may present envi- ronmental, dietary and occupational exposure concerns.
Several potential toxicity concerns are associ- ated with exposure to ETU. These include car- cinogenicity, developmental toxicity, thyroid toxi- city and mutagenicity. The evidence for carcino- genicity demonstrates that ETU increases the incidence of thyroid follicular cell adenomas and adenocarcinomas in both sexes of mice and rats
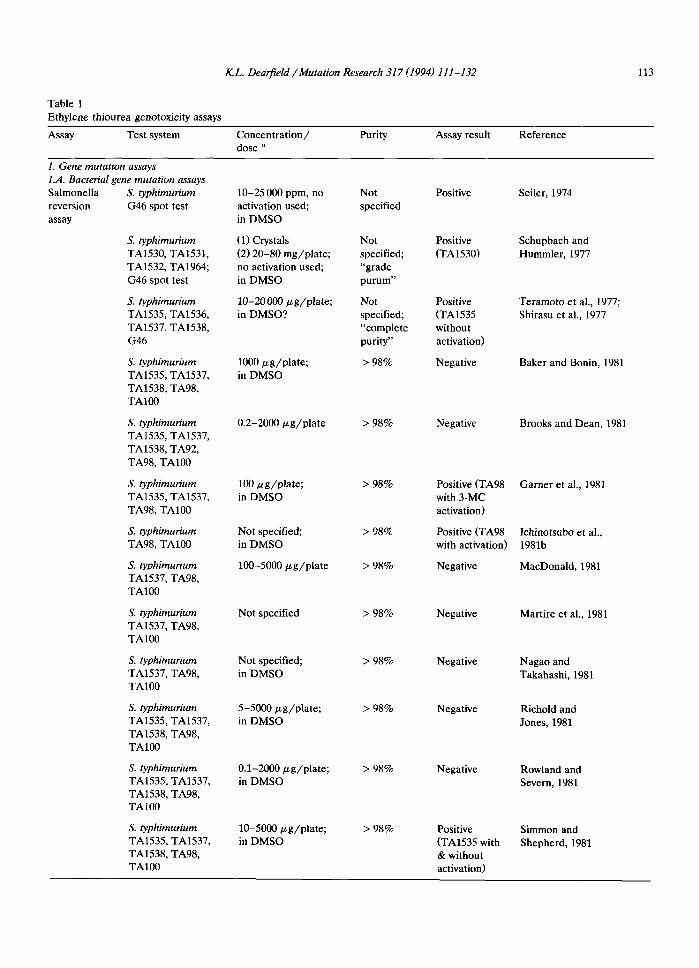
I( L. DearfieM / Mutation Research 317 (1994) 111 - 132 113
Table 1 Ethylene thiourea genotoxicity assays
Assay Test system Concentration/ Purity Assay result Reference dose a
1. Gene mutation assays 1.A. Bacterial gene mutation assays Salmonella S. typhimurium 10-25 000 ppm, no Not reversion G46 spot test activation used; specified assay in DMSO
Positive Seiler, 1974
S. typhimurium (1) Crystals Not Positive TA1530, TA1531, (2) 20-80 mg/plate; specified; (TA1530) TA1532, TA1964; no activation used; "grade G46 spot test in DMSO purum"
S. typhimurium 10-20000 #g/p la te ; Not Positive TA1535, TA1536, in DMSO? specified; (TA1535 TA1537, TA1538, "complete without G46 purity" activation)
S. typhimurium 1000/~g/plate; > 98% Negative TA1535, TA1537, in DMSO TA1538, TA98, TA100
S. typhimurium 0.2-2000/.tg/plate > 98% Negative TA1535, TA1537, TA1538, TA92, TA98, TA100
S. typhimurium 100 ~g/pla te ; > 98% Positive (TA98 TA1535, TA1537, in DMSO with 3-MC TA98, TA100 activation)
S. typhimurium Not specified; > 98% Positive (TA98 TA98, TA100 in DMSO with activation)
S. typhimurium 100-5000 ~g /p la te > 98% Negative TA1537, TA98, TA100
S. typhimurium Not specified > 98% Negative TA1537, TA98, TA100
S. typhimurium Not specified; > 98% Negative TA1537, TA98, in DMSO TA100
S. typhimurium 5-5000/zg/plate; > 98% Negative TA1535, TA1537, in DMSO TA1538, TA98, TA100
S. typhimurium 0.1-2000/~g/plate; > 98% Negative TA1535, TA1537, in DMSO TA1538, TA98, TA100
S. typhimurium 10-5000/zg/plate; > 98% Positive TA1535, TA1537, in DMSO (TA1535 with TA1538, TA98, & without TA100 activation)
Schupbach and Hummler, 1977
Teramoto et al., 1977; Shirasu et al., 1977
Baker and Bonin, 1981
Brooks and Dean, 1981
Garner et al., 1981
Ichinotsubo et al., 1981b
MacDonald, 1981
Martire et al., 1981
Nagao and Takahashi, 1981
Richold and Jones, 1981
Rowland and Severn, 1981
Simmon and Shepherd, 1981
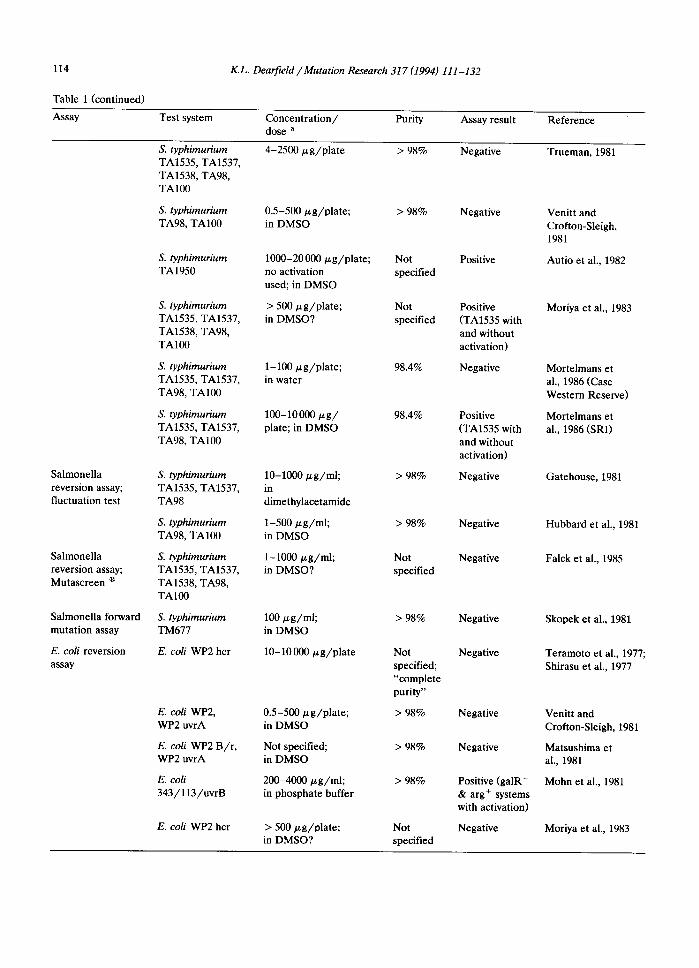
114 K.L. Dearfield /Mutation Research 317 (1994) 111-132
Table 1 (continued)
Assay
Salmonella reversion assay; fluctuation test
Salmonella reversion assay; Mutascreen ®
Salmonella forward mutation assay
E. coli reversion assay
Test system
S. typhimurium TA1535, TA1537, TA1538, TA98, TA100
S. typhimurium TA98, TA100
S. typhimurium TA1950
S. typhimurium TA1535, TA1537, TA1538, TA98, TA100
S. typhimurium TA1535, TA1537, TA98, TA100
S. typhimurium TA1535, TA1537, TA98, TA100
S. typhimurium TA1535, TA1537, TA98
S. typhimurium TA98, TA100
S. typhimurium TA1535, TA1537, TA1538, TA98, TA100
S. typhimurium TM677
E. coli WP2 hcr
E. coli WP2, WP2 uvrA
E. coli WP2 B/r , WP2 uvrA
E. coli 343/113/uvrB
E. coli WP2 hcr
Concentration/ Purity dose a
4-2500/xg/plate > 98%
0.5-500/zg/plate; > 98% in DMSO
1000-20 000 #g/pla te ; Not no activation specified used; in DMSO
> 500/~g/plate; Not in DMSO? specified
1-100/zg/plate; 98.4% in water
100-10 000/zg / 98.4% plate; in DMSO
10-1000/zg/ml; > 98% in dimethylacetamide
1-500 gg /ml ; > 98% in DMSO
1-1000 brg/ml; Not in DMSO? specified
100 gg /ml ; > 98% in DMSO
10-10 000 p,g/plate Not specified; "complete purity"
0.5-500 gg/pla te ; > 98% in DMSO
Not specified; > 98% in DMSO
200-4000 ~g /ml ; > 98% in phosphate buffer
> 500/zg/plate; Not in DMSO? specified
Assay result
Negative
Negative
Positive
Positive (TA1535 with and without activation)
Negative
Positive (TA1535 with and without activation)
Negative
Negative
Negative
Negative
Negative
Negative
Negative
Positive (galR- & arg + systems with activation)
Negative
Reference
Trueman, 1981
Venitt and Crofton-Sleigh, 1981
Autio et al., 1982
Moriya et al., 1983
Mortelmans et al., 1986 (Case Western Reserve)
Mortelmans et al., 1986 (SRI)
Gatehouse, 1981
Hubbard et al., 1981
Falck et al., 1985
Skopek et al., 1981
Teramoto et al., 1977; Shirasu et al., 1977
Venitt and Crofton-Sieigh, 1981
Matsushima et al., 1981
Mohn et al., 1981
Moriya et al., 1983
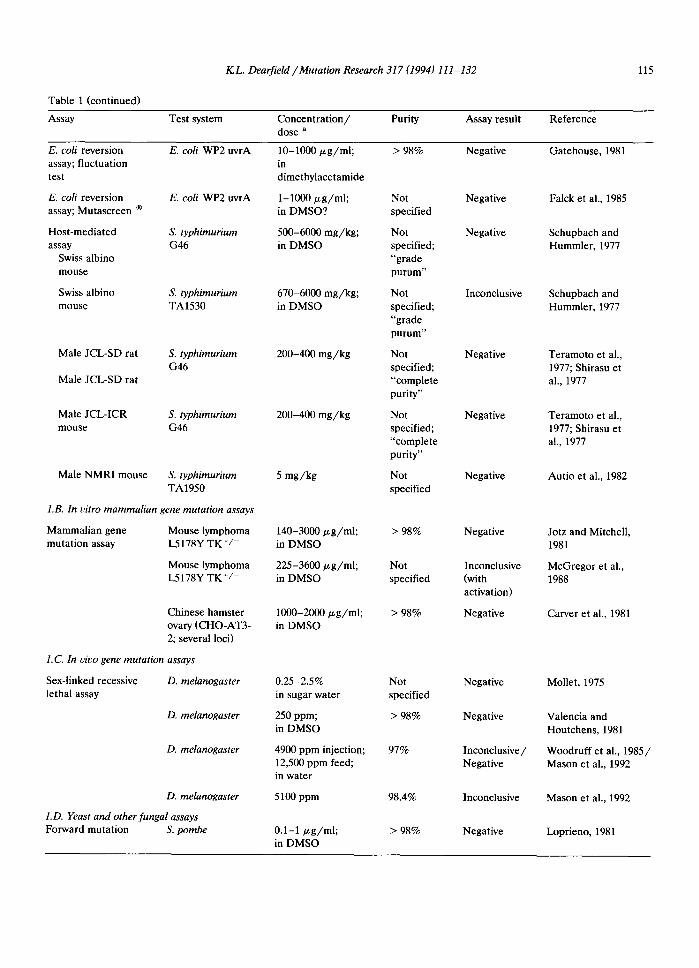
KL. Dearfield /Mutation Research 317 (1994) 111-132 115
Table 1 (continued)
Assay Test system Concentration/ Purity Assay result Reference dose a
E. coli reversion E. coli WP2 uvrA 10-1000 p,g/ml; > 98% Negative Gatehouse, 1981 assay; fluctuation in test dimethylacetamide
E. coli reversion E. coli WP2 uvrA 1-1000 ~g/ml; Not Negative Falck et al., 1985 assay; Mutascreen ® in DMSO? specified
Host-mediated S. typhimurium 500-6000 mg/kg; Not Negative Schupbach and assay G46 in DMSO specified; Hummler, 1977
Swiss albino "grade mouse purum"
Swiss albino S. typhirnurium 670-6000 mg/kg; Not Inconclusive Schupbach and mouse TA1530 in DMSO specified; Hummler, 1977
Male JCL-SD rat S. typhimurium 200-400 mg/kg G46
Male JCL-SD rat
Male JCL-ICR S. typhimurium 200-400 mg/kg mouse G46
Male NMRI mouse S. typhimurium TA1950
1.B. In vitro mammalian gene mutation assays
Mammalian gene Mouse lymphoma mutation assay L5178Y TK ÷/-
Mouse lymphoma L5178Y TK +/-
"grade purum"
Not specified; "complete purity"
Not specified; "complete purity"
Not specified
Chinese hamster ovary (CHO-AT3- 2; several loci)
1.C. In vivo gene mutation assays
Sex-linked recessive D. melanogaster lethal assay
D. melanogaster
1). melanogaster
1). melanogaster
I.D. Yeast and other fungal assays Forward mutation S. pombe
5 mg/kg
140-3000/.~g/ml; in DMSO
225-3600/xg/ml; in DMSO
1000-2000/.~g/ml; in DMSO
0.25-2.5% in sugar water
250 ppm; in DMSO
4900 ppm injection; 12,500 ppm feed; in water
5100 ppm
0.1-1/xg/ml; in DMSO
> 98%
Not specified
> 98%
Not specified
> 98%
97%
98.4%
> 98%
Negative
Negative
Negative
Negative
Inconclusive (with activation)
Negative
Negative
Negative
Inconclusive/ Negative
Inconclusive
Negative
Teramoto et al., 1977; Shirasu et al., 1977
Teramoto et al., 1977; Shirasu et al., 1977
Autio et al., 1982
Jotz and Mitchell, 1981
McGregor et al., 1988
Carver et al., 1981
Mollet, 1975
Valencia and Houtchens, 1981
Woodruff et al., 1985/ Mason et al., 1992
Mason et al., 1992
Loprieno, 1981
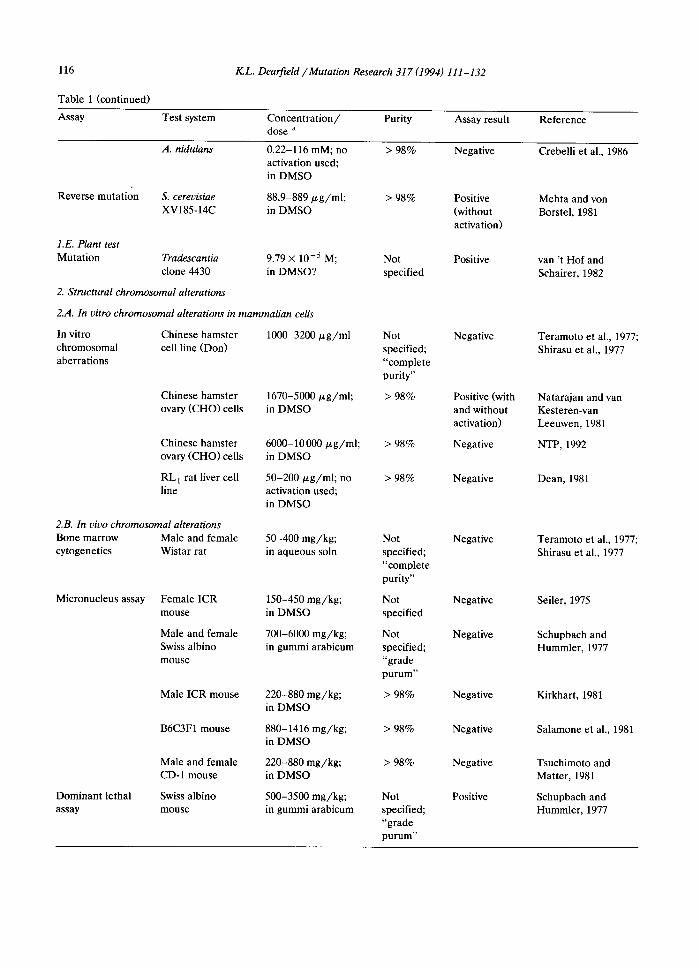
116
Table 1 (continued)
Assay Test system
K.L. DearfieM /Mutation Research 317 (1994) 111-132
Concentration/ Purity Assay result dose a
Reference
Reverse mutation
A. nidulans
S. cerevisiae XV185-14C
0.22-116 mM; no activation used; in DMSO
> 98% Negative
88.9-889/zg/ml; > 98% Positive in DMSO (without
activation)
I.E. Plant test Mutation Tradescantia 9.79 x 10 -5 M;
clone 4430 in DMSO?
2. Structural chromosomal alterations
2M. In vitro chromosomal alterations in mammalian cells
In vitro Chinese hamster 1000-3200/xg/ml chromosomal cell line (Don) aberrations
Chinese hamster 1670-5000/zg/ml; ovary (CHO) cells in DMSO
Chinese hamster 6000-10000/zg/ml; ovary (CHO) cells in DMSO
RL I rat liver cell 50-200/zg/ml; no line activation used;
in DMSO
50-400 mg/kg; in aqueous soln
2.B. In vivo chromosomal alterations Bone marrow Male and female cytogenetics Wistar rat
Micronucleus assay Female ICR 150-450 mg/kg; mouse in DMSO
Male and female 700-6000 mg/kg; Swiss albino in gummi arabicum mouse
Male ICR mouse 220-880 mg/kg; in DMSO
B6C3F1 mouse 880-1416 mg/kg; in DMSO
Male and female 220-880 mg/kg; CD-1 mouse in DMSO
Dominant lethal Swiss albino 500-3500 mg/kg; assay mouse in gummi arabicum
Not Positive specified
Not specified; "complete purity"
> 98%
> 98%
> 98%
Not specified; "complete purity"
Not specified
Not specified; "grade purum"
> 98%
> 98%
> 98%
Not specified; "grade purum"
Negative
Positwe(with and without activation)
Negative
Negative
Negative
Negatwe
Negatwe
Negative
Negative
Negative
Positive
Crebelliet a1.,1986
Mehta and von Borstel, 1981
van 't Hofand Schairer, 1982
Teramoto et al., 1977; Shirasu et al., 1977
Natar~an and van Kesteren-van Leeuwen, 1981
NTP, 1992
Dean, 1981
Teramoto et al., 1977; Shirasu et al., 1977
Seiler, 1975
Schupbach and Hummler, 1977
Kirkhart, 1981
Salamone et al., 1981
Tsuchimoto and Matter, 1981
Schupbach and Hummler, 1977
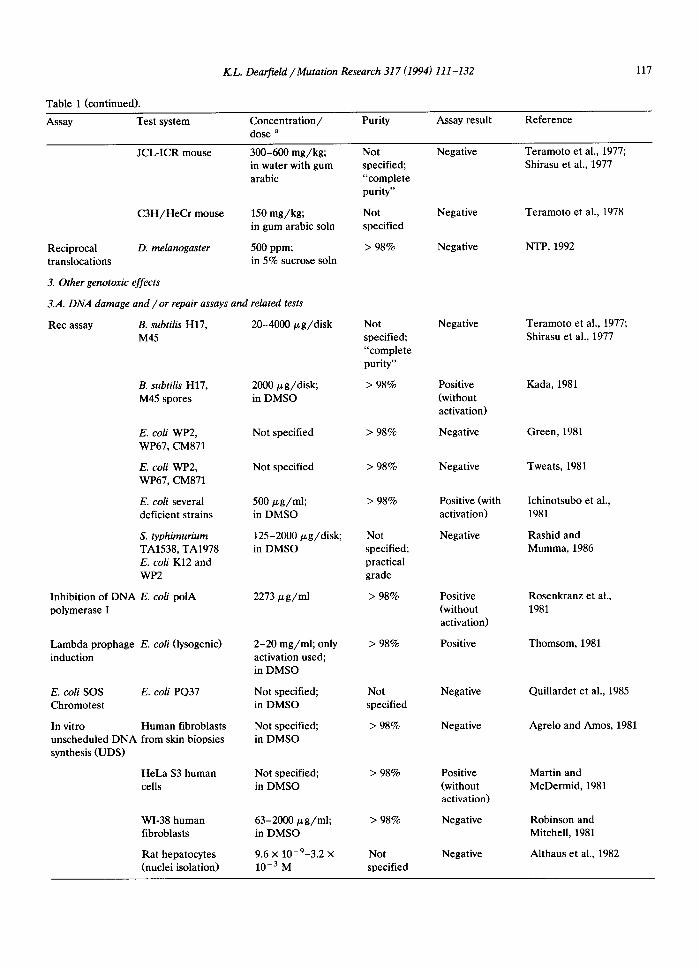
K.L. Dearfield /Mutation Research 317 (1994) 111-132 117
Table 1 (continued).
Assay Test system Concentration/ Purity Assay result Reference dose a
JCL-ICR mouse 300-600 mg/kg; Not Negative Teramoto et al., 1977; in water with gum specified; Shirasu et al., 1977 arabic "complete
purity"
C3H/HeCr mouse 150 mg/kg; Not Negative Teramoto et al., 1978 in gum arabic soln specified
Reciprocal D. melanogaster 500 ppm; > 98% Negative NTP, 1992 translocations in 5% sucrose soln
3. Other genotoxic effects
3.4. DNA damage a n d / o r repair assays and related tests
Rec assay B. subtilis H17, 20-4000 ~g/disk M45
B. subtilis H17, 2000/zg/disk; M45 spores in DMSO
E. coli WP2, WP67, CM871
E. coli WP2, WP67, CM871
E. coli several deficient strains
S. typhimurium TA1538, TA1978 E. coli K12 and WP2
Inhibition of DNA E. coli polA polymerase I
Lambda prophage E. coli (lysogenic) induction
Not specified
Not specified
500 ~g/ml; in DMSO
125-2000/zg/disk; in DMSO
2273/zg/ml
2-20 mg/ml; only activation used; in DMSO
E. coli SOS E. coli PQ37 Not specified; Chromotest in DMSO
In vitro Human fibroblasts Not specified; unscheduled DNA from skin biopsies in DMSO synthesis (UDS)
HeLa $3 human Not specified; cells in DMSO
WI-38 human fibroblasts
Rat hepatocytes (nuclei isolation)
63-2000/.L g/ml; in DMSO
9.6 × 10-9-3.2 × 10 -3 M
Not specified; "complete purity"
> 98%
> 98%
> 98%
> 98%
Not specified; practical grade
> 98%
> 98%
Not specified
> 98%
> 98%
> 98%
Not specified
Negative Teramoto et al., 1977; Shirasu et al., 1977
Positive Kada, 1981 (without activation)
Negative Green, 1981
Negative Tweats, 1981
Positive (with Ichinotsubo et al., activation) 1981
Negative Rashid and Mumma, 1986
Positive Rosenkranz et al., (without 1981 activation)
Positive Thomsom, 1981
Negative Quillardet et al., 1985
Negative Agrelo and Amos, 1981
Positive Martin and (without McDermid, 1981 activation)
Negative Robinson and Mitchell, 1981
Negative Althaus et al., 1982
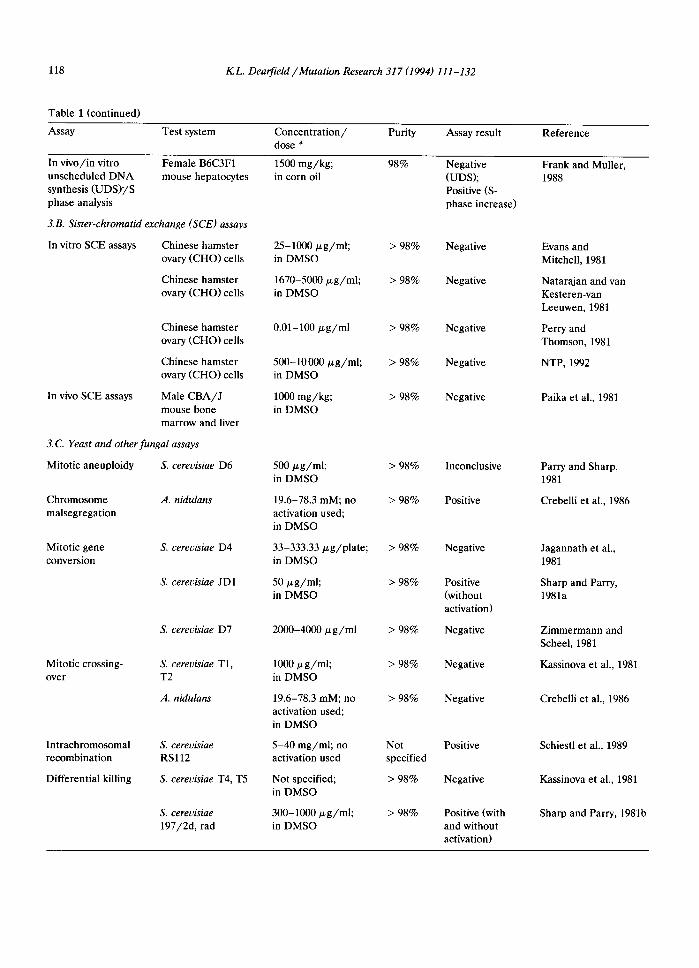
118 K L . Dearfield /Mutat ion Research 317 (1994) 111-132
Table 1 (continued)
Assay Test system Concentration/ Purity Assay result Reference dose a
In vivo/in vitro Female B6C3F1 1500 mg/kg; 98% Negative Frank and Muller, unscheduled DNA mouse hepatocytes in corn oil (UDS); 1988 synthesis (UDS)7'S Positive (S- phase analysis phase increase)
3.B. Sister-chromatid exchange (SCE) assays
In vitro SCE assays Chinese hamster 25-1000/zg/ml; > 98% Negative Evans and ovary (CHO) cells in DMSO Mitchell, 1981
Chinese hamster 1670-5000/xg/ml; > 98% Negative Natarajan and van ovary (CHO) cells in DMSO Kesteren-van
Leeuwen, 1981
Perry and Thomson, 1981
NTP, 1992
Chinese hamster ovary (CHO) cells
Chinese hamster ovary (CHO) cells
In vivo SCE assays Male CBA/J mouse bone marrow and liver
3.C. Yeast and other fungal assays
Mitotic aneuploidy S. cerevisiae D6
Chromosome A. nidulans malsegregation
Mitotic gene S. cerevisiae D4 conversion
S. cerevisiae JD1
Mitotic crossing- over
S. cerevisiae D7
S. cerevisiae T1, T2
A. nidulans
Intrachromosomal recombination
Differential killing
S. cerevisiae RSl12
S. cerevisiae T4, T5
S. cerevisiae 197/2d, rad
0.01-100/zg/ml > 98% Negative
500-10000/xg/ml; > 98% Negative in DMSO
1000 mg/kg; > 98% Negative in DMSO
500 ~g/ml; > 98% Inconclusive in DMSO
19.6-78.3 mM; no > 98% Positive activation used; in DMSO
33-333.33/zg/plate; > 98% Negative in DMSO
50/xg/ml; > 98% Positive in DMSO (without
activation)
2000-4000/zg/ml > 98% Negative
1000/xg/ml; > 98% Negative in DMSO
19.6-78.3 mM; no > 98% Negative activation used; in DMSO
5-40 mg/ml; no Not Positive activation used specified
Not specified; > 98% Negative in DMSO
300-1000 tzg/ml; > 98% Positive (with in DMSO and without
activation)
Paika et al., 1981
Parry and Sharp, 1981
Crebelli et al., 1986
Jagannath et al., 1981
Sharp and Parry, 1981a
Zimmermann and Scheel, 1981
Kassinova et al., 1981
Crebelli et al., 1986
Schiestl et al., 1989
Kassinova et al., 1981
Sharp and Parry, 1981b

K.L. Dear)qeld / Mutation Research 317 (1994) 111-132 119
Table 1 (continued)
Assay Test s y s t e m Concentration/ Purity Assay result Reference dose a
3.D. Cell transformation assays
Cell transformation C3H/10T 1/2 cells
Cell transformation with "promotion"
3.E. Germ cell effects Inhibition of testicular DNA synthesis
Spermhead abnormalities
Syrian hamster embryo (SHE) cells/adenovirus SA7
Syrian hamster embryo (SHE) cells/adenovirus SA7
C3H/10T 1/2 cells
100-1000 p.g/ml; 99.8% in DMSO
62-1000 ~g/ml Not specified
1-24 mM Not specified
Negative McGlynn-Kreft and McCarthy, 1984
Negative Casto, 1975, 1976 in: Heidelberger et al., 1983
Inconclusive Hatch et al., 1986
333 p.g/ml; 99.8% Negative McLeod and in DMSO Doolittle, 1985
Mouse 100 mg/kg Not Negative Seiler, 1977a specified
(CBA X BALB/c) 250-2000 mg/kg > 98% Negative Topham, 1981 F1 mouse in Tween 80
B6C3F1/CRL 166-2655 mg/kg; > 98% N e g a t i v e Wyrobek et al., 1981 mouse in DMSO
a In vitro assays performed with and without exogenous activation unless indicated otherwise or the test system does not normally use such supplementation; solvent is provided if specified in the report.
(Graham et al., 1973, 1975; NTP, 1992; Ulland et al., 1972). ETU also increased the incidence of liver adenomas and carcinomas in both sexes of mice (Innes et al., 1969; NTP, 1992).
ETU has demonstrated developmental toxicity in exposed rodents. A variety of central nervous system and skeletal effects have been induced by ETU exposure to litters of exposed rats (Chernoff et al., 1979; Khera, 1973, 1987). Reduced pup birth weight, litter size and number of viable litters have been observed after exposure to preg- nant mice (Hardin et al., 1987). ETU has been shown to be a developmental toxicant at doses lower than those that produced no apparent ma- ternal toxicity or fetal lethality (Khera, 1973).
Thyroid effects are caused by exposure to ETU (for review, see Hill et al., 1989). Thyroid toxicity includes thyroid hyperplasia, decreased uptake of radioactive iodine by the thyroid and decreased serum levels of triiodothyronine (T3) and te- traiodothyronine (T4). The ability to inhibit io- dine localization in the thyroid and the thyroid
toxicity seen may contribute to the thyroid car- cinogenic effects induced by ETU (Hill et al., 1989).
ETU has been tested in a wide variety of mutagenicity assays that cover gene mutations, chromosomal alterations and other genotoxic ef- fects. For example, it was included in the Interna- tional Collaborative Program to evaluate short- term tests for carcinogens (de Serres and Ashby, 1981).
The purpose of this review is to examine the available mutagenicity information and data on ETU. The results are tabulated into Table 1. The studies are discussed in the following presenta- tion of the assays.
2. Assay evaluations
(A) Bacterial assays
(1) Salmonel la assay. Many studies have been performed with ETU in the Salmonella assay

120 K.L. Dearfield / Mutation Research 317 (1994) 111-132
with and without metabolic activation. It appears that ETU produces predominantly base substitu- tions as evidenced by positive results in strains TA1535, TA1530, TA1950 and sometimes in strain G46. The EPA's Gene-Tox panel of ex- perts in the Salmonella assay has concluded that ETU is positive in the Salmonella assay based on positive results in strain TA1535 from the Ter- amoto et al. (1977) study (Kier et al., 1986). The addition of metabolic activation does not appear to alter the mutagenic response seen without activation. ETU usually does not induce muta- tions in the frameshift sensitive strains, e.g. TA1531, TA1532, TA1537, TA1538 and TA1964 with the possible exception of an occasional posi- tive in TA98 (e.g. see several studies as part of the International Collaborative Program, report edited by de Serres and Ashby, 1981; Moriya et al., 1983; Schupbach and Hummler, 1977; Ter- amoto et al., 1977).
Seiler (1974) provided an early indication that ETU was active in the Salmonella assay. ETU at moderate concentrations, 100 and 1000 ppm, pro- duced about a 2.5-fold increase in relative muta- genic activity in a spot test with strain his G46 without activation. This positive result in strain G46 was not confirmed by Teramoto et al. (1977) or Schupbach and Hummler (1977) at concentra- tions up to 80 m g / p l a t e in plate tests. However, both of these studies were consistent with a base-substitution mechanism as both found muta- genic activity in other base-substitution strains. Schupbach and Hummler (1977) obtained a con- centrat ion-dependent increase in the number of revertants induced by ETU, but at fairly high concentrations, 20-80 m g / p l a t e in strain TA 1530. Teramoto et al. (1977) also fQund an in- crease in revertant frequency in strain TA1535 from 5000 ixg /p la te to 20000 /zg/p la te . Strain TA100 was not responsive to ETU. While strain TA100 was engineered from TA1535, it is well recognized that a differential response between these two strains is not unusual. Further studies by this same laboratory reenforce this finding (e.g. Shirasu et al., 1977). Autio et al. (1982) also found ETU caused a concentrat ion-dependent increase in revertants in strain 1950 at concentra- tions of 5000 ~ g / p l a t e and above.
Additional studies add to the weight-of-evi- dence that ETU is mutagenic in Salmonella strain TA1535, usually at concentrations around 1000 izg /p la te and above. Simmon and Shepherd (1981), Moriya et al. (1983) and Mortelmans et al. (1986) all found induced responses of 2-4-fold over background. All other strains tested in these laboratories, including TA1537, TA1538, TA98 and TA100, were negative with and without acti- vation. It is of note that ETU has limited solubil- ity in water and negative results with ETU in this assay were found when dissolved at the limit in water and applied to Salmonella plates at con- centrations up to 100 izg /p la te (Mortelmans et al., 1986); ETU is usually dissolved in DMSO for testing (the positive result from Mortelmans et al. was for ETU in DMSO).
ETU was one of the chemicals used in the International Collaborative Program that evalu- ated short-term tests for carcinogens (de Serres and Ashby, 1981). Several laboratories tested ETU in several Salmonella assays as well as other short-term tests. One positive result, Simmon and Shepherd (1981), mentioned above, showed ETU to produce a concentration related and repro- ducible increase in TA1535. Two other studies in this program also found positive results with ETU, but with other strains. Garner et al. (1981) found that ETU did not induce revertants in any tested strains (TA98, TA100, TA1535, TA1537) without or with liver postmitochondrial supernatant acti- vation mix from phenobarbital induced rats. However, when activation mix from 3-methyl- cholanthrene induced rats was used, a response was detected in TA98 at 100 ~ g / p l a t e . Ichino- tsubo et al. (1981b) also found ETU to induce revertants over background in TA98 with activa- tion mix from Aroclor 1254 induction. The Pro- gram report did not address these divergences (Bridges et al., 1981).
The studies found in the International Collab- orative Program report appear to support the suggestion that the overwhelming majority of gene mutation studies in Salmonella were negative. However, upon closer examination of the results with ETU in the laboratories that participated in this program, there are a few caveats that would appear to diminish the impact of much of the

I~L. Dearfield /Mutation Research 317 (1994) 111-132 121
negative studies. There were 15 separate report studies involving the use of Salmonella in the Program report (Chapters 21-35 inclusive; de Serres and Ashby, 1981). Three of the reports indicated positive results for ETU (detailed above). Four reports indicated negative results for ETU; however, three of these studies may not have high enough concentrations to detect the weak genotoxic effect of ETU: Baker and Bonin (1981) tested to 1000/zg/p la te ; Brooks and Dean (1981) and Rowland and Severn (1981) tested to 2000/xg/p la te ; Richold and Jones (1981) did test to 5000 /zg/pla te . Four of the studies in the Program used 3 or fewer strains in their evalua- tions, not including TA1535, which appears to be the strain where ETU most consistently induces mutagenic activity; this restricts the value of the reported negative results for ETU in these stud- ies (Martire et al., 1981; MacDonald, 1981; Na- gao and Takahashi, 1981; Venitt and Crofton- Sleigh, 1981). It should be noted that the Ichinot- subo et al. study mentioned above also did not use strain TA1535. Another study, Trueman (1981) reported that ETU was negative in this Program effort, but ETU was positive in this laboratory when tested on other occasions. Of the remaining three studies, two were not Salmonella plate-incorporation tests but rather fluctuation tests, one using only strains TA98 and TA100 (Hubbard et al., 1981) and one using concentra- tions up to 1000 / zg /ml (Gatehouse, 1981). The final study was a forward mutation assay using Salmonella strain TM677, a derivation of TA1535 which has the pKM101 R factor plasmid (similar to TA100) (Skopek et al., 1981). This forward mutation assay, which measures resistance to 8- azaguanine, found ETU-negative, but at a highest concentration of 100 /zg/ml . So it appears that the evidence is not overwhelmingly negative, but rather that ETU has weak activity in the Salmonella assay, albeit at concentrations usually above 1000 /xg /p la te usually in strain TA1535.
In a test with an automated assay system (Mutascreen®), ETU was found negative at con- centrations up to 1000 / x g / m l in TA1535, TA1537, TA1538, TA98 and TA100 (Falck et al., 1985). This top concentration, again, is at a level where a positive response is usually not seen.
(2) Escherichia coli assays. Several reverse muta- tion assays with different E. coli strains have been performed with ETU. Teramoto et al. (1977) found ETU to be negative when tested in two strains, WP2 hcr ÷ and WP2 hcr - up to 10000 tzg /p la te with and without activation from phe- nobarbital induced rat liver. Moriya et al. (1983) and Shirasu et al. (1977) found similar findings in strain WP2 hcr up to 5000 /xg /p la te with and without activation. With the Mutascreen ®, Falck et al. (1985) found a negative result with WP2 uvrA at concentrations up to 1000 / zg /ml with and without activation.
Several studies were performed with E. coli in the International Collaborative Program (de Ser- res and Ashby, 1981). Venitt and Crofton-Sleigh (1981) found ETU-negative in two strains, WP2(P) (pKM101) up to 100/.~g/plate and WP2 uvrA(P) (pKM101) up to 500 /zg/pla te , both with and without activation. Matsushima et al. (1981) per- formed a preincubation before plating the bacte- ria, but also foundnegat ive results in strains WP2 uvrA ( t rp-) , WP2 uvrA (pKM101) ( t rp - ) and WP2 B / r ( t rp - ) with and without activation; the concentrations used were not specified. Mohn et al. (1981) utilized a liquid suspension assay with strain 3 4 3 / l 1 3 / u v r B at the ga lR- and arg + sys- tems. They obtained positive results with both systems under activated conditions at 0.2-1.0 m g / m l which suggests a base-pair substitution mechanism. Higher concentrations (up to 4.0 m g / m l ) produced values lower than control val- ues. Another study, Gatehouse (1981), utilized a microtiter fluctuation assay with WP2 uvrA with and without activation and found no induced activity up to 1000/zg /ml .
Two additional studies were performed in E. coli in the International Collaborative Program. The preferential ability of ETU to inhibit DNA polymerase I deficient E. coli was determined (Rosenkranz et al., 1981). ETU produced a weak positive result up to 2273 ~ g / m l by preferen- tially inhibiting the pol A~- strain (P3478) over that of the pol A + strain (W3110) without metabolic activation in a liquid suspension assay; this activity was not observed in the presence of activation. The induction of prophage lambda in lysogenic E. coli due to mutagenic action that

122 K.L. Dearfield /Mutation Research 317 (1994) 111-132
induces the "SOS" DNA-repair system was ex- amined by Thomson (1981). ETU induced lambda prophage after bacterial exposure to 1 0 0 0 0 / z g / ml in the presence of metabolic activation.
(3) Host-mediated assays. ETU has been tested in several host-mediated assays, using Salmonella strains inoculated into ETU-exposed rodents. Since ETU appeared to be mutagenic to strain G46 (Seiler, 1974), Teramoto et al. (1977) and Schupbach and Hummler (1977) used this strain in the host-mediated assay. Teramoto et al. (1977) obtained negative results in JCL-SD male rats and JCL-ICR male mice after 3 consecutive doses of 200 or 400 m g / k g given orally at 3-h intervals. Shirasu et al. (1977) found similar results in the ICR male mice at a dose of 150 mg/kg . Schup- bach and Hummler (1977) obtained negative re- suits with G46 in ETU-exposed Swiss albino mice up to 6000 mg/kg . However, at this same dose (which is above their reported LD50 of 5400 mg/kg) , a slight induction of the reversion fre- quency by a factor of 2.37 was noted when Salmonella strain TA1530 was used as the indica- tor organism. The reviewers of the host-mediated assay for the EPA's Gene-Tox program evaluated this result and reported that this was evidence for a positive decision in the host-mediated assay (Legator et al., 1982). Autio et al. (1982) report that they found a negative result in this assay with 5 m g / k g ETU in male NMRI mice. However, this dose was lower than the one where ETU induced a slight increase.
(4) Bacterial rec assays. Bacterial rec assays were performed with Salmonella, Bacillus subtilis and E. coli strains. Strains either proficient or defi- cient in DNA repair are exposed to ETU and if the repair-deficient strain is affected to a greater extent, this suggests that the test compound has DNA-damaging activity. Teramoto et al. (1977) did not find apparent differential activity between B. subtilis strains H17 (rec +) and M45 ( rec - ) after exposure to 4000 ~g /d i sk . Rashid and Mumma (1986) assayed ETU in three bacterial repair assays using Salmonella strains TA1538/ TA1978 (to 2000 /~g/disc), E. coli K-12 strains (polA~-/polA~-) and the E. coli WP2 system
(WP2, WP2 uvrA, WP67, CM611, CM571) (con- centrations not provided for E. coli systems). They obtained negative results in these three repair assays.
Several studies were performed in the Interna- tional Collaborative Program. Kada (1981) used spores of H17 and M45 instead of the usual vegetative cells and found a reproducible weak positive result without activation at 2 mg/disk. Green (1981) and Tweats (1981) both used E. coli WP2 (DNA-repair-proficient) and the repair-de- ficient E. coli strains WP67 uvrA polA and CM871 uvrA recA lexA and obtained negative results with and without activation; exact concen- trations used not specified. Ichinotsubo et al. (1981a) used recombinational deficient E. coli strains JC2921, JC9238, JC8471 and JC5519 and 2 strains with suppressor genes, JC7689 and JC7623, either with or without metabolic activa- tion. ETU induced a positive response in all strains with activation but JC7689 at a concentra- tion of 500 p~g/ml.
(B) Yeast assays Several yeast assays were performed under the
auspices of the International Collaborative Pro- gram which used ETU as a test chemical. In a yeast reversion assay via mutagenic action, Mehta and von Borstel (1981) found a greater than 2-fold increase in reversion frequency in Saccharomyces cerevisiae strain XV185-14C after exposure to 88.9 and 889 l zg /ml in stationary phase cells. Negative results were obtained in the presence of metabolic activation and questionable increases were noted when logarithmic growing cells were exposed without activation at 100 Izg/ml. Using Schizosaccharomyces pombe, Loprieno (1981) did not find an increase in mutation frequency in this forward mutation assay measuring five different genetic loci with and without activation up to 1 /~g/ml.
Other laboratories in the Program examined endpoints such as mitotic gene conversion, mi- totic aneuploidy, mitotic crossing-over and pref- erential inhibition of growth in repair-deficient strains. Three laboratories tested for mitotic gene conversion, but with different yeast strains and obtained different results. Jagannath et al. (1981)

K.L. Dearfield / Mutation Research 317 (1994) 111-132 123
tested ETU in S. cerevisiae strain D4 at the ade2 and trp5 loci with a preincubation method with and without activation to 333.33 t zg /p la te and obtained negative results. Zimmermann and Scheel (1981) also obtained negative results in S. cerevisiae strain D7 trp 5 locus with and without activation at 2 mg /m l . These latter authors sug- gest their testing was performed at insufficient concentrations. Sharp and Parry (1981a) however, using strain JD1 of S. cerevisiae at the his4 po- laron and trp5 locus found a 2-4-fold increase in gene conversion with stationary phase ceils with- out activation with a minimum effective concen- tration of 50 ~ g / m l (activation was not required). Parry and Sharp (1981) also examined for mitotic aneuploidy in S. cerevisiae strain D6. They report that ETU produced aneuploidy at 500 /xg /ml without activation after treatment with either ini- tially stationary or logarithmic phase ceils. How- ever, the report of the EPA's Aneuploidy effort suggests that this result is inconclusive since con- firmation of aneuploidy was not performed as genetic events that mimic the aneuploid pheno- type were not considered (Resnick et al., 1986); nonetheless, there appears to be an effect caused by ETU that may be indicative of an aneuploidy event (Parry, 1986). Mitotic crossing-over with S. cerevisiae strains T1 and T2 was examined by Kassinova et al. (1981) and no increase was found with and without activation to 1000/zg/ml . These same authors also examined the preferential inhi- bition of a repair-deficient strain, T4, versus strain T5, which would indicate DNA damage. No dif- ference between the two strains was found (con- centration not indicated). Sharp and Parry (1981b) also examined the preferential inhibition of growth in the repair-deficient rad strain versus the repair-proficient 197/2d strain. They, on the other hand, obtained positive results with and without activation to 1000/zg /ml .
Schiestl et al. (1989) developed an intrachro- mosomal recombination assay in the yeast S. cerevisiae. ETU induced recombination activity by about 3-fold over solvent control at concentra- tions of 20 and 40 mg /ml ; the increase and high concentrations do not indicate a potent effect. It was also positive by the same fold increase for interchromosomal recombination at 40 mg /ml .
(C) Drosophila assays ETU has been assayed in Drosophila melano-
gaster as the test organism. The first available report was an abstract from Mollet (1975). With a 2.0% concentration of ETU in 5% sugar water, Drosophila mortality was about 50%, but fertility was not reduced and sterility not induced among survivors. There was no induction of sex-linked recessive lethals although the number of lethal tests per dose (up to 2.5% ETU) was too low to be very sensitive, between 1000 and 2000 tests. Dominant lethal effects were increased slightly although the author reports this was not statisti- cally significant. Furthermore, he suggests that ETU affects XY-bearing sperm differently from sperm lacking sex chromosomes.
Valencia and Houtchens (1981; part of Inter- national Collaborative Program) examined ETU in the Drosophila sex-linked recessive lethal as- say. They found negative results after a feeding exposure to 250 ppm and examining a total of 10118 lethal tests. However, these authors sug- gest that this assay was not tested under optimal conditions under the rigid conditions of the Pro- gram. These results have been confirmed by the National Toxicology Program effort. In a pub- lished study (Woodruff et al., 1985), ETU was reported to be equivocal after feeding exposure and negative after injection exposure. However, these results have now been classified as negative by the NTP upon reexamination of these tests at doses up to 5100 ppm (Mason et al., 1992). The NTP (1992) also reports negative results for re- ciprocal translocations in Drosophila after a 500 ppm exposure.
(D) Mammalian cells in culture - - gene mutation assays
The NTP has examined the mutagenicity of ETU at the tk locus of mouse lymphoma cells (McGregor et al., 1988). ETU was negative in the assay without activation at concentrations up to 3600 /zg /ml , the apparent limit of solubility in DMSO. However, under activated conditions, just over a 2-fold increase was noted at around 3000- 3 6 0 0 /x g /m l with relative total growth of > 73% observed. This single weak response presents in- conclusive evidence for ETU genotoxicity in this

124 I£L. Dearfield /Mutation Research 317 (1994) 111-132
assay system. In contrast, Jotz and Mitchell (1981), as part of the International Collaborative Pro- gram, found negative results with and without activation to concentrations of 3000 / z g / m l with a relative total growth of > 71.7%. This top concentration was where the NTP results began to observe suggestive activity.
Carver et al. (1981; part of International Col- laborative Program) tested E T U in the Chinese hamster ovary (CHO) cell line that was heterozy- gous at aprt and tk loci. Other loci examined were hgprt and ATPase loci. The selection agents used were 8-azaadenine, 5-fluorodeoxyuridine, 6-thioguanine and ouabain. There was not an increased mutant frequency at any loci after ex- posure to 2000 / z g / m l with and without activa- tion with relative cell survival at 0.86 and above.
(E) Mammal ian cells in culture - - aberration as- says
Teramoto et al. (1977) exposed the Chinese hamster cell line, Don, to up to 3200 / xg / m l of E T U for 6, 24 or 48 h. The results for aberra- tions, hyperploidy and polyploidy were negative. Shirasu et al. (1977) reports the same results from this same laboratory. The EPA-sponsored Aneu- ploidy Commit tee agrees with the hyperploidy and polyploidy conclusions (Galloway and Ivett, 1986). The NTP reports that E T U was negative for inducing aberrations in the C H O cell line at concentrations up to 10000 / z g / m l (NTP, 1992).
Two reports are found from the International Collaborative Program. Natarajan and van Kes- teren-van Leeuwen (1981) report that E T U does not produce a concentrat ion-dependent increase in aberrations in cultured C H O cells treated for 1 h with and without activation. However, aberra- tions were induced at concentrations of 3.33 and 5.0 m g / m l with and without activation. There was a negative response at 1.67 m g / m l , but large increases at 3.33 were seen, with a larger re- sponse with activation (e.g. 24 b r e a k s / 1 0 0 cells versus 0.3 for control with $9). At 5.0 m g / m l , there was still an elevated increase, but less than that seen at 3.33 m g / m l . Although positive, this drop at the high concentration is the basis for the author 's no concentrat ion-dependent increase re- port. Dean (1981) exposed the rat liver (RL 1) cell
line to ETU. This cell line is reportedly able to possess metabolizing capability, therefore, cul- tures with $9 were not tested. Negative results were tested to 200 ~ g / m l , but this concentration was significantly lower than that used in other studies where activity was observed. The author states that this result is inconclusive as higher concentrations should have been tested, but were not due to constraints to conserve material pro- vided him.
Information from an abstract of an unavailable report (Pilinskaya, 1982) indicates that ETU (1 m g / m l ) incubated with human lymphocytes for 28 h induced about a 2-fold increase in aberra- tions over that found in control lymphocytes.
(F) In vivo cytogenetics Teramoto et al. (1977) and Shirasu et al. (1977)
report on in vivo aberration results from male and female Wistar rats exposed to ETU. Rats of 3 or 10 weeks of age were given either a single oral dose of 200 or 400 m g / k g or 2-5 (50 m g / kg x 5, 100 m g / k g X 5, or 400 m g / k g x 2) con- secutive oral doses at 24-h intervals. Bone mar- row was harvested 24 and 6 h after single and multiple treatments, respectively. These investi- gators did not find an increased aberration fre- quency in the 3 or 10 week old animals; however, they suggest that the frequency of numerically aberrant cells (including aneuploid and polyploid cells) was higher, though not statistically signifi- cant. Sram (1975) reports that E T U increased the frequency of chromosomal aberrations in mouse bone marrow, but no data or concentrations were available for review.
There are several studies that examined for induced micronuclei in exposed animals. Three of these reported studies all exposed mice to 2 treat- ments of ETU, given 24 h apart and then har- vested for bone marrow 6 h after the second dose. All three of these studies reported negative results: in female ICR mice to 150 m g / k g / d a y (Seiler, 1975), in male and female Swiss albino mice to 6000 m g / k g / d a y (Schupbach and Hummler , 1977) and male and female CD-1 mice to 880 m g / k g / d a y (Tsuchimoto and Matter, 1981; part of International Collaborative Pro- gram). The EPA sponsored Gene-Tox program,

K~L. Dearfield /Mutation Research 317 (1994) 111-132 125
in the review of the micronucleus test suggests this sampling time is inappropriate (Heddle et al., 1983). They state the reason is that enucleation of the erythrocyte takes place about 6 -8 h after the final mitosis; this would not allow an opportunity for micronucleus formation in the PCE. As many chemicals also induce cell-cycle delay, this 6-h interval may be especially inappropriate. There- fore, it is recommended that sampling take place no earlier than 12 h after the second treatment. These three studies then may not be examining for micronuclei in an appropriate manner.
Two studies from the International Collabora- tive Program examined for micronuclei with more appropriate sampling times. Salamone et al. (1981) exposed B6C3F1 hybrid mice (sex not stated) to ETU under 2 protocols. Mice were given two i.p. doses of ETU 24 h apart and then sampled at 48 and 72 h for the 1416 m g / k g / d a y doses and at 24, 72 and 96 h for the 880 m g / k g / day doses. The second protocol involved a single i.p. t reatment with 1328 m g / k g and sampling at 48 h. The 1416 m g / k g dose was reported to represent 80% of the LD50. Under both proto- cols, the authors concluded the results were nega- tive. A slight increase was seen at the top dose at 48 h in the first protocol, but was not seen in a confirmatory test. Kirkhart (1981) dosed male ICR mice with up to 880 m g / k g one time and then sampled at 30 and 48 h. Negative results were also reported. However, in both of these reports, there was uncertainty as evaluated by the Gene-Tox panel of experts, as these two studies did not examine the recommended minimum number of samples (PCE) for determining an actual effect (Heddle et al., 1983). Also, it was not clear if animals from both sexes were used.
(G) Dominant lethal assays Schupbach and Hummler (1977) exposed Swiss
albino male mice to an oral dose of 500, 1000 or 3500 m g / k g and then mated them to 3 females / m a l e / w e e k for 10 weeks. There were statistically significant increases in number of dead implants per female at several doses in the first 5 weeks after exposure (not found at every week at every dose, so the authors suggest that without consis- tent response, these results are probably nega-
tive). The Gene-Tox panel of experts for the dominant lethal assay assessed these results as positive (Green et al., 1985); a consistent dose response is not necessary for a positive response in this assay.
Teramoto et al. (1977) and Shirasu et al. (1977) report that JCL-ICR mice were given oral doses of 300 or 600 mg E T U / k g / d a y for 5 consecutive days and then mated for 5 weeks with 1 female per mating period (2-4 per iod /week) . There was a negative dominant lethal effect, but no appar- ent effect on fertility or toxicity as these doses were at the low end of the dose range used in the Schupbach and Hummler (1977) study that ob- tained positive results. Teramoto et al. (1978), in another dominant lethal study using C 3 H / H e C r mice exposed to 150 m g / k g for 5 days, obtained similar results to their earlier study (Teramoto et al., 1977). Sram (1975) reports that ETU did not increase the frequency of dominant lethal effects after a single or repeated administration in mice. No details or data from this report were available for review.
(H) Unscheduled DNA synthesis (UDS) assays Three reports from the International Collabo-
rative Program examined UDS after ETU expo- sure. All three utilized human cultured cells and scintillation counting after DNA extraction to quantitate the level of UDS. After a hydroxyurea pretreatment, Robinson and Mitchell (1981) ex- posed WI-38 human fibroblasts to ETU for 3 h without activation (63-1000 i zg /ml ) and for 4 h with activation (125-2000 tzg/ml) . They report negative findings, but suggest that ETU was not able to be tested at concentrations capable of inducing cytotoxic effects. Martin and McDermid (1981) exposed HeLa $3 cells, pretreated with hydroxyurea, with and without activation for 2.5 h. They report positive results without activation, negative with activation, but no concentrations or data were provided. Agrelo and Amos (1981) also provided no ETU concentrations for their UDS assay that examined human fibroblasts from skin biopsies, pretreated with hydroxyurea and ex- posed to ETU for 1 h without activation only. Negative results were reported.

126 K.L. Dearfield /Mutation Research 317 (1994) 111-132
Althaus et al. (1982) performed a rat hepato- cyte DNA-repair assay using a nuclei isolation procedure for determination of DNA-repair syn- thesis. Cultures were treated with ETU (3.2 × 10 -3 M to 9.6 × 10 -9 M) for 18 h. Results ob- tained were negative after liquid scintillation counting of isolated nuclei.
Frank and Muller (1988) obtained negative results for UDS after an in vivo exposure of 1500 m g / k g ETU in corn oil to female B6C3F1 mice and in vitro culture of hepatocytes. On the other hand, an induction of the number of S-phase cells was found, suggesting an effect on proliferation.
(I) S&ter-chromatid exchange (SCE) assays There are several reports that examined for
SCE in cultured Chinese hamster ovary (CHO) cells. The NTP (1992) reports they obtained neg- ative results for SCE in CHO cells at concentra- tions up to 10000 /zg /ml . Three studies from the International Collaborative Program also report negative results for SCE in CHO ceils with and without activation. Evans and Mitchell (1981) ex- posed CHO cells for 21.5 h without activation and 2 h with activation with a total culture time of 24 h. Concentrations used were 2 5 - 4 0 0 / x g / m l without activation and 62.5-1000 / zg /ml with activation. These authors suggest that dosing may not have been high enough and no signs of toxic- ity were reported. Perry and Thomson (1981) exposed CHO cells to ETU for 1 h with and without activation up to 1 0 0 / z g / m l and obtained negative results. However, these authors also suggested that the restriction in concentration range may fail to identify activity as no toxicity was reported. Natarajan and van Kesteren-van Leeuwen (1981) also obtained negative results after CHO cells were exposed to up to 5.0 m g / m l for 1 h with and without activation (it should be noted that this study saw induced aberrations, see above).
Paika et al. (1981, part of International Collab- orative Program) examined for SCE in male C B A / J mice after an in vivo exposure to ETU. Bone marrow and liver were obtained after a partial hepatectomy and bone marrow was also obtained without a partial hepatectomy per-
formed. Negative results were found under all conditions at doses up to 1000 mg/kg .
(J) Cell transformation assays There were two submitted studies to the
USEPA's Office of Pesticide Programs (OPP) examining for cell transformation effects in C 3 H / 1 0 T 1 / 2 ceils (McGlynn-Kreft and Mc- Carthy, 1984; McLeod and Doolittle, 1985). The transformation frequency was not increased for Type III foci in these cells at concentrations up to 1000 # g / m l . These results were considered ac- ceptable. The second study reported to examine for promotion effects where C 3 H / 1 0 T 1 / 2 cells were pretreated with acetone or MNNG and then exposed to 3 3 3 / x g / m l of ETU. The results were negative, but considered unacceptable as more concentrations are required as promoters fre- quently induce erratic and non-dose related ef- fects.
Hatch et al. (1986) reported a slight enhance- ment of adenovirus SA7 transformation of ham- ster embryo cells at a top concentration of 12 mM, but this result was not reproducible in a second laboratory. In this interlaboratory study, therefore, they considered these results for ETU-negative. There were two transformation studies reported in the International Collabora- tive Program (Daniel and Dehnel, 1981; Styles, 1981) that used the BHK21 cell transformation assay. Both studies found positive results for in- ducing ability of BHK21 cells to form colonies in soft agar medium. However, it is known that this assay is not performed much anymore.
(K) Germ cell assays Seiler (1977a) found that ETU at a 100 m g / k g
i.p. dose did not inhibit testicular DNA synthesis in male mice as detected by uptake of tritiated thymidine. Two studies from the International Collaborative Program assayed ETU for sperm- head abnormalities. Wyrobek et al. (1981) dosed B 6 C 3 F 1 / C R L mice once a day i.p. for 5 consec- utive days and then sacrificed animals 35 days after the first dose. Sperm were examined and no effects on morphology were seen after dosing up to 2655 m g / k g / d a y . In a similar experiment, Topham (1981) examined ( C B A × BALB/c)F1

K.L. Dearfield / Mutation Research 317 (1994) 111-132 127
mice at doses up to 2000 m g / k g / d a y and found negative results.
(L) Other genotoxicity assays Other genotoxicity assays have been per-
formed with ETU. Crebelli et al. (1986) looked for forward mutations (methionine suppressors), mitotic crossing-over and chromosome malsegre- gation in Aspergillus nidulans. They found that E T U induced the frequency of mitotic segrega- tion at concentrations to 58.7 mM without appar- ent effect on gene mutations and mitotic cross- ing-over. The authors suggest that E T U distur- bance of the mitotic segregation of chromosomes may occur via interaction with non-DNA targets. The SOS Chromotest did not find an induction of the gene sfiA as a consequence of induced D N A damage in the SOS system in E. coli (Quillardet et al., 1985). van ' t Hof and Schairer (1982) found that ETU was effective at a minimum concentra- tion of 9.79 x 10 -s M after a 24-h exposure was able to produce mutations in the plant Trades- cantia, clone 4430. This assay involves detecting a phenotypic change in flower color from blue to pink as detected as sectors in the petals or cells in the stamen hairs.
(M) ETU nitrosation products There are suggestions that ureas can be ni-
trosated with sodium nitrite at appropriate pH's. As many ethylene bisdi thiocarbamate-treated products are cooked and eaten, E T U as a metabolite and a contaminant can be exposed to humans under conditions where nitrosation in the stomach may occur. The following reports exam- ine the potential mutagenic activity by such nitro- sation products.
In two studies, Seiler (1975, 1977b) examined the effect a combination of ETU and sodium nitrite would have on inducing Salmonella muta- tion, micronuclei in mice and aberrations in Chi- nese hamsters. In Salmonella strain G46, E T U by itself was found to be a weak mutagen, but in combination with sodium nitrite (1 mM E T U + 5 mM sodium nitrite), very large increases in mu- tant frequency were obtained. In the micronu- cleus assay, performed with sampling times that
may be considered inappropriate to detect weak effects (see above discussion, section F), E T U itself was negative. However, E T U + sodium ni- trite given p.o. induced very large increases in micronuclei. It was presumed that nitrosation oc- curred in the stomach, yet genotoxic reaction products were able to reach the bone marrow. E T U (300 m g / k g ) + sodium nitrite (100 m g / k g ) given p.o. also induced large increases in chromo- somal aberrations in Chinese hamster bone mar- row. An exposure to N-ni t roso-ETU (the ni- trosated derivative of ETU) at 100 m g / k g i.p. also induced a large increase in Chinese hamster bone marrow aberrations.
Shirasu et al. (1977) found that under acidic conditions ETU + sodium nitrite added together induced large increases in mutant frequency in Salmonella strains TA1535 (160-fold over back- ground) and TA100 (15-fold) and in E. coli strain WP2 hc r - (30-fold). Also, in a host-mediated assay with Salmonella strain G46 as indicator organism, male ICR mice were exposed p.o. to E T U (150 m g / k g ) + sodium nitrite (50 m g / k g ) or N-ni t roso-ETU (125 m g / k g ) . Large increases in mutant frequency were seen where negative results were seen with E T U alone. A dose re- sponse was found if the concentration of sodium nitrite was fixed (50 m g / k g ) and E T U doses were increased (1-25 m g / k g ) .
The interaction of E T U and sodium nitrite was examined in the dominant lethal assay (Teramoto et al., 1978). Male C 3 H / H e C r mice were given test article p.o. once a day for 5 days and then mated for 6 weeks. E T U (300 m g / k g ) + sodium nitrite (100 m g / k g ) killed half the ani- mals; N-ni t roso-ETU at 400 m g / k g was lethal. There was no apparent effect with ETU alone, but E T U (150 m g / k g ) + sodium nitrite (50 m g / kg) reduced the percentage pregnancy, number of implants and number of live embryos at weeks 5 a n d / o r 6. Post-implantation losses were not ob- served. Similar results were seen with 100 m g / k g N-nitroso-ETU. The authors suggest that interac- tion of ETU and sodium nitrite is capable of adverse effects in mouse germ cells at pre-meiotic stages.
E T U is metabolized in mice by oxidation of the sulfur atom to 2-imidazolin-2-yl sulfenate.

128 K.L. Dearfield / Mutation Research 317 (1994) 111-132
Autio et al. (1982) tested ETU, this oxidation product and their nitrosated derivatives in the Salmonella assay and the host-mediated assay. E TU by itself was weakly positive and the sulfe- hate was negative in the plate-incorporation as- say. However, both of their nitrosated derivatives induced large increases in the Salmonella assay strain TA1950 ( > 5000 revertants per plate) with the ETU derivative more active. In the host- mediated assay with NMRI mice and Salmonella strain TA1950, negative results were obtained with the parent compounds. The nitrosated derivatives produced positive results with the ETU derivative again more active.
It appears from this body of evidence that the nitrosated form of ETU is a mutagenic product and presents a mutagenicity concern.
3. Other considerations and conclusion
Although the available studies on ethylene thiourea (ETU) show many negative results, there is evidence to suggest that ETU has weak geno- toxic activity. This is based on responses that are found in several different assay systems (e.g. strain TA1535 in the Salmonella assay, aberrations in cultured mammalian cells, a dominant lethal as- say, bacterial rec assay and several yeast assays). The responses are not considered potent and are usually found at higher concentra t ions /doses in the assay systems. Because of the weak activity and potency of ETU, many negative studies may not have been performed with high enough con- centrations to adequately test ETU. Higher con- centrations in some of these studies might reveal ETU to be weakly positive with additional test- ing.
This conclusion is consistent with the final position of the U.S. Environmental Protection Agency's (USEPA's) Office of Pesticide Programs (OPP) in its conclusion of the special review of EBDCs (USEPA, 1992). This special review ex- amined the toxicity profile of ETU as well as the EBDCs. These two conclusions are similar be- cause they are derived from the same review effort.
Other groups have made inferences about the ETU genotoxicity data base and while on the surface appear conflicting, they generally do not diametrically disagree with the basic assumption that ETU has some genotoxic activity. During the OPP consideration of EBDCs and ETU, the OPP's Scientific Advisory Panel (SAP; a panel of external peer reviewers) opined " that the evi- dence for genotoxicity was equivocal both in prokaryotic and eukaryotic test systems" (USEPA, 1992). This opinion was delivered in the context of the carcinogenicity consideration of ETU and was part of the overall evidence for cancer induc- tion. Since the SAP was not examining the geno- toxicity information in depth, this opinion recog- nizes the occurrence of positive and negative results in the data base by their equivocal conclu- sion. This suggests that this is not a definitive decision and one at least cannot conclude that ETU does not have any genotoxicity activity at all.
The United Kingdom's Ministry of Agricul- ture, Fisheries and Food (MAFF) also examined some of the genotoxicity information on EBDCs and ETU (MAFF, 1990). They concluded that " the mutagenicity s tudies . . . show no evidence that E T U . . . is genotoxic in vivo." Although there are many genotoxicity tests performed with ETU, in actuality, there are very few available in vivo studies in which there were not problems (dis- cussed in assay sections above). Also, one of the dominant lethal studies was judged to have a positive effect (Green et al., 1985). This suggests at the very least that there may be activity for dominant lethal activity. Due to the paucity of well conducted in vivo studies and the suggestive dominant lethal result, it is clear that additional in vivo studies are warranted before a definitive conclusion can be reached on the in vivo activity of ETU.
The International Agency for Research on Cancer (IARC) presented genotoxicity data on ETU (IARC, 1987) in the genetic activity profile methodology as developed by Waters and co- workers (e.g. Garret t et al., 1986). The ETU profile also shows the mixed results of the ETU data base with negative and positive results. In their deliberations on the ETU genotoxicity data

K.L. Dearfield / Mutation Research 317 (1994) 111-132 129
base, I A R C conc luded that the evidence for ac- tivity in shor t - te rm tests (i.e. mutagenici ty) is lim- i ted ( IARC, 1982). Limited is def ined a s . . . " w h e n there were at least two positive results, e i ther for different endpo in t s or in systems represen t ing two levels of biological complexity." So it again suggests f rom these de l ibera t ions that E T U is capable of some genotoxic activity.
The Nat ional Toxicology Program also dis- cussed the mutagenic i ty data in a summary form in the Technica l Repor t for the carcinogenici ty studies on E T U (NTP, 1992). The Technica l Re- port also recognizes the many negative studies with some positive results. The test ing pe r fo rmed u n d e r the auspices of the NTP and repor ted in the Technica l Repor t show positive results in the Sa lmonel la assay that are consis tent with other reports. Also, inconclusive activity is seen with the mouse lymphoma assay and negat ive results for Drosophi la tests and aber ra t ions and SCE in C H O cells. Dur ing the del ibera t ions that led to the carcinogenici ty conclusions found in the Technica l Repor t , the genotoxicity evidence was discussed. In the summary minu te s of the NTP ' s Board of Scientific Counse lors ' peer review on drafts of the Technica l Repor t (November 20-21, 1989), the op in ion was p resen ted that " the evi- dence suppor ted E T U as be ing mutagen ic . " This is not inconsis tent with the overall conclusion
p resen ted in this review. It is clear from these de l ibera t ions and this
review that E T U exhibits weak genotoxic poten- tial. At the very least, it canno t be conc luded that E T U is devoid of any genotoxic activity. In terms of how this may impact on her i table risk and the induc t ion of tumors, such as E T U - i n d u c e d thy- roid tumors, is unclear . These are areas of future research that may shed insight on the potent ia l role of E T U in these processes.
4. Acknowledgement
The au thor acknowledges the helpful com- ments and review of this manuscr ip t by Dr. A lb in Kocialski, Office of Pesticide Programs, U.S. En- v i ronmenta l Protec t ion Agency.
5. References
Agrelo, C., and H. Amos (1981) DNA repair in human fibro- blasts, Prog. Mutation Res., 1,528-532.
Althaus, F., S. Lawrence, G. Sattler, D. Longfellow and H. Pitot (1982) Chemical quantification of unscheduled DNA synthesis in cultured hepatocytes as an assay for the rapid screening of potential chemical carcinogens, Cancer Res., 42, 3010-3015.
Autio, K., A. von Wright and H. Pyysalo (1982) The effect of oxidation of the sulfur atom on the mutagenicity of eth- ylenethiourea, Mutation Res., 106, 27-31.
Baker, R., and A. Bonin (1981) Study of 42 coded compounds with the Salmonella/mammalian microsome assay, Prog. Mutation Res., 1,247-260.
Bridges, B., D. MacGregor and E. Zeiger (1981) Summary report on the performance of bacterial mutation assays, Prog. Mutation Res., 1, 49-67.
Brooks, T., and B. Dean (1981) Mutagenic activity of 42 coded compounds in the Salmonella/microsome assay with preincubation, Prog. Mutation Res., 1,261-270.
Carver, J., E. Salazar, M. Knize and D. Wandres (1981) Mutation induction at multiple gene loci in Chinese ham- ster ovary cells: the genetic activity of 15 coded carcino- gens and noncarcinogens, Prog. Mutation Res., 1,594-601.
Chernoff, N., R. Kavlock, E. Rogers, B. Carver and S. Murray (1979) Perinatal toxicity of maneb, ethylene thiourea, and ethylene-bisisothiocyanate sulfide in rodents, J. Toxicol. Environ. Health, 5, 821-834.
Crebelli, R., D. Bellincamp, G. Conti, L. Conti, G. Morpurgo and A. Carere (1986) A comparative study on the selected chemical carcinogens for chromosome malsegregation, mi- totic crossing-over and forward mutation induction in As- pergillus nidulans, Mutation Res., 172, 139-150.
Daniel, M., and J. Dehnel (1981) Cell transformation test with baby hamster kidney cells, Prog. Mutation Res., 1,626-637.
Dean, B. (1981) Activity of 27 coded compounds in the RL 1 chromosome assay, Prog. Mutation Res., 1,570-579.
de Serres, F., and J. Ashby (Eds.) (1981) Evaluation of Short- Term Tests for Carcinogens, Report of the International Collaborative Program, Progress in Mutation Research, Vol. 1, Elsevier, New York.
Engst, R., and W. Schnaak (1974) Residues of dithiocarba- mate fungicides and their metabolites on plant foods, Residue Rev., 52, 45-67.
Evans, E., and A. Mitchell (1981) Effects of 20 coded chemi- cals on sister-chromatid exchange frequencies in cultured Chinese hamster cells, Prog. Mutation Res., 1,538-550.
Falck, K., P. Partanen, M. Sorsa, O. Suovaniemi and H. Vainio (1985) Mutascreen ®, an automated bacterial muta- genicity assay, Mutation Res., 150, 119-125.
Frank, J., and G. Muller (1988) ETU: in vivo/in vitro UDS/S-phase assay in mice, Report No. 88R-47, Unpub- lished study by Rohm and Haas submitted to the U.S. Environmental Protection Agency, Office of Pesticide Pro- grams, Washington, DC, MRID No. 42908001.
Galloway, S., and J. Ivett (1986) Chemically induced aneu-

130 K.L. Dearfield /Mutation Research 317 (1994) 111-132
ploidy in mammalian cells in culture, Mutation Res., 167, 89-105.
Garner, R.D., A. Welch and C. Pickering (1981) Mutagenic activity of 43 coded compounds in the Salmonella/ microsome assay, Prog. Mutation Res., 1,280-284.
Garrett, N., H.F. Stack and M. Waters (1986) Evaluation of the genetic activity profiles of 65 pesticides, Mutation Res., 168, 301-325.
Gatehouse, D. (1981) Mutagenic activity of 42 coded com- pounds in the "microtiter" fluctuation test, Prog. Muta- tion Res., 1,376-386.
Graham, S., W. Hansen, K. Davis and C. Perry (1973) Effects of one-year administration of ethylenethiourea upon the thyroid of the rat, J. Agric. Food Chem., 21,324-329.
Graham, S., K. Davis, W. Hansen and C. Graham (1975) Effects of prolonged ETU ingestion on the thyroid of the rat, Food Cosmet. Toxicol., 13, 493-499.
Green, M.H.L. (1981) A differential killing test using an improved repair-deticient strains of Escherichia coli, Prog. Mutation Res., 1, 183-194.
Green, S., A. Auletta, J. Fabricant, R. Kapp, M. Manandhar, C. Sheu, J. Springer and B. Whitfield (1985) Current status of bioassays in genetic toxicology - - the dominant lethal assay. A report of the U.S. Environmental Protec- tion Agency Gene-Tox Program, Mutation Res., 154, 49- 67.
Hardin, B., R. Schuler, J. Burg, G. Booth, K. Hazelden, K. MacKenzie, V. Piccirillo and K. Smith (1987) Evaluation of 60 chemicals in a preliminary developmental toxicity test, Teratogen. Carcinogen.Mutagen., 7, 29-48.
Hatch, G., T. Anderson, R. Lubet, R. Houri, D. Putman, J. Cameron, R. Nims, B. Most, J. Spalding, R. Tennant and L. Schechtman (1986) Chemical enhancement of SA7 virus transformation of hamster embryo cells: evaluation by interlaboratory testing of diverse chemicals, Environ. Mu- tagen., 8, 515-531.
Heddle, J., M. Hite, B. Kirkhart, K. Mavournin, J. MacGre- gor, G. Newell and M. Salamone (1983) The induction of micronuclei as a measure of genotoxicity. A report of the U.S. Environmental Protection Agency Gene-Tox Pro- gram, Mutation Res., 123, 61-118.
Heidelberger, C., A. Freeman, R. Pienta, A. Sivak, J. Bertram, B. Casto, V. Dunkel, M. Francis, T. Kakunaga, J. Little and L. Schechtman (1983) Cell transformation by chemical agents - - a review and analysis of the literature, A report of the U.S. Environmental Protection Agency Gene-Tox Program, Mutation Res., 114, 283-385.
Hill, R., L. Erdreich, O. Paynter, P. Roberts, S. Rosenthal and C. Wilkinson (1989) Thyroid follicular cell carcinogen- esis, Fund. Appl. Toxicol., 12, 629-697.
Hubbard, S., M.H.L. Green, B. Bridges, A. Wain and J. Bridges (1981) Fluctuation test with $9 and hepatocyte activation, Prog. Mutation Res., 1,361-370.
IARC (1982) IARC Monographs on the Evaluation of the Carcinogenic Risk of Chemicals to Humans; Chemicals, Industrial Processes and Industries Associated with Can- cer in Humans, IARC Monographs, Vols. 1-29, Suppl. 4,
International Agency for Research on Cancer, Lyon, France.
IARC (1987) IARC Monographs on the Evaluation of Car- cinogenic Risks to Humans: Genetic and Related Effects: an Update of Selected IARC Monographs from Vols. 1-42, Suppl. 6, International Agency for Research on Cancer, Lyon, France.
Ichinotsubo, D., H. Mower and M. Mandel (1981a) Testing of a series of paired compounds (carcinogen and noncarcino- genic structural analog) by DNA repair-deficient E. coli strains, Prog. Mutation Res., 1, 195-198.
Ichinotsubo, D., H. Mower and M. Mandel (1981b) Mutagen testing of a series of paired compounds with the Ames Salmonella testing system, Prog. Mutation Res., 1, 298- 301.
Innes, J., B. Ulland, M. Valerio, L. Petrucelli, L. Fishbein, E. Hart, A. Pallotta, R. Bates, H. Falk, J. Gart, M. Klein, I. Mitchell and J. Peters (1969) Bioassay of pesticides and industrial chemicals for tumorigenicity in mice: a prelimi- nary note, J. Natl. Cancer Inst., 42, 1101-1114.
Jagannath, D., D. Vultaggio and D. Brusick (1981) Genetic activity of 42 coded compounds in the mitotic gene conver- sion assay using Saccharomyces cerevisiae strain D4, Prog. Mutation Res., 1,456-467.
Jordan, L., and R. Neal (1979) Examination of the in vivo metabolism of maneb and zineb to ethylenethiourea (ETU) in mice, Bull. Environ. Contam. Toxicol., 22, 271-277.
Jotz, M.M., and A. Mitchell (1981) Effects of 20 coded chemi- cals on the forward mutation frequency at the thymidine kinase locus in L5178Y mouse lymphoma cells, Prog. Mu- tation Res., 1, 580-593.
Kada, T. (1981) The DNA-damaging activity of 42 coded compounds in the rec-assay, Prog. Mutation Res., 1, 175- 182.
Kassinova, G., S. Kovaltsova, S. Martin and I. Zakharov (1981) Activity of 40 coded compounds in differential inhibition and mitotic crossing-over assays in yeast, Prog. Mutation Res., 1,434-455.
Khera, K. (1973) Teratogenic effects of ethylenethiourea in rats and rabbits, Toxicol. Appl. Pharmacol., 25, 455-456.
Khera, K. (1987) Ethylenethiourea: a review of teratogenicity and distribution studies and an assessment of reproduction risk, CRC Crit. Rev. Toxicol., 18, 129-139.
Kier, L., D. Brusick, A. Auletta, E. von Halle, M. Brown, V. Simmon, V. Dunkel, J. McCann, K. Mortelmans, M. Pri- val, T. Rao and V. Ray (1986) The Salmonella ty- phimurium/mammalian microsomal assay, A report of the U.S. Environmental Protection Agency Gene-Tox Pro- gram, Mutation Res., 168, 69-240.
Kirkhart, B. (1981) Micronucleus test on 21 compounds, Prog. Mutation Res., 1, 698-704.
Legator, M., E. Beuding, R. Batzinger, T. Connor, E. Eisen- stadt, M. Farrow, G. Ficsor, A. Hsie, J. Seed and R. Stafford (1982) An evaluation of the host-mediated assay and body fluid analysis. A report of the U.S. Environmen- tal Protection Agency Gene-Tox Program, Mutation Res., 98, 319-374.

K.L. Dearfield /Mutation Research 317 (1994) 111-132 131
Loprieno, N. (1981) Screening of coded carcinogenic/ noncarcinogenic chemicals by a forward-mutation system with the yeast Schizosaccharomyces pombe, Prog. Mutation Res., 1,424-433.
MacDonald, D. (1981) Salmonella/microsome tests on 42 coded chemicals, Prog. Mutation Res., 1,285-297.
MAFF (1990) Ministry of Agriculture, Fisheries, and Food, Position document on consumer risk arising from the use of ethylene bisdithiocarbamates, MAFF, London, United Kingdom, January 31, 1990.
Marshall, W. (1977) Thermal decomposition of ethylenebis- dithio-carbamate fungicides to ethylenethiourea in aque- ous media, J. Agric. Food Chem., 25, 357-361.
Martin, C., and A. McDermid (1981) Testing of 42 coded compounds for their ability to induce unscheduled DNA repair synthesis in HeLa cells, Prog. Mutation Res., 1, 533-537.
Martire, G., G. Vricella, A.M. Perfumo and F. DeLorenzo (1981) Evaluation of the mutagenic activity of coded com- pounds in the Salmonella test, Prog. Mutation Res., 1, 271-279.
Mason, J., R. Valencia and S. Zimmering (1992) Chemical mutagenesis testing in Drosophila, VIII. Reexamination of equivocal results, Environ. Mol. Mutagen., 19, 227-234.
Matsushima, T., Y. Takamoto, A. Shirai, M. Sawamura and T. Sugimura (1981) Reverse mutation test of 42 coded com- pounds with the E. coli WP2 system, Prog. Mutation Res., 1,387-395.
McGlynn-Kreft, A., and K. McCarthy (1984) Ethylenethiourea mammalian cell transformation test, Report No. 84R 0056, Unpublished study by Rohm and Haas Co. submitted to the U.S. Environmental Protection Agency, Office of Pes- ticide Programs, Washington, DC, MRID No. 00148231.
McGregor, D., A. Brown, P. Cattanach, I. Edwards, D. McBride, C. Riach and W. Caspary (1988) Responses of the L5178Y t k + / t k - mouse lymphoma cell forward muta- tion assay, III. 72 coded chemicals, Environ. Mol. Muta- gen., 12, 85-154.
McLeod, P., and D. Doolittle (1985) Ethylenethiourea mam- malian cell transformation test for promotion, Report No. 84R-298, Unpublished study by Rohm and Haas Co. sub- mitted to the U.S. Environmental Protection Agency, Of- fice of Pesticide Programs, Washington, DC, MRID No. 00148241.
Mehta, R., and R. von Borstel (1981) Mutagenic activity of 42 encoded compounds in the haploid yeast reversion assay, strain XV185-14C, Prog. Mutation Res., 1,414-423.
Mohn, G., S. Vogels-Bouter and J. van der Horst-van der Zon (1981) Studies on the mutagenic activity of 20 coded compounds in liquid tests using the multipurpose strain Escherichia coli K-12/343/113 and derivatives, Prog. Mu- tation Res., 1,396-413.
Mollet, P. (1975) Toxicity and mutagenicity of ethylene- thiourea (ETU) in Drosophila, Mutation Res., 29, 254.
Moriya, M., T. Ohta, K. Watanabe, T. Miyazawa, K. Kato and Y. Shirasu (1983) Further mutagenicity studies on pesti-
cides in bacterial reversion assay systems, Mutation Res., 116, 185-216.
Mortelmans, K., S. Haworth, T. Lawlor, W. Speck, B. Tainer and E. Zeiger (1986) Salmonella mutagenicity tests, II. Results from the testing of 270 chemicals, Environ. Muta- gen., 8 (Suppl. 7), 1-119.
Nagao, M., and Y. Takahashi (1981) Mutagenic activity of 42 coded compounds in the Salmonella/microsome assay, Prog. Mutation Res., 1, 302-313.
Natarajan, A., and A. van Kesteren-van Leeuwen (1981) Mu- tagenic activity of 20 coded compounds in chromosome aberrations/sister-chromatid exchanges assay using Chi- nese hamster ovary (CHO) cells, Prog. Mutation Res., 1, 551-559.
NTP (1992) National Toxicology Program, Toxicology and carcinogenesis studies of ethylene thiourea in F344/N rats and B6C3F1 mice (Feed Studies), U.S. Department of Health and Human Services, Public Health Service, Na- tional Institutes of Health, NIH Publication No. 92-2843, National Toxicology Program Technical Report Series No. 388, March 1992.
Paika, I., M. Beauchesne, M. Randall, R. Schreck and S. Latt (1981) In vivo SCE analysis of 20 coded compounds, Prog. Mutation Res., 1,673-681.
Parry, J. (1986) The expression of recessive markers in pre- sumptive monosomic colonies derived from Saccha- romyces cerevisiae, strain D6, Mutagenesis, 1,299-300.
Parry, J., and D. Sharp (1981) Induction of mitotic aneuploidy in the yeast strain D6 by 42 coded compounds, Prog. Mutation Res., 1,468-480.
Perry, P., and E. Thomson (1981) Evaluation of the sister chromatid exchange method in mammalian cells as a screening system for carcinogens, Prog. Mutation Res., 1, 560-569.
Pilinskaya, M. (1982) Analysis of the cytogenetic activity of ethylenethiourea (ETU) in human peripheral lymphoblast culture in vitro, Dopov. Akad. Nauk. Ukr., RSR, Ser. B: Geol., Khim. Biol. Nauki., 10, 66-68.
Quillardet, P., C. de Bellecombe and M. Hofnung (1985) The SOS Chromotest, a colorimetric bacterial assay for geno- toxins: validation study with 83 compounds, Mutation Res., 147, 79-95.
Rashid, K., and R. Mumma (1986) Screening pesticides for their ability to damage bacterial DNA, J. Environ. Sci. Health Part B Pestic. Food Contam. Agric. Wastes, 21, 319-334.
Resnick, M., V. Mayer and F. Zimmermann (1981) The detection of chemically induced aneuploidy in Saccha- romyces cerevisiae: an assessment of mitotic and meiotic systems, Mutation Res., 167, 47-60.
Richold, M., and E. Jones (1981) Mutagenic activity of 42 coded compounds in the Salmonella/microsome assay, Prog. Mutation Res., 1,314-322.
Robinson, D., and A. Mitchell (1981) Unscheduled DNA synthesis response of human fibroblasts, WI-38 cells, to 20 coded chemicals, Prog. Mutation Res., 1,517-527.

132 K. L. Dearfield / Mutation Research 317 (1994) 111-132
Rosenkranz, H., J. Hyman and Z. Leifer (1981) DNA poly- merase deficient assay, Prog. Mutation Res., 1,210-218.
Rowland, I., and B. Severn (1981) Mutagenicity of carcino- gens and noncarcinogens in the Salmonella/microsome test, Prog. Mutation Res., 1,323-332.
Salamone, M., J. Heddle and M. Katz (1981) Mutagenic activity of 41 compounds in the in vivo micronucleus assay, Prog. Mutation Res., 1,686-697.
Schiestl, R., R.D. Gietz, R. Mehta and P. Hastings (1989) Carcinogens induce intrachromosomal recombination in yeast, Carcinogenesis, 10, 1445-1455.
Schupbach, M., and H. Hummler (1977) A comparative study on the mutagenicity of ethylenethiourea in bacterial and mammalian test systems, Mutation Res., 56, 111-120.
Seiler, J. (1974) Ethylenethiourea (ETU), a carcinogenic and mutagenic metabolite of ethylenebis-dithiocarbamate, Mu- tation Res., 26, 189-191.
Seiler, J. (1975) In vivo mutagenic interaction of nitrite and ethylenethiourea, Experientia, 31, 214-215.
Seiler, J. (1977a) Inhibition of testicular DNA synthesis by chemical mutagens and carcinogens, Preliminary results in the validation of a novel short term test, Mutation Res., 46, 305-310.
Seiler, J. (1977b) Nitrosation in vitro and in vivo by sodium nitrite, and mutagenicity of nitrogenous pesticides, Muta- tion Res., 48, 225-236.
Sharp, D., and J. Parry (1981a) Induction of mitotic gene conversion by 41 coded compounds using the yeast culture JD1, Prog. Mutation Res., 1,491-501.
Sharp, D., and J. Parry (1981b) Use of repair-deficient strains of yeast to assay the activity of 40 coded compounds, Prog. Mutation Res., 1,502-516.
Shirasu, Y., M. Moriya, K. Kato, F. Lienard, H. Tezuka, S. Teramoto and T. Kada (1977) Mutagenicity screening on pesticides and modification products: a basis of carcino- genicity evaluation, Cold Spring Harbor Conference Cell Proliferation, Vol. 4, pp. 267-285.
Simmon, V., and G. Shepherd (1981) Mutagenic activity of 42 coded compounds in the Salmonella/microsome assay, Prog. Mutation Res., 1,333-342.
Skopek, T., B. Andon, D. Kaden and W. Thilly (1981) Muta- genic activity of 42 coded compounds using 8-azaguanine resistance as a genetic marker in Salmonella typhimurium, Prog. Mutation Res., 1,371-375.
Sram, R. (1975) Genetic risk from chemicals: mutagenicity studies and evaluation, Rev. Czech. Med., 21, 186-193.
Styles, J. (1981) Activity of 42 coded compounds in the BHK-21 cell transformation test, Prog. Mutation Res., 1, 638-646.
Teramoto, S., M. Moriya, K. Kato, H. Tezuka, S. Nakamura, A. Shingu and Y. Shirasu (1977) Mutagenicity testing on ethylenethiourea, Mutation Res., 56, 121-129.
Teramoto, S., A. Shingu and Y. Shirasu (1978) Induction of dominant lethal mutations after administration of eth- ylenethiourea in combination with nitrite or of N-nitroso- ethylenethiourea in mice, Mutation Res., 56, 335-340.
Thomson, J. (1981) Mutagenic activity of 42 coded compounds in the lambda induction assay, Prog. Mutation Res., 1, 224-235.
Topham, J. (1981) Evaluation of some chemicals by the sperm morphology assay, Prog. Mutation Res., 1, 718-720.
Trueman, R. (1981) Activity of 42 coded compounds in the Salmonella reverse mutation test, Prog. Mutation Res., 1, 343-350.
Truhaut, R., M. Fujita, N. Lich and M. Chaigueau (1973) Study of metabolic transformations of zineb, C.R. Acad. Sci. (Paris), Ser. D, 276, 229-233.
Tsuchimoto, T., and B. Matter (1981) Activity of coded com- pounds in the micronucleus test, Prog. Mutation Res., 1, 705-711.
Tweats, D. (1981) Activity of 42 coded compounds in a differential killing test using Escherichia coli strains WP2, WP67 (uvrA polA), and CM871 (uvrA lexA recA), Prog. Mutation Res., 1, 199-209.
Ulland, B., J. Weisberger, E. Weisberger, J. Rice and R. Cypher (1972) Thyroid cancer in rats from ethylene thiourea intake, J. Natl. Cancer Inst., 49, 583-584.
USEPA (1992) Ethylene bisdithiocarbamates (EBDCs); notice of intent to cancel and conclusion of special review, Fed- eral Register, 57, 7484-7530.
Valencia, R., and K. Houtchens (1981) Mutagenic activity of 10 coded compounds in the Drosophila sex-linked reces- sive lethal test, Prog. Mutation Res., 1,651-659.
van 't Hof, J., and L. Schairer (1982) Tradescantia assay system for gaseous mutagens. A report of the U.S. Envi- ronmental Protection Agency Gene-Tox Program, Muta- tion Res., 99, 303-315.
Venitt, S., and C. Crofton-Sleigh (1981) Mutagenicity of 42 coded compounds in a bacterial assay using Escherichia coli and Salmonella typhimurium, Prog. Mutation Res., 1, 351-360.
Woodruff, R., J. Mason, R. Valencia and S. Zimmering (1985) Chemical mutagenesis testing in Drosophila, V. Results of 53 coded compounds tested for the National Toxicology Program, Environ. Mutagen., 7, 677-702.
Wyrobek, A., L. Gordon and G. Watchmaker (1981) Effect of 17 chemical agents including 6 carcinogen/noncarcinogen pairs on sperm shape abnormalities in mice, Prog. Muta- tion Res., 1, 712-717.
Zimmermann, R., and I. Scheel (1981) Induction of mitotic gene conversion in strain D7 of Saccharomyces cerevisiae by 42 coded chemicals, Prog. Mutation Res., 1,481-490.





























