Embed Size (px)
Citation preview

T H E JOURNAL OF BIOLOGICAL CHEMISTRY ‘ c 1992 by The American Society for Blochemistry and Molecular Biology , Inc
Vol. 267, No. 17, Issue of June IS, pp. 12082-12089, 1992 Printed in U S A
Expression Cloning of @1,4 N-Acetylgalactosaminyltransferase cDNAs That Determine the Expression of G M z and GD2 Gangliosides*
(Received for publication, December 17, 1991)
Yasuhiko NagataS, Shuji YamashiroS, Junji Yodois, Kenneth 0. Lloydll, Hiroshi ShikuS, and Koichi FurukawaSII From the $Department of Oncology, Nagasaki University School of Medicine, Nagasaki 852, Japan, the §Institute for Virus Research, Kyoto University, Kyoto 606, Japan, and the 7Sloan-Kettering Cancer Center, New York, New York 10021
Gwn and G D ~ gangliosides are sialic acid-containing glycosphingolipids expressed in some normal tissues such as brain and in various tumors such as neuroblas- tomas, astrocytomas, and malignant melanomas. We used a eukaryotic cell transient expression system to isolate cDNA clones that determine GMz expression. We developed a new cell line from murine melanoma line B16 by transfecting with the polyoma T antigen gene that was suitable for this purpose. Two cDNA clones, both of which have a continuous open reading frame of 1683 base pairs, were isolated. Although the cloned cDNAs had no primary sequence similarity to reported glycosyltransferases, the deduced amino acid sequence predicted a type I1 transmembrane protein with an overall structure similar to other glycosyltransferases. The cDNA clones, when stably transfected, determined the expression of GMz in B16 cells and G M Z and GDz in the human melanoma line MeWo. Northern blot analy- sis revealed two transcripts in all cells that expressed either G M ~ or Gp2 or both. These findings indicate that the cDNAs catalyze the transfer of GalNAc onto GM3 and GD:s by a @1,4 linkage, resulting in the synthesis of GMv and GDZ, respectively. Namely they suggest that these cDNAs derive from the UDP-GalNAc: G M ~ / G D ~ @l,4 N-acetylgalactosaminyltransferase (EC 2.4.1.92) gene.
The roles of glycoprotein and glycolipid sugar chains in biological recognition have become increasingly appreciated (1, 2). In particular, changes in gangliosides, which are gly- cosphingolipids containing sialic acid in their carbohydrate moiety, have been intensively studied with regard to cell differentiation, development, and malignant transformation (3-5). Although they have been considered as receptors for exogenous agents such as toxins and viruses and also act as antigens, their true biological function is poorly understood. Two general functions for gangliosides have been studied; their ability to modify the function of transmembrane signal transducers, resulting in regulation of cell proliferation and
* This work was supported by a grant-in-aid for Scientific Research
the Ministry of Health and Welfare of Japan, and by Grants CA from the Ministry of Education, Science and Culture, by a grant from
21445 and CA 08478 from the United States Public Health Service (National Institutes of Health). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore he hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
to the GenBank‘rM/EMBL Data Bank with accession numberfs) The nucleotide sequencefs) reported in thispuper has been submitted
M836FjI. 11 To whom correspondence should he addressed. Tel.: 958-47-2111
(ext. 4800); Fax: 958-49-3695.
their role as mediators for cell-cell or cell-substratum recog- nition (1, 5 ) .
The biological roles of carbohydrates as mentioned above are considered to be controlled by the cell type specific expres- sion of each structure under the regulation of glycosylation machinery (6). It is generally believed that the terminal glycosylation sequences produced by a cell are determined by the specificity of glycosyltransferases present in the intracel- lular Golgi apparatus. Namely, cell type-specific glycosylation may be due to differential expression of the 100-150 different glycosyltransferases estimated to be required for the synthesis of known carbohydrate structures on glycolipids and glyco- proteins (7).
DNA-mediated transfer of expression of carbohydrate an- tigens has been described previously (8, 9) . Recently several glycosyltransferase cDNAs have been cloned (7, lO-l2), and the regulation of cell type-specific expression of glycosyltrans- ferases is being intensively investigated (13). However, the cDNA of glycosyltansferases playing major roles in the syn- thesis of gangliosides have not yet been cloned. It is, therefore, important to isolate the genes coding for these enzymes and to analyze the regulation of their expression for further un- derstanding of the biological roles of gangliosides.
In order to clone the cDNA of glycosyltransferases, we used a eukaryotic cell expression cloning system using a derivative of the mouse melanoma line, B16, as the recipient cell. By means of this system, cDNAs coding the gene which deter- mines the expression of GM2’ and GD2 ganglioside were isolated from a cDNA library prepared from mRNA of the YT lymph- oid cell line. Analysis of the newly expressed gangliosides in transfected cell lines indicates that they can catalyze the transfer of N-acetylgalactosamine by a p1,4 linkage onto both GMic and Gu:i resulting in the generation of GW and G D ~ , respectively. These findings indicate that this cDNA codes for the UDP-GalNAc: GM:?/Gui3 p1,4 N-acetylgalactosaminyl- tranferase (GalNAc-T) (EC 2.4.1.92) gene.
MATERIALS AND METHODS
cDNA Library-A cDNA library prepared from poly(A)’ RNA isolated from a human NK-like cell line YT, using the procedure of
’ The abbreviations used are: Ganglioside nomenclature is based on that of Svennerholm (62 ) : G,,, GalNAcp1-+4 (NeuAcaB--t3) GallJl-4 Glc-Cer; Gw.<, NeuAcaZ-3 Ga181-4 Glc-Cer; GI)<, NeuAca2-8 NeuAcn2+3 Galp1-4Glc-Cer; Gw, GalNAcp1+4 (NeuAca2-8 NeuAcaB”t3) Galpl“t4 Glc-Cer. Other glycolipids were: gangliotriaosylceramide (asialo-Ghd, GalNAcpl+4 Gal@l+4 Glc-Cer; NeuAc, N-acetylneuraminic acid mAb, monoclonal anti- body; TLC, thin layer chromatography; FCM, flow cytometry; GalNAc-T, N-acetylgalactosaminyltransferase. Le‘, Fucnl- ZGal@l-4 (Fucml“t3) GlcNAclJl-R; Le”, Ga181+4 (Fuccul-9) GlcNAc$l-+R; IF, immunofluorescence; FITC, fluorescein isothio- cyanate; SDS, sodium dodecyl sulfate; hp, base paids); kb, kilobase pair(s).
12082

Expression Cloning of G M 2 / G D 2 Synthase 12083
Seed et al. (14, 15), was a gift from Drs. M. Hatakeyama and T. 'I'aniguchi of Osaka University. The mammalian expression vector pCDM8 was modified by deleting the M13 ori from the original vector. The library contained 5 X 10' independent colonies. The strain of bacterial host used was MC1061/P3 as described previously (14).
Cell Lines-All human melanoma and neuroblastoma lines were obtained from Dr. L. J. Old at the Sloan-Kettering Cancer Center, New York and maintained in Dulbecco's modified Eagle's minimal essential medium containing 7.5% fetal bovine serum. The B16 mouse melanoma (clone 78) was a gift from Dr. A. Albino (Sloan-Kettering Cancer Center), and the mouse melanoma JB-RH was provided by Dr. P. 0. Livingston (Sloan-Kettering Cancer Center). They were also maintained in Dulbecco's modified Eagle's minimal essential medium supplemented with 7.5% fetal bovine serum. The mouse melanoma line, designated KF3027, used as a recipient cell in the transient expression system was prepared by cotransfection of B16 melanoma line with pd13027 plasmid containing polyoma T antigen gene (16) (a gift from Dr. C. Basilico, New York University, New York) and pSV2 neo. KF3027 was selected from neo-resistant trans- lectants by screening with a rat anti-polyoma T serum presented by Dr. C. Basilico. The cells were fixed in 60-well microplates with methanol/acetone (1:l) a t -20 "C for 15 min. Expression of T antigen was detected by an indirect immunofluorescence (IF) assay. Positive clones were subcloned and maintained in Dulbecco's modified Eagle's minimal essential medium containing 7.5% fetal bovine serum and (;418 (750 pglml). The usefulness of the transfectant, KF3027, in transient expression system with pCDM8 vector was confirmed by transfection of LFA-3 cDNA in CDM8 vector presented by Dr. B. Seed and detection by anti-LFA-3 monoclonal antibody (mAb), TS2/ 9 (a gift from Dr. T. Springer, Harvard Medical School).
Flow Cytometry Analysis-Ganglioside expression was analyzed using mouse mAb 10-11 (anti-GM2; Ref. 17) given by Dr. P. 0. Livingston, mAb 3F8 (anti-Gpl; Ref. 18) given by Dr. N. K. Cheung (Sloan-Kettering Cancer Center), mAb R24 (anti-GD?; Ref. 19), and M2590 (anti-G,,,; Ref. 20; Meiji Seika Co.). Cells were incubated with mAbs for 45 min on ice and stained with FITC-conjugated goat anti- mouse IgM (heavy and light) (Cappel) for mAbs 10-11 and M2590 or with FITC-labeled protein A (E-Y Labs.) for mAbs 3F8 and R24. Cells were analyzed by flow cytometry on a FACScan (Becton Dick- inson, Mountain View, CA). Cell staining was measured in arbitrary units as the log of fluorescent intensity and displayed on a 4 decade scale.
Isolation of a Human @1,4 N-AcetylgalactosaminyltransferasecDNA ('lone-Plasmid DNA was prepared (21) from an amplified portion of the library and was transfected into KF3027 cells using DEAE- dextran (22). Forty samples of 1.2 X 10' KF3027 cells in 10-cm dishes (Corning) were transfected with 12 pg of plasmid DNA. After 64-72 h, the transfected KF3027 cells were harvested and incubated with mAb 10-11 a t a 1:200 dilution on ice for 45 min. After washing twice, reactive cells were plated on dishes coated with goat anti-mouse IgG (heavy and light) (Zymed) according to the method of Wysocki and Sato (23). Plasmid DNA was rescued from adherent cells by Hirt extraction (24) and transformed into MC1061/P3 by electroporation. Plasmid DNA was prepared from these transformants and transfected again by the same procedure, which was repeated four times. There- after 16 pools containing 10 colonies each were prepared and screened by the expression of mAb 10-11 binding activity. Finally 20 clones l'rom two positive pools were screened, and two single colonies were isolated using microscale transfection and IF assay.
DNA Sequencing-Single-stranded DNA was prepared using M13 phage containing cDNA inserts of various lengths generated with a deletion kit (Takara Shuzo Corp.). Dideoxynucleotide termination sequencing reactions (25) were performed using M13 single-stranded DNA, with -21 M13 Dye Primers, for the Applied Biosystems model 370A DNA sequencing system (AB1 Corp.). Dideoxynucleotide ter- mination sequencing using the Sequenase kit (United States Bio- chemical Corp.) and '"S-dCTP (26) was also performed to confirm the results. SDC Genetyx Sequence Analysis Software was used for sequence analysis.
Southern Blot Hybridization-Genomic DNA was prepared using standard procedures (27). Ten micrograms of genomic DNA was digested overnight with the appropriate restriction endonuclease and
denatured (30 min) in 0.5 N NaOH and 1.5 M NaC1, neutralized (30 loaded onto a 0.8% agarose gel. After electrophoresis, the gels were
min) in 0.5 M Tris-HCI (pH 7.5) and 3 M NaC1, and then the DNA was transferred onto a Nylon membrane (Genescreen Plus, Du Pont- New England Nuclear).
Filters were prehybridized in 50% formamide, 5 X SSC, 5 X Denhardt's, and 0.1% SDS solution at 42 "C for 2 h and then hybrid- ized overnight at 42 "C with a probe of XbaI fragments from the pMZT1-1 insert labeled with using the random priming method (Amersham). Filters were washed in 2 X SSC, 0.1% SDS at room temperature twice, and then in 1 x SSC, 0.1% SDS at 68 "C for 1 h. The membrane was exposed to XAR-5 film (Kodak, Rochester, NY) for 24 h a t -70 "C.
Northern Blot Hybridization-RNA poly(A)+ was isolated by the method of Badley et al. (28). Briefly, cell pellets were lysed in 0.2 M Tris-HCI (pH 7.5) containing 0.2 M NaC1, 1.5 mM MgCl,, 2% SDS, and 200 wg/ml of proteinase K. The extracts were mixed with oligo(dT)-cellulose in 0.01 M Tris-HCI (pH 7.5) containing 0.5 M NaC1. After incubation for 20 min a t room temperature, the beads were washed and packed in a diethylpyrocarbonate-treated column. The packed column was washed until the A26(, of the eluates reached less than 0.05. Poly(A)' mRNA was eluted by 0.01 M Tris-HC1 (pH 7.5). Formaldehyde-denatured RNA was separated in an agarose gel and transferred to a Nylon membrane (Genescreen Plus, Du Pont- New England Nuclear). After baking, the membrane was prebybrid- ized for 3 h and hybridized with XbaI fragment containing 2.7k bp of cDNA labeled by random priming (Amersham) a t 42 "C for 15 h. The membrane was washed twice with 2 X SSC a t room temperature, 2 X SSC, and 1% SDS at 60 "C twice and finally once with 0.1 X SSC at room temperature, and then exposed to a XAR-5 film for 1-3 days at -70 "C.
Extraction of Glycolipids and TLC Immunostaining-Stable trans- formants of the @1,4 N-acetylgalactosaminyltransferase gene were obtained by cotransfecting B16 melanoma with the cloned plasmid and pSV2neo by calcium phosphate precipitation (29). Among neo- resistant clones, GM2-expressing clones were selected based upon the results of IF assay and then subcloned by limiting dilution. Glyco- lipids were isolated as described previously (30). Briefly, cells were extracted from about 1 ml of packed cells of transfectants and control clones containing the neo gene alone using chloroform/methanol(2:1, 1:1, 1:2) sequentially. After acetylation, the glycolipid fraction was isolated using a Florisil column. After deacetylation and desalting, gangliosides were isolated by DEAF-Sephadex A-50 (Pharmacia LKB) ion exchange chromatography. Thin layer chromatography was performed on high performance TLC plates (Merck) using the chlo- roform/methanol (C/M)/2.5 N NHdOH (60:35:8). The components were visualized by spraying with resorcinol.
The identity of new gangliosides was confirmed by TLC immuno- staining using aluminum-backed silica plates (Merck) as described previously (30). After TLC, the plate was air-dried and fixed with polyisobutylmethacrylate in hexane. After blocking in 2% bovine serum albumin in phosphate-buffered saline for 2 h, the plates were incubated with mAbs for 2 h at room temperature and then with rabbit anti-mouse IgG (heavy and light) (Cappel) for 1.5 h. Finally, '2sI-conjugated protein A was added to detect reactive bands. Washed and dried plates were exposed to XAR-5 film (Kodak) for 12 h at -70 "C.
RESULTS
Cloning Approach-To clone the genes coding for the gly- cosyltransferases involved in ganglioside biosynthesis we used the eukaryotic gene expression system of Seed et al. (14, 15) that allowed us to isolate cDNA clone without the need to purify the enzymes first.
This approach requires a recipient host cell with specific properties that allow selection of the appropriate cDNA. The host cell must not express the enzyme to be cloned, nor the cognate surface antigen. However, it must synthesize the appropriate substrates for the enzyme to be cloned. These substrates include the sugar nucleotides and surface-ex- pressed glycoconjugates that may serve as oligosaccharide acceptors for the transglycosylation reaction. As shown in Fig. 1 and Table I, the mouse melanoma line B16 contained abundant G M ~ and a negligible level of GM2 which was essen- tially undetectable in an IF assay and not identified by anti- G M M ~ mAbs. Neither Go:, nor GD, could be detected in B16 by surface serology assay or TLC immunostaining of extracted glycolipids (data not shown). These properties of B16 make it an excellent host recipient cell for cDNA cloning of many
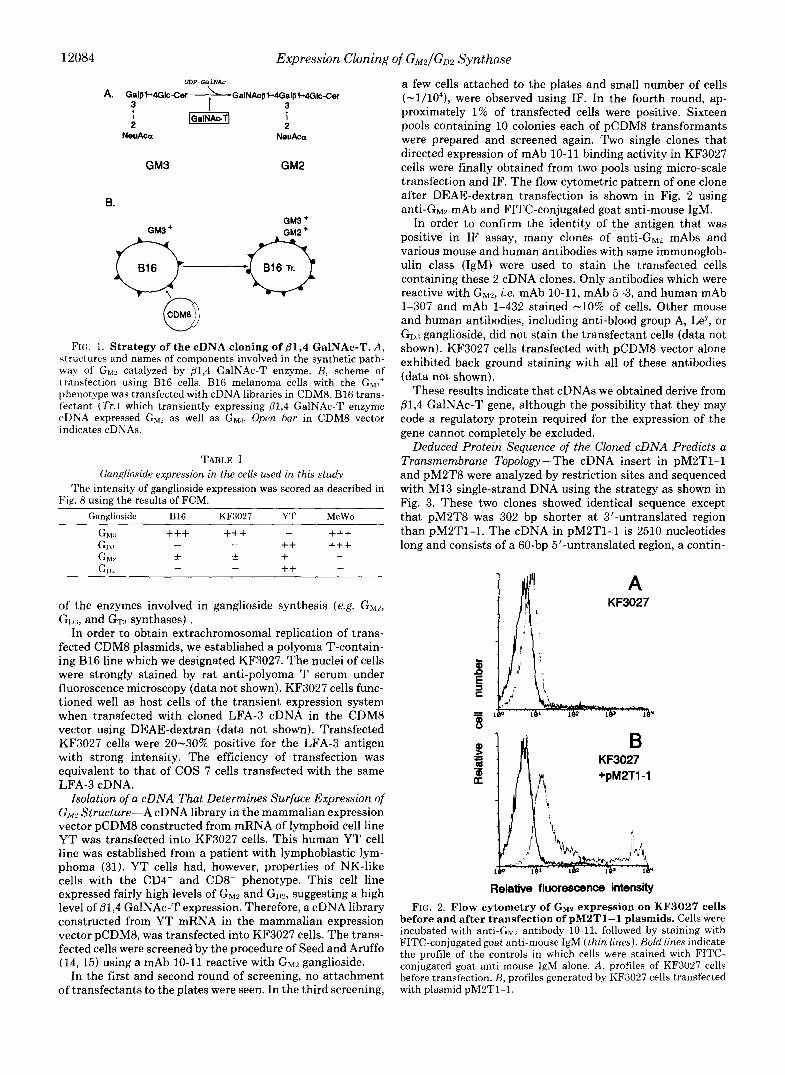
12084 Expression Cloning of G M 2 / G D 2 Synthase
GM3 GM2
B. OM3 +
FIG. 1. Strategy of the cDNA cloning of 01,4 GalNAc-T. A , h~ructures and names of components involved in the synthetic path- way of GW catalyzed by @1,4 GalNAc-T enzyme. B, scheme of transfection using B16 cells. B16 melanoma cells with the GM:~' phenotype was transfected with cDNA libraries in CDMS. B16 trans- lectant (Tr . ) which transiently expressing @1,4 GalNAc-T enzyme cDNA expressed G,, as well as GM.~. Open bar in CDM8 vector indicates cDNAs.
TABLE I Ganglioside expression in the cells used in this study
The intensity of ganglioside expression was scored as described in Fig. 8 using the results of FCM.
Ganglioside B16 KF3027 YT MeWo
of the enzymes involved in ganglioside synthesis (e.g. GMZ, GI),,, and GTTI synthases) .
In order to obtain extrachromosomal replication of trans- fected CDMS plasmids, we established a polyoma T-contain- ing B16 line which we designated KF3027. The nuclei of cells were strongly stained by rat anti-polyoma T serum under fluorescence microscopy (data not shown). KF3027 cells func- tioned well as host cells of the transient expression system when transfected with cloned LFA-3 cDNA in the CDM8 vector using DEAE-dextran (data not shown). Transfected KF3027 cells were 20-30% positive for the LFA-3 antigen with strong intensity. The efficiency of transfection was equivalent to that of COS 7 cells transfected with the same
Isolation of a cDNA That Determines Surface Expression of G,, Structure-A cDNA library in the mammalian expression vector pCDM8 constructed from mRNA of lymphoid cell line YT was transfected into KF3027 cells. This human YT cell line was established from a patient with lymphoblastic lym- phoma (31). YT cells had, however, properties of NK-like cells with the CD4- and CDS- phenotype. This cell line expressed fairly high levels of GM2 and GD?, suggesting a high level of p1,4 GalNAc-T expression. Therefore, a cDNA library constructed from YT mRNA in the mammalian expression vector pCDM8, was transfected into KF3027 cells. The trans- fected cells were screened by the procedure of Seed and Aruffo (14, 15) using a mAb 10-11 reactive with Gw2 ganglioside.
In the first and second round of screening, no attachment of transfectants to the plates were seen. In the third screening,
LFA-3 cDNA.
a few cells attached to the plates and small number of cells (-1/104), were observed using IF. In the fourth round, ap- proximately 1% of transfected cells were positive. Sixteen pools containing 10 colonies each of pCDM8 transformants were prepared and screened again. Two single clones that directed expression of mAb 10-11 binding activity in KF3027 cells were finally obtained from two pools using micro-scale transfection and IF. The flow cytometric pattern of one clone after DEAE-dextran transfection is shown in Fig. 2 using anti-GM2 mAb and FITC-conjugated goat anti-mouse IgM.
In order to confirm the identity of the antigen that was positive in IF assay, many clones of anti-Gw2 mAbs and various mouse and human antibodies with same immunoglob- ulin class (IgM) were used to stain the transfected cells containing these 2 cDNA clones. Only antibodies which were reactive with GMZ, i.e. mAb 10-11, mAb 5-3, and human mAb 1-307 and mAb 1-432 stained -10% of cells. Other mouse and human antibodies, including anti-blood group A, Ley, or Go:i ganglioside, did not stain the transfectant cells (data not shown). KF3027 cells transfected with pCDM8 vector alone exhibited back ground staining with all of these antibodies (data not shown).
These results indicate that cDNAs we obtained derive from p1,4 GalNAc-T gene, although the possibility that they may code a regulatory protein required for the expression of the gene cannot completely be excluded.
Deduced Protein Sequence of the Cloned cDNA Predicts a Transmembrane Topology-The cDNA insert in pM2T1-1 and pM2T8 were analyzed by restriction sites and sequenced with M13 single-strand DNA using the strategy as shown in Fig. 3. These two clones showed identical sequence except that pM2T8 was 302 bp shorter at 3"untranslated region than pM2T1-1. The cDNA in pM2T1-1 is 2510 nucleotides long and consists of a 60-bp 5"untranslated region, a contin-
KF3027
v
KF3027
Relative fluorescence intensity FIG. 2. Flow cytometry of GW expression on KF3027 cells
before and after transfection of pM2TI-1 plasmids. Cells were incubated with anti-GH2 antibody 10-11, followed by staining with FITC-conjugated goat anti-mouse IgM ( thin l ines) . Bold lines indicate the profile of the controls in which cells were stained with FITC- conjugated goat anti-mouse IgM alone. A , profiles of KF3027 cells before transfection. B, profiles generated by KF3027 cells transfected with plasmid pM2T1-1.

Expression Cloning of G M 2 / G D 2 Synthase 12085
pM2T1-1
pM2T8
f- - c- "m - c- 4
f-
f- - - f-
f- 4
f- "m f- &
c- 4 f- f- "m - c- - f- - - c- 4
"m 4
f- "m - f-"m c- 4
FIG. 3. Restriction endonuclease cleavage map and se- quencing strategy of the cloned cDNAs pM2T1-1 and pM2TS. The dotted box represents the open reading frame. The black box indicates untranslated region. Restriction sites are indicated for XbaI ( X b ) , SmaI (Srn), PstI (Ps), Sac1 (Sc ) , EcoRI ( R ) , and KpnI (Kp) .
uous open reading frame of 1683 bp, and a 3"untranslated region of 827 bp that does not contain a poly A tail (Fig. 4). The initiation codon at the beginning of the long open reading frame is embedded within a sequence similar to the Kozak consensus initiation sequence (32). The long reading frame predicts a 561-amino acid protein with a molecular mass of 61,001.89 daltons. Searches of currently available protein and nucleic acid data bases (SWISS-PROT Release 18.0 and EMBL-GDB Release 27.0) identified no other gene with significant sequence similar to this cDNA. Comparison be- tween the amino acid sequence predicted by the insert in pM2T1-1 and different cloned mammalian glycosyltransfer- ases revealed no obvious primary sequence similarities. In- spection and hydropathy analysis of the predicted protein sequence, however, suggested that this protein maintains a similar structural organization to those of known glycosyl- transferases (Fig. 5). There is a single hydrophobic segment near the amino terminus which is comprised of 18 amino acids and flanked by basic residues. This putative signal- anchor sequence would place 536 amino acids within the Golgi lumen and 7 residues within the cytosolic compartment. N-Acetylgalactosaminyltransferase Can Transfer N-Acetyl-
galactosamine onto Two Distinct Acceptor Structures-Clones pM2T1-1 and pM2T8 were isolated by an enrichment ap- proach using a GMZ ganglioside-specific monoclonal antibody, 10-11. In order to confirm the neo-synthesis of the GM2 structure by this cDNA on a biochemical basis, stable trans- fectants of B16 containing this cDNA were established by cotransfection of suitable cell lines with pSV2neo. From one cDNA transfectant and one neo-resistant clone without GM2 expression, ganglioside fractions were isolated and compared with that from parental B16 cells by TLC. As shown in Fig. 6A, the transfected line expressed high levels of GM2 at an almost equivalent level as GM3, whereas the clone containing the neo gene alone was identical to the parent line. The G M ~ level in the transfectant carrying the cloned cDNA did not necessarily decrease when compared with that in control cells.
Since biochemical studies have indicated that GM2 synthase is identical to GD2 and asialo-GM2 synthases (33), we examined whether or not these clones could convert G D ~ to GD2 as well as G M ~ to GM2. As mentioned above, KF3027 cells, prepared from mouse melanoma B16, contained no G D ~ . Therefore we attempted to establish stable transformants using the human melanoma cell line MeWo. This line contained abundant G D ~ as well as GM3, but no GD2 or GM2, which made it suitable for transfection and to investigate the possible expression of GD2
induced with these GalNAc-T cDNAs. As shown in Fig. 7, FCM analysis revealed that one trans-
fectant expressed high levels of surface GDz as well as GM2, as recognized by mAbs 3F8 and 10-11, respectively. Analysis of extracted ganglioside fractions on TLC revealed intense GM2 and Go2 bands in addition to G M ~ and G D ~ (Fig. 6B). The structures newly synthesized by the transfected cDNA were confirmed in TLC immunostaining using appropriate mAbs (data not shown). Consequently, it was demonstrated that this enzyme transferred N-acetylgalactosamine onto the Gal residue of both NeuAca2+3Ga1/31+4Glc-(G~~) and NeuAca2+8NeuAca2-3Gal/31+ 4Glc-(GD3) structures with the same type of linkage (/31,4).
Northern Blots Revealed two Different Transcripts-As shown in Fig. 8, almost all cells expressing G M ~ and/or GDz expressed two transcripts, one of 5.2 kb and the other of 3.0 kb. Another faint band was detectable in some samples at -2.0 kb, the intensity of which was much lower thanthe other two. The intensity of main transcripts does not exactly cor- respond to that of ganglioside expression assayed by FCM. This is partly because the ganglioside expression was scored simply by the percent of positive cells without considering the fluorescence intensity of positive cells. Plasmid pM2T1-1 was the longest cDNA (2.5 kb) among the clones selected and contained neither a poly(A) tail nor a poly(A) signal. There- fore, we have not yet determined which transcript corresponds with these clones. Although the SK-MEL-37 mRNA had a more intense band compared with the upper, both transcripts were equally intense in all the other cell lines. The significance and the regulation of these two transcripts remain to be investigated.
Southern Blot Analysis-Genomic DNA from two human and two mouse cell lines was Southern blotted. The XbaT fragment of pM2T1-1 was used as a probe after 32P labeling. As shown in Fig. 9, this probe hybridized not only human but with mouse DNA also, although with a different profile. Human placental DNA hybridized in the same manner, indi- cating that there was no rearrangement of the genomic se- quence (data not shown).
DISCUSSION
The cloning of mammalian glycosyltransferases has been achieved mainly by two approaches: (i) using synthetic oli- gonucleotides predicted from partial amino acid sequences of soluble, extractable, or pure enzyme (34-36) and (ii) using antibodies reactive with the enzyme molecules, to select col- onies or plaques containing the cDNA of the enzyme in an appropriate expression vector system (37-39).
These methods are, however, not applicable to crude, insol- uble, or nonextractable enzymes. Cloning methods that do not require enzyme purification have recently been successful (11, 12, 40, 41). Although sugar sequences are not the direct products of glycosyltransferase genes, the carbohydrate struc- tures generated by the enzymes could be expressed on the cells and detected with appropriate antibodies or lectins. Thus, this approach allows the detection of cell surface oligo- saccharide products of these enzymes and provides for specific assay of their enzymatic activity. Isolation of an enzyme gene is, however, possible only when the antigen epitope can be made in a cell by transfer and expression of single gene. A few glycosyltransferases have been cloned by cDNA expres- sion cloning in COS cells already (10-12, 40).
We also adopted the mammalian cDNA expression cloning system developed by Seed et al. (14,15) to develop this system, because it would facilitate rescue of the relevant transfected cDNAs. We believed that transfer and expression of the

12086 Expression Cloning of GM2/GI12 Synthase
TPCA""" F T A R T R L E R L V D V L E R T P L D L V G G A V R E I S G F A T T Y R O L L 361
-CCAC- 1200
S V E P G A P G L G J J C S G K G A A S T T S S S A S O A A W S P T A W L T S S W h m - C C K U = P " l T M V 1320
401
CGCajA"""rn" R G L T R C A R S V S T P P O P R G S S G I L L G W A W F P S G W L L L R R R G
-- 1440 441
-CACmm""" L L Q T P A A V H D L P V M A R W G F L T V R L G L P P C P C O E F P T N P T T
TIXCMCAAACCCCACCACC 1680
521
FIG. 4. DNA and deduced amino acid sequence of the cDNA insert in pM2Tl-1. The putative transmembrane domain is denoted hy a double underline. Potential N-glycosylation sites are underlined.
functional cDNAs that determine p1,4 GalNAc-T expression should be detected by virtue of de nouo surface expression of oligosaccharide structures produced by the enzymes. However, COS cells could not be used as the recipient to isolate @1,4 GalNAc-T cDNA, because GM2 and GD2, which are the prod- ucts of the enzyme were present in COS cells, suggesting that 01,4 GalNAc-T is already expressed. Moreover, COS cells do not express much G M ~ , which is a GM2 precursor and which would be used as p1,4 GalNAc-T acceptor. As shown in Table I, the mouse melanoma cell line B16 expresses almost exclu-
sively GM3 ganglioside. In order to obtain sufficient expression of ganglioside structure on the cell surface, B16 cells were transfected with pd13027 containing the polyoma T antigen gene (42). The presence of polyoma T antigen would allow the replication of transiently transfected CDM8-cDNA li- braries and the expression of very high levels of products of the enzymatic synthesis, since the vector has polyoma ori as well as SV-40 ori (42). The B16 variant, KF3027, was useful for our purpose. The ganglioside profile was suitable and the efficiency of transfection, extrachromosomal replication of
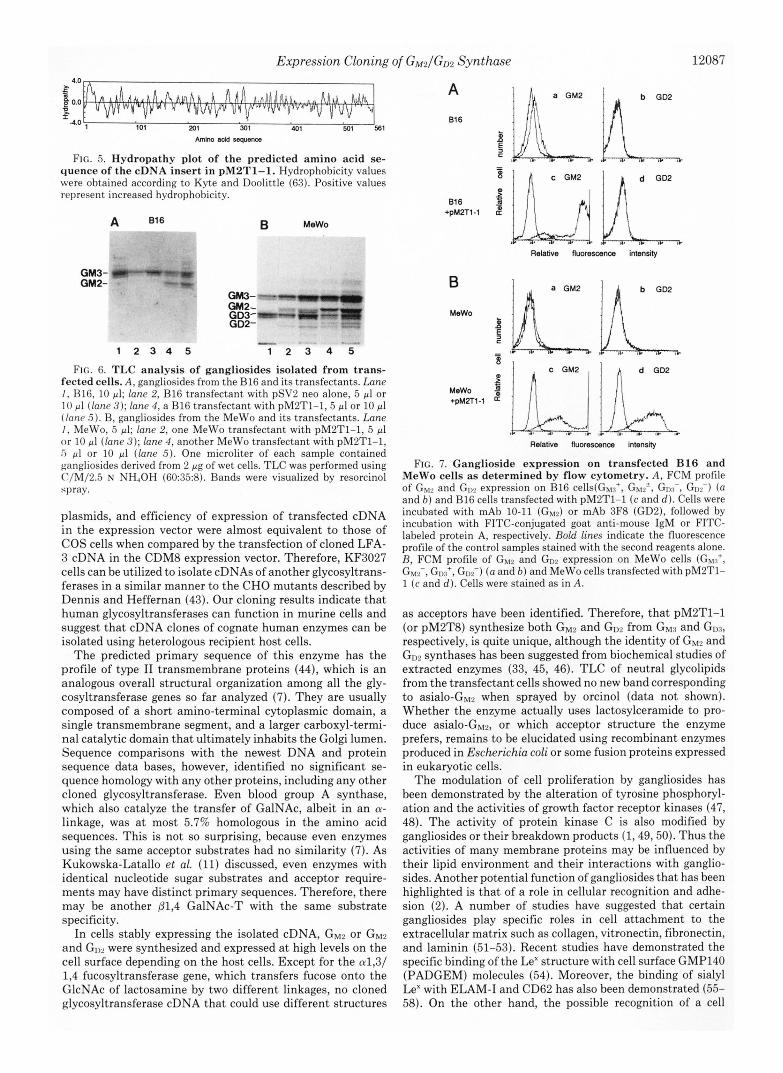
Expression Cloning of G M 2 / G D 2 Synthase 12087
I 101 201 301 401 501 Ea1
Amh add seqdenm
FIG. 5. Hydropathy plot of the predicted amino acid se- quence of the cDNA insert in pMPT1-1. Hydrophobicity values were obtained according to Kyte and Doolittle (63). Positive values represent increased hydrophobicity.
GM3 GM2
B MeWo
Relative fluorexana, intensity
GM3- GM2- GD3- W- Z GD2-
1 2 3 4 5 -
1 2 3 4 5 , I,
FIG. 6. TLC analysis of gangliosides isolated from trans- fected cells. A, gangliosides from the B16 and its transfectants. Lane I , HlG, 10 pl; lane 2, B16 transfectant with pSV2 neo alone, 5 p1 or 1 0 pl (lane 3); lane 4 , a B16 transfectant with pMLT1-1,5 pl or 10 pl (Inne 5 ) . B, gangliosides from the MeWo and its transfectants. Lane I , MeWo, 5 pl; lane 2, one MeWo transfectant with pM2T1-1, 5 pl o r 10 pl (lane 3); lane 4, another MeWo transfectant with pM2T1-1, 5 pl or 10 pl (lane 5). One microliter of each sample contained gangliosides derived from 2 pg of wet cells. TLC was performed using C/M/2.5 N NH,OH (60:35:8). Bands were visualized by resorcinol spray.
plasmids, and efficiency of expression of transfected cDNA in the expression vector were almost equivalent to those of COS cells when compared by the transfection of cloned LFA- 3 cDNA in the CDM8 expression vector. Therefore, KF3027 cells can be utilized to isolate cDNAs of another glycosyltrans- ferases in a similar manner to the CHO mutants described by Dennis and Heffernan (43). Our cloning results indicate that human glycosyltransferases can function in murine cells and suggest that cDNA clones of cognate human enzymes can be isolated using heterologous recipient host cells.
The predicted primary sequence of this enzyme has the profile of type I1 transmembrane proteins (44), which is an analogous overall structural organization among all the gly- cosyltransferase genes so far analyzed (7). They are usually composed of a short amino-terminal cytoplasmic domain, a single transmembrane segment, and a larger carboxyl-termi- nal catalytic domain that ultimately inhabits the Golgi lumen. Sequence comparisons with the newest DNA and protein sequence data bases, however, identified no significant se- quence homology with any other proteins, including any other cloned glycosyltransferase. Even blood group A synthase, which also catalyze the transfer of GalNAc, albeit in an a- linkage, was a t most 5.7% homologous in the amino acid sequences. This is not so surprising, because even enzymes using the same acceptor substrates had no similarity (7). As Kukowska-Latallo et al. (11) discussed, even enzymes with identical nucleotide sugar substrates and acceptor require- ments may have distinct primary sequences. Therefore, there may be another /31,4 GalNAc-T with the same substrate specificity.
In cells stably expressing the isolated cDNA, GM2 or GM2 and GIy2 were synthesized and expressed at high levels on the cell surface depending on the host cells. Except for the a1,3/ 1,4 fucosyltransferase gene, which transfers fucose onto the GlcNAc of lactosamine by two different linkages, no cloned glycosyltransferase cDNA that could use different structures
Relative RuorescenQ) intensity
FIG. 7. Ganglioside expression on transfected B16 and MeWo cells as determined by flow cytometry. A, FCM profile of Gur and expression on B16 cells(Gy:%+, Gw~', GI", Go,-) ( a and b) and B16 cells transfected with pM2T1-1 (c and d) . Cells were incubated with mAb 10-11 (GM,) or mAb 3F8 (GD2), followed by incubation with FITC-conjugated goat anti-mouse IgM or FITC- labeled protein A, respectively. Bold lines indicate the fluorescence profile of the control samples stained with the second reagents alone. R, FCM profile of Gu2 and GD.' expression on MeWo cells (GM:~', G&, &+, GIp2-) ( a and b ) and MeWo cells transfected with pM2T1- 1 (c and d). Cells were stained as in A .
as acceptors have been identified. Therefore, that pM2T1-1 (or pM2T8) synthesize both GM2 and GD2 from G M ~ and G D ~ , respectively, is quite unique, although the identity of GW and GD2 synthases has been suggested from biochemical studies of extracted enzymes (33, 45, 46). TLC of neutral glycolipids from the transfectant cells showed no new band corresponding to asialo-GM? when sprayed by orcinol (data not shown). Whether the enzyme actually uses lactosylceramide to pro- duce asialo-Gw2, or which acceptor structure the enzyme prefers, remains to be elucidated using recombinant enzymes produced in Escherichia coli or some fusion proteins expressed in eukaryotic cells.
The modulation of cell proliferation by gangliosides has been demonstrated by the alteration of tyrosine phosphoryl- ation and the activities of growth factor receptor kinases (47, 48). The activity of protein kinase C is also modified by gangliosides or their breakdown products (1,49,50). Thus the activities of many membrane proteins may be influenced by their lipid environment and their interactions with ganglio- sides. Another potential function of gangliosides that has been highlighted is that of a role in cellular recognition and adhe- sion (2). A number of studies have suggested that certain gangliosides play specific roles in cell attachment to the extracellular matrix such as collagen, vitronectin, fibronectin, and laminin (51-53). Recent studies have demonstrated the specific binding of the Le" structure with cell surface GMP140 (PADGEM) molecules (54). Moreover, the binding of sialyl Le" with ELAM-I and CD62 has also been demonstrated (55- 58). On the other hand, the possible recognition of a cell
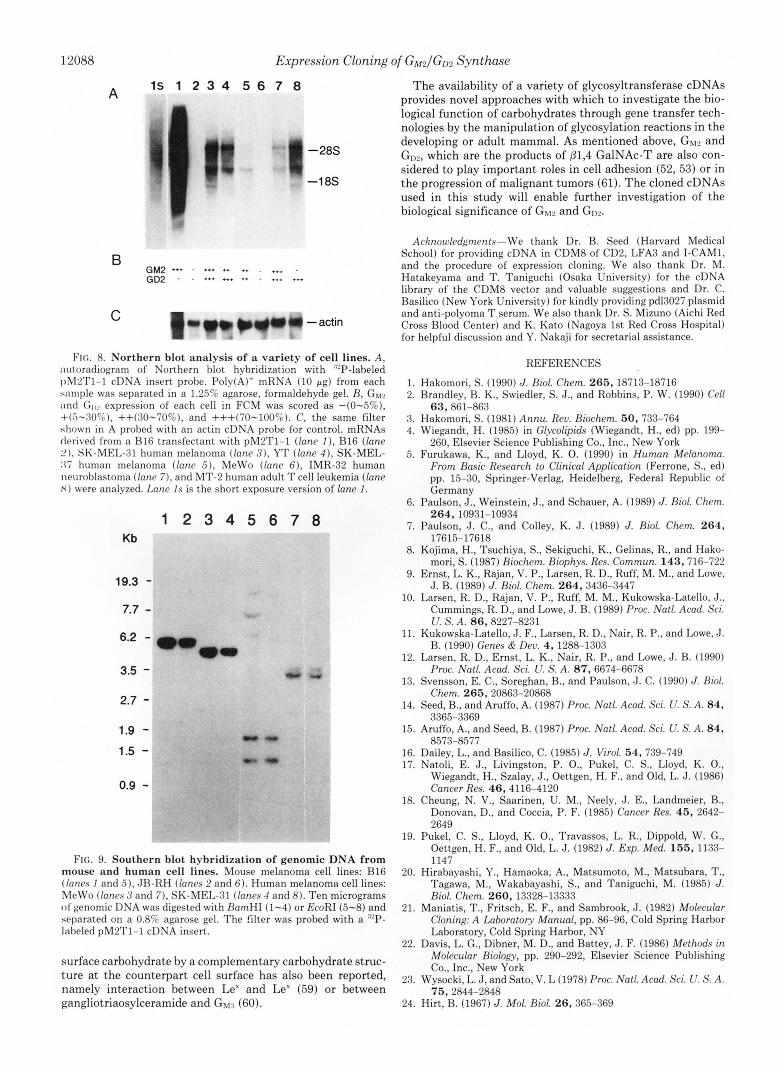
12088
A
B
C
Expression Cloning of GW2/GI,2 Synthase
1s 1 2 3 4 5 6 7 8 The availability of a variety of glycosyltransferase cDNAs provides novel approaches with which to investigate the bio- logical function of carbohydrates through gene transfer tech- nologies by the manipulation of glycosylation reactions in the developing or adult mammal. As mentioned above, GW and
which are the products of @1,4 GalNAc-T are also con- sidered to play important roles in cell adhesion (52, 53) or in
used in this study will enable further investigation of the biological significance of GM2 and Gw.
-28s
-1 8s the progression of malignant tumors (61). The cloned cDNAs
Acknoukdgmmts-We thank Dr. B. Seed (Harvard Medical School) for providing cDNA in CDM8 of CD2, LFA3 and I-CAM1,
GM2 +** - +*+ ++ ++ . ... . and the procedure of expression cloning. We also thank Dr. M. GD2 . . *++ - +++ Hatakeyama and T. Taniguchi (Osaka University) for the cDNA
library of the CDM8 vector and valuable suggestions and Dr. C. Rasilico (New York University) for kindly providing pd13027 plasmid 1" w * m Y - a c t i n and anti-polyoma T serum. We also thank Dr. S. Mizuno (Aichi Red Cross Blood Center) and K. Kat0 (Nagoya 1st Red Cross Hospital) for helpful discussion and Y. Nakaji for secretarial assistance.
FIG. 8. Northern blot analysis of a variety of cell lines. A , autoradiogram of Northern blot hybridization with "'P-labeled I)M2T1-1 cDNA insert probe. Polv(A)+ mRNA (10 pg) from each sample was separated in a 1.25% agarose, formaldehyde gel. R, Gbj2 ;Ind GI,, expression of each cell in FCM was scored as -(0-5%), +(5-:30%), ++(:10-70%), and +++(70-100%). C, the same filter shown in A probed with an actin cDNA probe for control. mRNAs derived from a €316 transfectant with pM2T1-1 (lane I ) , B16 (lane 21, SK-MEL-31 human melanoma (lane 3 ) , YT (lane 4 ) , SK-MEL- :I7 human melanoma (lane ,5), MeWo (lane 6 ) , IMR-32 human neurohlastoma (lane 7), and MT-2 human adult T cell leukemia (lane X ) were analyzed. Lane Is is the short exposure version of lane I .
1 2 3 4 5 6 7 8 Kb
19.3
7.7
6.2
3.5
2.7
1.9
1.5
0.9
FIG. 9. Southern blot hybridization of genomic DNA from mouse and human cell lines. Mouse melanoma cell lines: B16 (/anc.s I and 5 ) , JR-RH (/anes 2 and 6 ) . Human melanoma cell lines: MeWo (lanes 3 and 7), SK-MEL-31 (lanes 4 and 8). Ten micrograms o f genomic DNA was digested with RamHI (1-4) or EcoRI (5-8) and separated on a 0.8% agarose gel. The filter was probed with a '"P- labeled pM2T1-1 cDNA insert.
surface carbohydrate by a complementary carbohydrate struc- ture at the counterpart cell surface has also been reported, namely interaction between Le" and Le" (59) or between gangliotriaosylceramide and GM:, (60).
REFERENCES
1. Hakomori, S. (1990) J. Riol. Chem. 265, 18713-18716 2. Brandley, B. K., Swiedler, S. J., and Robbins, P. W. (1990) Cell
3. Hakomori, S. (1981) Annu. Reo. Biochem. 50, 733-764 4. Wiegandt, H. (1985) in Glycolipids (Wiegandt, H., ed) pp. 199-
260, Elsevier Science Publishing Co., Inc., New York 5. Furukawa, K., and Lloyd, K. 0. (1990) in Human Melanoma.
From Basic Research to Clinical Application (Ferrone, S., ed) pp. 15-30, Springer-Verlag, Heidelberg, Federal Republic of Germany
6. Paulson, J., Weinstein, J., and Schauer, A. (1989) J. Bioi. Chem.
7. Paulson, J. C., and Colley, K. J. (1989) J. Riol. Chem. 264,
8. Kojima, H., Tsuchiya, S., Sekiguchi, K., Gelinas, R., and Hako- mori, S. (1987) Riochem. Riophys. Res. Commun. 143,716-722
9. Ernst, L. K., Rajan, V. P., Larsen, R. D., Ruff, M. M., and Lowe, J. B. (1989) J . Riol. Chem. 264,3436-3447
10. Larsen, R. D., Rajan, V. P., Ruff, M. M., Kukowska-Latello, J., Cummings. R. D., and Lowe, J. B. (1989) Proc. Natl. Acad. Sci.
63,861-863
264,10931-10934
17615-17618
U. S. A . 8 6 , 8227-8231 11. Kukowska-Latello. J. F.. Larsen. R. D.. Nair. R. P.. and Lowe. J .
B. (1990) Genes k De;. 4, 1288-1303
Proc. Natl. Acad. Sci. U. S. A . 87, 6674-6678
Chem. 265,20863-20868
12. Larsen, R. D., Ernst, L. K., Nair, R. P., and Lowe, J. B. (1990)
13. Svensson, E. C., Soreghan, B., and Paulson, J . C. (1990) J. Riol.
14. Seed, B., and Aruffo, A. (1987) Proc. Natl. Acad. Sci. U. S. A. 84,
15. Aruffo, A., and Seed, B. (1987) Proc. Natl. Acad. Sci. U. S. A. 84,
16. Dailey, L., and Basilico, C. (1985) J. Virol. 54, 739-749 17. Natoli, E. J., Livingston, P. O., Pukel, C. S., Lloyd, K. O.,
Wiegandt, H., Szalay, J., Oettgen, H. F., and Old, L. J. (1986) Cancer Res. 46,4116-4120
18. Cheung, N. V., Saarinen, U. M., Neely, J. E., Landmeier, B., Donovan, D., and Coccia, P. F. (1985) Cancer Res. 45, 2642- 2649
19. Pukel, C. S., Lloyd, K. O., Travassos, L. R., Dippold, W. G., Oettgen, H. F., and Old, L. J. (1982) J. Exp. Med. 155, 1133- 1147
20. Hirabayashi, Y., Hamaoka, A., Matsumoto, M., Matsubara, T., Tagawa, M., Wakabayashi, S., and Taniguchi, M. (1985) J. Riol. Chem. 260, 13328-13333
21. Maniatis, T., Fritsch, E. F., and Sambrook, J. (1982) Molecular Cloning: A Laboratory Manual, pp. 86-96, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY
22. Davis, L. G., Dibner, M. D., and Battey, J. F. (1986) Methods in Molecular Biology, pp. 290-292, Elsevier Science Publishing Co., Inc., New York
23. Wysocki, L. J, and Sato, V. L (1978) Proc. Natl. Acad. Sci. U. S. A.
24. Hirt, B. (1967) J . Mol. Bioi. 26, 365-369
3365-3369
8573-8577
75,2844-2848

25.
26.
27.
28.
29.
30.
31.
32. 33.
34.
35.
36.
37.
38.
39.
40.
41.
Expression Cloning Sanger, F., Nicklen, S., and Coulson, A. R. (1977) Proc. Natl.
Acad. Sci. U. S. A. 74,5463-5467 Maniatis, T., Fritsch, E. F., and Sambrook, J. (1982) Molecular
Cloning, A Laboratory Manual, pp. 76-85, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY
Maniatis, T., Fritsch, E. F., and Sambrook, J. (1982) Molecular Cloning, A Laboratory Manual, pp. 280-281, Cold Spring Har- bor Laboratory, Cold Spring Harbor, NY
Badley, J., Bishop, G., Joho, T., and Frelinger, J. (1988) Biotech- niqws 6 , 114-116
Davis, L. G., Dibner, M. D., and Battey, J. F.(1986) Basic Methods in Molecular Biology, pp. 285-289, Elsevier Science Publishing Co., Inc., New York
Furukawa, K., Clausen, H., Hakomori, S., Sakamoto, J., Look, K., Lundblad, A., Mattes, M. J., and Lloyd, K. 0. (1985) Biochemistry 24,7820-7826
Yodoi, J., Teshigawara, K., Nikaido, T., Fukui, K., Noma, T., Honjo, T., Takigawa, M., Sasaki, M., Minato, N., Tsudo, M., Uchiyama, T., and Maeda, M. (1985) J. Immunol. 134, 1623- 1630
Kozak, M. (1984) Nucleic Acid Res. 12, 3873-3893 Pohlentz, G., Klein, D., Schwarzmann, G., Schmitz, D., and
Sandhoff, K. (1988) Proc. Natl. Acad. Sci. U. S. A. 8 5 , 7044- 7048
Narimatsu, H., Sinha, S., Brew, K., Okayama, H., and Quasba, P. K. (1986) Proc. Natl. Acad. Sci. U. S. A. 83,4720-4724
Yamamoto, F., Marken, J., Tsuji, T., White, T., Clausen, H., and Hakomori, S. (1990) J. Biol. Chem. 265, 1146-1151
Sarker, M., Hull, E., Nishikawa, Y., Simpson, R. J., Moritz, R. L., Dunn, D., and Schchater, H. (1991) Proc. Natl. Acad. Sci.
Shaper, N. L., Shaper, J. H., Meuth, J. L., Fox, J. L., Chang, H., Kirsch, I. R., and Hollis, G. F. (1986) Proc. Natl. Acad. Sci.
Joziasse, D. H., Shaper, J. H., Van den Eijnden, D. H., Van Tunen, A. J., and Shaper, N. L (1989) J. Biol. Chem. 2 6 4 ,
Weinstein, J., Lee, E. U., McEntee, K., Lai, P.-H., and Paulson, J . C. (1987) J. Biol. Chem. 262, 17735-17743
Goelz, S. E., Hession, C., Goff, D., Griffiths, B., Tizard, R., Newman, B., Chi-Rosso, G., and Lobb, R. (1990) Cell 63,1349- 1356
Kumar, R., Yang, J., Larsen, R. D., and Stanley, P. (1990) Proc.
U. S. A. 88,234-238
U. S. A. 83, 1573-1577
14290-14297
of G M , / G ~ , Synthase 12089
Natl. Acad. Sci. U. S. A. 87,9948-9952 42. Seed, B. (1987) Nature 329,840-842 43. Heffernan, M., and Dennis, J. W. (1991) Nucleic Acid Res. 19 ,
44. Wickner, W. T., and Lodish, H. F. (1985) Science 230,400-407 45. Cumar, F. A., Tallman, J. F., and Brady, R. 0. (1972) J. Biol.
Chem. 247,2322-2327 46. Pacuszka, T., Duffard, R. O., Nishimura, R. N., Brady, R. O., and
Fishman, P. H. (1978) J. Bwl. Chem. 253,5839-5846 47. Bremer, E., Hakomori, S., Bowen-Pope, D. F., b i n e s , E., and
Ross, R. (1984) J. Biol. Chem. 259,6818-6825 48. Bremer, E., Schlessinger, J., and Hakomori, S. (1986) J. Biol.
Chem. 261,2434-2440 49. Kreutter, D., Kim, J. Y. H., Goldenring, J. R., Rasmussen, H.,
Ukomadu, C., DeLorenzo, R. J., and Yu, R. K. (1987) J. Biol. Chem. 262, 1633-1637
85-92
50. Hannum, Y. A., and Bell, R. M. (1987) Science 236,670-674 51. Kleinman, H. K., Martin, G. R., and Fishman, P. H. (1979) Proc.
Natl. Acad. Sci. U. S. A. 76,3367-3371 52. Cheresh, D. A., Harper, J. R., Schulz, G., and Reisfeld R. A.
(1984) Proc. Natl. Acad. Sci. U. S. A. 81, 5767-5771 53. Cheresh, D. A., Pierschbacher, M. D., Herzig, M. A., and Mujoo,
K. (1986) J. Cell Biol. 102 , 688-696 54. Larsen, E., Palabrica, T., Sajer, S., Gilbert, G., Wagner, D., Furie,
B., and Furie, B. (1990) Cell 6 3 , 467-474 55. Lowe, J., Stoolman, L., Nair, R., Larsen, R., Berhend, T., and
Marks, M. (1990) Cell 63,475-484 56. Phillips, M. L., Nudelman, E., Gaeta, F. C. A., Perez, M., Singhal,
A. K., Hakomori, S., and Paulson, J. C. (1990) Science 250 ,
57. Walz, G., Aruffo, A., Kolanus, W., Bevilacqua, M., and Seed, B. (1990) Science 250, 1132-1135
58. Polley, M. J., Phillips, M. L., Wayner, E., Singhal, A. K., Hako- mori, S., and Paulson, J. C. (1991) Proc. Natl. Acad. Sci. U. S. A.
59. Eggens, I., Fenderson, B., Toyokuni, T., Dean, B., Stroud, M., and Hakomori, S. (1989) J. Biol. Chem. 264 , 9476-9484
60. Kojima, N., and Hakomori, S. (1989) J. Biol. Chem. 264,20159- 20162
61. Thurin, J., Thurin, M., Herlyn, M., Elder, D. E., Steplewski, Z., Clark, W. H., Jr., and Koprowski, H. (1986) FEBS Lett. 208 ,
1130-1132
88,6224-6228
17-22 62. Svennerholm, L. (1963) J. Neurochem. 10 , 613-623 63. Kyte, J., and Doolittle, R. F. (1982) J. Mol. Biol. 157 , 105-132





























