Embed Size (px)
Citation preview

1
Evaluation of Molecular Typing Methods in Characterizing a European
Collection of Epidemic Methicillin-Resistant Staphylococcus aureus (MRSA) -
the HARMONY collection
Barry D. Cookson1, D. Ashley Robinson
2, Alastair B. Monk
3, Stephen Murchan
4,
Ariane Deplano5, Rafaël de Ryck
5, Marc J. Struelens
5, Christina Scheel
6, Vivian
Fussing6, Saara Salmenlinna
7, Jaana Vuopio-Varkila
7, Christina Cuny
8, Wolfgang
Witte8, Panayotis T. Tassios
9, Nikolas J. Legakis
9, Willem van Leeeuwen
10, Alex van
Belkum10
, Anna Vindel11
, Javier Garaizer12
, Sara Haeggman13
, Barbro Olsson-
Liljequist13
, Ulrika Ransjo14
, Manica Muller-Premru15
, Waleria Hryniewicz16
, Angela
Rossney17
, Brian O’Connell17
, Benjamin D. Short18
, Jonathan Thomas18
, Simon
O’Hanlon18
and Mark C. Enright18
.
1Laboratory of Hospital Infection, Centre for Infections, Health Protection Agency,
London, UK; 2New York Medical School, New York, USA;
3Virginia
Commonwealth University, Virginia, USA; 4Health Protection Surveillance Centre,
Dublin, Ireland; 5Laboratoire de Microbiologie, Hopital Erasme, Bruxelles, Belgium;
6Statens Serum Institut, Copenhagen, Denmark;
7National Public Health Laboratory
(KTL), Helsinki, Finland; 8Robert Koch-Institut, Wernigerode (Harz), Germany;
9
Department of Microbiology, University of Athens, Athens, Greece; 10
Erasmus MC
Center, Rotterdam, The Netherlands; 11
Centro Nacional de Microbiología, Instituto de
Salud Carlos III, Madrid, Spain; 12
Dpt. Immunol., Microbiol. y Parasitol., F.
Farmacia, UPV/EHU, Vitoria-Gasteiz, Spain; 13
Swedish Institute for Infectious
Disease Control, Solna, Sweden; 14
Swedish Institute for Infectious Disease Control,
ACCEPTED
Copyright © 2007, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.J. Clin. Microbiol. doi:10.1128/JCM.02402-06 JCM Accepts, published online ahead of print on 11 April 2007
on August 30, 2018 by guest
http://jcm.asm
.org/D
ownloaded from

2
Bacteriology, Solna, Sweden; 15
Institute of Microbiology, University of Ljubljana,
Ljubljana, Slovenia; 16
Sera and Vaccines Central Research Laboratory, Warsaw,
Poland; 17
National MRSA Reference Laboratory, St James’s Hospital, James’s St.,
Dublin 8, Ireland 18
Department of Infectious Disease Epidemiology, Imperial College,
London.
Correspondence to: Dr Mark C. Enright, Department of Infectious Disease
Epidemiology, Imperial College, London, United Kingdom. E-mail:
[email protected]; Telephone +44 207 5943417; Fax +44 207 594 3693.
ACCEPTED
on August 30, 2018 by guest
http://jcm.asm
.org/D
ownloaded from

3
Abstract
We analyzed a representative sample of methicillin-resistant Staphylococcus aureus
(MRSA) from 11 European countries (referred to as the HARMONY collection)
using three molecular typing methods used within the HARMONY group to examine
their usefulness for large, multicentre MRSA surveillance networks that use these
different laboratory methodologies.
MRSA isolates were collected based on their prevalence in each centre and their
genetic diversity, assessed by pulsed-field gel electrophoresis (PFGE). PFGE
groupings (≤ 3 bands difference between patterns) were compared to those made by
sequencing of the variable repeats in the protein A gene spa and clonal designations
based on multilocus sequence typing (MLST) combined with PCR analysis of the
staphylococcal chromosome cassette containing the mec genes involved in methicillin
resistance (SCCmec).
A high level of discrimination was achieved using each of the three methodologies
with discriminatory indices of between 89.5% and 91.9% with overlapping 95%
confidence intervals. There was also a high level of concordance of groupings made
using each method. MLST / SCCmec typing distinguished 10 groups containing at
least two isolates and these correspond to the majority of nosocomial MRSA clones
described in the literature. PFGE and spa typing resolved 34 and 31 subtypes
respectively within these 10 MRSA clones with each subtype differing only slightly
from the most common pattern using each method.
ACCEPTED
on August 30, 2018 by guest
http://jcm.asm
.org/D
ownloaded from

4
The HARMONY group has found that the methods used in this study differ in their
availability and affordability to European centres involved in MRSA surveillance.
Here we demonstrate that the integration of such technologies is achievable, although
common protocols (as we have developed for PFGE) may also be important, as is the
use of centralised internet sites to facilitate data analysis. PFGE and spa typing data
from analysis of MRSA isolates from the many centres that have access to the
relevant equipment can be compared to reference patterns / sequences and clonal
designations made. In the majority of cases these will correspond to those made by
the (more expensive) method of choice - MLST / SCCmec typing and these
alternative methods can therefore be used as frontline typing systems for multicentre
surveillance of MRSA.
Introduction
Methicillin-resistant Staphylococcus aureus (MRSA) is among the most common
nosocomial pathogens globally and is generally acknowledged as the most significant
due to the burden of disease it causes and to the evolution and global spread of
multidrug resistant clones. MRSA isolation rates in the USA, parts of Europe and
Asia have been increasing for more than four decades (36) and recent figures show
that in some areas >50% of S. aureus bacteraemias are caused by MRSA (1, 6, 7).
Emerging intermediate, and more recently high-level (vanA-encoded), vancomycin
resistance (2, 22) and increasing numbers of multidrug-resistant MRSA emphasize the
importance of effective antimicrobial prescribing and infection control measures that
can be informed usefully by molecular typing results.
Epidemic spread of S. aureus in hospitals and intercontinental spread of a
particular clone was first demonstrated in the 1950s using bacteriophage typing (37).
ACCEPTED
on August 30, 2018 by guest
http://jcm.asm
.org/D
ownloaded from

5
This method has largely been replaced in centres with low workloads as an
epidemiological tool by genotyping approaches that index chromosomal variation and
offer advantages in typeability, discrimination and reproducibility. Many different
genotyping methods are currently in use, but the most popular is pulsed-field gel
electrophoresis (PFGE) of SmaI–digested genomic DNA (27). Other techniques that
have been explored include ribotyping (34) and PCR-based methods such as
repetitive-element PCR (13), AFLP (amplified fragment length polymorphism) typing
(23) and more recently DNA sequencing approaches (16, 35, 39, 40). Typing methods
should have high and relevant discriminatory power and typeability, good
reproducibility, applicability to all organisms of interest, ease of use, portability (that
is, they should produce data that can easily be transferred between laboratories or
presented in published work) and low cost (43).
PFGE is the most commonly used method when studying local or short-term
S. aureus epidemiology. It has proven invaluable in investigations of nosocomial
outbreaks but difficulties in reproducibility and inter laboratory reliability has limited
its application to relatively small studies (10). Multicentre studies using PFGE are
now possible due to recent advances in the standardization of electrophoresis
conditions (10, 32) and the development of normalization and analysis software (15).
Interpretative criteria for use in comparing complex PFGE patterns in outbreaks have
been applied to non-outbreak situations to track the national and international
dissemination of S. aureus clones (44). The use of PFGE typing with adjusted
interpretation criteria for grouping patterns with < 7 bands difference has been shown
to correspond to clonal assignments made by other methods (12). The main criticisms
of this technique for S. aureus are that PFGE may, on occasion, be too discriminatory
for other than local or short-term epidemiological analyses, the arbitrary nature of the
ACCEPTED
on August 30, 2018 by guest
http://jcm.asm
.org/D
ownloaded from

6
interpretive criteria used and the requirement for occasional subjective analysis of
complex band patterns (30).
Multilocus sequence typing (MLST) (26) has had a large impact on the field
of bacterial typing and it has been used as an investigatory tool in many studies of S.
aureus evolution and epidemiology (4, 11, 16, 17, 28). MLST has a major advantage
over PFGE as a reference method due to the unambiguous nature of DNA sequences
which can be stored easily along with corresponding clinical information on each
isolate in internet-linked databases. The S. aureus MLST website (www.mlst.net)
currently contains information on > 1500 isolates from humans and animals from 40
different countries and represents a useful global resource for the study of the
epidemiology of this species and the surveillance of hyper-virulent and / or antibiotic
resistant clones.
In order to define MRSA clones, MLST has been used in conjunction with
PCR analysis of the element conferring methicillin resistance – SCCmec
(staphylococcal chromosomal cassette mec (17, 25). An international nomenclature
for MRSA clones using the MLST and SCCmec designations has been accepted by
the International Union of Microbiological Societies subcommittee on S. aureus
typing. This unambiguous nomenclature is characterizes clones based on their MLST
sequence type, SCCmec type and phenotype. For example members of the United
Kingdom Epidemic MRSA -15 clone would be referred to as ST22-MRSA-IV –
MLST sequence type 22, MRSA, SCCmec type IV. Vancomycin-intermediately
susceptible descendants would be referred to as ST22-VISA-IV. This scheme allows
the efficient worldwide tracking and surveillance of MRSA clones.
MLST / SCCmec typing is widely regarded as the reference method for
defining MRSA clones. However, it is not as widely applicable as other methods for
ACCEPTED
on August 30, 2018 by guest
http://jcm.asm
.org/D
ownloaded from

7
high volume analysis of MRSA due to the cost of DNA sequencing (MLST requires 7
PCR reactions plus 14 DNA sequencing reactions per isolate). In addition SCCmec
typing requires an ever increasing number of primers as new alleles are found (9). spa
typing has been proposed as a rapid sequence-based approach to characterize MRSA.
It has high portability, discrimination and ease of use (39). An automatically curated,
internet-accessible spa sequence database is now available (3). Several recent
epidemiological studies have used PFGE and / or spa typing to determine the genetic
relatedness of large numbers of isolates, and MLST / SCCmec typing for further
characterization of representative isolates of each genotype found (5, 12, 18).
The EU-funded HARMONY project (29) standardized PFGE in eight
European centres to facilitate transfer of molecular typing data between laboratories.
It now has the support of the International Union of Microbiological Societies
subcommittee on S. aureus typing with the longer term goal to allow the development
of networks of Reference Laboratories and Research Centres to provide
epidemiologically useful information about the emergence and spread of MRSA
clones. The potential benefits of such networks would be most obvious in the early
detection of MRSA with new or multiple antibiotic resistance or pathogenic potential
and determining whether such strains had emerged from parallel evolution and in
monitoring their intra- or inter-country spread.
In this study we analysed a broadly representative collection of MRSA isolates
from 11 European countries. Isolates were assigned to MRSA clones on the basis of
MLST / SCCmec typing and the congruence between these groupings and those made
using PFGE and spa typing was assessed.
ACCEPTED
on August 30, 2018 by guest
http://jcm.asm
.org/D
ownloaded from

8
Materials and methods
Bacterial isolates
The HARMONY collection of MRSA was used in this study. It consists of 98 clinical
isolates collected from 11 European countries between 1981 and 1998 as part of the
HARMONY project (contract number BMH4-CT96) (29). The countries that
submitted isolates were Belgium, Finland, France, Germany, Greece, Ireland, Poland,
Slovenia, Spain, Sweden and the United Kingdom. Details on each isolate are
available from the HARMONY website (www.harmony-microbe.net). The isolates
were chosen by each contributing centre to represent currently circulating epidemic or
otherwise important nosocomial clones from the mid-1980s to 1998. These isolates
still represent the major clones currently causing hospital-acquired MRSA outbreaks
in Europe (17). Countries were invited where possible to submit isolates that
represented closely related PFGE types and important variants of these representative
clones.
All isolates were stored in 30% glycerol / nutrient broth at -80°C and grown on blood
agar plates at 37°C overnight.
PFGE
PFGE following SmaI digestion was performed and analyzed according to the
HARMONY protocol (29). Comparisons and groupings of PFGE patterns by Dice
coefficients (14) and UPGMA (41), respectively, was performed using GelCompar
and BioNumerics software (Applied Maths, Belgium). Results were represented both
as distinct PFGE patterns (indistinguishable band patterns were assigned the same
ACCEPTED
on August 30, 2018 by guest
http://jcm.asm
.org/D
ownloaded from

9
PFGE pattern number) and as PFGE types (each type included closely related PFGE
patterns that differed by ≤ 3 bands). By using this definition of PFGE types, isolates
assumed to be closely related according to the guidelines proposed by Tenover et al
(44) for short-term outbreak-associated isolates were grouped.
MLST
MLST was performed as described previously (16). Briefly, seven housekeeping gene
fragments (~500 bp) were sequenced and compared to known alleles at each locus on
the MLST website (www.mlst.net). Allelic profiles, each consisting of seven allele
numbers, defined sequence types (STs). STs sharing 100% genetic identity in at least
five of seven MLST loci were grouped into a clonal complex (CC) named after it’s
presumed ancestral genotype, as described previously (19).
SCCmec typing
The four main SCCmec structures were differentiated by PCR detection of the SCC
type (I, II, III, IV) and the class of the mec region (A or B) using conventional (25,
31) and multiplex PCR (33).
spa sequence typing
Sequencing of the short sequence repeats region of the spa gene was performed as
described previously (39). Sequences obtained were compared to those held on the
SpaServer (http://www.ridom.de/spaserver/query) (21).
Diversity index
ACCEPTED
on August 30, 2018 by guest
http://jcm.asm
.org/D
ownloaded from

10
The discriminatory power of each typing method was represented by an index of
diversity (DI) (24) which represents the probability that two isolates selected from the
sample at random will have different types. 95% confidence intervals were calculated
as described in Grundmann et al (20).
Results
PFGE
PFGE following SmaI digestion resolved 81 PFGE patterns whose profiles differed by
at least 1 band (Table 1). PFGE patterns 8 and 13 were the most common, shared by 4
isolates each. PFGE pattern 8 isolates were from four different countries - Finland,
Germany, Sweden and Spain and those having PFGE pattern 13 were from France
and Belgium only. Three PFGE patterns (16, 18 and 70) were shared by three isolates
each and five patterns (1, 7, 37, 40 and 50) by two isolates each. The majority of
study isolates (71/98 or 72.4%) had unique PFGE patterns.
The number of PFGE types (PFGE patterns that differed at ≤ 3 band positions)
was 34. These were numbered (1-34) arbitrarily. Nineteen of these types comprised
only one PFGE pattern each. Twenty-four isolates from seven countries belonged to
PFGE type 3 which contained all isolates with PFGE patterns 8 and 13 detailed
above. Eight isolates (from Germany and Slovenia) were of type 13 (the Southern
German clone), seven isolates (Sweden, Ireland and UK) were of type 25, six isolates
(France) were of type 1, five isolates (Greece and France) were of type 7 and five
isolates (Germany) were of type 24 (the Berlin clone). Four isolates (from Belgium,
Sweden and Finland) belonged to type 22 and four isolates (Finland and UK) were of
type 27 (UK EMRSA-16). Three isolates each were of types 5 (Finland, Poland) and 8
ACCEPTED
on August 30, 2018 by guest
http://jcm.asm
.org/D
ownloaded from

11
(Germany) and two isolates each were of types 4 (Finland, Slovenia), 9 (Finland,
Slovenia), 14 (Germany), 15 (Slovenia) and 19 (Belgium).
spa typing
Thirty-one spa sequence types were resolved and identified by interrogating the
RIDOM SpaServer. Eighteen spa types were unique to one isolate. spa type t051 was
found in 18 isolates, t001 and t037 in 13 isolates each, t008 in nine, t004 in five, t018
and t030 in four each, t009 and t022 in three each and t015, t032, t038 and t045 in
two isolates each.
MLST / SCCmec typing
Seventeen MLST STs were resolved, the most common of which was sequence type
ST247 shared by 23 isolates (Table 1). Six isolates were not typeable using published
primers recommended for analysis of SCCmec (25, 31) and may therefore represent
novel types of this element. Combined MLST and SCCmec typing resolved 24 MRSA
clones, which included five STs with >1 type of SCCmec element (Table 1). The
common clone names that have been used in other published studies of MRSA
epidemiology are also shown in Table 1.
Diversity Index
Table 2 details the discriminatory power of PFGE, MLST, MLST/SCCmec, and spa
typing for the 98 MRSA isolates of the HARMONY collection. PFGE pattern
analysis gave a DI of 99.5% (95% CI, 99.1-99.9) indicating that when two isolates
were chosen at random they would have different patterns 99.5% of the time. The four
other methods gave lower levels of discrimination of between 88.7% and 92.1% with
ACCEPTED
on August 30, 2018 by guest
http://jcm.asm
.org/D
ownloaded from

12
overlapping 95% CIs. spa typing was the second most discriminatory method
followed by PFGE type using the ≤ 3 bands difference criterion, then MLST /
SCCmec typing followed by MLST alone.
Comparison of clonal assignments
MLST / SCCmec typing identified 10 clones containing at least two isolates. These
represent the majority of pandemic nosocomial MRSA clones described in the
scientific literature (17) (Tables 1 and 3). ST247-MRSA-I commonly referred to as
the Iberian clone (38), was the most frequently represented clone in this study with
isolates coming from seven countries (Table 1). Twenty-one of 23 isolates belonging
to the ST247-MRSA-I clone were of PFGE type 3 but PFGE type 3 isolates were not
exclusive to this clone as three PFGE type 3 isolates were ST8-MRSA-IV. However,
ST247 and ST8 are closely related, both belonging to CC8. Two isolates of ST247-
MRSA-I were PFGE type 4. Of the 21 ST247-MRSA-I isolates, 18 had the same spa
type (t051) and three were unique (t008, t052 and t200).
PFGE was the most discriminatory technique followed by spa typing and then PFGE
using ≤ 3 bands difference although the latter two methods differed by very little in
their discriminatory power (Table 2). MLST and MLST augmented with SCCmec
typing were also very similar in their discriminatory power due to the fact that only
ST5 isolates were found with different SCCmec elements (disregarding SCCmec non-
typeables). However study isolates were not randomly seleced as some participating
centres submitted several PFGE variants of their main types. There was perfect
concordance between MLST CC and PFGE analysis at the 65% level (data not
shown) and this might be explored as a way of designing a nomenclature (it has been
adopted by some member of the HARMONY group).
ACCEPTED
on August 30, 2018 by guest
http://jcm.asm
.org/D
ownloaded from

13
spa sequence types were not shared between isolates from different clonal
complexes (Table 1). The same spa type, however, was shared between different
clones in two cases: t008 was present in ST247-MRSA-I and ST8-MRSA-IV (both
CC8) and t001 was found in both ST228-MRSA-I and ST5-MRSA-I (both CC5).
Discussion
The choice of typing method used to investigate the epidemiology of MRSA should
primarily be dictated by the environment and / or timescale being examined and the
aims of the investigation. For a single MRSA outbreak in one hospital, for example, a
highly discriminatory typing method that could differentiate between index case-
associated isolates and isolates which are not associated with the index case would be
required. As a short timescale is being examined, little genomic variation would be
expected to occur in index case-associated isolates during the course of the outbreak
and minimal data interpretation would be required. PFGE typing has been the method
applied most commonly in such scenarios. Interpretation of the banding patterns
generated by PFGE was refined by the suggested guidelines of Tenover (44), which
allows variant PFGE patterns to be included in outbreak assignments. These criteria
are biologically-based, validated in outbreak investigations (8, 42) and have been used
in numerous studies of MRSA epidemiology. However, they should only be used for
short-term outbreak investigation and need validation outside of such scenarios (42).
An approach recommended for more widespread epidemiological typing is to
initially use PFGE and then select isolates for clonal confirmation by SCCmec
analysis and MLST. spa typing could be used instead of PFGE if sequencing capacity
and expense is not an issue. spa typing provides data that are as concordant with the
MLST designation as PFGE but use of spa typing does not obviate the need for
ACCEPTED
on August 30, 2018 by guest
http://jcm.asm
.org/D
ownloaded from

14
SCCmec analysis to determine the clonal designation of an isolate.
A potential problem with spa typing is that it involves sequencing of only one small
region of the chromosome which is subject to recombination between unrelated
clones. This could result in isolates exhibiting the same spa type when they are shown
to be unrelated by other methods. PFGE and MLST both investigate multiple
locations around the chromosome. However, spa typing and MLST are portable and
can be compared directly using global databases.
Potential difficulties may arise when using PFGE to analyze epidemiological
scenarios involving large numbers of study centres or long timescales. PFGE has been
used in many studies to show international spread of MRSA and frequently the
criteria used for microepidemiological analyses have been employed. SmaI PFGE
patterns of MRSA clones are known to change considerably during transmission of
some strains over the course of years. In a number of cases, isolates assigned to
different clones by PFGE have been shown to be genetically highly-related using
MLST. These limitations should not hinder the continued development of networks of
centres using methods such as PFGE and spa typing as long as examples of each
genotype are assigned to MRSA clones using MLST and SCCmec typing.
Acknowledgements.
This paper is dedicated to the memory of Névine El Solh. MCE is a Royal Society
University Research Fellow. This work was funded by the Wellcome Trust (grant
GR073363 to MCE) and a EUDGXII grant (contract No BMH4-CT96b) to BC
(project leader). Further details of HARMONY can be obtained from
ACCEPTED
on August 30, 2018 by guest
http://jcm.asm
.org/D
ownloaded from

15
References
1. 2002. National Nosocomial Infections Surveillance (NNIS) System Report,
data summary from January 1992 to June 2002, issued August 2002. Am J
Infect Control 30:458-75.
2. 2004. Vancomycin-resistant Staphylococcus aureus--New York, 2004.
MMWR Morb Mortal Wkly Rep 53:322-3.
3. Aires-de-Sousa, M., K. Boye, H. de Lencastre, A. Deplano, M. C. Enright,
J. Etienne, A. Friedrich, D. Harmsen, A. Holmes, X. W. Huijsdens, A. M.
Kearns, A. Mellmann, H. Meugnier, J. K. Rasheed, E. Spalburg, B.
Strommenger, M. J. Struelens, F. C. Tenover, J. Thomas, U. Vogel, H.
Westh, J. Xu, and W. Witte. 2006. High interlaboratory reproducibility of
DNA sequence-based typing of bacteria in a multicenter study. J Clin
Microbiol 44:619-21.
4. Aires de Sousa, M., C. Bartzavali, I. Spiliopoulou, I. S. Sanches, M. I.
Crisostomo, and H. de Lencastre. 2003. Two international methicillin-
resistant Staphylococcus aureus clones endemic in a university hospital in
Patras, Greece. J Clin Microbiol 41:2027-32.
5. Aires de Sousa, M., T. Conceicao, C. Simas, and H. de Lencastre. 2005.
Comparison of genetic backgrounds of methicillin-resistant and -susceptible
Staphylococcus aureus isolates from Portuguese hospitals and the community.
J Clin Microbiol 43:5150-7.
6. Anonymous. 2003. European Antimicrobial Resistance Surveillance System
(EARSS) annual report 2003.
7. Bell, J. M., and J. D. Turnidge. 2002. High prevalence of oxacillin-resistant
Staphylococcus aureus isolates from hospitalized patients in Asia-Pacific and
South Africa: results from SENTRY antimicrobial surveillance program,
1998-1999. Antimicrob Agents Chemother 46:879-81.
8. Blanc, D. S., M. J. Struelens, A. Deplano, R. De Ryck, P. M. Hauser, C.
Petignat, and P. Francioli. 2001. Epidemiological validation of pulsed-field
gel electrophoresis patterns for methicillin-resistant Staphylococcus aureus. J
Clin Microbiol 39:3442-5.
9. Chongtrakool, P., T. Ito, X. X. Ma, Y. Kondo, S. Trakulsomboon, C.
Tiensasitorn, M. Jamklang, T. Chavalit, J. H. Song, and K. Hiramatsu. 2006. Staphylococcal cassette chromosome mec (SCCmec) typing of
methicillin-resistant Staphylococcus aureus strains isolated in 11 Asian
countries: a proposal for a new nomenclature for SCCmec elements.
Antimicrob Agents Chemother 50:1001-12.
10. Chung, M., H. de Lencastre, P. Matthews, A. Tomasz, I. Adamsson, M.
Aries de Sousa, T. Camou, C. Cocuzza, A. Corso, I. Couto, A. Dominguez,
M. Gniadkowski, R. Goering, A. Gomes, K. Kikuchi, A. Marchese, R.
Mato, O. Melter, D. Oliveira, R. Palacio, R. Sa-Leao, I. Santos Sanches, J.
H. Song, P. T. Tassios, and P. Villari. 2000. Molecular typing of methicillin-
resistant Staphylococcus aureus by pulsed-field gel electrophoresis:
comparison of results obtained in a multilaboratory effort using identical
protocols and MRSA strains. Microb Drug Resist 6:189-98.
11. Coombs, G. W., G. R. Nimmo, J. M. Bell, F. Huygens, G. O'Brien F, M. J.
Malkowski, J. C. Pearson, A. J. Stephens, and P. M. Giffard. 2004.
Genetic Diversity among Community Methicillin-Resistant Staphylococcus
ACCEPTED
on August 30, 2018 by guest
http://jcm.asm
.org/D
ownloaded from

16
aureus Strains Causing Outpatient Infections in Australia. J Clin Microbiol
42:4735-43.
12. Denis, O., A. Deplano, C. Nonhoff, R. De Ryck, R. de Mendonca, S.
Rottiers, R. Vanhoof, and M. J. Struelens. 2004. National surveillance of
methicillin-resistant Staphylococcus aureus in Belgian hospitals indicates
rapid diversification of epidemic clones. Antimicrob Agents Chemother
48:3625-9.
13. Deplano, A., A. Schuermans, J. Van Eldere, W. Witte, H. Meugnier, J.
Etienne, H. Grundmann, D. Jonas, G. T. Noordhoek, J. Dijkstra, A. van
Belkum, W. van Leeuwen, P. T. Tassios, N. J. Legakis, A. van der Zee, A.
Bergmans, D. S. Blanc, F. C. Tenover, B. C. Cookson, G. O'Neil, and M.
J. Struelens. 2000. Multicenter evaluation of epidemiological typing of
methicillin-resistant Staphylococcus aureus strains by repetitive-element PCR
analysis. The European Study Group on Epidemiological Markers of the
ESCMID. J Clin Microbiol 38:3527-33.
14. Dice, L. R. 1945. Measures of the amount of ecologic association between
species. Ecology 26:297-302.
15. Duck, W. M., C. D. Steward, S. N. Banerjee, J. E. McGowan, Jr., and F.
C. Tenover. 2003. Optimization of computer software settings improves
accuracy of pulsed-field gel electrophoresis macrorestriction fragment pattern
analysis. J Clin Microbiol 41:3035-42.
16. Enright, M. C., N. P. Day, C. E. Davies, S. J. Peacock, and B. G. Spratt.
2000. Multilocus sequence typing for characterization of methicillin-resistant
and methicillin-susceptible clones of Staphylococcus aureus. J Clin Microbiol
38:1008-15.
17. Enright, M. C., D. A. Robinson, G. Randle, E. J. Feil, H. Grundmann, and
B. G. Spratt. 2002. The evolutionary history of methicillin-resistant
Staphylococcus aureus (MRSA). Proc Natl Acad Sci U S A 99:7687-92.
18. Faria, N. A., D. C. Oliveira, H. Westh, D. L. Monnet, A. R. Larsen, R.
Skov, and H. de Lencastre. 2005. Epidemiology of emerging methicillin-
resistant Staphylococcus aureus (MRSA) in Denmark: a nationwide study in a
country with low prevalence of MRSA infection. J Clin Microbiol 43:1836-
42.
19. Feil, E. J., J. E. Cooper, H. Grundmann, D. A. Robinson, M. C. Enright,
T. Berendt, S. J. Peacock, J. M. Smith, M. Murphy, B. G. Spratt, C. E.
Moore, and N. P. Day. 2003. How clonal is Staphylococcus aureus? J
Bacteriol 185:3307-16.
20. Grundmann, H., S. Hori, and G. Tanner. 2001. Determining confidence
intervals when measuring genetic diversity and the discriminatory abilities of
typing methods for microorganisms. J Clin Microbiol 39:4190-2.
21. Harmsen, D., H. Claus, W. Witte, J. Rothganger, D. Turnwald, and U.
Vogel. 2003. Typing of methicillin-resistant Staphylococcus aureus in a
university hospital setting by using novel software for spa repeat
determination and database management. J Clin Microbiol 41:5442-8.
22. Hiramatsu, K., H. Hanaki, T. Ino, K. Yabuta, T. Oguri, and F. C.
Tenover. 1997. Methicillin-resistant Staphylococcus aureus clinical strain
with reduced vancomycin susceptibility. J Antimicrob Chemother 40:135-6.
23. Hookey, J. V., V. Edwards, S. Patel, J. F. Richardson, and B. D. Cookson.
1999. Use of fluorescent amplified fragment length polymorphism (fAELP) to
ACCEPTED
on August 30, 2018 by guest
http://jcm.asm
.org/D
ownloaded from

17
characterise methicillin-resistant Staphylococcus aureus. J Microbiol Methods
37:7-15.
24. Hunter, P. R., and M. A. Gaston. 1988. Numerical index of the
discriminatory ability of typing systems: an application of Simpson's index of
diversity. J Clin Microbiol 26:2465-6.
25. Ito, T., Y. Katayama, K. Asada, N. Mori, K. Tsutsumimoto, C.
Tiensasitorn, and K. Hiramatsu. 2001. Structural comparison of three types
of staphylococcal cassette chromosome mec integrated in the chromosome in
methicillin-resistant Staphylococcus aureus. Antimicrob Agents Chemother
45:1323-36.
26. Maiden, M. C., J. A. Bygraves, E. Feil, G. Morelli, J. E. Russell, R. Urwin,
Q. Zhang, J. Zhou, K. Zurth, D. A. Caugant, I. M. Feavers, M. Achtman,
and B. G. Spratt. 1998. Multilocus sequence typing: a portable approach to
the identification of clones within populations of pathogenic microorganisms.
Proc Natl Acad Sci U S A 95:3140-5.
27. Maslow, J. N., A. M. Slutsky, and R. D. Arbeit. 1993. Application of
pulsed-field gel electrophoresis to molecular epidemiology., p. 563-572. In T.
J. White (ed.), Diagnostic molecular microbiology: principles and
applications. American Society for Microbiology, Washington D.C.
28. Mato, R., F. Campanile, S. Stefani, M. I. Crisostomo, M. Santagati, S. I.
Sanches, and H. de Lencastre. 2004. Clonal types and multidrug resistance
patterns of methicillin-resistant Staphylococcus aureus (MRSA) recovered in
Italy during the 1990s. Microb Drug Resist 10:106-13.
29. Murchan, S., M. E. Kaufmann, A. Deplano, R. de Ryck, M. Struelens, C.
E. Zinn, V. Fussing, S. Salmenlinna, J. Vuopio-Varkila, N. El Solh, C.
Cuny, W. Witte, P. T. Tassios, N. Legakis, W. van Leeuwen, A. van
Belkum, A. Vindel, I. Laconcha, J. Garaizar, S. Haeggman, B. Olsson-
Liljequist, U. Ransjo, G. Coombes, and B. Cookson. 2003. Harmonization
of pulsed-field gel electrophoresis protocols for epidemiological typing of
strains of methicillin-resistant Staphylococcus aureus: a single approach
developed by consensus in 10 European laboratories and its application for
tracing the spread of related strains. J Clin Microbiol 41:1574-85.
30. Murchan, S., H. M. Aucken, L. O'Neill G, M. Ganner, and B. D. Cookson.
2004. Emergence, spread, and characterization of phage variants of epidemic
methicillin-resistant Staphylococcus aureus 16 in England and Wales. J Clin
Microbiol 42:5154-60.
31. Okuma, K., K. Iwakawa, J. D. Turnidge, W. B. Grubb, J. M. Bell, F. G.
O'Brien, G. W. Coombs, J. W. Pearman, F. C. Tenover, M. Kapi, C.
Tiensasitorn, T. Ito, and K. Hiramatsu. 2002. Dissemination of new
methicillin-resistant Staphylococcus aureus clones in the community. J Clin
Microbiol 40:4289-94.
32. Oliveira, D. C., A. Tomasz, and H. de Lencastre. 2001. The evolution of
pandemic clones of methicillin-resistant Staphylococcus aureus: identification
of two ancestral genetic backgrounds and the associated mec elements. Microb
Drug Resist 7:349-61.
33. Oliveira, D. C., and H. de Lencastre. 2002. Multiplex PCR strategy for rapid
identification of structural types and variants of the mec element in
methicillin-resistant Staphylococcus aureus. Antimicrob Agents Chemother
46:2155-61.
ACCEPTED
on August 30, 2018 by guest
http://jcm.asm
.org/D
ownloaded from

18
34. Preheim, L., D. Pitcher, R. Owen, and B. Cookson. 1991. Typing of
methicillin resistant and susceptible Staphylococcus aureus strains by
ribosomal RNA gene restriction patterns using a biotinylated probe. Eur J Clin
Microbiol Infect Dis 10:428-36.
35. Robinson, D. A., and M. C. Enright. 2003. Evolutionary models of the
emergence of methicillin-resistant Staphylococcus aureus. Antimicrob Agents
Chemother 47:3926-34.
36. Robinson, D. A., and M. C. Enright. 2004. Multilocus sequence typing and
the evolution of methicillin-resistant Staphylococcus aureus. Clin Microbiol
Infect 10:92-7.
37. Rountree, P. M., and B. M. Freeman. 1955. Infections Caused by a
Particular Phage Type of Staphylococcus aureus. Med. J. Aust. 2:157.
38. Sanches, I. S., M. Ramirez, H. Troni, M. Abecassis, M. Padua, A. Tomasz,
and H. de Lencastre. 1995. Evidence for the geographic spread of a
methicillin-resistant Staphylococcus aureus clone between Portugal and Spain.
J Clin Microbiol 33:1243-6.
39. Shopsin, B., M. Gomez, S. O. Montgomery, D. H. Smith, M. Waddington,
D. E. Dodge, D. A. Bost, M. Riehman, S. Naidich, and B. N. Kreiswirth. 1999. Evaluation of protein A gene polymorphic region DNA sequencing for
typing of Staphylococcus aureus strains. J Clin Microbiol 37:3556-63.
40. Shopsin, B., M. Gomez, M. Waddington, M. Riehman, and B. N.
Kreiswirth. 2000. Use of coagulase gene (coa) repeat region nucleotide
sequences for typing of methicillin-resistant Staphylococcus aureus strains. J
Clin Microbiol 38:3453-6.
41. Sneath, P. H., and R. R. Sokal. 1962. Numerical taxonomy. Nature 193:855-
60.
42. Struelens, M. J., A. Deplano, C. Godard, N. Maes, and E. Serruys. 1992.
Epidemiologic typing and delineation of genetic relatedness of methicillin-
resistant Staphylococcus aureus by macrorestriction analysis of genomic DNA
by using pulsed-field gel electrophoresis. J Clin Microbiol 30:2599-605.
43. Struelens, M. J. 1996. Consensus guidelines for appropriate use and
evaluation of microbial epidemiologic typing systems. Clin Microbiol Infect
2:2-11.
44. Tenover, F. C., R. D. Arbeit, R. V. Goering, P. A. Mickelsen, B. E.
Murray, D. H. Persing, and B. Swaminathan. 1995. Interpreting
chromosomal DNA restriction patterns produced by pulsed-field gel
electrophoresis: criteria for bacterial strain typing. J Clin Microbiol 33:2233-9.
ACCEPTED
on August 30, 2018 by guest
http://jcm.asm
.org/D
ownloaded from
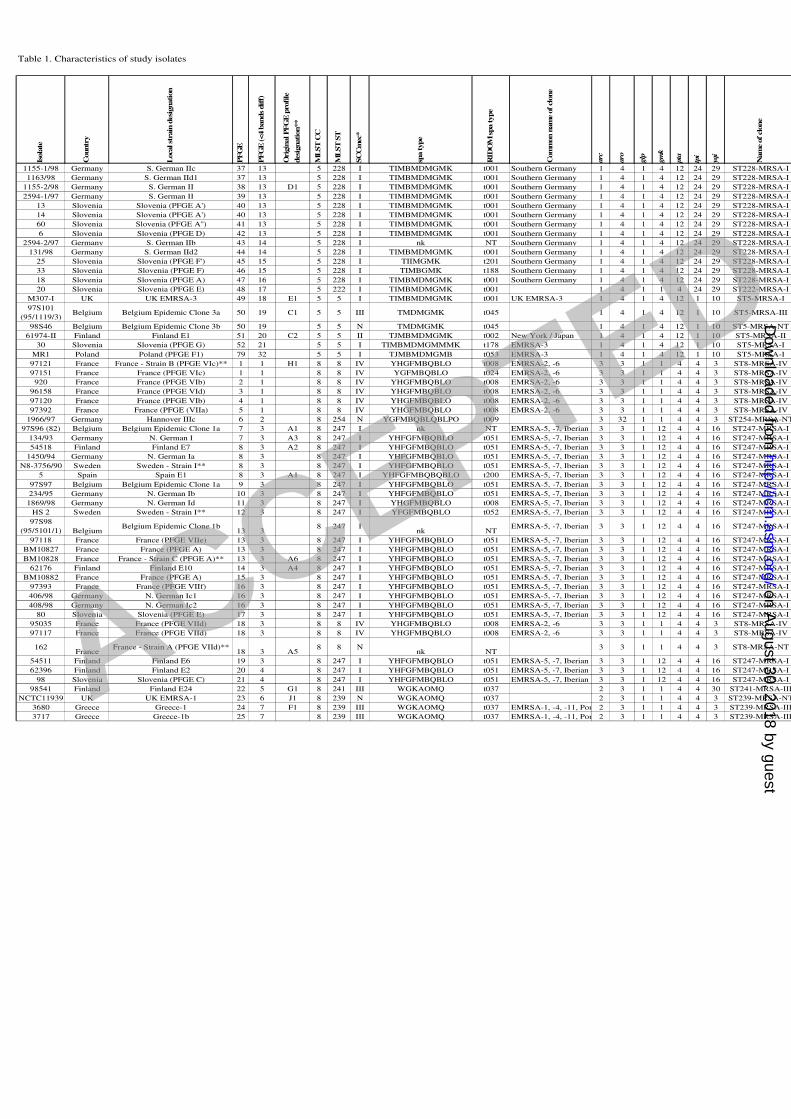
Table 1. Characteristics of study isolates
Isola
te
Countr
y
Loca
l st
rain
desi
gnati
on
PF
GE
PF
GE
(<4 b
ands
dif
f)
Orig
inal PF
GE
pro
file
desi
gnation**
ML
ST
CC
ML
ST
ST
SC
Cm
ec*
spa type
RID
OM
spa type
Com
mon n
am
e of cl
one
arc
aro
glp
gm
k
pta
tpi
yqi
Nam
e of cl
one
1155-1/98 Germany S. German IIc 37 13 5 228 I TIMBMDMGMK t001 Southern Germany 1 4 1 4 12 24 29 ST228-MRSA-I
1163/98 Germany S. German IId1 37 13 5 228 I TIMBMDMGMK t001 Southern Germany 1 4 1 4 12 24 29 ST228-MRSA-I
1155-2/98 Germany S. German II 38 13 D1 5 228 I TIMBMDMGMK t001 Southern Germany 1 4 1 4 12 24 29 ST228-MRSA-I
2594-1/97 Germany S. German II 39 13 5 228 I TIMBMDMGMK t001 Southern Germany 1 4 1 4 12 24 29 ST228-MRSA-I
13 Slovenia Slovenia (PFGE A') 40 13 5 228 I TIMBMDMGMK t001 Southern Germany 1 4 1 4 12 24 29 ST228-MRSA-I
14 Slovenia Slovenia (PFGE A') 40 13 5 228 I TIMBMDMGMK t001 Southern Germany 1 4 1 4 12 24 29 ST228-MRSA-I
60 Slovenia Slovenia (PFGE A'') 41 13 5 228 I TIMBMDMGMK t001 Southern Germany 1 4 1 4 12 24 29 ST228-MRSA-I
6 Slovenia Slovenia (PFGE D) 42 13 5 228 I TIMBMDMGMK t001 Southern Germany 1 4 1 4 12 24 29 ST228-MRSA-I
2594-2/97 Germany S. German IIb 43 14 5 228 I nk NT Southern Germany 1 4 1 4 12 24 29 ST228-MRSA-I
131/98 Germany S. German IId2 44 14 5 228 I TIMBMDMGMK t001 Southern Germany 1 4 1 4 12 24 29 ST228-MRSA-I
25 Slovenia Slovenia (PFGE F') 45 15 5 228 I TIIMGMK t201 Southern Germany 1 4 1 4 12 24 29 ST228-MRSA-I
33 Slovenia Slovenia (PFGE F) 46 15 5 228 I TIMBGMK t188 Southern Germany 1 4 1 4 12 24 29 ST228-MRSA-I
18 Slovenia Slovenia (PFGE A) 47 16 5 228 I TIMBMDMGMK t001 Southern Germany 1 4 1 4 12 24 29 ST228-MRSA-I
20 Slovenia Slovenia (PFGE E) 48 17 5 222 I TIMBMDMGMK t001 1 4 1 1 4 24 29 ST222-MRSA-I
M307-I UK UK EMRSA-3 49 18 E1 5 5 I TIMBMDMGMK t001 UK EMRSA-3 1 4 1 4 12 1 10 ST5-MRSA-I
97S101
(95/1119/3)Belgium Belgium Epidemic Clone 3a 50 19 C1 5 5 III TMDMGMK t045 1 4 1 4 12 1 10 ST5-MRSA-III
98S46 Belgium Belgium Epidemic Clone 3b 50 19 5 5 N TMDMGMK t045 1 4 1 4 12 1 10 ST5-MRSA-NT
61974-II Finland Finland E1 51 20 C2 5 5 II TJMBMDMGMK t002 New York / Japan 1 4 1 4 12 1 10 ST5-MRSA-II
30 Slovenia Slovenia (PFGE G) 52 21 5 5 I TIMBMDMGMMMK t178 EMRSA-3 1 4 1 4 12 1 10 ST5-MRSA-I
MR1 Poland Poland (PFGE F1) 79 32 5 5 I TJMBMDMGMB t053 EMRSA-3 1 4 1 4 12 1 10 ST5-MRSA-I
97121 France France - Strain B (PFGE VIc)** 1 1 H1 8 8 IV YHGFMBQBLO t008 EMRSA-2, -6 3 3 1 1 4 4 3 ST8-MRSA-IV
97151 France France (PFGE VIc) 1 1 8 8 IV YGFMBQBLO t024 EMRSA-2, -6 3 3 1 1 4 4 3 ST8-MRSA-IV
920 France France (PFGE VIb) 2 1 8 8 IV YHGFMBQBLO t008 EMRSA-2, -6 3 3 1 1 4 4 3 ST8-MRSA-IV
96158 France France (PFGE VId) 3 1 8 8 IV YHGFMBQBLO t008 EMRSA-2, -6 3 3 1 1 4 4 3 ST8-MRSA-IV
97120 France France (PFGE VIb) 4 1 8 8 IV YHGFMBQBLO t008 EMRSA-2, -6 3 3 1 1 4 4 3 ST8-MRSA-IV
97392 France France (PFGE (VIIa) 5 1 8 8 IV YHGFMBQBLO t008 EMRSA-2, -6 3 3 1 1 4 4 3 ST8-MRSA-IV
1966/97 Germany Hannover IIIc 6 2 8 254 N YGFMBQBLQBLPO t009 3 32 1 1 4 4 3 ST254-MRSA-NT
97S96 (82) Belgium Belgium Epidemic Clone 1a 7 3 A1 8 247 I nk NT EMRSA-5, -7, Iberian 3 3 1 12 4 4 16 ST247-MRSA-I
134/93 Germany N. German I 7 3 A3 8 247 I YHFGFMBQBLO t051 EMRSA-5, -7, Iberian 3 3 1 12 4 4 16 ST247-MRSA-I
54518 Finland Finland E7 8 3 A2 8 247 I YHFGFMBQBLO t051 EMRSA-5, -7, Iberian 3 3 1 12 4 4 16 ST247-MRSA-I
1450/94 Germany N. German Ia 8 3 8 247 I YHFGFMBQBLO t051 EMRSA-5, -7, Iberian 3 3 1 12 4 4 16 ST247-MRSA-I
N8-3756/90 Sweden Sweden - Strain I** 8 3 8 247 I YHFGFMBQBLO t051 EMRSA-5, -7, Iberian 3 3 1 12 4 4 16 ST247-MRSA-I
5 Spain Spain E1 8 3 A1 8 247 I YHFGFMBQBQBLO t200 EMRSA-5, -7, Iberian 3 3 1 12 4 4 16 ST247-MRSA-I
97S97 Belgium Belgium Epidemic Clone 1a 9 3 8 247 I YHFGFMBQBLO t051 EMRSA-5, -7, Iberian 3 3 1 12 4 4 16 ST247-MRSA-I
234/95 Germany N. German Ib 10 3 8 247 I YHFGFMBQBLO t051 EMRSA-5, -7, Iberian 3 3 1 12 4 4 16 ST247-MRSA-I
1869/98 Germany N. German Id 11 3 8 247 I YHGFMBQBLO t008 EMRSA-5, -7, Iberian 3 3 1 12 4 4 16 ST247-MRSA-I
HS 2 Sweden Sweden - Strain I** 12 3 8 247 I YFGFMBQBLO t052 EMRSA-5, -7, Iberian 3 3 1 12 4 4 16 ST247-MRSA-I
97S98
(95/5101/1) BelgiumBelgium Epidemic Clone 1b
13 38 247 I
nk NTEMRSA-5, -7, Iberian 3 3 1 12 4 4 16 ST247-MRSA-I
97118 France France (PFGE VIIe) 13 3 8 247 I YHFGFMBQBLO t051 EMRSA-5, -7, Iberian 3 3 1 12 4 4 16 ST247-MRSA-I
BM10827 France France (PFGE A) 13 3 8 247 I YHFGFMBQBLO t051 EMRSA-5, -7, Iberian 3 3 1 12 4 4 16 ST247-MRSA-I
BM10828 France France - Strain C (PFGE A)** 13 3 A6 8 247 I YHFGFMBQBLO t051 EMRSA-5, -7, Iberian 3 3 1 12 4 4 16 ST247-MRSA-I
62176 Finland Finland E10 14 3 A4 8 247 I YHFGFMBQBLO t051 EMRSA-5, -7, Iberian 3 3 1 12 4 4 16 ST247-MRSA-I
BM10882 France France (PFGE A) 15 3 8 247 I YHFGFMBQBLO t051 EMRSA-5, -7, Iberian 3 3 1 12 4 4 16 ST247-MRSA-I
97393 France France (PFGE VIIf) 16 3 8 247 I YHFGFMBQBLO t051 EMRSA-5, -7, Iberian 3 3 1 12 4 4 16 ST247-MRSA-I
406/98 Germany N. German Ic1 16 3 8 247 I YHFGFMBQBLO t051 EMRSA-5, -7, Iberian 3 3 1 12 4 4 16 ST247-MRSA-I
408/98 Germany N. German Ic2 16 3 8 247 I YHFGFMBQBLO t051 EMRSA-5, -7, Iberian 3 3 1 12 4 4 16 ST247-MRSA-I
80 Slovenia Slovenia (PFGE E) 17 3 8 247 I YHFGFMBQBLO t051 EMRSA-5, -7, Iberian 3 3 1 12 4 4 16 ST247-MRSA-I
95035 France France (PFGE VIId) 18 3 8 8 IV YHGFMBQBLO t008 EMRSA-2, -6 3 3 1 1 4 4 3 ST8-MRSA-IV
97117 France France (PFGE VIId) 18 3 8 8 IV YHGFMBQBLO t008 EMRSA-2, -6 3 3 1 1 4 4 3 ST8-MRSA-IV
162France
France - Strain A (PFGE VIId)**18 3 A5
8 8 Nnk NT
3 3 1 1 4 4 3 ST8-MRSA-NT
54511 Finland Finland E6 19 3 8 247 I YHFGFMBQBLO t051 EMRSA-5, -7, Iberian 3 3 1 12 4 4 16 ST247-MRSA-I
62396 Finland Finland E2 20 4 8 247 I YHFGFMBQBLO t051 EMRSA-5, -7, Iberian 3 3 1 12 4 4 16 ST247-MRSA-I
98 Slovenia Slovenia (PFGE C) 21 4 8 247 I YHFGFMBQBLO t051 EMRSA-5, -7, Iberian 3 3 1 12 4 4 16 ST247-MRSA-I
98541 Finland Finland E24 22 5 G1 8 241 III WGKAOMQ t037 2 3 1 1 4 4 30 ST241-MRSA-III
NCTC11939 UK UK EMRSA-1 23 6 J1 8 239 N WGKAOMQ t037 2 3 1 1 4 4 3 ST239-MRSA-NT
3680 Greece Greece-1 24 7 F1 8 239 III WGKAOMQ t037 EMRSA-1, -4, -11, Por 2 3 1 1 4 4 3 ST239-MRSA-III
3717 Greece Greece-1b 25 7 8 239 III WGKAOMQ t037 EMRSA-1, -4, -11, Por 2 3 1 1 4 4 3 ST239-MRSA-III
ACCEPTED
on August 30, 2018 by guest
http://jcm.asm
.org/D
ownloaded from
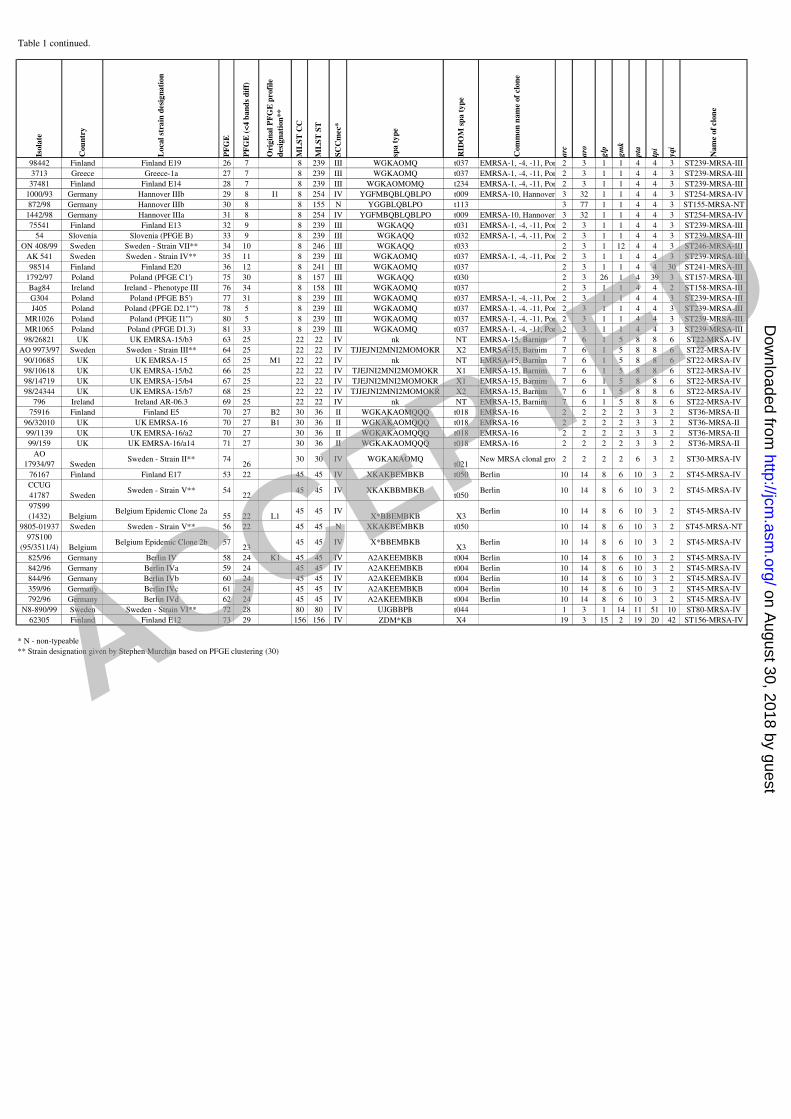
Table 1 continued.Is
ola
te
Cou
ntr
y
Local
stra
in d
esi
gn
ati
on
PF
GE
PF
GE
(<
4 b
an
ds
dif
f)
Ori
gin
al
PF
GE
profi
le
desi
gn
ati
on
**
ML
ST
CC
ML
ST
ST
SC
Cm
ec*
spa
ty
pe
RID
OM
sp
a t
yp
e
Com
mon
nam
e of
clo
ne
arc
aro
glp
gm
k
pta
tpi
yqi
Nam
e of
clo
ne
98442 Finland Finland E19 26 7 8 239 III WGKAOMQ t037 EMRSA-1, -4, -11, Por 2 3 1 1 4 4 3 ST239-MRSA-III
3713 Greece Greece-1a 27 7 8 239 III WGKAOMQ t037 EMRSA-1, -4, -11, Por 2 3 1 1 4 4 3 ST239-MRSA-III
37481 Finland Finland E14 28 7 8 239 III WGKAOMOMQ t234 EMRSA-1, -4, -11, Por 2 3 1 1 4 4 3 ST239-MRSA-III
1000/93 Germany Hannover IIIb 29 8 I1 8 254 IV YGFMBQBLQBLPO t009 EMRSA-10, Hannover 3 32 1 1 4 4 3 ST254-MRSA-IV
872/98 Germany Hannover IIIb 30 8 8 155 N YGGBLQBLPO t113 3 77 1 1 4 4 3 ST155-MRSA-NT
1442/98 Germany Hannover IIIa 31 8 8 254 IV YGFMBQBLQBLPO t009 EMRSA-10, Hannover 3 32 1 1 4 4 3 ST254-MRSA-IV
75541 Finland Finland E13 32 9 8 239 III WGKAQQ t031 EMRSA-1, -4, -11, Por 2 3 1 1 4 4 3 ST239-MRSA-III
54 Slovenia Slovenia (PFGE B) 33 9 8 239 III WGKAQQ t032 EMRSA-1, -4, -11, Por 2 3 1 1 4 4 3 ST239-MRSA-III
ON 408/99 Sweden Sweden - Strain VII** 34 10 8 246 III WGKAQQ t033 2 3 1 12 4 4 3 ST246-MRSA-III
AK 541 Sweden Sweden - Strain IV** 35 11 8 239 III WGKAOMQ t037 EMRSA-1, -4, -11, Por 2 3 1 1 4 4 3 ST239-MRSA-III
98514 Finland Finland E20 36 12 8 241 III WGKAOMQ t037 2 3 1 1 4 4 30 ST241-MRSA-III
1792/97 Poland Poland (PFGE C1') 75 30 8 157 III WGKAQQ t030 2 3 26 1 4 39 3 ST157-MRSA-III
Bag84 Ireland Ireland - Phenotype III 76 34 8 158 III WGKAOMQ t037 2 3 1 1 4 4 2 ST158-MRSA-III
G304 Poland Poland (PFGE B5') 77 31 8 239 III WGKAOMQ t037 EMRSA-1, -4, -11, Por 2 3 1 1 4 4 3 ST239-MRSA-III
J405 Poland Poland (PFGE D2.1'''') 78 5 8 239 III WGKAOMQ t037 EMRSA-1, -4, -11, Por 2 3 1 1 4 4 3 ST239-MRSA-III
MR1026 Poland Poland (PFGE I1''') 80 5 8 239 III WGKAOMQ t037 EMRSA-1, -4, -11, Por 2 3 1 1 4 4 3 ST239-MRSA-III
MR1065 Poland Poland (PFGE D1.3) 81 33 8 239 III WGKAOMQ t037 EMRSA-1, -4, -11, Por 2 3 1 1 4 4 3 ST239-MRSA-III
98/26821 UK UK EMRSA-15/b3 63 25 22 22 IV nk NT EMRSA-15, Barnim 7 6 1 5 8 8 6 ST22-MRSA-IV
AO 9973/97 Sweden Sweden - Strain III** 64 25 22 22 IV TJJEJNI2MNI2MOMOKR X2 EMRSA-15, Barnim 7 6 1 5 8 8 6 ST22-MRSA-IV
90/10685 UK UK EMRSA-15 65 25 M1 22 22 IV nk NT EMRSA-15, Barnim 7 6 1 5 8 8 6 ST22-MRSA-IV
98/10618 UK UK EMRSA-15/b2 66 25 22 22 IV TJEJNI2MNI2MOMOKR X1 EMRSA-15, Barnim 7 6 1 5 8 8 6 ST22-MRSA-IV
98/14719 UK UK EMRSA-15/b4 67 25 22 22 IV TJEJNI2MNI2MOMOKR X1 EMRSA-15, Barnim 7 6 1 5 8 8 6 ST22-MRSA-IV
98/24344 UK UK EMRSA-15/b7 68 25 22 22 IV TJJEJNI2MNI2MOMOKR X2 EMRSA-15, Barnim 7 6 1 5 8 8 6 ST22-MRSA-IV
796 Ireland Ireland AR-06.3 69 25 22 22 IV nk NT EMRSA-15, Barnim 7 6 1 5 8 8 6 ST22-MRSA-IV
75916 Finland Finland E5 70 27 B2 30 36 II WGKAKAOMQQQ t018 EMRSA-16 2 2 2 2 3 3 2 ST36-MRSA-II
96/32010 UK UK EMRSA-16 70 27 B1 30 36 II WGKAKAOMQQQ t018 EMRSA-16 2 2 2 2 3 3 2 ST36-MRSA-II
99/1139 UK UK EMRSA-16/a2 70 27 30 36 II WGKAKAOMQQQ t018 EMRSA-16 2 2 2 2 3 3 2 ST36-MRSA-II
99/159 UK UK EMRSA-16/a14 71 27 30 36 II WGKAKAOMQQQ t018 EMRSA-16 2 2 2 2 3 3 2 ST36-MRSA-II
AO
17934/97 SwedenSweden - Strain II** 74
2630 30 IV WGKAKAOMQ
t021New MRSA clonal grou 2 2 2 2 6 3 2 ST30-MRSA-IV
76167 Finland Finland E17 53 22 45 45 IV XKAKBEMBKB t050 Berlin 10 14 8 6 10 3 2 ST45-MRSA-IV
CCUG
41787 SwedenSweden - Strain V** 54
2245 45 IV XKAKBBMBKB
t050Berlin 10 14 8 6 10 3 2 ST45-MRSA-IV
97S99
(1432) BelgiumBelgium Epidemic Clone 2a
55 22 L145 45 IV
X*BBEMBKB X3Berlin 10 14 8 6 10 3 2 ST45-MRSA-IV
9805-01937 Sweden Sweden - Strain V** 56 22 45 45 N XKAKBEMBKB t050 10 14 8 6 10 3 2 ST45-MRSA-NT
97S100
(95/3511/4) BelgiumBelgium Epidemic Clone 2b 57
2345 45 IV X*BBEMBKB
X3Berlin 10 14 8 6 10 3 2 ST45-MRSA-IV
825/96 Germany Berlin IV 58 24 K1 45 45 IV A2AKEEMBKB t004 Berlin 10 14 8 6 10 3 2 ST45-MRSA-IV
842/96 Germany Berlin IVa 59 24 45 45 IV A2AKEEMBKB t004 Berlin 10 14 8 6 10 3 2 ST45-MRSA-IV
844/96 Germany Berlin IVb 60 24 45 45 IV A2AKEEMBKB t004 Berlin 10 14 8 6 10 3 2 ST45-MRSA-IV
359/96 Germany Berlin IVc 61 24 45 45 IV A2AKEEMBKB t004 Berlin 10 14 8 6 10 3 2 ST45-MRSA-IV
792/96 Germany Berlin IVd 62 24 45 45 IV A2AKEEMBKB t004 Berlin 10 14 8 6 10 3 2 ST45-MRSA-IV
N8-890/99 Sweden Sweden - Strain VI** 72 28 80 80 IV UJGBBPB t044 1 3 1 14 11 51 10 ST80-MRSA-IV
62305 Finland Finland E12 73 29 156 156 IV ZDM*KB X4 19 3 15 2 19 20 42 ST156-MRSA-IV
* N - non-typeable
** Strain designation given by Stephen Murchan based on PFGE clustering (30)
ACCEPTED
on August 30, 2018 by guest
http://jcm.asm
.org/D
ownloaded from

Table 2. Resolution of typing methods for MRSA
No. of % isolates most Index of diversity (DI)
Typing method types frequent type (95% CI)
PFGE 81 4.1 99.50 (99.11-99.88)
PFGE (≤3 bands diff.) 34 24.5 91.94 (88.27-95.61)
MLST 17 23.5 88.72 (85.78-91.66)
MLST + SCCmec 24 24.7 89.46 (86.32-92.59)
spa sequencing 27 19.8 91.31 (90.82-91.79)
CI - Confidence Interval
ACCEPTED
on August 30, 2018 by guest
http://jcm.asm
.org/D
ownloaded from

Table 3. PFGE and spa sequence typing subdivisions of MLST / SCCmec clones
Clone No. of isolates PFGE type(Frequency) spa sequence type(Frequency)
ST247-MRSA-I 23 3(21); 4(2) t051(18); t008(1); t052(1); t200(1); nd(2)
ST228-MRSA-I 13 13(8); 14(2); 15(2); 16(1) t001(10); t188(1); t201(1); nd(1)
ST239-MRSA-III 12 7(5): 5(2); 9(2); 11(1): 31(1); 33(1) t037(9); t031(1); t032(1); t234(1)
ST45-MRSA-IV 9 24(5); 22(3); 23(1) t004(5); t050(2); X3(2)
ST8-MRSA-IV 8 1(6); 3(2) t008(7); t024(1)
ST22-MRSA-IV 7 25(7) X1(2); X2(2); nd(4)
ST36-MRSA-II 4 27(4) t018(4)
ST5-MRSA-I 3 18(1); 21(1); 32(1) t001(1); t053(1); t178(1)
ST241-MRSA-III 2 5(1); 12(1) t037(2)
ST254-MRSA-IV 2 8(2) t009(2)
ACCEPTED on August 30, 2018 by guest
http://jcm.asm
.org/D
ownloaded from





























