Embed Size (px)
Citation preview

of September 25, 2018.This information is current as
CellsExpression and Function of CD300 in NK
Levi-Schaffer, Stipan Jonjic and Ofer MandelboimDikla Lankry, Hrvoje Simic, Yair Klieger, Francesca
http://www.jimmunol.org/content/185/5/2877doi: 10.4049/jimmunol.0903347July 2010;
2010; 185:2877-2886; Prepublished online 23J Immunol
MaterialSupplementary
7.DC1http://www.jimmunol.org/content/suppl/2010/07/23/jimmunol.090334
Referenceshttp://www.jimmunol.org/content/185/5/2877.full#ref-list-1
, 13 of which you can access for free at: cites 44 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2010 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on September 25, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on September 25, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
Expression and Function of CD300 in NK Cells
Dikla Lankry,* Hrvoje Simic,† Yair Klieger,* Francesca Levi-Schaffer,‡
Stipan Jonjic,† and Ofer Mandelboim*
The killing activity of NK cells is regulated by signals derived from inhibitory and activating NK cell receptors, including the
CD300 family of proteins. CD300a was reported to be expressed on all NK cells and to deliver an inhibitory signal upon binding to
a yet unknown ligand/s. The CD300a protein contains four ITIMs and is highly homologous to CD300c. Little is known about the
function and distribution of these two receptors and the identity of their ligand/s. In this article, we show that CD300a is indeed an
inhibitory receptor expressed by human NK cells, but surprisingly, we show that not all NK clones are inhibited in a CD300a-
dependent manner. We demonstrate, using a panel of 13 new anti-CD300a and CD300c Abs that we generated, that CD300a and
CD300c are indistinguishable on the surface of NK cells. Using mutational-analysis survey, we show that tyrosine 267 located in
the third ITIM motif of the CD300a protein is important for the inhibitory function of CD300a. The Journal of Immunology,
2010, 185: 2877–2886.
Natural killer cells are bone marrow-derived lymphocytesthat make up 5–15% of the PBLs (1, 2). NK cells areable to kill a broad spectrum of pathogens and tumors
without prior specific stimulation (3–5), although recent evidencesuggests that NK cells possess some adaptive properties. The NKcytotoxicity is controlled by multiple activating and inhibitory sur-face receptors and intracellular signal-transduction molecules (6–9). In humans, the major NK-triggering receptors include theNKp80, NKG2D, CD16, and the natural cytotoxic receptors, whichinclude NKp46, NKp44, and NKp30 (3, 6, 10–13).Inhibition of NK cell activity is mediated mainly by the two
major families ofMHC-specific inhibitory receptors, which includethe Ig superfamily receptors (killer Ig-related receptor [KIR] andleukocyte Ig-like receptor) and the C-type lectin (CD94/NKG2A)receptor superfamily (14–16). The different inhibitory receptorsshow diverse specificity and can discriminate between differentclass I MHC proteins. An important feature of all inhibitory NKreceptors that interact with MHC class I is that they are expressedin a variegated fashion so that each NK cell expresses multiplereceptors in a complex combinatorial repertoire (16).
The KIR gene family is divided, based on functional activity,into inhibitory and stimulating KIRs (17, 18). Products of in-hibitory KIR genes are characterized by long cytoplasmic tailsfeaturing ITIMs that, upon engagement, transmit inhibitory sig-nals. In contrast, stimulating KIRs have short cytoplasmic tailslacking ITIMs but instead they carry a positively charged aminoacid in the transmembrane region, which provides a docking sitefor the activating adaptor molecule DAP12 (17, 18). In addition toMHC class I binding receptors, other inhibitory receptors alsoexist. However, interestingly, as opposed to the class I inhibitoryreceptors, the inhibitory receptors that do not bind MHC class Iare sometimes expressed on all NK cells (9).The CD300 proteins are a family of leukocyte membrane reg-
ulatory molecules that modulate leukocyte responses. There areseven members in the family, named CD300a–g (19). The CD300aand CD300c receptors, which are the subject of this work, are cellsurface glycoproteins that are encoded by independent, but closelylinked, genes on human chromosome 17q 22–24. Both moleculesare members of the Ig superfamily and each contains a single V-like Ig domain, a membrane proximal region, a transmembraneregion, and a cytoplasmic region. The Ig domains of the CD300aand CD300c molecules show 80% identity at the amino acid level(20–23). The CD300a molecule has a long cytoplasmic tail of 100aa. This region contains four tyrosines, three of which are foundwithin the ITIM consensus sequences (20, 21). The CD300a cy-toplasmic region also contains a di-leucine motif that is associatedwith endocytosis and delivery to lysosomes (20, 22). The CD300cmolecule has a proline-rich membrane proximal region, a trans-membrane region containing a charged glutamic acid residue, anda short cytoplasmic tail of only 18 aa.The CD300a and CD300c receptors demonstrate several unique
characteristics. First, like the KIR gene family, they share a highdegree of homology but might have different functions (23),perhaps similar to the pairwised receptors (17, 18). Second, mostof the inhibitory receptors expressed by NK cells have one or twoITIM motifs at the cytoplasmic tail, with the exception of leu-kocyte Ig-like receptor 1 and CD300a, which have four ITIMmotifs (20). Thus, it will be interesting to investigate which ofthe ITIMs are important for CD300a function. Third, none of theactivating receptors identified contains a charged glutamic acidresidue in the transmembrane, suggesting that CD300c deliversits activating signals via signaling proteins other than those pre-
*Lautenberg Center for General and Tumor Immunology, BioMedical Research In-stitute, Hadassah Medical School and ‡Department of Pharmacology and Experimen-tal Therapeutics, Faculty of Medicine, School of Pharmacy, Hebrew University,Jerusalem, Israel; and †Department of Histology and Embryology, Center for Pro-teomics, Faculty of Medicine, University of Rijeka, Rijeka, Croatia
Received for publication October 15, 2009. Accepted for publication June 24, 2010.
This work was supported by Israeli Science Foundation and The Israeli Science Foun-dation (Morasha) Grants, a Croatia Israel Research Grant, a MOST-DKFZ ResearchGrant, EuropeanConsortiumGrantMRTN-CT-2005, theRosetreesTrust, Israel CancerAssociationGrant 20100003, theAssociation for International Cancer Research, a Cro-atia-Israel Joint Research Grant (all to O.M.), and European Union Grant FP7-REGPOT-2008-1 (to S.J.). O.M is a Crown Professor of Molecular Immunology.
Address correspondence and reprint requests to Dr. Ofer Mandelboim, The Lauten-berg Center for General and Tumor Immunology, Hadassah Medical School, HebrewUniversity, P.O. Box 12272, Jerusalem, Israel or Dr. Stipan Jonjic, Department ofHistology and Embryology Center for Proteomics, Faculty of Medicine, University ofRijeka, Brace Branchetta 2051 000, Rijeka, Croatia. E-mail addresses: [email protected] or [email protected]
The online version of this article contains supplemental material.
Abbreviations used in this paper: BulkNK, bulkNKcultures; HA, hemagglutinin; INH,inhibited; KIR, killer Ig-related receptor; MIX, mixed population of inhibited andnoninhibited clones; NK Fresh, freshly isolated NK cells; non-INH, noninhibited;WT, wild-type.
Copyright� 2010 by The American Association of Immunologists, Inc. 0022-1767/10/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.0903347
by guest on September 25, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

viously observed or that CD300c is not an activating receptor.Finally, the ligands for both receptors are still unknown.In this study, we investigated the activity of CD300a. We dem-
onstrate that CD300a is actually functional only on a subset of NKclones and that CD300a and CD300c are indistinguishable onthe cell surface, and we identified a critical ITIM that is importantfor CD300a function.
Materials and MethodsAbs and cells
The following mouse anti-human Abs were used: anti-CD300a/c (P192)(20), anti-CD300a/c (MEM-260, Cedarlane Laboratories, Hornby, Ontario,Canada), anti-MHCI (W6/32), anti-KIR2DL1 (HP3E4), anti-KIR2DL3(CD158b, Beckman Coulter, Fullerton, CA), anti-CD16 (B73.1.1), anti-CD99 (12E7; a kind gift fromA.Bernard,Hopital deL’Archet,Nice, France),anti-hemagglutinin (HA) tag (12C45), anti-2B4 (CD244 clone C1.7, Beck-manCoulter), andanti-CD3 (T3D).Togenerate theanti-CD300aandCD300cmAbs, BALB/c mice were injected s.c. with the fusion proteins in CFA. Twoweeks later, mice were boosted with the same protein in IFA. After an addi-tional 2wk, the sera of immunizedmicewere screened for theAb titer againstthe immunogen using ELISA. The best responders were additionally boostedi.p. with fusion protein dissolved in PBS. Three days later, spleen cells werecollected, and after lysis of erythrocytes, they were fused with SP2/0 mye-loma cells in a 1:1 ratio. The cellswere seeded on96-well tissue-culture platesin 20% RPMI 1640 medium with hypoxanthine, aminopterine, and thymi-dine. The cultures were fed every 4 d; after 12 d, each well was screened forAbs reactive against fusion proteins in ELISA. Positive mother wells wereexpanded and cloned.
The cell lines used in the current study were mouse BW cells, a murinethymoma cell line that lacks TCR expression; YTS cells, a subline of the YTNK leukemia cell line (24) transfected with the ecotropic murine retrovirusreceptor alone (YTS/eco) or the ecotropic receptor; and CD300a: wild-type(WT) or mutated receptor (YTS CD300a or YTS Y231A, YTS Y255A,YTS Y267A, and YTS Y293A) (25).
Isolation of human NK cells
Primary NK cells were isolated from PBLs using the human NK cell iso-lation kit and the autoMACS instrument (Miltenyi Biotec, Auburn, CA). Toobtain NK clones, freshly separated NK cells were diluted in NK medium,based on the number of cells desired per well, and seeded in 96U plates. NKclones were collected and split every 2–3 d in a 1:2 ratio. The validation ofNK cell phenotype was performed by FACS staining using the mouse anti-human Abs anti-CD56 (BD Biosciences) and anti-CD3 (BioLegend, SanDiego, CA) or anti-NKp46 (R&D Systems, Minneapolis, MN).
YTS and BW transfectants
The cDNA of CD300a used for transfection into YTS/eco cells wasamplified by PCR from cDNA isolated from human NK clones. Theprimers used for the PCR of the CD300a were 59-CCG GAA TTC GCCGCC ACC ATG TGG CTG CCT TGG GCT CTG (including an EcoRIrestriction site) and 39-TT CC GCG GCC GCT ATG GCC GAC GTCGAC CTA TGT CTT CCT TAT CAC ACT (including an SalI restrictionsite). The tyrosine residues located in positions 231, 255, 267, and 293 ofthe cytoplasmic tail of the CD300a protein were mutated to alanine bytwo-step PCR amplification using the following primers: 39-CTC CAGATT TGC GGC GTG CAG CTC, 39-AGT GCT GGC TTC CAC CTCCAC CTC, 39-GAA CTT CAC GCT GCC TCG GTG GTG, 59-GAGCTG CAC GCC GCA AAT CTG GAG, 59-GAG GTG GAG GTGGAA GCC AGC ACT, and 59-GAA CTT CAC GCT GCC TCG GTGGTG. For Y293A, one-step PCR amplification was done using the fol-lowing primer: 59-TT CC GCG GCC GCT ATG GCC GAC GTC GACCTA TGT CTT CTT TAT CAC ACT GGC ATC TGA. The WT ormutated CD300a constructs were cloned into a pBABE-puro retrovirusvector, and YTS cells were transfected with cDNA encoding for theCD300a protein, as previously described (26, 27).
The generation of chimeric CD300c HA z cDNA used for transfectioninto BW cells was performed by two-step PCR amplification using cDNAisolated from human NK clones. The primers used for the PCR of theextracellular portion of CD300c were: 59-CCC AAG CTT GCC GCCACC ATG ACT GCC AGG GCC TGG GCC (including a HindIII restric-tion site) and 39-GTA GCA GAG GCG GAC ATT GCT GAA CAG GGA(including the first seven nucleotides of mouse z-chain transmembraneportion). The mouse z-chain was amplified by PCR using the followingprimers: 59-AAT GTC CGC CTC TGC TAC TTG CTA GAT GGA
(including the last seven nucleotides of human CD300c extracellular por-tion) and 39-CC GAA TTC TTA GCG AGG GGC CAG GGT CTG (in-cluding an EcoRI restriction site). The two amplified fragments weremixed, and PCR was performed with the 59 HindIII primer and the 39EcoRI primer for the generation of the CD300c z construct. To insertthe HA tag, the following primers were used: 59HindIII and 39-AGCGTA ATC TGG AAC ATC GTA TGG GTA AGG AAA ATA GCC TGGGAC AAG (including the HA tag fragment) and 59-TAC CCA TAC GATGTT CCA GAT TAC GCT CTG AGC CAC CCC ATG ACC GTG (in-cluding the HA tag fragment) and 3EcoRI. The CD300c HA z constructwas cloned into a pcDNA3 expression vector (Invitrogen, Carlsbad, CA)and stably transfected into BW cells.
Ig-fusion proteins
The Ig fusion proteins CD300a-Ig, CD300c-Ig, and D1-Ig were generated asdescribed previously (28). Briefly, the sequence encoding the extracellularportion of the receptor was amplified by PCR from cDNA isolated fromhuman NK clones. The primers used for the PCR of the extracellularportion of CD300a/c were CD300a: 59-CCC AAG CTT GCC GCC ACCATG TGG CTG CCT TGG GCT CTG (including a HindIII restriction site)and 39-GA AG ATC TGA GTT CAC CAC CTC CTC AGT (includinga BglII restriction site); CD300c: 59-CCC AAG CTT GCC GCC ACCATG ACT GCC AGG GCC TGG GCC (including a HindIII restrictionsite) and 39-GA AG ATC TCG GAC ATT GCT GAA CAG GGA (in-cluding a BglII restriction site). These PCR-generated fragments werecloned into a mammalian expression vector containing the Fc portion ofa human IgG1. The constructs were transfected into COS-7 cells, and theprotein produced was purified using a protein G column.
Redirected killing assay
The inhibitory activity of CD300a in NK cells was assessed in a redirectedkilling assay, in which NK line cells, NK clones, or various YTS cells werecocultured with 35S-labeled P815 (murine mastocytoma) target cells thatwere precoated with 0.2 mg/well various mAbs: anti-CD16, anti-CD99,anti-KIR2DL3, or anti-CD300 Abs. The E:T ratio was 3:1 or 5:1 for NKline/clones cells or YTS cells, respectively. In all presented cytotoxicityassays, the spontaneous release was ,20% of maximal release. The levelof cytotoxicity was determined as previously described (29).
PCR and quantitative real-time PCR
The sequence encoding the CD300a/c receptor was amplified by PCR fromcDNA isolated from human NK clones. The primers used for the PCR of theCD300a/c receptor were CD300a: 59-CCG GAA TTC GCC GCC ACCATG TGG CTG CCT TGG GCT CTG (including an EcoRI restriction site)and 39-TT CC GCG GCC GCT ATG GCC GAC GTC GAC CTA TGTCTT CCT TAT CAC ACT (including an SalI restriction site); CD300c: 59-CCG GAA TTC GCC GCC ACC ATG ACT GCC AGG GCC TGG GCC(including an EcoRI restriction site) and 39-TT CC GCG GCC GCT ATGGCC GAC GTC GAC CTA CTG GTT CTC ACC CTT GGG (including anSalI restriction site).
For quantitative real-time PCR analysis, total RNAwas isolated with Per-fectPure RNA Cultured cell Kit (5 PRIME), and cDNAwas generated usingthe SuperScript II kit (Invitrogen). DNAwas amplified with specific primers:CD300a: 59-GCA TTG TTG CTG CTT CTG TTG and 39-CAC CAG CTTTGATCC ATT TCTG; CD300c: 59-ACA CCC TGG CTC CCT GTTC and39-CTA GAG CTT CTC TGA GGT CTG TTC AC. Real-time PCR wasperformed with a DyNAmo SYBR Green qPCR kit (Invitrogen) on an ABIPRISM 7500 Real-Time PCR system (Applied Biosystems, Foster City, CA).
Western blot and immunoprecipitation
CD300a/c detection.NK cells (20M cell) were washed four times with coldPBS containing 1 mMMgCl2 and 0.1 mM CaCl2. For immunoprecipitationwith anti-CD300a/c (CD300c Hybridoma #9), the cells were biotinylatedwith EZ-Link Sulfo-NHS-LC-Biotin (Pierce, Rockford, IL) for 30 min at4˚C and washed four times with cold PBS. After washing, cells were lysedwith lysis buffer (PBS containing 150 mM NaCl, 50 mM Tris [pH 7.6],0.5% Nonidet P-40, 9 mM iodoacetamide, 5 mM EDTA, 1 mM PMSF,1:100 aprotinin). Cell lysates were then immunoprecipitated overnight at4˚C with Protein-G plus beads (Pierce) conjugated with 100 ml anti-CD300a/c (CD300c Hybridoma #9). The immunoprecipitates were washedwith lysis buffer, and the proteins were eluted by using IgG elution buffer(Pierce). All samples were loaded on SDS-PAGE and transferred to nitro-cellulose membrane. The blotted proteins were visualized by streptavidinHRP (Jackson ImmunoResearch Laboratories, West Grove, PA) using ECLdetection (Biological Industries, Bet-haemek, Israel).
2878 EXPRESSION AND FUNCTION OF CD300 IN NK CELLS
by guest on September 25, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

CD300a phosphorylation
NK cells (20M) were washed with TBS and activated with 500 mM H2O2
for 30 s and 5 min incubation at 37˚C. Control cells were not exposed toH2O2. Following incubation, cells were washed three times with coldTBS-PI (containing phosphate inhibitors). After washing, cells were lysedwith lysis buffer (TBS containing 0.5% Triton X-100, 50 mM Tris (pH 7.4),150 mM NaCl, and phosphatase and protease inhibitors). Cell lysates werethen loaded on SDS-PAGE and transferred to nitrocellulose membrane. Theblotted phosphorylated proteins were visualized by anti-phosphotyrosine(4G10; Upstate Biotechnology, Lake Placid, NY) after incubation over-night at 4˚C, goat anti-mouse biotin for 2 h at room temperature andstreptavidin HRP (Jackson ImmunoResearch Laboratories), using ECL de-tection (Biological Industries).
Internalization assay
Cells were resuspended at 105 in 100 ml RPMI 1640/10% FCS and in-cubated on ice for 30 min with 5 ml anti-CD300a/c (CD300c Hybridoma#9) mAb. Cells were then washed three times with RPMI 1640/10% FCSand incubated at 37˚C in 5% CO2. After various incubation times, aliquotsof cells were removed, fixed by the addition of FACS medium (0.1% azide/1% BSA/PBS), and kept on ice. After an appropriate number of aliquotshad been taken, cells were labeled with an excess of FITC-labeled goatanti-mouse IgG for 30 min and subsequently washed in FACS mediumbefore analysis by flow cytometry.
ResultsCD300a expression and function on NK cells
It was shown previously that CD300a is an inhibitory receptor (20,21). However, it is still unknown whether CD300a is an inhibitoryreceptor for all NK clones. Therefore, we initially tested whetherCD300a is expressed by all NK cells. For that purpose, we isolated
PBLs and NK cells from healthy donors and analyzed them forCD300a expression by using a commercial mAb P192, pre-sumably specific to CD300a. Indeed, as previously reported,a subset of PBLs expressed CD300a, and all NK cells tested, ei-ther freshly isolated or bulk NK cell cultures, which were grown inthe presence of IL-2 (NK bulk), expressed the CD300a receptor(Fig. 1A). C1R cells were used as a positive control and werealso stained with the anti-CD300a mAb P192 (Fig. 1A). To testwhether the uniform expression of CD300a was also observed invarious NK clones derived from different donors, we isolated NKclones from two different healthy donors and analyzed them forCD300a expression. As can be seen in Fig. 1B and 1C, all NKclones expressed CD300a, in contrast to KIR2DL1, which isexpressed by only some NK clones. Similar results were observedin all NK clones that we tested (.100). Thus, all NK cells testedexpressed CD300a, as detected by staining with the P192 mAb.We next tested the functionality of CD300a. Primary bulk
NK cell cultures were incubated with P815 cells, which wereprecoated with anti-CD16 mAb (to induce killing), in the presenceor absence of mAb P192, anti-CD99 (which was used as a negativecontrol), and anti-KIR2DL3 (which was used as a positive control)(Fig. 1D). As previously reported, the coincubation of NK cellswith the anti-CD16 and anti-CD300a/c mAb resulted in an inhi-bition of killing compared with the activation by anti-CD16 mAbalone (Fig. 1D). However, surprisingly, despite the observationsthat CD300a contains four ITIMs and is expressed on all NK cells,the CD300a-mediated inhibition seemed to be moderate comparedwith the inhibition mediated by the other NK receptors (e.g.,
FIGURE 1. CD300a expression and function. A, Flow-cytometric analysis of Bulk NK, NK Fresh, PBLs, and CIR cells. B and C, Flow-cytometric
analysis of Bulk NK and NK clones obtained from two healthy donors. Staining was performed with 5 ml/well anti-CD300a/c mAb (P192) and 100 ml anti-
KIR2DL1 (HP3E4 Hybridoma) (black line). Background (shaded graph) is the staining with FITC-conjugated F(ab9)2 goat anti-mouse IgG only. D,
Redirected experiments were performed with 0.1 mg/well anti-CD16 (B73.1), 0.1 mg/well anti-CD99 (12E7), 0.1 mg/well anti-KIR2DL3 (GL183b), and 5
ml/well anti-CD300a/c (P192). The E:T ratio was 3:1. The horizontal line represents the inhibition observed when the CD300a receptor was cross-linked
with CD16. pp # 0.05. Representative result of four separate experiments. Bulk NK, bulk NK cultures; NK Fresh, freshly isolated NK cells.
The Journal of Immunology 2879
by guest on September 25, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

KIR2DL3; Fig. 1D), and it was not observed in all NK clones(data not shown).
CD300a and CD300c are indistinguishable on the cell surface
Because the CD300a inhibition observed was weak (Fig. 1D) andwas not observed in all NK clones (data not shown), we hypoth-esized that CD300a is not expressed by all NK cells and that theanti-CD300a mAb (P192) used by us in the above experimentsand by other investigators (20, 30), cross-reacted with anotherprotein. Indeed, the CD300c, which was suggested to be a putativeactivating receptor, showed high similarity to CD300a at the DNAand protein levels (Fig. 2A). The expression pattern of CD300cand its precise activity are still unknown. To test whether the P192mAb cross-reacts with CD300a and CD300c, we constructed sev-eral reagents: YTS cells expressing CD300a and BW cells expres-sing CD300c in-frame with mouse z-chain. Because we did nothave an Ab for CD300c, the CD300c protein was tagged with HA,and its expression was confirmed with anti-HA mAb. Despite theuse of several plasmids (including lenti vectors and other expres-sion vectors) and several attempts, we could not express CD300a zin BW cells. However, we were able to express the full-lengthCD300a in BW cells and, thus, we assumed that the CD300a longcytoplasmic tail is required for stable expression of the protein onthe cell surface.In addition, we generated two reagents in which we fused the
extracellular portion of CD300a and CD300c to human IgG1 andproduced these fusion proteins in COS-7 cells. Equipped with thesereagents, we now generated new mAbs directed against CD300aand CD300c. Mice were injected with these two proteins, andthe emerging hybridomas were tested against the YTS and theBW cells expressing CD300a and CD300c, respectively, as well as
against NK cells. Using this strategy, we obtained four new anti-CD300a mAbs (CD300a Hybridoma #1–4) and nine new anti-CD300c mAbs (CD300c Hybridoma #6–14). Importantly, none ofthe mAbs generated, including the P192 mAb, could discriminatebetween CD300a and CD300c, and all Abs recognized all of the NKclones and bulk NK cultures tested (Fig. 3). Although it was pre-viously demonstrated that the anti-CMRF35 mAb recognizes theshort protein (CD300c) and the long protein (CD300a) (31–33),the question of whether this observation is restricted to this mAbor is a general phenomenon is still open. In this study, we show thatsuch cross-reactivity is also observed with a large panel of Abs, aswell as the commercial mAb P192.
NK clones exhibit different phenotypes upon CD300 cross-linking
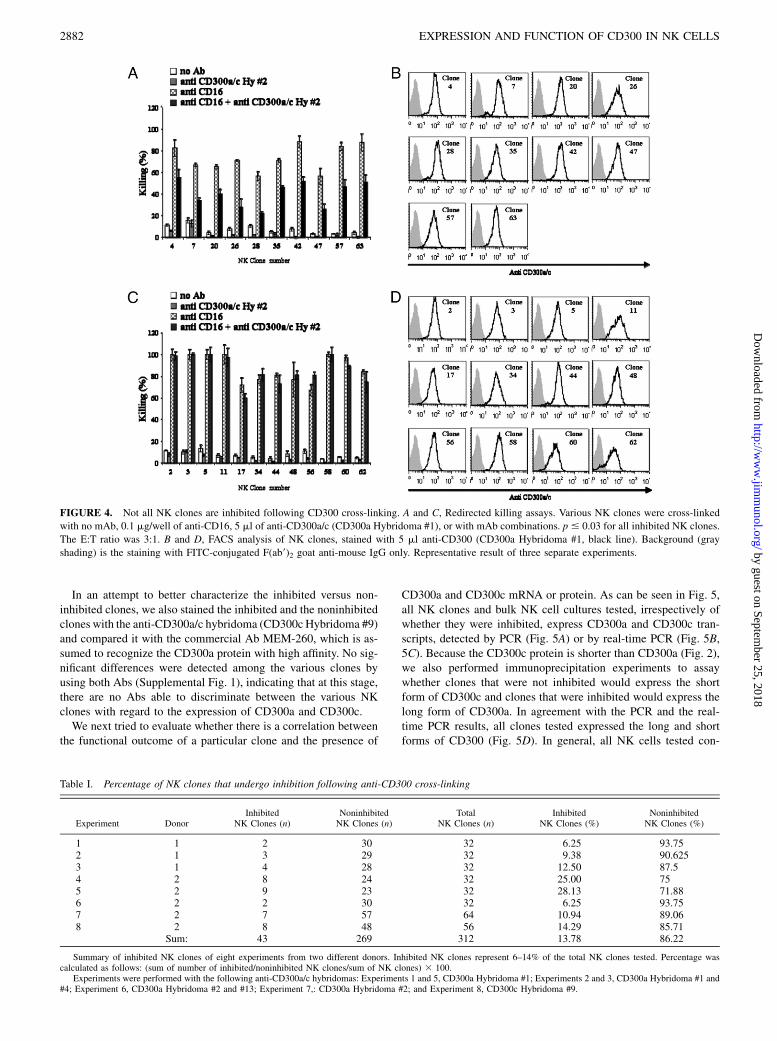
As demonstrated above, and even by using a large panel of newmAbs that we generated, we could not distinguish between CD300aand CD300c on the cell surface of various cells. In addition, theinhibition of CD300a was moderate compared with KIR inhibition(Fig. 1D). Thus, we next examined whether there would be a dif-ference in NK inhibition upon CD300a/c cross-linking of differentNK clones. Indeed, as shown in Fig. 4 and in Table I, only a smallportion of the NK clones tested was inhibited by CD300a/c cross-linking, whereas the majority of the clones tested could not beinhibited by the cross linking of CD300a/c (Fig. 4 and Table I).Although the percentages of the inhibitory clones vary betweendifferent donors, our conclusion, which is deduced from experi-ments performed in several donors, as well as with several differ-ent anti-CD300a/c hybridomas, is that most of the NK clones arenot inhibited by CD300a/c cross-linking (Table I). Interestingly,the CD16-mediated killing of clones that were not inhibited was
FIGURE 2. A, CD300a and CD300c share a high degree of homology. Sequence alignment of CD300a and CD300c using the Clustalw program. B,
Schematic presentation of the CD300a and CD300c receptors.
2880 EXPRESSION AND FUNCTION OF CD300 IN NK CELLS
by guest on September 25, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

not enhanced by the CD300 cross-linking (Fig. 4), and cross-linking of CD300 alone on these clones did not induce killing,suggesting that CD300c might not be an activating NK cell
receptor. As expected, all clones tested expressed CD300a/c; how-ever, because of the mAb cross-reactivity, it is not clear whetherCD300a, CD300c, or both are present on the cell surface (Fig. 4).
FIGURE 3. CD300a and CD300c are indistin-
guishable on the cell surface. FACS analysis of bulk
NK cultures (A), YTS cells expressing CD300a (B),
and BW cells expressing CD300c fused to HA9 z (C).
Staining is represented by the black line. Background
(shaded graph) is the staining with FITC-conjugated
F(ab9)2 goat anti-mouse IgG only. Representative re-
sult of three separate experiments.
The Journal of Immunology 2881
by guest on September 25, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
In an attempt to better characterize the inhibited versus non-inhibited clones, we also stained the inhibited and the noninhibitedclones with the anti-CD300a/c hybridoma (CD300c Hybridoma #9)and compared it with the commercial Ab MEM-260, which is as-sumed to recognize the CD300a protein with high affinity. No sig-nificant differences were detected among the various clones byusing both Abs (Supplemental Fig. 1), indicating that at this stage,there are no Abs able to discriminate between the various NKclones with regard to the expression of CD300a and CD300c.We next tried to evaluate whether there is a correlation between
the functional outcome of a particular clone and the presence of
CD300a and CD300c mRNA or protein. As can be seen in Fig. 5,all NK clones and bulk NK cell cultures tested, irrespectively ofwhether they were inhibited, express CD300a and CD300c tran-scripts, detected by PCR (Fig. 5A) or by real-time PCR (Fig. 5B,5C). Because the CD300c protein is shorter than CD300a (Fig. 2),we also performed immunoprecipitation experiments to assaywhether clones that were not inhibited would express the shortform of CD300c and clones that were inhibited would express thelong form of CD300a. In agreement with the PCR and the real-time PCR results, all clones tested expressed the long and shortforms of CD300 (Fig. 5D). In general, all NK cells tested con-
FIGURE 4. Not all NK clones are inhibited following CD300 cross-linking. A and C, Redirected killing assays. Various NK clones were cross-linked
with no mAb, 0.1 mg/well of anti-CD16, 5 ml of anti-CD300a/c (CD300a Hybridoma #1), or with mAb combinations. p # 0.03 for all inhibited NK clones.
The E:T ratio was 3:1. B and D, FACS analysis of NK clones, stained with 5 ml anti-CD300 (CD300a Hybridoma #1, black line). Background (gray
shading) is the staining with FITC-conjugated F(ab9)2 goat anti-mouse IgG only. Representative result of three separate experiments.
Table I. Percentage of NK clones that undergo inhibition following anti-CD300 cross-linking
Experiment DonorInhibited
NK Clones (n)NoninhibitedNK Clones (n)
TotalNK Clones (n)
InhibitedNK Clones (%)
NoninhibitedNK Clones (%)
1 1 2 30 32 6.25 93.752 1 3 29 32 9.38 90.6253 1 4 28 32 12.50 87.54 2 8 24 32 25.00 755 2 9 23 32 28.13 71.886 2 2 30 32 6.25 93.757 2 7 57 64 10.94 89.068 2 8 48 56 14.29 85.71
Sum: 43 269 312 13.78 86.22
Summary of inhibited NK clones of eight experiments from two different donors. Inhibited NK clones represent 6–14% of the total NK clones tested. Percentage wascalculated as follows: (sum of number of inhibited/noninhibited NK clones/sum of NK clones) 3 100.
Experiments were performed with the following anti-CD300a/c hybridomas: Experiments 1 and 5, CD300a Hybridoma #1; Experiments 2 and 3, CD300a Hybridoma #1 and#4; Experiment 6, CD300a Hybridoma #2 and #13; Experiment 7,: CD300a Hybridoma #2; and Experiment 8, CD300c Hybridoma #9.
2882 EXPRESSION AND FUNCTION OF CD300 IN NK CELLS
by guest on September 25, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

tained lower levels of CD300c and although it seems as if somedifferences could be observed among the various cells with regardto CD300c expression, they were not significant.A general defect in the inhibitory capacity of these clones might
explain why inhibition was not observed in some of the clones.Therefore, we isolated noninhibitory clones that express otherinhibitory receptors, such as KIR2DL1 and KIR2DL3 (Supple-mental Fig. 2), and redirected the killing of these clones by usinganti-CD300a/c Abs, as well as the anti-KIR2DL1 or -KIR2DL3mAb. Because the triggering of the inhibitory receptors, togetherwith CD16, resulted in efficient inhibition in these clones, whereasno inhibition was observed with CD300a/c (Supplemental Fig. 2),we concluded that noninhibited clones are normal functional cloneswith regard to NK inhibition.To further substantiate this point, we also tested the general
phosphorylation pattern of the inhibitory versus the noninhibitoryclones by treating them with H2O2. As can be seen in SupplementalFig. 3, there was no major difference in the phosphorylation patternbetween the clones, further indicating that the noninhibitory clonesare normal (Supplemental Fig. 3).Thus, it seems that CD300 activity is quite complex and that,
despite the observations that different clones express CD300a andCD300c transcripts and proteins, some are functionally inhibitedby anti-CD300a/c, whereas others are not.
The third ITIM motif of CD300a is important for its inhibitoryfunction
A common feature of the inhibitory receptors is the presence of oneor more ITIMs in their cytoplasmic tail. The cytoplasmic portion ofCD300a contains three motifs (LHYANL, VEYSTV, and LHYASV),which fits with the consensus sequence of a classical ITIM (I/V/L/S xYxx L/V) and a fourth nonclassical motif (SDYSVI) (20, 21).To study which of the four ITIMs is most important for the
inhibitory signal transduction, we inserted a point mutation in eachITIM, changing tyrosine to alanine. Once we constructed 15CD300a proteins, each with a different ITIM mutation (Fig. 6), weexpressed each of the proteins in the YTS cell line and performeda redirected killing assay. All proteins were expressed at similarlevels on the cell surface, except from the CD300a protein, in whichall four ITIMs were mutated (Fig. 6A), suggesting that the ITIMsare probably needed for efficient cell surface expression of CD300a.This result might also explain the lack of expression of CD300a z
on the surface of BW cells, because it lacks the tail and trans-membrane of CD300a. To test this hypothesis, we performed in-ternalization experiments in which the quadruple CD300a mutant,CD300c, and the parental CD300a-expressingYTS cellswere stainedwith anti-CD300a, and the spontaneous internalization ratewas mea-sured over time. As can be seen in Supplemental Fig. 4, the internal-ization rate of the quadruple CD300amutant and that of CD300cwasmore rapid compared with the parental CD300a-expressing cells,suggesting that the ITIM motifs are important for the stable expres-sion of CD300a on the cell surface.We next assessed the importance of each of the ITIM motifs in
the inhibition mediated by CD300a. Although single-point muta-tion in a single ITIM had no effect on the CD300a-mediated inhi-bition, double or triple mutation involving the third ITIM, Y267,significantly reduced the CD300a-mediated inhibition (Fig. 6).Combinations of double mutation, which did not include the thirdITIM, Y267, did not affect the CD300a inhibition, whereas triplemutations significantly downregulated the receptor’s inhibitoryfunction. In the triple mutant, reduced inhibitory activity ofCD300a was observed even when the third ITIM was not mutated(Fig. 6B, triple mutation Y231A + Y255A + Y293A), albeit, toa lower extent compared with other triple mutations that includedthe third ITIM. Finally, mutation in all four ITIMs completelyabolished the inhibition caused by the CD300a receptor. These
FIGURE 5. All clones tested express the mRNA and the protein of CD300a and CD300c. RT-PCR (A) and real-time RT-PCR analysis (B) were
performed on mRNA extracted from different NK clones. NK clones were divided into four groups: INH, non-INH, MIX, and NK bulk. Relative mRNA
abundance is shown as a percentage of the level of hypoxanthine phosphoribosyltransferase mRNA (encoding hypoxanthine guanine phosphoribosyl-
transferase), and error bars (SD) are derived from triplicates. D, All NK clones, INH and non-INH, express CD300a and CD300c proteins. The cells were
biotinylated, and cell lysates were immunoprecipitated overnight at 4˚C with Protein-G plus beads (Pierce) conjugated with 100 ml anti-CD300a/c. The
blotted proteins were visualized by streptavidin-HRP using ECL. Representative of two independent experiments. INH, inhibited; non-INH, noninhibited;
MIX, mixed population of inhibited and noninhibited clones; NK bulk, bulk NK cultures.
The Journal of Immunology 2883
by guest on September 25, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

results indicate that all four ITIMs are probably involved in theCD300a-mediated inhibition and that the third ITIM is the mostcrucial ITIM involved in this inhibition.
DiscussionNK cell activation results from a balance of opposing signals that,on the one hand, activate NK cells, allowing them to eradicatetumor or virally infected cells, and, on the other hand, inhibit theNK cells, to prevent killing of self (34, 35). The NK inhibitoryreceptors, which recognize MHC class I proteins, are the moststudied NK cell receptors with regard to NK cell inhibition;however, in recent years, it became apparent that MHC classI-independent inhibitory receptors also exist (9, 36, 37).The CD300a and CD300c receptors of the CD300 family belong
to the Ig super-family of proteins. Little is known about their cell-surface expression phenotype, their functionality, and the identityof their ligand/s. In this study, we tried to shed new light on theinvolvement of these two proteins in the regulation of humanNK cell activity. CD300a has been identified as a surface moleculeexpressed on all NK cells (20, 21), displaying an inhibitory func-tion upon binding to its unknown non-MHC class I ligand (20,21). As we showed in this article, CD300a might be expressed on
all NK clones; however, not all NK clones could deliver an inhib-itory signal, probably due to the presence of CD300c. Indeed,CD300a and CD300c share a high degree of homology (23). Sucha high degree of homology is a reminiscent of paired receptors, suchas KIR2DL and KIR2DS receptors. These two receptors, perhapssimilarly to the CD300a and CD300c pair, share a high degree ofsequence resemblance but differ in function; KIR2DL is an inhib-itory receptor, whereas KIR2DS is a coactivating receptor (17, 18).In contrast, although the function of CD300a as an inhibitory re-ceptor is well established [this study and (19, 20)], the function ofCD300c is yet to be determined. Furthermore, unlike other activat-ing KIR receptors that contain a positively charged amino acid inthe transmembrane region, which provides a docking site for theactivating adapter molecules (17, 18), CD300c has a negativecharged amino acid (glutamic acid) in the transmembrane region,and its associated partner/s remain to be revealed (19, 22).We show that although transcripts and proteins of CD300a and
CD300c are found within a particular clone, some NK clones areinhibited, whereas others are not. Furthermore, we show that thenoninhibitory clones are normal with regard to NK inhibition(i.e., the triggering of other inhibitory receptors on these clonesleads to NK cell inhibition). The different function of the various
FIGURE 6. Mutation of the CD300a ITIM motifs causes downregulation of the receptor’s inhibition. A, Flow-cytometric analysis of the different
CD300a mutants stained with 5 ml of anti-CD300a/c mAb (CD300a Hybridoma #1) (open graph). Background (shaded graph) is the staining with FITC-
conjugated F(ab9)2 goat anti-mouse IgG only. Representative result of three separate experiments. B, Redirected killing assay of YTS cells expressing the
different mutants and YTS CD300a (WT). CD300a receptor cross-linking was performed with 0.1mg/well anti-2B4 and 5 ml of anti-CD300a/c (CD300a
Hybridoma #1). The E:T ratio was 5:1. pp # 0.03; ppp # 0.00005. Representative result of three separate experiments. C, Table summarizing the various
mutations used.
2884 EXPRESSION AND FUNCTION OF CD300 IN NK CELLS
by guest on September 25, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

clones could be explained in several ways. Because there are nomAbs that could discriminate between CD300a and CD300c, it ispossible that, on the cell surface, the expression of these proteins isdifferentially controlled or modified. It is also possible that oneprotein antagonizes the activity of the other and that the activity ofCD300a is more dominant in the inhibited clones. Although un-likely, it is possible that the expression of CD300c in NK cells isirrelevant to the inhibitory activity of CD300a. Finally, it might bepossible that the activity of CD300a is somehow dependent uponCD300c. Clearly, further research needs to be performed with thisregard, including the generation of mAbs that could discriminatebetween the ectodomains of the two receptors.Indeed, because CD300a and CD300c are indistinguishable on
the cells surface, many questions still need to be answered, forexample: what is the expression pattern of CD300a and CD300creceptors on NK cells? Does it change following NK activation?Can it be influenced by cytokines?What is the function of CD300c?All of these questions cannot be precisely answered until specificmAbs for CD300a and CD300c are developed.The CD300a receptor is expressed on NK cells, as well as on
many other immune cells, such as plasmacytoid dendritic cells (38),T cells (39), neutrophils (40), mast cells (30, 41), and others (42,43). Different studies evaluated the CD300a activity in immuneregulation. CD300a Ab cross-linking resulted in reduced TNF-asecretion and increased IFN-g secretion in plasmacytoid dendriticcells (38), CD300a affected T cell proliferation and IFN-g secre-tion (39), coactivation of CD300a and CD32a inhibited CD32a-mediated signaling in neutrophils (40), and mAb neutralization ofLMIR1 (the mouse CD300a homolog) in BALB/c mice led to a sig-nificant augmentation of inflammatory mediators and eosinophilicinfiltration (30). In addition, a genetic link was established betweenthe CD300a and CD300c chromosomal location, 17q25, and psori-asis, which may link CD300a and CD300c as part of the dis-ease mediators (44). In view of the observasions that CD300a andCD300c are so far indistinguishable, these works should be reeval-uated because some of the effects seen might be due to the activationof CD300a or CD300c or both.We demonstrated that the ITIM motifs of CD300a are important
for its stability on the cell surface and that the tyrosine residue inthe third ITIM motif, Y267A, is the most important residue for theexecution of the inhibitory signal. Further understanding of thereceptor’s signal transduction might be useful for the identificationof unique downstream components, which can be targets for drugdevelopment and disease treatment.
DisclosuresThe authors have no financial conflicts of interest.
References1. Bisset, L. R., T. L. Lung, M. Kaelin, E. Ludwig, and R. W. Dubs. 2004. Ref-
erence values for peripheral blood lymphocyte phenotypes applicable to thehealthy adult population in Switzerland. Eur. J. Haematol. 72: 203–212.
2. Colucci, F., M. A. Caligiuri, and J. P. Di Santo. 2003. What does it take to makea natural killer? Nat. Rev. Immunol. 3: 413–425.
3. Biassoni, R., C. Cantoni, D. Pende, S. Sivori, S. Parolini, M. Vitale, C. Bottino,and A. Moretta. 2001. Human natural killer cell receptors and co-receptors.Immunol. Rev. 181: 203–214.
4. Cerwenka, A., and L. L. Lanier. 2001. Natural killer cells, viruses and cancer.Nat. Rev. Immunol. 1: 41–49.
5. Yokoyama, W. M., S. Kim, and A. R. French. 2004. The dynamic life of naturalkiller cells. Annu. Rev. Immunol. 22: 405–429.
6. Moretta, A., C. Bottino, M. Vitale, D. Pende, C. Cantoni, M. C. Mingari,R. Biassoni, and L. Moretta. 2001. Activating receptors and coreceptors involved inhuman natural killer cell-mediated cytolysis. Annu. Rev. Immunol. 19: 197–223.
7. Biassoni, R., C. Cantoni, M. Falco, D. Pende, R. Millo, L. Moretta, C. Bottino,and A. Moretta. 2000. Human natural killer cell activating receptors. Mol.Immunol. 37: 1015–1024.
8. Biassoni, R., C. Cantoni, D. Marras, J. Giron-Michel, M. Falco, L. Moretta, andN. Dimasi. 2003. Human natural killer cell receptors: insights into theirmolecular function and structure. J. Cell. Mol. Med. 7: 376–387.
9. Lebbink, R. J., and L. Meyaard. 2007. Non-MHC ligands for inhibitory immunereceptors: novel insights and implications for immune regulation. Mol. Immunol.44: 2153–2164.
10. Raulet, D. H. 2003. Roles of the NKG2D immunoreceptor and its ligands. Nat.Rev. Immunol. 3: 781–790.
11. Pessino, A., S. Sivori, C. Bottino, A. Malaspina, L. Morelli, L. Moretta,R. Biassoni, and A. Moretta. 1998. Molecular cloning of NKp46: a novelmember of the immunoglobulin superfamily involved in triggering of naturalcytotoxicity. J. Exp. Med. 188: 953–960.
12. Cantoni, C., C. Bottino, M. Vitale, A. Pessino, R. Augugliaro, A. Malaspina,S. Parolini, L. Moretta, A. Moretta, and R. Biassoni. 1999. NKp44, a triggeringreceptor involved in tumor cell lysis by activated human natural killer cells, isa novel member of the immunoglobulin superfamily. J. Exp. Med. 189: 787–796.
13. Pende, D., S. Parolini, A. Pessino, S. Sivori, R. Augugliaro, L. Morelli, E. Marcenaro,L. Accame A. Malaspina, R. Biassoni, et al. 1999. Identification and molecularcharacterization of NKp30, a novel triggering receptor involved in natural cy-totoxicity mediated by human natural killer cells. J. Exp. Med. 190: 1505–1516.
14. Long, E. O. 1999. Regulation of immune responses through inhibitory receptors.Annu. Rev. Immunol. 17: 875–904.
15. Lopez-Botet, M., and T. Bellon. 1999. Natural killer cell activation andinhibition by receptors for MHC class I. Curr. Opin. Immunol. 11: 301–307.
16. Natarajan, K., N. Dimasi, J. Wang, R. A. Mariuzza, and D. H. Margulies. 2002.Structure and function of natural killer cell receptors: multiple molecular sol-utions to self, nonself discrimination. Annu. Rev. Immunol. 20: 853–885.
17. Bashirova, A. A., M. P. Martin, D. W. McVicar, and M. Carrington. 2006. Thekiller immunoglobulin-like receptor gene cluster: tuning the genome for defense.Annu. Rev. Genomics Hum. Genet. 7: 277–300.
18. Uhrberg, M. 2005. The KIR gene family: life in the fast lane of evolution. Eur. J.Immunol. 35: 10–15.
19. Clark, G. J., X. Ju, C. Tate, and D. N. Hart. 2009. The CD300 family of mol-ecules are evolutionarily significant regulators of leukocyte functions. TrendsImmunol. 30: 209–217.
20. Cantoni, C., C. Bottino, R. Augugliaro, L. Morelli, E. Marcenaro, R. Castriconi,M. Vitale, D. Pende, S. Sivori, R. Millo, et al. 1999. Molecular and functionalcharacterization of IRp60, a member of the immunoglobulin superfamily thatfunctions as an inhibitory receptor in human NK cells. Eur. J. Immunol. 29:3148–3159.
21. Clark, G. J., B. J. Green, and D. N. Hart. 2000. The CMRF-35H gene structurepredicts for an independently expressed member of an ITIM/ITAM pair ofmolecules localized to human chromosome 17. Tissue Antigens 55: 101–109.
22. Clark, G. J., B. Cooper, S. Fitzpatrick, B. J. Green, and D. N. Hart. 2001. Thegene encoding the immunoregulatory signaling molecule CMRF-35A localizedto human chromosome 17 in close proximity to other members of the CMRF-35family. Tissue Antigens 57: 415–423.
23. Clark, G. J., S. Fitzpatrick, B. Kuo, C. Modra, L. Jamriska, and D. N. Hart. 2002.CMRF-35A, CMRF-35H: potential new CD. J. Biol. Regul. Homeost. Agents 16:233–235.
24. Yoneda, N., E. Tatsumi, S. Kawano, K. Teshigawara, T. Oka, M. Fukuda, andN. Yamaguchi. 1992. Detection of Epstein-Barr virus genome in natural-killer-like cell line, YT. Leukemia 6: 136–141.
25. Achdout, H., I. Manaster, and O. Mandelboim. 2008. Influenza virus infectionaugments NK cell inhibition through reorganization of major histocompatibilitycomplex class I proteins. J. Virol. 82: 8030–8037.
26. Cohen, G. B., R. T. Gandhi, D. M. Davis, O. Mandelboim, B. K. Chen,J. L. Strominger, and D. Baltimore. 1999. The selective downregulation of class Imajor histocompatibility complex proteins by HIV-1 protects HIV-infected cellsfrom NK cells. Immunity 10: 661–671.
27. Davis, D. M., I. Chiu, M. Fassett, G. B. Cohen, O. Mandelboim, andJ. L. Strominger. 1999. The human natural killer cell immune synapse. Proc.Natl. Acad. Sci. USA 96: 15062–15067.
28. Mandelboim, O., P. Malik, D. M. Davis, C. H. Jo, J. E. Boyson, andJ. L. Strominger. 1999. Human CD16 as a lysis receptor mediating direct naturalkiller cell cytotoxicity. Proc. Natl. Acad. Sci. USA 96: 5640–5644.
29. Mandelboim, O., H. T. Reyburn, M. Vales-Gomez, L. Pazmany, M. Colonna,G. Borsellino, and J. L. Strominger. 1996. Protection from lysis by naturalkiller cells of group 1 and 2 specificity is mediated by residue 80 in humanhistocompatibility leukocyte antigen C alleles and also occurs with empty majorhistocompatibility complex molecules. J. Exp. Med. 184: 913–922.
30. Bachelet, I., A. Munitz, A. Moretta, L. Moretta, and F. Levi-Schaffer. 2005. Theinhibitory receptor IRp60 (CD300a) is expressed and functional on humanmast cells. J. Immunol. 175: 7989–7995.
31. Jackson, D. G., D. N. Hart, G. Starling, and J. I. Bell. 1992. Molecular cloning ofa novel member of the immunoglobulin gene superfamily homologous to thepolymeric immunoglobulin receptor. Eur. J. Immunol. 22: 1157–1163.
32. Daish, A., G. C. Starling, J. L. McKenzie, J. C. Nimmo, D. G. Jackson, andD. N. Hart. 1993. Expression of the CMRF-35 antigen, a new member of theimmunoglobulin gene superfamily, is differentially regulated on leucocytes. Im-munology 79: 55–63.
33. Green, B. J., G. J. Clark, and D. N. Hart. 1998. The CMRF-35 mAb recognizesa second leukocyte membrane molecule with a domain similar to the poly Igreceptor. Int. Immunol. 10: 891–899.
34. Brumbaugh, K. M., B. A. Binstadt, and P. J. Leibson. 1998. Signal transductionduring NK cell activation: balancing opposing forces. Curr. Top. Microbiol.Immunol. 230: 103–122.
The Journal of Immunology 2885
by guest on September 25, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

35. Held, W., J. D. Coudert, and J. Zimmer. 2003. The NK cell receptor repertoire:formation, adaptation and exploitation. Curr. Opin. Immunol. 15: 233–237.
36. Stern, N., G. Markel, T. I. Arnon, R. Gruda, H. Wong, S. D. Gray-Owen, andO. Mandelboim. 2005. Carcinoembryonic antigen (CEA) inhibits NK killing viainteraction with CEA-related cell adhesion molecule 1. J. Immunol. 174: 6692–6701.
37. Stanietsky, N., H. Simic, J. Arapovic, A. Toporik, O. Levy, A. Novik, Z. Levine,M. Beiman, L. Dassa, H. Achdout, et al. 2009. The interaction of TIGIT withPVR and PVRL2 inhibits human NK cell cytotoxicity. Proc. Natl. Acad. Sci.USA 106: 17858–17863.
38. Ju, X., M. Zenke, D. N. Hart, and G. J. Clark. 2008. CD300a/c regulate type Iinterferon and TNF-alpha secretion by human plasmacytoid dendritic cells stim-ulated with TLR7 and TLR9 ligands. Blood 112: 1184–1194.
39. Clark, G. J., M. Rao, X. Ju, and D. N. Hart. 2007. Novel human CD4+T lymphocyte subpopulations defined by CD300a/c molecule expression. J.Leukoc. Biol. 82: 1126–1135.
40. Alvarez, Y., X. Tang, J. E. Coligan, and F. Borrego. 2008. The CD300a (IRp60)inhibitory receptor is rapidly up-regulated on human neutrophils in response to
inflammatory stimuli and modulates CD32a (FcgammaRIIa) mediated signaling.
Mol. Immunol. 45: 253–258.41. Munitz, A., I. Bachelet, R. Eliashar, A. Moretta, L. Moretta, and F. Levi-
Schaffer. 2006. The inhibitory receptor IRp60 (CD300a) suppresses the effects
of IL-5, GM-CSF, and eotaxin on human peripheral blood eosinophils. Blood
107: 1996–2003.42. Alvarez-Errico, D., H. Aguilar, F. Kitzig, T. Brckalo, J. Sayos, and M. Lopez-
Botet. 2004. IREM-1 is a novel inhibitory receptor expressed by myeloid cells.
Eur. J. Immunol. 34: 3690–3701.43. Bachelet, I., A. Munitz, D. Mankutad, and F. Levi-Schaffer. 2006. Mast cell
costimulation by CD226/CD112 (DNAM-1/Nectin-2): a novel interface in the
allergic process. J. Biol. Chem. 281: 27190–27196.44. Speckman, R. A., J. A. Wright Daw, C. Helms, S. Duan, L. Cao, P. Taillon-Miller,
P. Y. Kwok, A. Menter, and A. M. Bowcock. 2003. Novel immunoglobulin
superfamily gene cluster, mapping to a region of human chromosome 17q25,
linked to psoriasis susceptibility. Hum. Genet. 112: 34–41.
2886 EXPRESSION AND FUNCTION OF CD300 IN NK CELLS
by guest on September 25, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from




























