Embed Size (px)
Citation preview

Indian Journal of Experimental Biology Vol. 43, April 2005, pp. 335-341
Evidence for presence of female produced pheromone components in male scent brush extract of castor semi-looper moth Achaea janata L.
K N Jyothi, A L Prasuna & A R Prasad
Pheromone Laboratory, Organic Division I, Indian Institute of Chemical Technology, Tarnaka , Hyderabad 500 007, India.
Received 4 October 2004; revised 3 January 2005
Hexane extract of male terminalia (along with scent brushes) of castor semi-looper moth, Achaea janata L, elicited significant olfactory responses in both male and female insects by electroantennogram recording technique. However, male extract in the wind tunnel evoked noticeable behaviour responses in the female insects only. Orientation response of the males to the male extract was not evident in wind tunnel experiments. Two electrophysiologically-active compounds were identified from the male extract. Based on GC retention times and mi'!ss spectrometry the two compounds were confirmed as (Z, Z)-9, 12-octadecadienal and (Z, Z, Z)-3, 6, 9-heneicosatriene. These two compounds are also constituents of female produced fourcomponent blend of A. janata.
Keywords: Achaea janata, Electroantennogram, , Gas chromatography, (Z, Z, Z)-3, 6, 9-Heneicosatriene Male brush, (Z, Z)-9, 12-0ctadecadienal,Wind tunnel.
Achaea janata L, Castor semi-looper (Lepidoptera: Noctuidae) moth is an important pest of castor, Ricinus communis in India. Both caterpillars and adult moths cause the damage. In severe infestation, larvae consume entire foliage leaving only the midribs and stalks. Excessive loss of foliage causes reduction in seed yield. Adults are fruit sucking moths and cause serious damage to citrus crop.
Persoons et aI. l identified the sex pheromone system in female semi-looper moth as a fourcomponent blend consisting of --{I) Heneicosane; (II) (Z, Z)-6, 9-Heneicosadiene; (III) (Z, Z, Z)-3, 6, 9-Heneicosatriene and (IV) (Z, Z)-9, 12-0ctadecadienal in the ratio of l.0: l.0: 60 -70: l.0, respectively.
Generally there is overwhelming behavioural and electrophysiological evidence that female moths are non-responsive to the sex pheromones which they produce2
. However, our electroantennogram studies indicated that females are stimulated not only by the total pheromone blend, but also by each of the individual pheromone components (Unpublished observations, - Jyothi K N). Similar responsiveness of females to female produced pheromone has also been demonstrated in Trichoplusia ni 2,3 and Choristonuera jumiferana4
.
Correspondent author: Tel. No. 27193136 E-maillO: [email protected] nCT communication No. 041219
Electrophysiological studies of olfactory sensitivities to pheromones in insects are mostly concentrated on antennal receptor neurons of males, particularly on their sensitivity to female produced male attracting pheromones. Pheromones produced by males and their role in close range complex sexual interplay have been relatively neglected5
.
Males of many species of Lepidoptera have scent organs located on the abdomen, thorax, legs and wings, which vary from simple scales and hair tufts to complex eversible structures. The noctuid male scent organs commonly referred to as hair pencils typically constitute a pair of brushes, scent glands and storage pockets6
• The male scent brush apparatus of Ajanata also possesses typical Noctuid morphology. The brushes extend and spread to a fan shape when a male approaches a calling female. Sehgal et al. 7,8 have described the morphological details of the external genitalia of Ajanata in detail. Although, volatile chemicals from males of some of the noctuid species have been identified and biosynthetic pathways for some of the compounds were known, the actual role of most of the compounds is not clearly elucidated9
.
Aim of the present investigation was--{l) to ascertain the electrophysiological and behavioural function of the hairbrush extract against both males and females of Ajanata; and (2) to isolate and identify the chemical nature of secretions from male abdominal tip extract (along with brushes) which may

336 INDIAN J EXP BIOL, APRIL 2005
possibly explain the reason for female-female attraction.
Materials and Methods Insect culture--Culture of Ajanata was maintained
in the laboratory in an environmental chamber at 27°± 1 °C, 70 % RH and 10: 14 dark/light under reverse photoperiod conditions. The larvae were fed on fresh castor leaves until pupation. Pupae were segregated sex wise (based on the morphology of 8-10 abdominal segments) and maintained in cubical wooden adult cages (36 x 36cm x 36cm). The moths after emergence were fed on 10 % honey solution soaked on cotton pads in petri dishes.
Male extracts (abdominal terminalia along with brushes) preparation-For extract preparation, 3-5 day old male moths were utilized. Male terminalia were everted by gently pressunzmg terminal abdominal segments during the period of mating activity 10. With the help of micro scissors they were quickly excised into vials containing n-hexane. The hexane extracts were pooled and stored at -300 C until further use. The extracts were filtered through a short silica column and were concentrated at room temperature with a stream of nitrogen for chemical analysis, electrophysiological and wind tunnel bioassay studies.
Analytical procedures
Gas chromatography (GC)-Gas chromatography of male extract and standard compounds were performed on a Carlo Erba 5300 gas chromatograph fitted with a flame ionization detector and split injector. Separations were performed on a 2m x 2.5 mm id glass column packed with 3% CPSil-5 chromo sorb phase (100-120 mesh). A temperature programme of 50°C (2min delay) increased to final temperature of 260°C (5 min delay) at the rate of 10°C/min was followed. Nitrogen was used as the carrier gas with a column head pressure of 100 kPa. Injector and detector temperatures were set at 275°C. The column effluent was split to send 1/6th of the extract to FlD detector and 5/6th to the glass capillary embedded in dry ice for collection of volatiles ( a custom made device of TNO, The Netherlands gifted to nCT) . Two min fractions were collected in chilled capillaries and subsequently bio-assayed by EAG technique.
Gas chromatography-mass spectrometry (GC-MS) -The extract was subjected to GC-MS instrument for analysis. Agilent 6890 GC with 5973 NMSD with
capillary column HP-5 MS (0.25mm id; 30 m length at 0.25 f.Lm film thickness) was used. Ionization voltage was 70 e V and the temperature of the ion source and the interface was 2300 and 2800 C, respectively. Carrier gas was helium at a constant flow rate of 1.2ml/min. The extract was analyzed using the same temperature program as described above for Gc. The identity of the various peaks was confirmed by comparing the unknowns with both the standard spectra library and authentic samples (gift from TNO, The Netherlands).
Chemicals For identification and electrophysiological work,
two compounds-(Z,Z)-9,12 octadecadienal and (Z,Z,Z)-3,6,9-heneicosatriene were obtained from Organic Division, nCT. Detailed synthesis procedures are described elsewhere 1 I.
Electroantennogram recording technique (EAG) The EAG technique was employed to study the
sensitivity/selectivity of both male and female antennae of Ajanata to the crude male scent brush extract as well as the synthetic components. An EAG profile indicated the senSItIvIty and relative abundance of olfactory receptor neurons present on the antenna for a particular compound tested.
For electrophysiological recordings, Syntech EAG (Syntech, Hilversum, The Netherlands) was employed. The isolated antennae of different ages of males and females Ajanata were individually mounted onto a metal electrode holder (stainless steel) with the help of an electrically conductive gel. The antenna was continuously flushed with charcoal filtered and moistened air stream through a stainless tube (8mm id) ending 2 cm before the antennal preparation. The EAG signals were amplified and recorded with a data acquisition controller and software (Syntech, The Netherlands).
The test compound (5 male equivalents of male extract/5-100 f.L g of synthetic compound) was spread on a filter paper. After complete evaporation of the solvent, the filter paper strip was inserted into a Pasteur pipette. Stimuli were provided by connecting the pipette for 300 ms into the air stream flushing over the antenna. An equal volume of solvent alone (hexane) spread on the filter paper served as the control. Control stimulation was made at the beginning and after every 2-3 EAG recordings. EAG responses were evaluated by measuring the maximum amplitude of depolarization triggered by stimuli. At

JYOTHI et al.: PRESENCE OF FEMALE PHAROMORE COMPONENTS IN MALE 337
least 30 sec was allowed between two continuous stimuli for recovery of the antenna. _ EAG responses were recorded from five males and five females moths individually to the extract/synthetic compound. The EAG data in Figs. 2-4, represent corrected EAG figures (male extract/ synthetic compound minus control). The EAG responses were subjected to analysis of variance (ANOV A).
Wind-tunnel bioassay
An indoor cylindrical wind tunnel (30 cm diam x 2 m) was used to test the biological activity of crude extract/synthetic standards 12. The tunnel was housed in a room with temperature and relative humidity controls and an exhaust system from the tunnel to the outside of the room. The test conditions were maintained at 26° ± 2°C, RH 65-70 % and wind speed of 0.45 mlsec. For odour delivery, a piece of filter paper (2cm - Whatman International Ltd, England» was used as the release substrate. All the experiments were carried out during dark period with 3-5 day old males and females at 22 hr (ie., during their normal mating activity time). Males and females were tested in groups (2-5 insects/experiment). The following criteria were used to judge the behavioural response. For each experiment, 20 insects were used and the insects were used only once. Observations were made with the help of a torch light covered with red filter.
Crete ria to judge the behavioural response
• Activation characterized by. vigorous antennal movement and wing vibration
• Upwind orientation exhibiting typical zig zag flight characteristic of male moths
• Oriented upwind flight and arrival at the middle of the tunnel
• Completed flight • Source contact
Individual males were transferred into 2/4cm meshed plastic containers before their placement in the wind tunnel. The test compound loaded on the filter paper served as pheromone dispenser at the upwind section of the wind tunnel. The container holding the male moths and the pheromone source were placed at 12 cm height at the down -and upwind ends in the wind tunnel, respectively. After 2-5 min, when test moths settled to the conditions of the wind tunnel, the odour source was placed in the tunnel. The lid was then removed and the moths were observed for flight behaviour. Males were used only once.
Two kinds of stimuli were used in the wind tunnel. In the first test, the orientation behaviour of males and females separately towards the crude extract (five male equivalents) was evaluated. Filter paper loaded with same volume of hexane served as control. In the second test, the behavioural response of compounds identified from the male extract i.e., [Z, Z, Z)-3, 6, 9-heneicosatriene and (Z, Z) - 9, 12-octadecadienal were evaluated. The test compound (100 I1g of each) dissolved in n-hexane was used as the stimulus. Numbers of males exhibiting different levels Of upwind flight behaviour in the tunnel were counted. All the experiments were performed in triplicate.
Results
Isolation and identification of active compounds from male extract
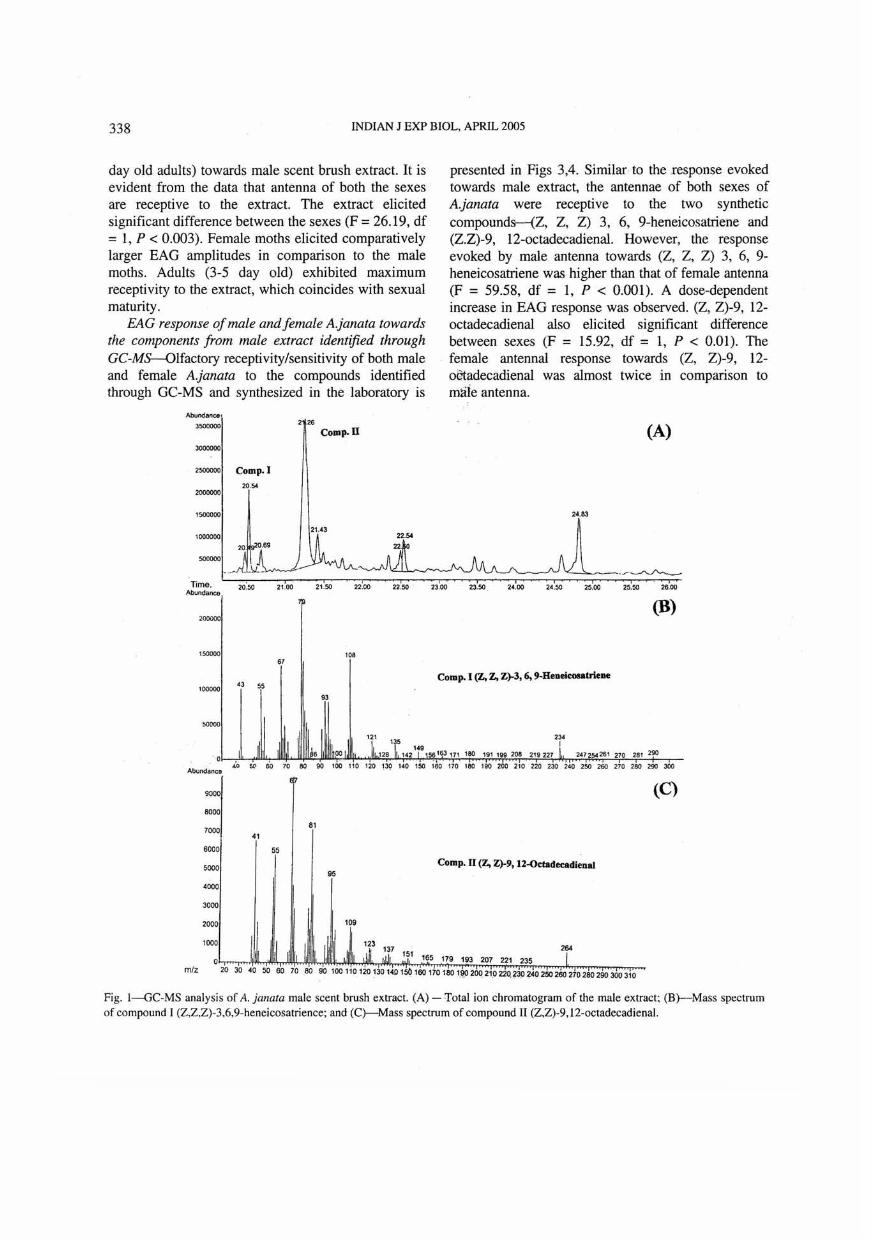
GC and GC-MS analysis-Gas chromatogram of 3-5 day old male extract is depicted in Fig. 1. In the chart depicting total ion chromatogram (lA), three main peaks and some trace peaks are evident. Based on the retention time and spectral analysis, the first peak was identified as hexadecanoic acid. The GC effluent of second peak with retention time 20.54 collected through chilled capillaries when tested for EAG activity elicited good EAG response against both males and females. The mass spectrum of this peak (lB) with characteristic signals at mlz=99 , mlz=108, mlz=234 and a molecular ion at mlz=290, is identical with the spectrum of (Z, Z, Z) 3, 6, 9-heneicosatriene. Earlier, we have identified this as the main component from female ovipositor extract of A. janata1
- The mass spectrum of third peak (IC) with strong molecular ion mlz 264 and evidence for two double bonds (mlz 67, 81 and 95) suggested an octadecadienal structure. The peak had an identical retention time on all columns to that of (Z, Z)-9, 12-octadecadienal synthesized 'in laboratory and was confmned as (Z, Z)-9, 12- octadecadienal through comparative GC-MS and electrophysiological studies. The trace peaks were identified as hydrocarbons whose mass spectra corresponded to tricosane and pentacosane, respectively which did not elicit any electrophysiological activity against both males and females.
Electroantennography Electroantennographic response of male and
female A.janata to male extract-Figure 2 depicts the EAG response of male and female A.janata (one-six
338 INDIAN J EXP BIOL, APRIL 2005
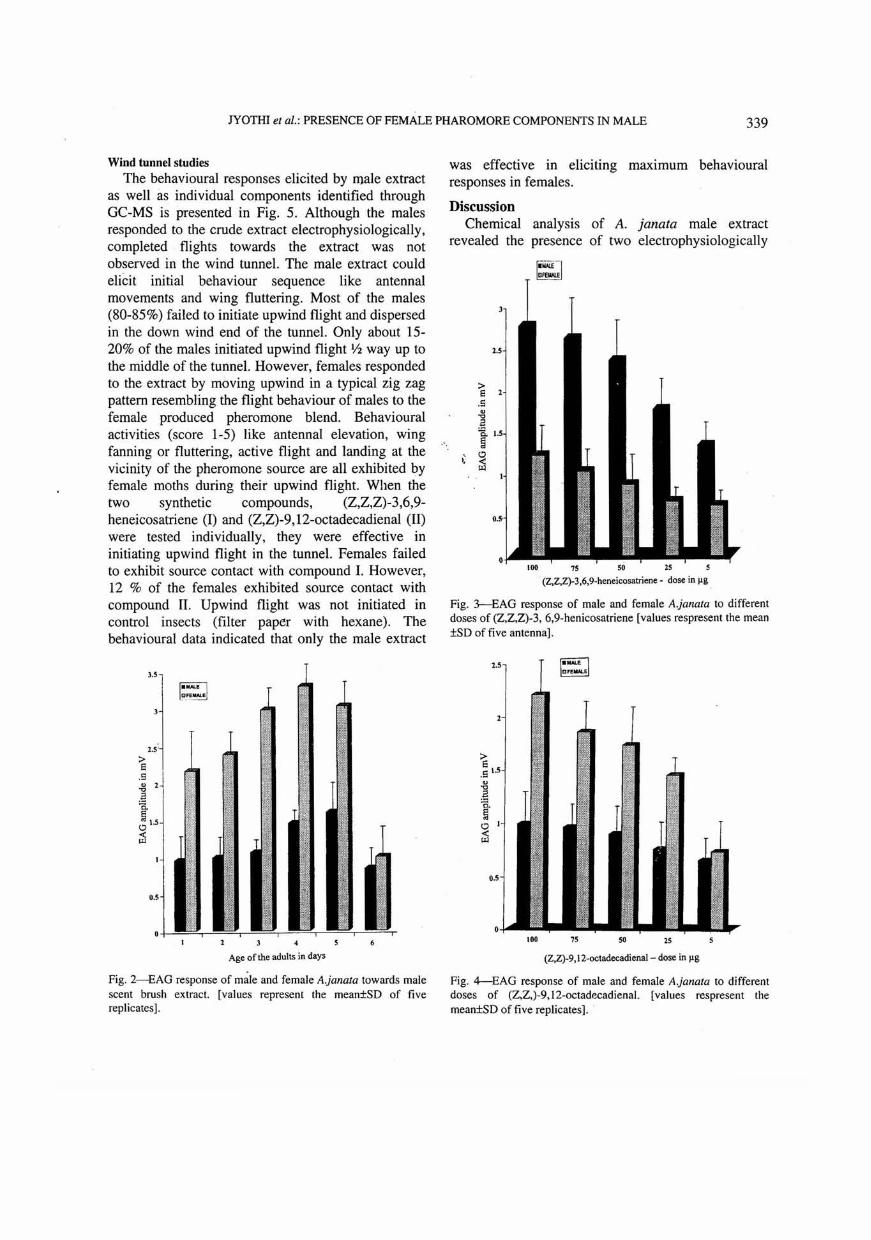
day old adults) towards male scent brush extract. It is evident from the data that antenna of both the sexes are receptive to the extract. The extract elicited significant difference between the sexes (F = 26.19, df = 1, P < 0.003). Female moths elicited comparatively larger EAG amplitudes in comparison to the male moths. Adults (3-5 day old) exhibited maximum receptivity to the extract, which coincides with sexual maturity.
EAG response of male andfemale Ajanata towards the components from male extract identified through GC-M5--0lfactory receptivity/sensitivity of both male and female Ajanata to the compounds identified through GC-MS and synthesized in the laboratory is
2 26
Comp.n
21 .43
presented in Figs 3,4. Similar to the response evoked towards male extract, the antennae of both sexes of Ajanata were receptive to the two synthetic compounds-{Z, Z, Z) 3, 6, 9-heneicosatriene and (Z.Z)-9, 12-octadecadienal. However, the response evoked by male antenna towards (Z, Z, Z) 3, 6, 9-heneicosatriene was higher than that of female antenna (F = 59.58, df = 1, P < 0.001). A dose-dependent increase in EAG response was observed. (Z, Z)-9, 12-octadecadienal also elicited significant difference between sexes (F = 15.92, df = 1, P < 0.01). The female antennal response towards (Z, Z)-9, 12-oetadecadienal was almost twice in comparison to rruiie antenna.
(A)
24.83
21 .50 22.00 22.50 . 23.00 23.50 . 24.00 24.50 25.00 25.50 26.00
108
93
Compo I (z, Z, Z}-3, 6, 9-HeDeicosatrieoe
234 I
(B)
149 128 142 156 163 171 180 191 199 208 219227 247254261 270 281 290
Abundance 40 5(> 60 70 80 90 100 110 120 130 140 150 160 170 180 190 200 210 220 230 240 250 260 270 280 290 JOO
1'/7 9000 (C) 8000
7000 81
41
6000 55
5000 95
Compo U (z, Z}-9, 12-OctadecadieDai
4000
3000
2000 109
II l000! .I. 1.~3 137 2f
o I, II/JI di'll! 1,1,11, 1,1.11 151 165 179 193 207 221 235 !.
m/z 20 30 40 5060 70 80 90 100 110 120 130 14D 150 180 170 180 1.!1O 2DQ21.0 22D.230 240 250 280 270280290 300 31 0
Fig. I-GC-MS analysis of A. janata male scent brush extract. (A) - Total ion chromatogram of the male extract; (B)--Mass spectrum of compound I (Z,Z,Z)-3,6,9-heneicosatrience; and (C)--Mass spectrum of compound II (Z,Z)-9,12-octadecadienal.
JYOTHI et al.: PRESENCE OF FEMALE PHAROMORE COMPONENTS IN MALE 339
Wind tunnel studies The behavioural responses elicited by male extract
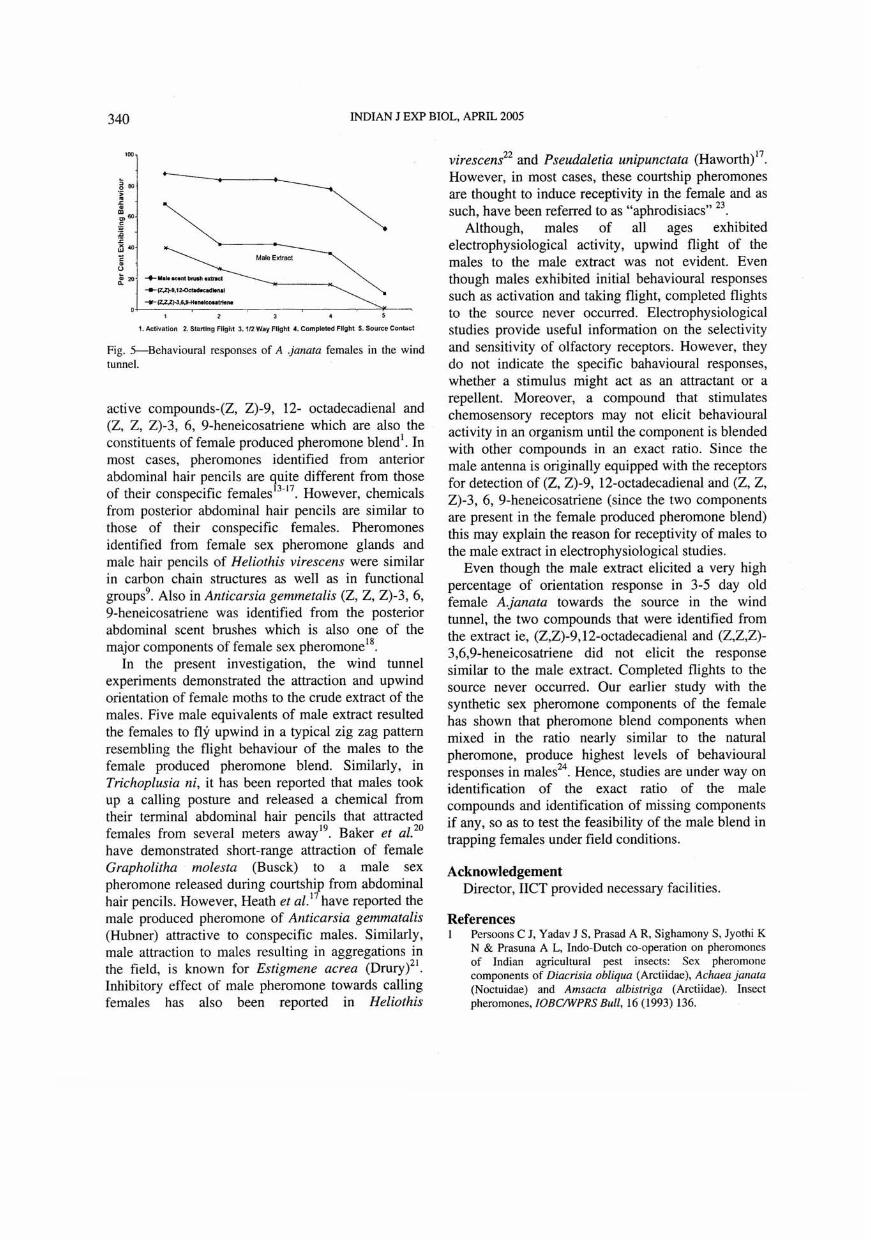
as well as individual components identified through GC-MS is presented in Fig. 5. Although the males responded to the crude extract electrophysiologically, completed flights towards the extract was not observed in the wind tunnel. The male extract could eliCit initial behaviour sequence like antennal movements and wing fluttering. Most of the males (80-85%) failed to initiate upwind flight and dispersed in the down wind end of the tunnel. Only about 15-20% of the males initiated upwind flight 'h way up to the middle of the tunnel. However, females responded to the extract by moving upwind in a typical zig zag pattern resembling the flight behaviour of males to the female produced pheromone blend. Behavioural activities (score 1-5) like antennal elevation, wing fanning or fluttering, active flight and landing at the vicinity of the pheromone source are all exhibited by female moths during their upwind flight. When the two synthetic compounds, (Z,Z,Z)-3,6,9-heneicosatriene (I) and (Z,Z)-9, 12-octadecadienal (II) were tested individually, they were effective in initiating upwind flight in the tunnel. Females failed to exhibit source contact with compound I. However, 12 % of the females exhibited source contact with compound II. Upwind flight was not initiated in control insects (filter paper with hexane). The behavioural data indicated that only the male extract
> E .5
" 2 -0 a i. §
<:] I.S
« ~
4
Age of the adults in days
Fig. 2-EAG response of male and female A.janata towards male scent brush extract. [values represent the mean±SD of five replicates] .
was effective in eliciting maximum behavioural responses in females.
Discussion Chemical analysis of A. janata male extract
revealed the presence of two electrophysiologically
100 7S 2S
. (Z,Z,Z)-3 ,6,9-heneicosatriene - dose in IIg .
Fig. 3--EAG response of male and female A.janata to different doses of (Z,Z,Z)-3, 6,9-henicosatriene [values respresent the mean ±SD of five antenna].
:> E .5
<:] « ~
100 7S so 2S
(Z,z)-9,12-octadecadienal- dose in IIg
Fig. 4--EAG response of male and female A.janata to different doses of (Z,Z,)-9,12-octadecadienal. [values respresent the mean±SD of five replicates]. .
340 INDIAN J EXP BIOL, APRIL 2005
100
-e-1Z,z)·t.12.oetod<cad1tnal
-¥- (z,z,z)-3,6.9-H.IMfcONtrieM
1. Activation 2. Starting Flight 3.1f2 Way Flight 4. Completed Flight 5. Source Contact
Fig. S--Behavioural responses of A .janata females in the wind tunneL
active compounds-(Z, Z)-9, 12- octadecadienal and (Z Z Z)-3 6 9-heneicosatriene which are also the
" " I constituents of female produced pheromone blend. In most cases, pheromones identified from anterior abdominal hair pencils are quite different from those of their conspecific females I3
.17
• However, chemicals from posterior abdominal hair pencils are similar to those of their conspecific females. Pheromones identified from female sex pheromone glands and male hair pencils of Heliothis virescens were similar in carbon chain structures as well as in functional groupS9. Also in Anticarsia gemmetalis (Z, Z, Z)-3, 6, 9-heneicosatriene was identified from the posterior abdominal scent brushes which is also one of the major components of female sex pheromone l8
.
In the present investigation, the wind tunnel experiments demonstrated the attraction and upwind orientation of female moths to the crude extract of the males. Five male equivalents of male extract resulted the females to fly upwind in a typical zig zag pattern resembling the flight behaviour of the males to the female produced pheromone blend. Similarly, in Trichop1usia ni, it has been reported that males took up a calling posture and released a chemical from their terminal abdominal hair pencils that attracted
19 ak 120 females from several meters away . B er et a . have demonstrated short-range attraction of female Grapholitha molesta (Busck) to a male sex pheromone released during courtship from abdominal hair pencils. However, Heath et a1. 1
? have reported the male produced pheromone of Anticarsia gemmatalis (Hubner) attractive to conspecific males. Similarly, male attraction to males resulting in aggregations in the field, is known for Estigmene acrea (Drury)21. Inhibitory effect of male pheromone towards calling females has also been reported in Heliothis
viresceni2 and Pseuda1etia unipunctata (Haworth) I?
However, in most cases, these courtship pheromones are thought to induce receptivity in the female and as
" h d" " 23 such, have been referred to as ap ro ISlacs . Although, males of all ages exhibited
electrophysiological activity, upwind flight of the males to the male extract was not evident. Even though males exhibited initial behavioural responses such as activation and taking flight, completed flights to the source never occurred. Electrophysiological studies provide useful information on the selectivity and sensitivity of olfactory receptors. However, they do not indicate the specific bahavioural responses, whether a stimulus might act as an attractant or a repellent. Moreover, a compound that stimulates chemosensory receptors may not elicit behavioural activity in an organism until the component is blended with other compounds in an exact ratio. Since the male antenna is originally equipped with the receptors for detection of (Z, Z)-9, 12-octadecadienal and (Z, Z, Z)-3, 6, 9-heneicosatriene (since the two components are present in the female produced pheromone blend) this may explain the reason for receptivity of males to the male extract in electrophysiological studies.
Even though the male extract elicited a very high percentage of orientation response in 3-5 day old female A.janata towards the source in the wind tunnel, the two compounds that were identified from the extract ie, (Z,Z)-9,12-octadecadienal and (Z,Z,Z)-3,6,9-heneicosatriene did not elicit the response similar to the male extract. Completed flights to the source never occurred. Our earlier study with the synthetic sex pheromone components of the female has shown that pheromone blend components when mixed in the ratio nearly similar to the natural pheromone, produce highest levels of behavioural
24 d' d responses in males . Hence, stu les are un er way on identification of the exact ratio of the male compounds and identification of missing components if any, so as to test the feasibility of the male blend in trapping females under field conditions.
Acknowledgement Director, IICT provided necessary facilities.
References 1 Persoons C J, Yadav J S, Prasad A R, Sighamony S, Jyothi K
N & Prasuna A L, Indo-Dutch co-operation on pheromones of Indian agricultural pest insects: Sex pheromone components of Diacrisia obliqua (Arctiidae), Achaea janata (Noctuidae) and Amsacta albistriga (Arctiidae). Insect pheromones, IOBC/wPRS Bull, 16 (1993) 136.

JYOTill et al.: PRESENCE OF FEMALE PHAROMORE COMPONENTS IN MALE 341
2 Light D M & Birch M C, Electrophysiological basis for the behavioural responses of male & female Trichoplusia ni to synthetic female pheromone, J Insect Physiol, 25 (1979) 161.
3 Seabrook W D, Linn C E, Dyer L J & Shorey H H, Comparison of electroantennograms from female and male cabbage looper moths (Trichoplusia ni) of different ages and for various pheromone concentrations, J Chem Eco!, 13 (1987) 1443.
4 Palaniswamy P & Seabrook W D, Behavioural responses of male eastern spruce bud worm, C.fumiferana (Lepidoptera:Tortricidae) to the sex pheromone of her own species, J Chem Ecol, 4 (1978) 649 .
5 Birch M C & Heftez A, Extrusible organs in male moths and their role in courtship behaviour, Bull Entomol Soc Am, 33 (1987) 222 .
6 Birch M C & Poppy G M, Scents and eversible scent structures of male moths, Annu Rev Entomol, 35 (1990) 25.
7 Sehgal S S, Neelam Sharma & Bhatti U S, Structure of external male genitalia of castor semi looper, Achaea janata (LINN), Bull Ent, 20 (1979 a) 8.
8 Sehgal S S, Bhatti U S & Anita D, Functional morphology and homology of musculature of external male genitalia of Achaeajanata (LINN), Bull Ent, 20 (1979 b) 12.
9 Teal PEA & Tumlinson J H, Isolation, identification and biosynthesis of compounds produced by male hair pencil glands of Heliothis virescens (F) (Lepidoptera : Noctuidae), J Chem Ecol, 15 (1989 ) 413.
10 Prasuna A L, Jyothi K N, Sighamony S, Prasad A R & Yadav J S, Activity patterns of Achaea janata L; Recording of insect actometer, MWDRD, Indian J Exp Bioi, 35 (1997) IllS.
11 Yadav J S, Rajasekhar K, Venkatram Reddy & Chandrasekhar S, Practical synthesis of pheromone components of Achaea janata (Noctuidae), Synth Commun, 28 (22) (1998 ) 4249.
12 Jyothi K N, Prasuna A L, Sighamony S, Prasad A R & Yadav J S, SEM of pheromone gland of Arctiid moth, Diacrisia obliqua Walker--evidence from EAG and windtunnel bio-assay, Indian Natl Sci Acad (B), 63 (1997) 421.
13 Alpin R T & Birch M C, Pheromones from the abdominal brushes of male noctuid Lepidoptera, Nature, 217 (1968) 1167.
14 Alpin R T & Birch M C, Identification of odorous compounds from male Lepidoptera, Experientia, 26 (1970) 1193.
15 Fitzpatrick S M, Miller D, Weatherston I, & NcNeil J N, Determination of pheromone content of hair pencils from individual males of Pseudaletia unipunctata (Haw) (Lepidoptera: Noctuidae), J Chem Ecol, II ( 1981) 207 .
16 Hirai K, Shorey H H & Gatson K K, Competition between courting male moths: Male-male inhibitory pheromone, Science, 202 (1978) 644.
17 Hirai K, Male scent emitted by armyworm, Pseudaletia unipunctata and P. separata (Lepidoptera: Noctuidae), Appl Entomol Zool, 15 (1980) 310.
18 Heath R R, Landolt P J, Leppla N C & Dueben B D, Identification of a male produced pheromone of Anticarsia gemmatalis (Hubner) (Lepidoptera: Noctuidae) attractive to con specific males, J Chem Eco!, 14 ( 1988) 1121.
19 Landolt P J & Heath R R, Attraction of female cabbage looper moths (Lepidoptera: Noctuidae) to male produced pheromone, Ann Entomo! Soc Am, 8 (1989) 520.
20 Baker T C, Nishida R & Roelofs W L, Close range attraction of female oriental fruit moths to herbal scent of mail hair pencils, Science, 214 (1981 ) 1359.
21 Willis M A & Birch M C, Male lek formation and female calling in a population of the arctiid moth Estigmene acrea, Science, . 218 (1982) 168.
22 Hendricks D E & Shaver T N, Tobacco budworm: Male pheromone suppressed emission of pheromone by the female, Environ Entomol, 4 (1975) 555.
23 Birch M C, Aphrodisiac pheromones in insects, in Pheromones, edited by MC Birch (American Elsevier, New York) 1974, 115
24 Krishnakumari, Prasuna A L, Jyothi K N, Valli M Y, Sighamony S, Prasad A R & Yadav J S, Behavioural and electrophysiological responses of Achaea janata Linn. (Lepidoptera: Noctuidae) males to synthetic female produced sex pheromone components, J Entomol Res, 22 (1988 ) 197.





























