Embed Size (px)
Citation preview

Evolution of resistance to white pine blisterrust in high-elevation pines
Mike Antolin, Stuart Field, Jen Klutsch, Anna Schoettle and Simon Tavener
Colorado State University
Rocky Mountain Research Station, USDA Forest Service
Thanks: NSF award 0734267, USDA Forest Service
Evolution of resistance – p. 1/74

The basic ecology
Evolution of resistance – p. 2/74

High-elevation White Pines
• Habitat: dry, exposed, high altitude sites (Rocky Mtns.)
• Occupies habitat other species cannot
• Long-lived species
• Keystone species
3

Threats to HEWPs
White pine blister rustClimate change
Mountain Pine Beetle
4

White Pine Blister Rust
• Cronartium ribicola
• Non-native fungal pathogen
• Lethal in HEWP
• Spreading despite extensive control efforts
5

Effects of WPBR
1

WPBR Life Cycleaeciospores
basidiospores
Two Obligate Hosts
• Ribes spp.• Spring• Sub-lethal
• HEWP• Fall• Lethal
6

B = Basic model
Evolution of resistance – p. 3/74

B: Basic n-stage model
Consider a stage-structured population model in the form of a nonlinear map
x{n}(t+ 1,p) = h{n}(x{n}(t,p),p)
x{n}(0,p) = x{n},0
where
h{n} : Rn × Rm 7→ R
n.
Notation
• The subscript {n} is used to denote the size of the system.
• x{n}(t,p) is the solution vector containing xi, the density of individuals
(individuals per hectare) in stage i = 1, . . . , n at times t = 0, 1, . . . , T
(years).
• p is the vector of parameters pj , j = 1, . . . ,M .
Nonlinear o.d.e. models are analogous.Evolution of resistance – p. 4/74

B: Order of operations
Assumption: Fecundity occurs after survivorship & transition.
Consequently,
x{n}(t+ 1) = g{n}(x{n}(t)) + f{n}
(g{n}(x{n}(t))
)
where f denotes fecundity and g denotes survivorship and transition.
Note: When considering a genetic model, a splitting between survivorship &
transition and fecundity arises naturally since survivorship & transition occurswithin genotypes while fecundity occurs across genotypes.
Note: For simplicity, we drop the explicit dependence of solution x{n}(t) on
parameters p.
Evolution of resistance – p. 5/74

Notation
We will use
• x to denote the outcome of survivorship & transition,
• y as the vector on which fecundity operates,
• x or y to denote intermediate quantities.
Assumption: Disease status and genotype affect the arguments (inputs) of thefunctions modeling survivorship & transition and fecundity.
Evolution of resistance – p. 6/74

D = Disease
Evolution of resistance – p. 7/74

D: Incorporating disease
Assumption: There are only susceptible and infected populations.
Let
x{2n} =
xS{n}
xI{n}
,
and
g{2n}(x{2n}) =
g{n}(xS{n})
g{n}(xI{n})
.
White bark pines do not recover from blister rust infection
Evolution of resistance – p. 8/74

D: Survivorship & transition
Assumption: The effect of disease can be described in terms of a linear
weighting applied to the argument of the function modeling survivorship &transition.
Let the cost of infection to survivorship & transition be
Cν{2n} =
I{n} 0
0 Cν{n}
, Cν{n} = diag(ci), i = 1, . . . , n ,
and define
x{2n} = g{2n}
(C
ν{2n} ∗ x{2n}
)=
g{n}(xS{n})
g{n}(Cν{n} ∗ xI
{n})
Evolution of resistance – p. 9/74

D: Fecundity
Fecundity is modeling through the action of the non-linear vector valued
function
f{2n} : R2n 7→ R2n,
but
f{2n} 6=
f{n}(xS{n})
f{n}(xI{n}).
Think vertical vs horizontal transmission.
Both susceptible and infected white bark pine produce susceptible seeds.
(Furthermore, seeds cannot become infected.)
Evolution of resistance – p. 10/74

D: Fecundity
Assumption: The effect of disease on fecundity can be described as a
weighting applied to the argument of the nonlinear function used to modelfecundity.
Let the cost of fecundity be
Cϕ
{2n} =
I 0
0 Cϕ
{n}
.
and define fecundity in the SI model to be
f{2n}
(C
ϕ
{2n} ∗ x{2n}
).
Evolution of resistance – p. 11/74

D: Infectivity
Assumption: Infection occurs after survivorship, transition and fecundity and
occurs at a constant rate.
Let
B{2n} =
I{n} −B{n} 0
B{n} I{n}
,
where
B{n} = diag(bi), i = 1, . . . , n
and I{n} is the n× n identity matrix.
Evolution of resistance – p. 12/74

D: SI disease model
Finally, the full SI disease model is the nonlinear map
x{2n}(t+ 1) = B{2n} ∗(x{2n} + f{2n}(y{2n})
),
where
x{2n} = g{2n}
(C
ν{2n} ∗ x{2n}
),
and
y{2n} = Cϕ
{2n} ∗ x{2n}.
Evolution of resistance – p. 13/74

D: SI disease model
The cost of disease on survival, transition, the cost of disease on fecundity and
infectivity could all be nonlinear.
The SI disease model would then become
x{2n}(t+ 1) = B{2n}
(x{2n} + f{2n}(y{2n})
),
where
x{2n} = Cν{2n}
(x{2n}
),
y{2n} = Cϕ
{2n}
(x{2n}
).
Conceptually this is no more complicated.
Evolution of resistance – p. 14/74

P = Pine model
Evolution of resistance – p. 15/74

P: Variables
The basic pine model has six stages or classes,
x{6} =
SEEDS
SD1
SD2
SA
YA
MA
=
x1
x2
x3
x4
x5
x6
.
Units: Individuals / hectare
Evolution of resistance – p. 16/74

P: The nonlinear map
Consistent with the general framework, we define the nonlinear map
x{6}(t+ 1) = g{6}(x{6}(t)) + f{6}(g{6}(x{6}(t)))
where f and g represent fecundity and survivorship & transition respectively.
Units: Time is measured in years.
Model structure and coefficient values is the result of extensive field studies.
Evolution of resistance – p. 17/74

P: Nonlinear seedling recruitment
Assumption: Seedling recruitment occurs before survivorship & transition
The modeling of the germination process involves a number of auxiliary
quantities, including properties of the surrounding forest, and the foraging andseed storing behaviors of the Clark’s nutcracker.
• Leaf Area Index (LAI)
• Seed caching
• Pfind
• . . .
Evolution of resistance – p. 18/74

P: Seedling recruitment. Leaf Area Index
Leaf Area Index (LAI) is used as a proxy for tree density and is defined as a
function of tree size as measured by the diameter at breast height.
Let
larea1= larea2
= 0, larea3= α1, lareai
= α2 (di)α3 , i = 4, 5, 6,
where
α1 = 0.456, α2 = 0.117, α3 = 1.925 ,
and the diameter at breast height (dbh) measurements for the six classes are
d1 = 0, d2 = 0, d3 = 0, d4 = 2.05, d5 = 12.5, d6 = 37.
Secondary seedlings have d3 = 0 but do possess leaf area, thus ~larea3is
calculated separately. Let
LAI(x6) =(larea, x6)
10000.
Evolution of resistance – p. 19/74

P: Seedling recruitment. Clark’s nutcracker
We define
SpB(x{6}) =x1
nBirds,
rcache(x{6}) =0.73
1 + exp((31000− SpB(x{6}))/3000
) + 0.27 ,
rALs(x{6}) =1
1 + exp(2 (LAI(x{6})− 3)),
where x1 is the number of seeds, and finally,
r2(x{6}) =
[(1− Pfind) (1− Pcons)
SpC
]rcache(x{6}) rALs(x{6}) .
The number of new seedlings is then given by
γ{6}(x{6}) = r2(x{6}) ∗ x1 ∗ e2 ,
where e2 ∈ R6 is the unit vector with a single nonzero entry in the second
position.Evolution of resistance – p. 20/74

P: Seedling recruitment
We define the intermediate vector
x{6} = x{6} + γ{6}(x{6}),
where γ{6}(x{6}) is the result of the nonlinear seedling recruitment process.
Evolution of resistance – p. 21/74

P: Linear survivorship & transition
Let
S{6} =
1 0 0 0 0 0
0 s2 0 0 0 0
0 t2 s3 0 0 0
0 0 t3 s4 0 0
0 0 0 t4 s5 0
0 0 0 0 t5 s6
.
The coefficients along the diagonal of S{6} are
s2 = 0.636, s3 = 0.8391, s4 = 0.9310, s5 = 0.9653, s6 = 0.995 .
Coefficients along the sub-diagonal of S{6} are
t2 = 0.212, t3 = 0.0559, t4 = 0.0490, t5 = 0.0197 .
Evolution of resistance – p. 22/74

P: Survivorship & transition
We define the outcome of both nonlinear and linear survivorship and transition
as
x{6} = g{6}(x{6}) = S{6} ∗ x{6}.
Evolution of resistance – p. 23/74

P: Fecundity
We define the nonlinear function
f{6} : R6 × ρ 7→ R6
where ρ is a measure of the effect of class on fecundity.
The modeling of the fecundity process involves a number of auxiliaryquantities.
• Maximum number of cones per tree
• Number of seeds per cone
• . . .
Evolution of resistance – p. 24/74

P: Fecundity
Seed production is determined by the number of seeds per cone, which is
determined as
Ctree(x{6}) =
[0.5
1 + exp(5 (LAI(x{6})− 2.25))+ 0.5
]Cmax
and
r1(x{6}) = Scone ∗ Ctree(x{6}).
The number of seeds produced is then given by
f{6} = r1(x{6}) ∗ (ρx5 + x6) ∗ e1
where e1 ∈ R6 is a unit vector with a single nonzero entry in the first position.
Evolution of resistance – p. 25/74

P: The nonlinear map
Finally
x{6}(t+ 1) = x{6} + f{6}(x{6}).
Evolution of resistance – p. 26/74

PD = Pine + disease
Evolution of resistance – p. 27/74

PD: Variables
We double the number of classes and define
x{12} =
xS{6}
xI{6}
=
SEEDS
susceptible SD1
susceptible SD2
susceptible SA
susceptible YA
susceptible MA
0
infected SD1
infected SD2
infected SA
infected YA
infected MA
=
x1
x2
x3
x4
x5
x6
x7
x8
x9
x10
x11
x12
.
Evolution of resistance – p. 28/74

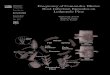
€
1) Seedling recruitment : r2
x {n}( )
€
2) Survival& transition : g {n}
€
3) Seed production : f {n}
€
4) Infection : B{n}
Fall
Spring

SEED
SD1
SD2SP
YA
MA
iSD1
iSD2
iSPiYA
iMA
Arrows:Survival & transitionInfectionFecundity
Nodes:SusceptibleInfected

PD: Seedling recruitment
Assumption: Seedling recruitment occurs before survivorship & transition.
We define
x{12} =
xS{6}
xI{6}
+
γD{6}(x{12})
0
where the nonlinear function γD{6} is the modification of γ{6}.
Both susceptible and diseased individuals provide shading.
Evolution of resistance – p. 29/74

PD: Survivorship & transition
Let
S{12} = diag(S{6},S{6}).
The cost of infection on survival & transition is given by
Cν{12} = diag(I{6},C
ν{6})
where
Cν{6} = diag(0, c2, . . . , c6),
with the cost of infection related to tree size, specifically
c2 = 0.01, c3 = 0.13, ci = 1− exp(−δ di), i = 4, 5, 6.
Evolution of resistance – p. 30/74

PD: Survivorship & transition
We define
x{12} = g{12}(x{12})
= S{12} ∗(C
ν{12} ∗ x{12}
)
=
S{6} 0
0 S{6}
∗
I{6} 0
0 Cν{6}
∗
xS{6}
xI{6}
=
S{6} 0
0 S{6}Cν{6}
∗
xS{6}
xI{6}
.
Evolution of resistance – p. 31/74

PD: Fecundity
Assumptions:
• Infection is not transmitted vertically. Susceptible and infected adults
produce susceptible seeds.
• Fecundity is modeled via cone production only because we assumefertilization occurs via a “pollen cloud", with each individual contributing
equally to the cloud. Thus pollen is not limiting and differences in fecunditycan be seen as a result of differences in available resources allocated to
cone production.
• Pollen production is unaffected by infection because the pollen producingbranches primarily compose the lower crown, while the seed cone
producing branches dominate the upper crown. Infectiondisproportionately affects the upper crown (top-kill), thus mitigating the
effect of infection on pollen relative to cone production.
Evolution of resistance – p. 32/74

PD: Fecundity
We define the nonlinear function
f{12} : R12 × (ρ, Cϕ)⊤ 7→ R12
where
• ρ is a measure of the effect of class on fecundity,
• Cϕ is a measure of the effect of disease on fecundity.
Evolution of resistance – p. 33/74

PD: Fecundity
Let y{12} = x{12} and
yS = ρy5 + y6, yI = ρy11 + y12,
and
z = yS + CϕyI ,
so that
f{12}(x{12}; ρ,Cϕ) = r1(x{12}) ∗ z ∗ e1,
where the function r1(x{12}) is an obvious extension of r1(x{6}). Thinkshading.
Evolution of resistance – p. 34/74

PD: Infectivity
Assumption: Infectivity occurs at a constant rate which may vary according to
age class.
We define
B{12} =
I{6} −B{6} 0
B{6} I{6}
where
B{6} = diag(0, b2, b3, b4, b5, b6)
and
b2 = b3 = b4 = b5 = b6 = 0.044.
Evolution of resistance – p. 35/74

PD: The nonlinear map
Assumption: Disease transmission occurs after fecundity.
Finally
x{12}(t+ 1) = B{12} ∗(x{12} + f{12}(x{12}; ρ, C
ϕ)),
where
x{12} = g{12}(x{12}).
Evolution of resistance – p. 36/74

Sensitivity analysis
Evolution of resistance – p. 37/74

Sensitivity wrt parameters
Using index notation,
xi(t+ 1,p) = hi(x(t,p), p),
xi(0) = zi
}
i = 1, . . . , n
Differentiating with respect to parameters, pk, gives
∂xi(t+ 1)
∂pk=
∂hi
∂xm
∗∂xm(t)
∂pk+
∂hi
∂pk
∂xi(0)
∂pk= 0
i = 1, . . . , n,
k = 1, . . . ,K .
Elasticities are relative sensitivities,
Ei,k =pkxi
∂xi
∂pk.
Evolution of resistance – p. 38/74

Sensitivity wrt initial conditions
To determine stability with respect to the initial conditions, we differentiate with
respect to the initial conditions to give
∂xi(t+ 1)
∂zk=
∂hi
∂xm
∗∂xm(t)
∂zk+
∂hi
∂zk
∂xi(0)
∂zj= δij
i = 1, . . . , n,
k = 1, . . . ,K ,
To solve this for the population and its stability with respect to parameters andinitial conditions we evolve all equations simultaneously.
Perform calculations using the MATLAB / MAPLE package SENSAI⋆.⋆Methods in Ecology and Evolution, 2, pp 560–575, (2011).
Evolution of resistance – p. 39/74

Results
Evolution of resistance – p. 40/74

Model Results IRegeneration from 1000 SD1
• Stages 2-6
• Eqm = 626
• No disease (β = 0)
0 200 400 600 800
0200
400
600
800
1000
1200
1400
time
No.
Indi
vidu
als
SD1SD2SAYAMA
Equilibrium Structure
14

Model Results II
0 200 400 600 800
0100
200
300
400
time
No.
Indi
vidu
als
SD1SD2SAYAMA
Equilibrium Structure: susceptible
0 200 400 600 800
050
100
150
200
250
300
350
time
No.
Indi
vidu
als
SD1SD2SAYAMA
Equilibrium Structure: infected
19
• Introduce rust into Eqm population
• Infection scenario (β = 0.044)

The Beta Effect
0 50 100 150 200
050
100
150
200
250
300
350
time
No.
Indi
vidu
als
! = 0.2
0 50 100 150 200
050
100
150
200
250
300
350
time
No.
Indi
vidu
als
! = 0
SD1SD2SAYAMA
0 50 100 150 200
050
100
150
200
250
300
350
time
No.
Indi
vidu
als
! = 0.016
0 50 100 150 200
050
100
150
200
250
300
350
time
No.
Indi
vidu
als
! = 0.044
20
0 0.016
0.0440.2

Beta vs. Delta
Beta
0.00
0.05
0.10
0.15
0.20
delta
0.00
0.05
0.10
0.15
0.20
0.25
0.30
Total Trees - 100 yrs 500
1000
1500
2000
2500
Beta
0.00
0.05
0.10
0.15
0.20
delta
0.00
0.05
0.10
0.15
0.20
0.25
0.30
Mature Adults - 100 yrs 0
100
200
300
25
Mature AdultsTotal Trees
100 years

Disease free sensitivities and elasticities
Mature adults and total population.Regenerating population after 100 years and equilibrium population.
Evolution of resistance – p. 41/74

Elasticities during transience
Elasticities for β = 0.044 after 100 years.
Mature adult population and total population.
Evolution of resistance – p. 42/74

Infectivity vs Prevalence of disease
Stage-specific and overall prevalence of blister rust for low (β = 0.016),
medium (β = 0.044) and high (β = 0.20) probability of infection.
Evolution of resistance – p. 43/74

Conclusions I
• Sustainability of HEWP stands infected with WPBR depends on twodominant effects:
(a) Infection probability
(b) Regeneration mediated by competition (e.g. LAI)
Parameters controlling these effects disproportionately remove smallerstages via infection induced mortality and reduced seedling establishment.
• Stand structure
Diseased equilibrium stands are less dominated by mature adults thandisease-free equilibrium stands.
• Disease prevalence
(a) Low prevalence in SD1 is due to high cost of infection, high natural
mortality and low residence time, not low susceptibility. This mayaccount for low seedling rust prevalence found in field surveys.
(b) High prevalence in mature adults due to low infection cost and low
natural mortality. Consistent with field observations.Evolution of resistance – p. 44/74

Conclusions II
• Transient vs equilibrium sensitivity
(a) 100 years after onset of infection, MA are sensitive to mortality and
infection probabilities of all stages
(a) At equilibrium, MA are sensitive to mortality and infection probabilitiesof the MA stage only
Evolution of resistance – p. 45/74

What insights can modeling provide?
Simulation
• Reinterpret existing observations
• Understand change in the structure of the forest
• Understand time scale for loss of forest ( » observation time)
• Understand time scale for control strategies ( » observation time)
• Predict behaviors of planted stands vs mature forests
Transient sensitivities
• Identify important parameters to estimate
• Identify important parameters to seek to control
• Equilibrium sensitivities are somewhat moot
Evolution of resistance – p. 46/74

Control strategies
Control efforts
1. Eradicate ribes†
2. Prune infected branches†
3. Select and breed rare resistant individuals
What insights might a genetic model provide?
• Understand time scale for spread of a resistant allele
• Estimate the effect of potential control strategies on pine stands
† = job creation schemeEvolution of resistance – p. 47/74

SLG = Single locus genetics
Evolution of resistance – p. 48/74

Single locus genetics
Assumption: Consider two distinct alleles at a single locus only.
Let
x{3n} =
xAA{n}
xaA{n}
xaa{n}
.
Survivorship & transition occur independently within each genotype, hence
g{3n}(x{3n}) =
g{n}(xAA{n})
g{n}(xaA{n})
g{n}(xaa{n})
.
Evolution of resistance – p. 49/74

SLG: Fecundity
Random mating is applied through the action of the nonlinear vector-valued
function
f{3n} : R3n 7→ R3n.
Evolution of resistance – p. 50/74

SLG: The nonlinear map
Assumption: Genotype affects survivorship & transition through fitness
function V{3n} : R3n 7→ R3n and affects fecundity through fitness function
W{3n} : R3n 7→ R3n.
Putting it all together,
x{3n}(t+ 1) = x{3n} + f{3n}(y{3n})
where
x{3n} = g{3n}
(V{3n}(x{3n})
)
y{3n} = W{3n}(x{3n}).
Evolution of resistance – p. 51/74

DSLG = Disease and single locus genetics
Evolution of resistance – p. 52/74

Disease and single-locus genetics
Combining our earlier ideas we define
x{6n} =
xAA{2n}
xaA{2n}
xaa{2n}
.
Evolution of resistance – p. 53/74

DSLG: Survivorship & transition
We extend g{2n} to create g{6n} : R6n 7→ R6n as
g{6n} =
g{2n}
g{2n}
g{2n}
.
Assumption: Genotype can influence the cost of disease on survivorship &transition.
Cν{6n} = diag
(C
ν,AA
{2n} ,Cν,aA
{2n} ,Cν,aa
{2n}
)
= diag(I{n},C
ν,AA
{n} , I{n},Cν,aA
{n} , I{n},Cν,aa
{n}
).
Evolution of resistance – p. 54/74

DSLG: Fecundity
We define
f{6n} : R6n 7→ R6n
to model random mating.
Assumption: Genotype can influence the effect of disease on fecundity.
Let
Cϕ
{6n} = diag(C
ϕ,AA
{2n} ,Cϕ,aA
{2n} ,Cϕ,aa
{2n}
)
= diag(I{n},C
ϕ,AA
{n} , I{n},Cϕ,aA
{n} , I{n},Cϕ,aa
{n}
).
Evolution of resistance – p. 55/74

DSLG: Infectivity
Assumption: Infectivity is independent of genotype.
Let
B{6n} = diag(B{2n},B{2n},B{2n}
).
Evolution of resistance – p. 56/74

DSLG: The nonlinear map
Putting it all together,
x{6n}(t+ 1) = B{6n} ∗(x{6n} + f{6n}(y{6n})
),
where
x{6n} = g{6n}(Cν{6n} ∗ x{6n})
and
y{6n} = Cϕ
{6n} ∗ x{6n}.
Evolution of resistance – p. 57/74

PDSLG = Pine + disease + single locusgenetics
Evolution of resistance – p. 58/74

PDSLG: Variables
Define x{36} as
x{36} =
xAA{12}
xaA{12}
xaa{12}
.
Recall that survivorship and transition occurs within each genotype.
Evolution of resistance – p. 59/74

PDSLG: Seedling recruitment
Assumption: Nonlinear seedling recruitment is independent of infection status
and genotype.
We define
x{12} = x{12} +
γDSLG{6} (x{36})
0
and
x{36} =
xAA{12}
xaA{12}
xaa{12}
.
Think shading.
Evolution of resistance – p. 60/74

PDSLG: Survivorship costs
Let
S{36} = diag(S{6}, S{6}, S{6}, S{6}, S{6}, S{6}),
and
Cν36 =
I6 0
... 0 0
... 0 0
0 I6... 0 0
... 0 0
. . . . . . . . . . . . . . . . . . . . . . . .
0 0
... I6 0
... 0 0
0 0
... 0 I6 − h ∗Cν6
... 0 0
. . . . . . . . . . . . . . . . . . . . . . . .
0 0
... 0 0
... I 0
0 0
... 0 0
... 0 I6 −Cν6
Evolution of resistance – p. 61/74

PDSLG: Survivorship & transition
Then
x{36} = g{36}
(x{36}
)
= S{36} ∗(C
ν{36} ∗ x{36}
).
Evolution of resistance – p. 62/74

PDSLG: Fecundity
We define the nonlinear function
f{36} : R36 × (ρ,Cϕ, h)⊤ 7→ R36
where
• ρ is a measure of the effect of class on fecundity
• Cϕ is a measure of the effect of disease on fecundity,
• h is the usual partial dominance parameter.
Evolution of resistance – p. 63/74

PDSLG: Pollen production
Assumptions
• Males do not have an infection status.
• The number of males linearly dependent on the number of females.
Let y = x{36} and
yAAmale = (y5 + y6) + (y11 + y12),
yaAmale = (y17 + y18) + (y23 + y24),
yaamale = (y29 + y30) + (y35 + y36),
male = yAAmale + yaA
male + yaamale,
so that
ppollen =yAAmale + 0.5 yaA
male
maleand qpollen =
0.5 yaAmale + yaa
male
male.
Evolution of resistance – p. 64/74

PDSLG: Females
Assumption: Stage has an effect on viability of females that is independent of
infection status and genotype.
Define
yAAS = ρy5 + y6, yAA
I = ρy11 + y12,
yaAS = ρy17 + y18, yaA
I = ρy23 + y24,
ySaa = ρy29 + y30, yaa
I = ρy35 + y36,
and
female = yAAS + yAA
I + yaAS + yaA
I + yaaS + yaa
I .
Evolution of resistance – p. 65/74

PDSLG: Egg viability
Assumption: Genotype affects egg viability of infected individuals.
Consider the effect of infection status and genotype on egg viability
zAA = yAAS + yAA
I ,
zaA = yaAS + (1− h ∗ (1− Cϕ) ∗ yaA
I ,
zaa = yaaS + Cϕ ∗ yaa
I .
Evolution of resistance – p. 66/74

PDSLG: Random mating
Random mating gives
f1(y36) = ppollen ∗ (zAA + 0.5 zaA),
f13(y36) = qpollen ∗ (zAA + 0.5 zaA) + ppollen ∗ (0.5 zaA + zaa),
f25(y36) = qpollen ∗ (0.5 zaA + zaa),
and
f{36} = f1(y36) ∗ e1 + f13(y36) ∗ e13 + f25(y36) ∗ e25.
Thus
f{36} = r1(y36) ∗ f36
where the function r1(x{36}) is an obvious extension of r1(x{6}).
Evolution of resistance – p. 67/74

PDSLG: Infectivity
Assumption: Infectivity is independent of genotype.
B{36} =
B{12} 0 0
0 B{12} 0
0 0 B{12}
.
Evolution of resistance – p. 68/74

PDSLG: The nonlinear map
Putting it all together,
x{36}(t+ 1) = B{36} ∗(x{36} + f{36}
(x{36}; ρ, C
ϕ, h))
.
Evolution of resistance – p. 69/74

Results
Evolution of resistance – p. 70/74

Preliminary studies
0 500 1000 15000
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1QoI vs Time
Time
QoI
LAIb=0, rsite=0.2LAIb=0, rsite=0.4LAIb=0, rsite=1LAIb=2, rsite=0.2LAIb=2, rsite=0.4LAIb=2, rsite=1
0 500 1000 15000
0.05
0.1
0.15
0.2
0.25
0.3
0.35
0.4
0.45
0.5QoI vs Time
Time
QoI
LAIb=0, rsite=0.2LAIb=0, rsite=0.4LAIb=0, rsite=1LAIb=2, rsite=0.2LAIb=2, rsite=0.4LAIb=2, rsite=1
Fixation of advantageous allele with and without background pollen
Evolution of resistance – p. 71/74

Preliminary studies
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
0 200 400 600 800 1000
5025
0
Time Step
Tota
l # T
rees
p = 0.005, LAI.b = 2, r.site=0.4, Beta = 0.044
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
0 50 100 150 200
010
0
Time Step
# of
Adu
lt Tr
ees Quantity of Interest
Parameter
Sen
sitiv
ity
−60
000
Sensitivity of # Adult Trees, TimeStep = 200
1 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31 33
Parameter
Ela
stic
ity
−3
02
Elasticity of # Adult Trees, TimeStep = 200
1 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31 33
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
0 200 400 600 800 1000
5025
0
Time Step
Tota
l # T
rees
p = 0.005, LAI.b = 2, r.site=0.4, Beta = 0.044
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●
●●●●●●●●●●●
●●●●●●●●●●
●●●●
0 50 100 150 200
0.00
0.06
Time Step
p (F
req.
of R
) Quantity of Interest
ParameterS
ensi
tivity
05
15
Sensitivity of p = f(R), TimeStep = 200
1 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31 33
Parameter
Ela
stic
ity
−8
−2
Elasticity of p = f(R), TimeStep = 200
1 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31 33
Number of adults Frequency of advantageous alleleModerate competition and low quality site.
Evolution of resistance – p. 72/74

Preliminary studies
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
0 200 400 600 800 1000
400
800
Time Step
Tota
l # T
rees
p = 0.005, LAI.b = 0, r.site=1, Beta = 0.044
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●
0 50 100 150 200
150
300
Time Step
# of
Adu
lt Tr
ees Quantity of Interest
Parameter
Sen
sitiv
ity
−40
0040
00
Sensitivity of # Adult Trees, TimeStep = 200
1 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31 33
Parameter
Ela
stic
ity
−12
−6
0
Elasticity of # Adult Trees, TimeStep = 200
1 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31 33
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
0 200 400 600 800 1000
400
800
Time Step
Tota
l # T
rees
p = 0.005, LAI.b = 0, r.site=1, Beta = 0.044
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●
●●●●●●●●●●●●
●●●●●●●●●●
●●●●●●●●●●
●●●●●●●●●●
0 50 100 150 200
0.00
0.15
Time Step
p (F
req.
of R
) Quantity of Interest
ParameterS
ensi
tivity
020
Sensitivity of p = f(R), TimeStep = 200
1 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31 33
Parameter
Ela
stic
ity
−6
−2
2Elasticity of p = f(R), TimeStep = 200
1 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31 33
Number of adults Frequency of advantageous alleleLow competition, high quality site.
Evolution of resistance – p. 73/74

Future directions
• Comprehensive study of single locus genetics including control strategies
• Multilocus genetics
1. Linkage disequilibrium for small numbers of loci and alleles
2. Recombination vs selection for small numbers of loci
3. Parametrization ???
Evolution of resistance – p. 74/74

Future directions
• Comprehensive study of single locus genetics including control strategies
• Multilocus genetics
1. Linkage disequilibrium for small numbers of loci and alleles
2. Recombination vs selection for small numbers of loci
3. Parametrization ???
There is something fascinating about science. One gets such wholesalereturns of conjecture out of such a trifling investment of fact. ... Mark Twain
Evolution of resistance – p. 74/74


























![Assessing the Impact of Blister Rust Infected Whitebark Pine in the Alpine Treelines of Glacier National Park and the Beartooth Plateau, U.S.A. [Emily Smith-Mckenna]](https://img.pdfslide.net/doc/110x75/555564c0b4c9052b208b56ae/assessing-the-impact-of-blister-rust-infected-whitebark-pine-in-the-alpine-treelines-of-glacier-national-park-and-the-beartooth-plateau-usa-emily-smith-mckenna.jpg)