Embed Size (px)
Citation preview

Expression of Extracellular Matrix Molecules and Cell Surface Molecules in
the Olfactory Nerve Pathway During Early Development
QIZHI GONG AND MICHAEL T. SHIPLEY Rockefeller University, New York, New York 10021 (Q.G.); Department of Anatomy,
University of Maryland School of Medicine, Baltimore, Maryland 21201 (M.T.S.)
ABSTRACT The spatial and temporal expression patterns of several extracellular matrix molecules-
laminin and fibronectin and cell surface molecules, neural cell adhesion molecule (NCAM), L1, tenascin, chondroitin sulfate proteoglycan, and peanut agglutinin (PNA) binding sites-were investigated during early olfactory nerve development. NCAM and L1 have similar patterns: They are expressed in the olfactory nerve and on the olfactory receptor neurons (ORNs) commencing with the earliest olfactory axon outgrowth (E12-El5). Their expression patterns suggest that both NCAM and L1 are associated with extension and fasciculation of olfactory axons. A comparison of L1 and olfactory marker protein suggests that L1 is expressed predominantly on immature ORNs. Laminin has an unique punctate staining pattern in the developing olfactory pathway as early as E12. These laminin puncta might play a role in olfactory neurite outgrowth and guidance. At E14, when pioneer olfactory axons enter the brain, the laminin-positive meninges on the surface of the olfactory bulb primordium break down but remain intact in the rest of the telencephalon. This suggests a functional interaction between the olfactory axons and the glial-pial barrier. Fibronectin staining is diffuse throughout the cranial mesenchyme but is absent from the olfactory nerve pathway. No specific patterns of tenascin or chondroitin sulfate, were observed during early olfactory development. PNA binding sites were associated with olfactory axon fasciculation. The expression of several extracellular matrix molecules and cell surface molecules is spatially and temporally regulated in the developing olfactory system. These molecules, thus, may play functional roles in olfactory axon outgrowth, fasciculation, and/or guidance. 8 I W ~ ~ i ~ e y - ~ i s s . Inc
Indexing terms: NCAM, L1, laminin, fibronectin, telencephalon
The development of neuronal connections is a highly regulated process. Axonal growth and guidance are influ- enced by both promoting and inhibitory growth factors (Kapfhammer and Raper, 1987a,b; Patterson, 1988; Silver et al., 1987). Extracellular matrix molecules (ECMs) and cell surface molecules (CSMs) play important roles in these processes (Bixby, 1991; Sanes, 1989). We have previously shown that early olfactory axons grow from the olfactory epithelium to the telencephalon along a highly specific pathway. The initial olfactory axons do not appear to contact olfactory Schwann cells (Gong et al., 1994). There- fore, guidance cues for early axon outgrowth may derive from other cell types. To investigate this issue further, we have characterized the expression patterns of several extra- cellular matrix molecules and cell surface molecules sus- pected of playing a role in axon outgrowth during early olfactory axon pathfinding.
The role of CSMs in nervous system development has been extensively studied. The most thoroughly investigated molecules are neural adhesion molecule (NCAM) and L1 (Appel et al., 1993; Bixby et al., 1988; Jacobson and Rutishauser, 1986; Landmesser et a]., 1988; Lemmon et al., 1992, 1989; Neugebauer et al., 1988; Rutishauser et al., 1983; Silver and Rutishauser, 1984; Thiery et al., 1982). NCAM and L1 are both cell surface glycoproteins that belong to the immunoglobulin superfamily (Cunningham et al., 1987; Moos et al., 1988; Williams and Barclay, 1988). Although they both act through homophilic binding, L1 also has heterophilic binding ability (Appel et al., 1993;
Accepted October 9. 1995. Address reprint requests to Michael T. Shipley, Department of Anatomy,
University of Maryland, School of Medicine, 655 W. Baltimore St., Ralti- more, MD 21201.
o 1996 WILEY-LISS. INC.

2 Q. (;ON(; AND M.T. SHIPLEY
buffer, pH 7.4, for 24 hours. Embryos at E l 6 and older were fixed by transcardiac perfusion using 4% paraformalde- hyde. Embryonic tissues were postfixed overnight. All tissues were cryoprotected in 30% sucrose (4°C J overnight following postfixation.
Immunocytochemistry The brains were placed in embedding matrix (Lipshaw)
and frozen in dry ice powder before sectioning. Parasagittal cryostat sections were cut at 20 pm, collected on chrome- alum-coated slides, air dried for 10 minutes, and then postfixed in 4V paraformaldehyde for 10 minutes at room temperature. After rinsing in phosphate buffer saline (PBS) with 0.2% Triton X-100, the sections were incubated in 0.5% hydrogen peroxide for 10 minutes. Following a I-hour incubation in 2% normal serum (room temperature), sec- tions were incubated in primary antibody overnight at 4°C. Immunoreactivity was visualized with a biotinylated second- ary antibody, and avidinibiotin-peroxidase complex (Vec- tastain Elite kit, Vector Laboratories) reacted with 3,3'- diaminobenzidine. Peanut agglutinin histochemistry was done by incubating biotinylated peanut agglutinin ( 1:400, Vector Labs) with the sections overnight and followed by reaction with avidinibiotin-peroxidase complex (Vec- tastain Elite kit, Vector Labs?. The reaction product was visualized with 3,3'-diaminobenzidine reaction. Sections were dehydrated, cleared in Hemo-De, and coverslipped with DPX. Omission of primary antisera from the incuba- tions was done to control for antibody specificity. Sections were examined by brightfield microscopy.
Bixby et al., 1988; Landmesser et al., 1988; Rutishauser et al., 1983; Silver and Rutishauser, 1984). NCAM has been implicated in multiple functions during neuronal pattern formation. It facilitates axon-axon interaction and fascicu- lation of neurites through homophilic adhesion and has also been shown to promote axonal outgrowth (Chang et al., 1987: Doherty et al., 1990a,b; Grumet et al., 1982). Like NCAM, L1 increases neurite outgrowth on L1-positive partner cells and has been implicated in axonal fascicula- tion (Apple et al., 1993; Changet al., 1987; Rutishauser and Landmesser, 1991). L1 is also important in neuron-glia interactions (Bixby, 1991; Seilheimer and Schachner, 1988).
Extracellular matrix molecules are a diverse class of factors that promote and guide neurite outgrowth (Bixby, 1991; Sanes, 1989). Laminin and fibronectin are abundant components of the extracellular matrix. There is evidence that these two molecules influence neuronal differentia- tion, neurite outgrowth, and neuronal survival (Hammar- back et al., 1985; Jousimaa et al., 1984; Lander et al., 1985; Liesi et al., 1984; Pixley and Cotman, 1986). Laminin produced by astrocytes may be a prerequisite for axonal growth and regeneration in the adult central nervous system (Zhou, 1990).
In a recent report, we defined four stages of the early rat olfactory axon development (Gong and Shipley, 1995): 1) Olfactory axons extend out of the olfactory placode and grow to the telencephalon along a highly stereotypic trajec- tory; 2 ) next, the earliest olfactory axons contact the telencephalon; 3) some of the earliest olfactory axons penetrate to the ventricular mitotic zone of the olfactory primordium-the arrival of these pioneer axons appears to trigger the formation of the olfactory bulb; 4) and finally, the pioneer olfactory axons disappear from the ventricular zone, and the olfactory bulb develops. Together, these findings indicate that early olfactory axon outgrowth is a highly regulated process and that these early axons may play a critical role in the induction of the olfactory bulb. The molecular mechanism(s) of olfactory nerve pathfinding and targeting are not known. Early olfactory axons express growth-associated protein GAP43 (Gongand Shipley, 1995). Previous studies in mouse olfactory system have demon- strated the expression of adhesion molecules (NCAM and L1) on developing olfactory axons (Miragall et a]., 1989). However. the temporal and spatial expression patterns of NCAM and L1 during the earliest stages of olfactory axon outgrowth are not known. Furthermore, the expression patterns of extracellular matrix molecules implicated in axonal outgrowth have not been characterized in detail. To explore possible mechanisms of olfactory axon pathfinding and targeting, we have characterized the spatio-temporal expression patterns of several ECMs and CSMs during olfactory system development.
MATERIALS AND METHODS Animal preparation
Embryos were taken from timed pregnant Sprague- Dawley rats (Zivic Miller?. The presence of a vaginal plug was checked for at 9 am; embryonic day 0 (EOJ was defined as the day the plug was found, and succeeding days of gestation were numbered in order. Embryos were taken from E l 2 to E21. Pregnant female rats were anesthetized with sodium pentobarbital (71.5 mgikg, ip). Embryos at E l 2 to E l 5 were removed from the uterus and fixed by immersion in 4% paraformaldehyde in 0.1 M phosphate
Antibodies NCAM antibody used in this study is a mouse monoclonal
antibody (a generous gift from Dr. R. Akeson), also known as 3F4. The supernatant was diluted to 1:6; The rabbit polyclonal L1 antibody was a generous gift from Dr. V. Lemmon (used at 1:1,000); laminin antibody was used at a dilution of 1:1,000 (rabbit antibody obtained from E Y Laboratories, Inc. j ; A rabbit polyclonal antibody against fibronectin (Collaborative Biomedical Product) was used in this study at a dilution of 1:500; Monoclonal anti-GAP43 (Boehringer and MannheimJ was used at 1:4,000 dilution; Antibody against OMP was a generous gift from Dr. F. Margolis (goat polyclonal, 1:4,000); Tenascin antibody (a gift from Dr. A. Faissner) was used at 1:200; Chondroitin sulfate antibody, clone CS-56, was used at 1:200 (Sigma).
RESULTS ECM and CSM expression pattern in the
developing olfactory nerve The earliest olfactory axons are readily identified by their
robust expression of GAP43. In the following account, the development of early olfactory axons is reiterated briefly to facilitate comparison of stages of axon growth with the expression of ECMs and CSMs (Gong and Shipley, 19951.
El2 GAP43. The first GAP43-positive olfactory axons grow
out of the olfactory placode and head directly toward the telencephalon. When they reach a zone which is = 50 pm away from the telencephalic surface, the axons make a 90" turn and grow toward the most rostra1 part of the telen- cephalon. The olfactory axons do not contact the telence- phalic surface at this time (Fig. 1AJ.
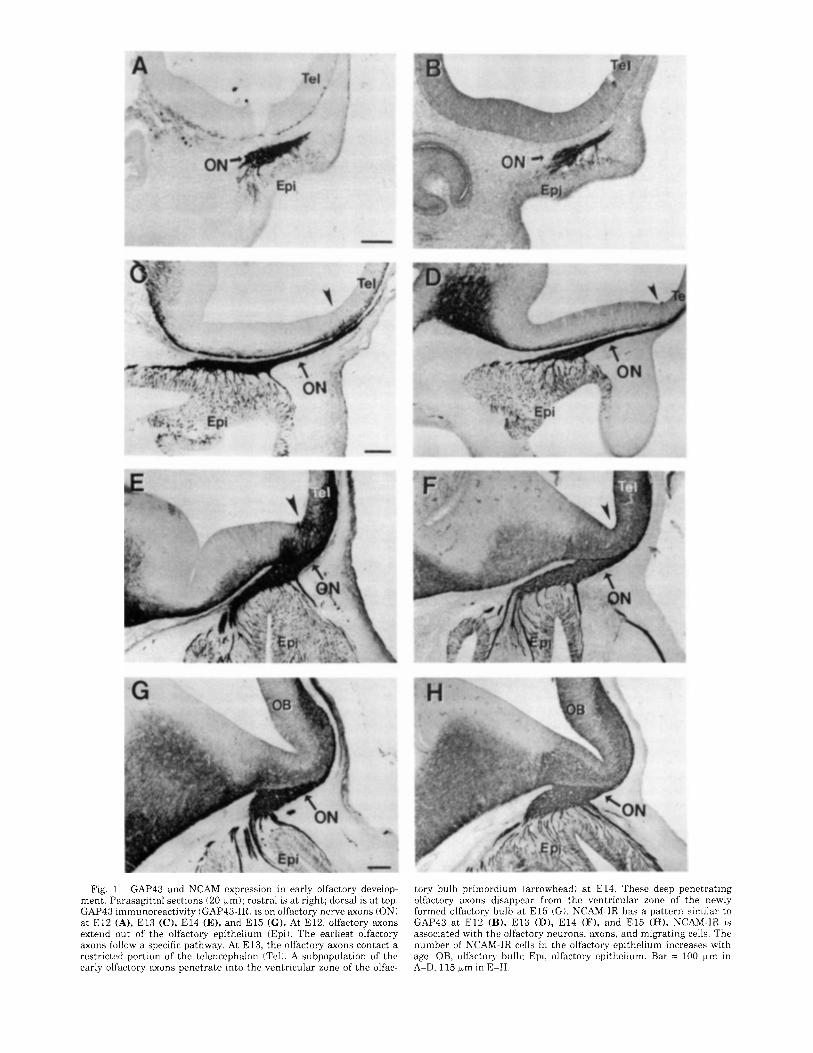
Fig. 1. GAP43 and NCAM expression in early olfactory develop- ment. Parasagt ta l sections (20 km): rostra1 is a t right; dorsal is at top. GAP43 immunoreactivity tGAP43-IRl is on olfactory nerve axons (ON) a t E l 2 (A), E l 3 (C), E l 4 (E), and E l 5 ( G ) . At E12, olfactory axons extend out of the olfactory epithelium (Epil. The earliest olfactory axons follow a specific pathway. At E13, the olfactory axons contact a restricted portion of the telencephalon !Tell. A subpopulation of the early olfactory axons penetrate into the ventricular zone of the olfac-
tory bulb primordium (arrowhead) a t E14. These deep penetrating olfactory axons disappear from the ventricular zone of the newly formed olfactory bulb at E l 5 ! G ) . NCAM-IR has a pattern similar to GAP43 a t E l 2 (B), E l 3 (D), E l 4 (F), and E l 5 (H). NCAM-IR is associated with the olfactory neurons, axons, and migrating cells. The number of NCAM-IR cells in the olfactory epithelium increases with age. OB, olfactory bulb: Epi, olfactory epithelium. Bar = 100 pm in A-D, 115 km in E-H.

4 Q. GONG AND M.T. SHIPLEY
the olfactory bulb primordium is not histologically different from the telencephalon, and the PNA staining pattern does not indicate any difference between the olfactory bulb primordium and the adjacent neocortex.
El4 GAP43. Pioneer olfactory axons penetrate into the ven-
tricular zone of the telencephalon at E14. The olfactory primordium starts to evaginate (Fig. 1E).
The olfactory nerve still has high levels of NCAM immunoreactivity. NCAM-positive olfactory axons penetrate into the ventricular zone of the telencephalon; some NCAM-positive fibers reach the ventricular surface. The number of NCAM-positive fibers in the telencephalon is similar to that of GAF'43. More NCAM-positive cells are observed in the olfactory epithelium than at E l 3 (Fig. 1F).
L1-positive olfactory axons are present within the olfactory nerve, in the mantle layer and ventricular zone of the telencephalon. The number of L1-positive olfactory axons is less than GAP43 and NCAM-positive axons (Fig. 2E).
Punctate laminin staining is still present along the olfactory pathway (Fig. 5C). In the telencephalon, small puncta are present, especially in the mantle layer of the neuroepithelium (Fig. 5D). There is a distinct gap in the laminin staining of the meninges where the olfactory axons enter into the telencephalon (Figs. 3E, 5C). In addition, the punctate laminin staining continues where the olfactory axons enter the telencephalon; similar, but smaller, puncta are also present in the mantle layer of the olfactory primordium (Fig. 5C).
Fibronectin is completely absent from the olfactory nerve pathway although it is still present in the mesenchyme surrounding the olfactory nerve (Figs. 3F, 5G).
At E14, PNA staining (Fig. 2F) increases in the olfactory nerve especially in the proximal parts of the nerve. Staining is slightly decreased in the mantle layer in the olfactory primordium compared to the adjacent cortex region. In contrast to the axonal staining with GAF'43, NCAM, and L1, none of the deep penetrating fibers in the ventricular zone are stained with PNA.
NCAM.
L1.
Laminin and fibronectin.
PNA.
El5 GAP43. At E15, pioneer olfactory axons disappear from
the ventricular zone of the telencephalon, and the number of olfactory axons increases significantly in the mantle layer of the olfactory bulb (Fig. 1G).
The NCAM staining pattern is similar to that of GAF'43. NCAM-positive axons disappear from the ventricu- lar zone of the olfactory bulb. Both NCAM-positive fibers and some positive cells are found in the mantle layer of the olfactory bulb, indicating that the staining might be on newly differentiated olfactory bulb neurons and their pro- cesses (Fig. 1H).
The olfactory nerve still has high levels of L1 immunoreactivity. The staining pattern of L l is similar to that of GAP43 at this stage. L1 axons are not seen in the ventricular zone of the olfactory bulb although a high level of L1 immunoreactivity is found in the mantle layer of the olfactory bulb (Fig. 2G). Unlike NCAM expression pattern, there are no L1-positive cells in the mantle layer of the olfactory bulb.
At E15, punctate staining begins to diminish in the part of the olfactory nerve nearest the olfactory epithelium (Fig. 3G). Punctate laminin stain- ing remains in the large olfactory nerve bundle closer to the olfactory bulb (Fig. 5E). Multiple gaps in the laminin-
NCAM.
L1.
Laminin and fibronectin.
NCAM. NCAM is expressed on the olfactory axons at this early stage. The staining pattern of NCAM is similar to that of GAP43. In addition, several cells in the olfactory placode are NCAM positive, and numerous NCAM positive cells were observed among the olfactory axons migrating along the trajectory of the olfactory nerve as reported previously (Schwanzel-Fukuda et al., 1992) (Fig. 1B).
The earliest olfactory axons also have high levels of L1 immunoreactivity. The staining pattern of L1 is similar to NCAM and GAP43 (Fig. 2A). L1-positive cell bodies were found in the olfactory placode at E12. L1 is also expressed on cells along the olfactory pathway.
Laminin appears as a diffuse background stain throughout the cranial mesenchyme. The meninges surrounding the telencephalon are densely stained (Figs. 3A, 4A). In the olfactory epithelium (OE), there is no specific laminin staining. However, laminin has a distinc- tive staining pattern along the trajectory of the olfactory axons (Figs. 3A, 4A). Here, laminin immunoreactivity has a unique punctate appearance (Fig. 4A). The staining inten- sity of the laminin puncta was higher than the diffuse reactivity of the cranial mesenchyme surrounding the olfactory pathway.
Diffuse fibronectin staining is present throughout the cranial portion of the mesenchyme; there is no specific pattern in the olfactory nerve pathway (Fig. 3B).
Along the olfactory pathway, PNA staining is very light and appears to be associated with olfactory axons. The cranial mesenchyme has no detectable PNA binding (Fig. 2B).
El.? GAP43. At this stage, most of the olfactory axons
coalesce to form a single olfactory nerve fascicle, which grows along the ventral surface of the telencephalon out- side of the meninges. GAP43-positive olfactory axons con- tact the telencephalon at a distinct point on its surface where a few fibers enter the telencephalon (Fig. 1C).
NCAM is still present on the olfactory nerve at this age. NCAM immunoreactivity has the same pattern as GAP43 staining. NCAM-positive olfactory axons form a single nerve bundle that contacts the rostra1 portion of the telencephalon. NCAM-positive cells are associated with the nerve bundle (Schwanzel-Fukuda et al., 1992). There are more NCAM-positive cells present in the olfactory epithe- lium at E l 3 than at E12; these cells are located throughout the thickness of the olfactory epithelium (Fig. 1D).
L1 stains the olfactory nerve along its entire length from the OE to the telencephalon. The staining pattern of L1 is similar to that of GAP43 and NCAM. Ll-positive cells are located throughout the thickness of the olfactory epithe- lium in a pattern similar to NCAM immunoreactivity (Fig. 2C).
Punctate laminin staining is still present along the entire trajectory of the olfactory pathway (Figs. 3C, 4B). The OE is devoid of laminin immunoreactivity. Blood vessels and the meninges are densely stained. Laminin-positive meninges are intact (Figs. 3C, 4B, 5A,B).
Fibronectin is also present throughout the cranial mesen- chyme in a diffuse form, but there is slightly less fibronectin immunoreactivity in the olfactory nerve pathway (Fig. 3D). The olfactory epithelium lacks fibronectin staining.
PNA staining (Fig. 2D) is still very light along the olfactory nerve. The mantle layer of the telencephalon has more PNA binding than the ventricular zone. At this stage,
L1.
Laminin and fibronectin.
PNA.
NCAM.
L1.
Laminin and fibronectin.
PNA.
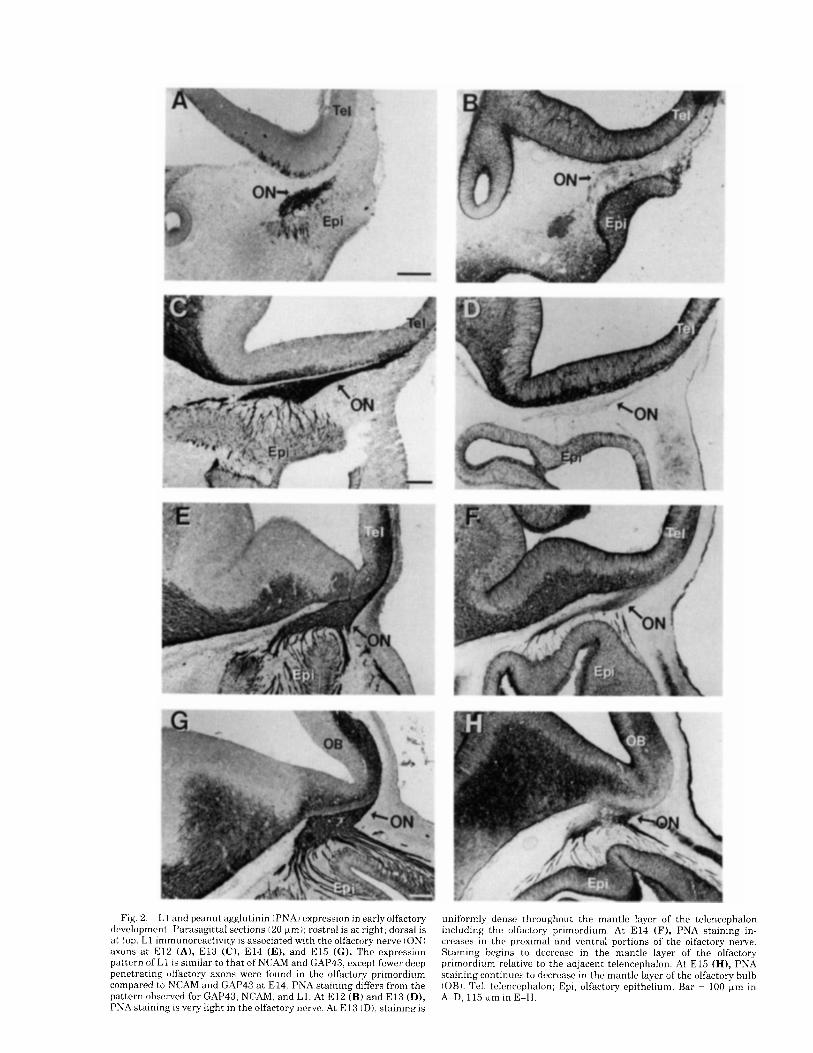
Fig. 2 . L 1 and peanut agglutinin (PNAI expression in early olfactory development. Parasagittal sections (20 Fm); rostra1 is at right; dorsal is a t top. L1 immunoreactivity is associated with the olfactory nerve (ON) axons a t E l 2 (A), E l 3 (C), E l 4 (E), and E l 5 ( G ) . The expression pattern of L1 is similar to that of NCAM and GAP43, except fewer deep penetrating olfactory axons were found in the olfactory primordium compared to NCAM and GAP43 a t E14. PNA staining differs from the pattern observed for GAP43, NCAM, and L1. At E l 2 (B) and E l 3 (D), PNA staining is very light in the olfactory nerve. At E l 3 ( D ) , staininn is
uniformly dense throughout the mantle layer of the telencephalon including the olfactory primordium. At E l 4 (F), PNA staining in- creases in the proximal and ventral portions of the olfactory nerve. Staining begins to decrease in the mantle layer of the olfactory primordium relative to the adjacent telencephalon. At E l 5 (HI, PNA staining continues to decrease in the mantle layer of the olfactory bulb (OBI. ?'el, tclcncephalon; Epi, olfactory epithelium. Bar = 100 pn in A-D, 115 wm in E-H.
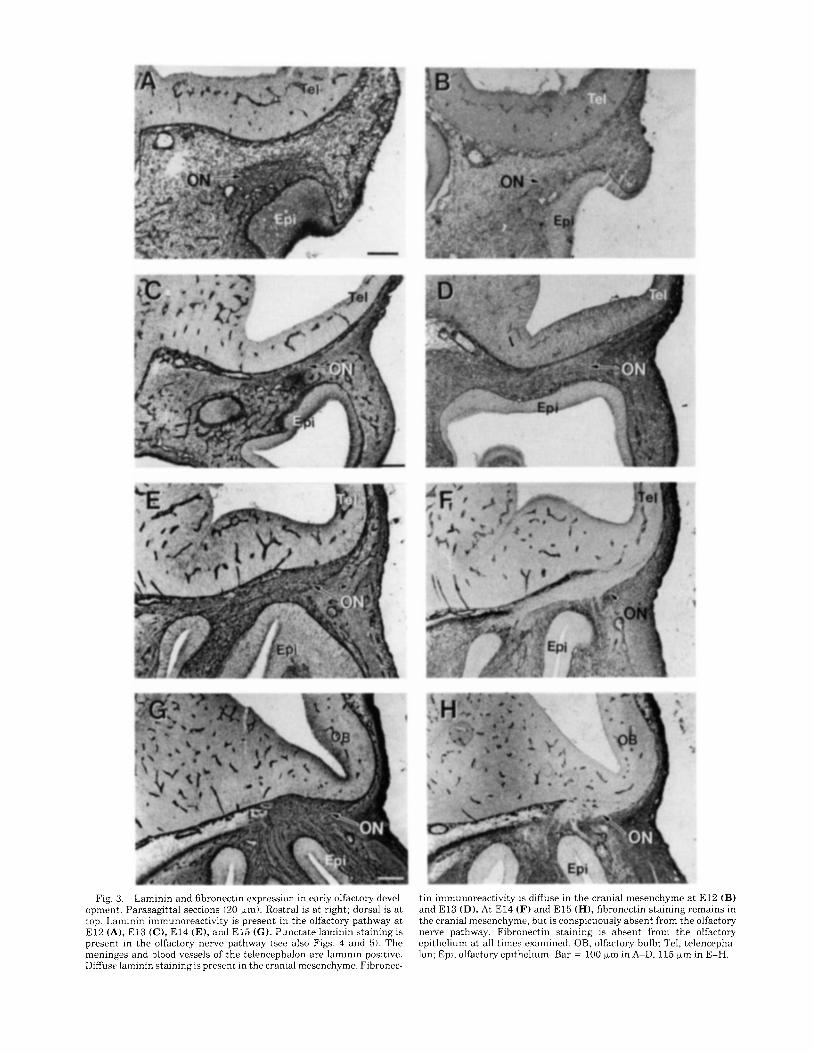
Fig. 3. Laminin and fihronectin expression in early olfactory devel- opment. Parasagittal sections (20 pm): Rostra1 is a t right; dorsal is a t top. Laminin immunoreactivity is present in the olfactory pathway a t E l 2 (A), El3 (C), E l 4 (E), and E l 5 ( C ) . Punctate laminin staining is present in the olfactory nerve pathway (see also Figs. 4 and 5). The meninges and blood vessels of the telencephalon are laminin positive. Diffuse laminin staining is present in the cranial mesenchyme. Fibronec-
tin immunoreactivity is diffuse in the cranial mesenchyme a t E l 2 (B) and E l 3 (D). At E l4 (F) and E l 5 (H), fibronectin staining remains in the cranial mesenchyme, hut is conspicuously absent from the olfactory nerve pathway. Fibronectin staining is absent from the olfactory epithelium a t all times examined. OB, olfactory bulb; Tel, telencepha- Ion; Epi, olfactory epithelium. Bar = 100 pm in A-D, 115 pm in E-H.

ECMS 4ND CSMS IN EARLY OLFACTORY DEVELOPMENT i
stained meninges are visible where olfactory nerves enter the olfactory bulb compared to the intact laminin meninges surrounding the rest of the telencephalon (Figs. 3G, 5E). Within the olfactory bulb and the rest of the telencephalon, small laminin puncta are present in the mantle layer (Fig. 5Fj. Blood vessels are laminin positive.
Fibronectin staining is uniform within the cranial mesen- chyme but is absent from the olfactory nerve pathway (Fig. 3H).
At E15, PNA staining (Fig. 2H) is high within a subpopulation of the olfactory nerve axons. PNA staining is highest in olfactory axons near the olfactory epithelium and on the ventrally located axons in the olfactory nerve. PNA staining is lower where the nerve contacts the olfactory bulb. PNA staining level is low in the mantle layer of the olfactory bulb compared to the adjacent neocortex.
PNA.
Tenascin and chondroitin sulfate A number of studies have implicated tenascin and chon-
droitin sulfate in barrier or growth inhibitory functions during neural development. As we have previously reported (see also Fig lA), early olfactory axons initially grow directly toward the telencephalon, but at a distinct point, they turn 90" to grow rostrally. It is reasonable to suspect that the axons encounter either a barrier or growth- inhibitory molecules causing them to turn. With this in mind, we investigated the expression patterns of tenascin and chondroitin sulfate during early olfactory axon out- growth. No specific staining patterns were associated with the olfactory nerve pathway for either of these epitopes at the ages examined (E12-El5).
In summary, extracellular matrix molecules and cell surface molecules are expressed with specific temporal and spatial patterns during early development of the olfactory system. L1 and NCAM are associated with developing olfactory axons. Laminin is present as a unique punctate staining pattern along the olfactory nerve trajectory. The presence of these molecules suggests that they may be involved in early olfactory nerve outgrowth and targeting.
L l is expressed on immature ORNs L1 staining in the olfactory epithelium suggests that this
epitope might be selectively expressed by immature olfac- tory neurons. Olfactory marker protein (OMP) is generally accepted as hallmark for the mature olfactory receptor neurons. Thus, we compared the distribution of OMP- and L1-positive cells from E l 2 to postnatal day 30 (P30). We refer to the part of the OE closer to the lamina propria as the deep portion and the region closer to the epithelial surface as the superficial portion. L1-positive cells are already present in OE at E l 2 (Fig. 2Aj; no OMP cells or axons are observed in the OE a t this age. At E16, L1- posit,ive cells are located throughout the OE but are most heavily concentrated in the deep half of the OE. OMP- positive cells are first seen at this age and are located at the superficial half to one-third of the olfactory epithelium (Fig. 6A,B). At E21, Ll-positive cells are restricted to the deep half of the OE; OMP cells are mostly in the superficial half of the OE. At this age, there is little overlap in the distributions of L1 and OMP immunoreactivity (Fig. 6C,D). At P30. which is similar to the adult picture, L1 cells are still restricted to the deep half of the OE. More OMP cells are found in the OE compared to earlier ages and are mostly located in the middle third of the OE. L1- and OMP-positive
cells partially overlap only in the middle portion of OE (Fig 6E,F). Olfactory receptor neurons are generated in the deepest part of the olfactory epithelium. As they mature their cell bodies migrate to more superficial parts of the OE and start to express OMP. The present findings indicate that L1 is expressed mainly on cells in the deep part of the epithelium. Therefore, we conclude that L1 is expressed primarily, if not exclusively, by immature olfactory receptor neurons.
DISCUSSION The results of this study indicate that the patterns of
expression of several extracellular matrix molecules and cell surface molecules are temporally and spatially regu- lated in the olfactory nerve pathway during early olfactory axon pathfinding and targeting. NCAM and L1 are present on the earliest olfactory axons but not on early Schwann cells. We also observed that L1 and OMP are expressed in largely different complementary populations of cells in the developing olfactory epithelium. L1 appears to express on immature olfactory receptor neurons. We show for the first time that laminin is expressed with a unique punctate staining pattern in the developing rat olfactory nerve. The spatio-temporal expression patterns of these extracellular matrix molecules and cell surface molecules on the olfac- tory axons suggest that they participate in olfactory axon outgrowth and fasciculation. By contrast, the lack of any temporal-spatial pattern of expression of tenascin and condroitin sulfate proteoglycan in the early olfactory system suggests that these barrier or growth-inhibitory molecules might not play roles in early olfactory system development.
Pioneer rat olfactory axons grow out of the olfactory placode at E l 2 and follow a specific stereotypic pathway to reach a restricted region of the rostra1 telencephalon, the olfactory bulb primordium, during early olfactory system development (Gong and Shipley, 1995). The initial growth of the olfactory axons is probably guided by specific cellular and molecular cues, as we observe the same axonal trajec- tory in all animals.
The development of axonal pathways involves several coordinated events: neurite extension, axonal guidance, fasciculation, target contact, and synapse formation. The interaction of growth cones with molecules present in the local environment is crucial in these events. Both positive and negative factors are important for axonal guidance (Kapfhammer and Raper, 1987a,b; Patterson, 1988). Posi- tive factors promote neurite outgrowth; negative influences from surrounding tissues can constrain axons to certain routes (Silver et al., 1987; Tosney and Oakley, 1990). Integrin and neuronal cell adhesion molecules NCAM, N-cadherin, and LI are found on growth cones (Bixby, 1991; Bixby et al., 1988; Doherty and Walsh, 1992; Rei- chardt and Tomaselli, 1991). Integrins are receptors for extracellular matrix molecules such as laminin and fibronec tin. Both NCAM and L1 function through homophilic binding, and thus they may be important in axon-axon interaction (Doherty et al., 1990b; Grumet et al., 1982; Rutishauser and Landmesser, 1991 ) , To investigate poten- tial mechanism(s) of olfactory axon pathfinding, we ana- lyzed the presence of extracellular matrix molecules (lami- nin and fibronectinj and adhesion molecules (NCAM and L1) in the olfactory axonal pathway during the earliest stages of rat olfactory system development.
8 Q. GONG AND M.T. SHIPLEY
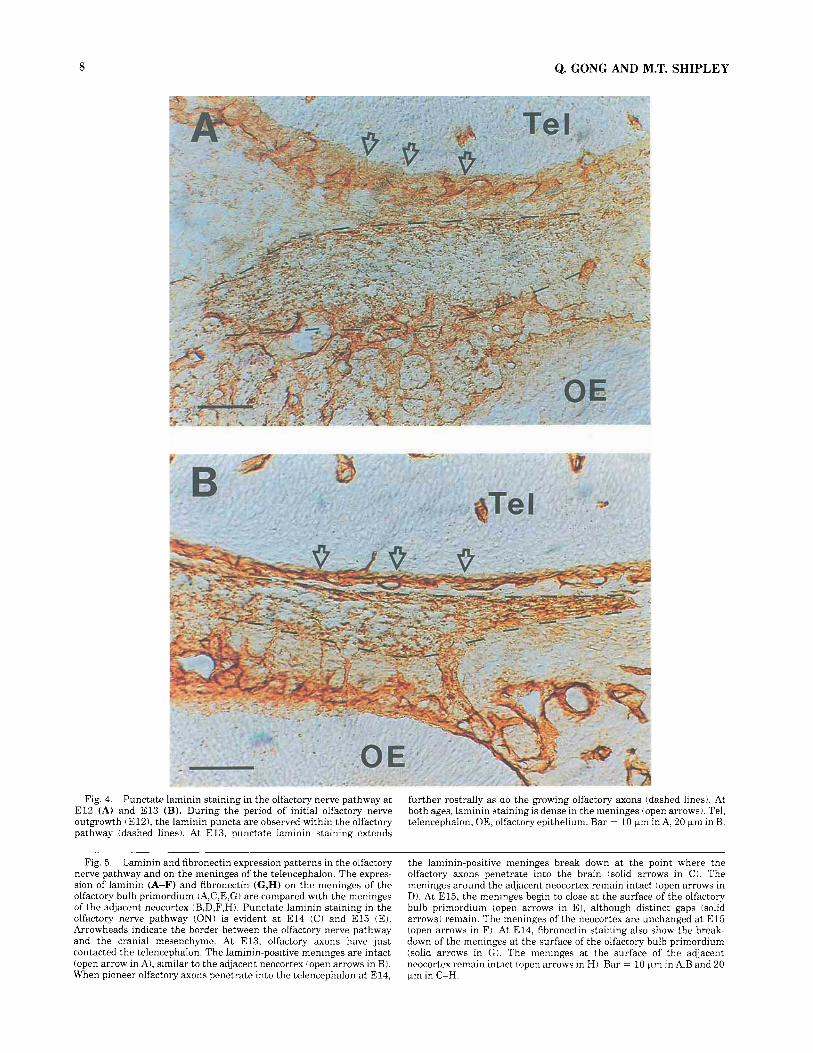
Fig. 4. Punctate laminin staining in the olfactory nerve pathway at E l 2 (A) and E l 3 (B). During the period of initial olfactory nerve outgrowth tE121, the laminin puncta are observed within the olfactory pathway (dashed lines). At E13, punctate laminin staining extends
further rostrally as do the growing olfactory axons (dashed lines). At both ages, laminin staining is dense in the meninges (open arrows). Tel, telencephalon; OE, olfactory epithelium. Bar = 10 pm in A, 20 pm in B.
Fig. 5. Laminin and fibronectin expression patterns in the olfactory nerve pathway and on the meninges of the telencephalon. The expres- sion of laminin (A-F) and fibronectin (G,H) on the meninges of the olfactory bulb primordium (A,C,E,G) are compared with the meninges of the adjacent neocortex (B,D,F,H). Punctate laminin staining in the olfactory nerve pathway (ON) is evident at E l 4 (Cl and E l 5 (El. Arrowheads indicate the border between the olfactory nerve pathway and the cranial mesenchyme. At E13, olfactory axons have just contacted the telencephalon. The laminin-positive meninges are intact (open arrow in A), similar to the adjacent neocortex (open arrows in B). When pioneer olfactory s o n s penetrate into the telencephalon a t E14,
the laminin-positive meninges break down at the point where the olfactory axons penetrate into the brain (solid arrows in C). The meninges around the adjacent neocortex remain intact (open arrows in D). At E15, the meninges begin to close a t the surface of the olfactory bulb primordium (open arrows in El, although distinct gaps (solid arrows) remain. The meninges of the neocortex are unchanged a t E l 5 (open arrows in Fi. At E14, fibronectin staining also show the break- down of the meninges a t the surface of the olfactory bulb primordium (solid arrows in G). The meninges at the surface of the adjacent neocortex remain intact (open arrows in Hi. Bar = 10 pm in A,B and 20 pm in C-H.
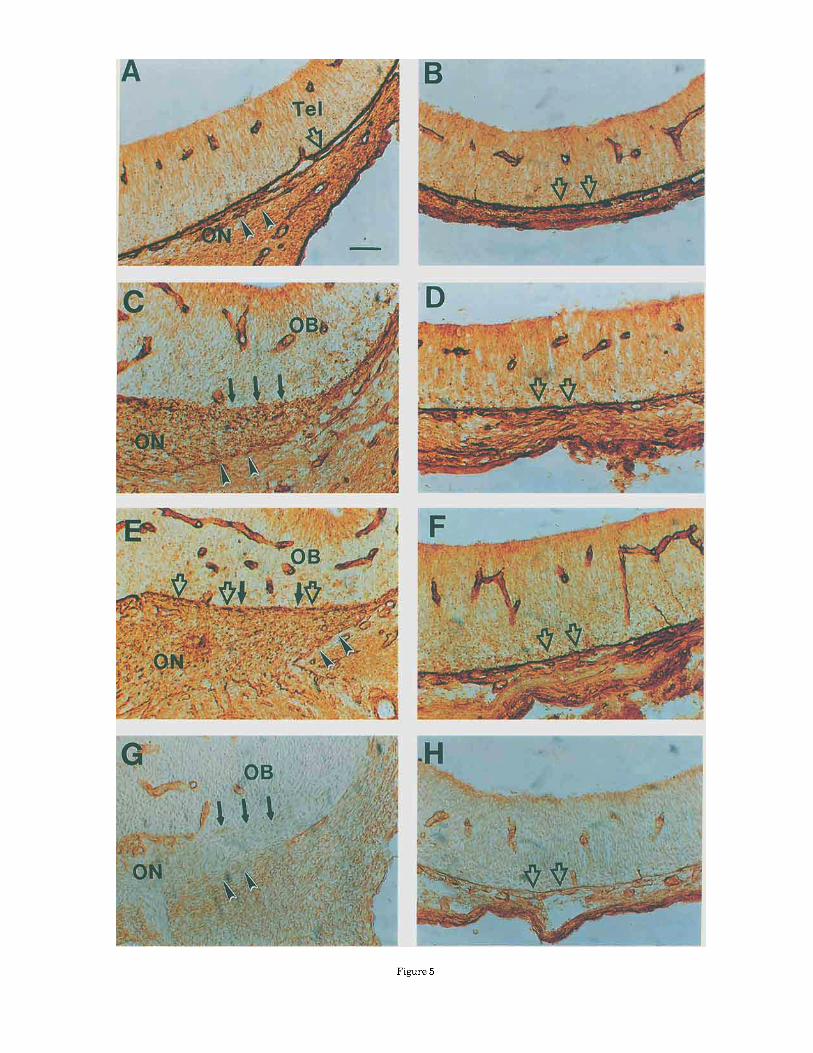
Figure 5

&. GONG AND M.T. SHIPLEY
Fig. 6. L1 and olfactory protein marke (OMPI are expressed in different cell populations in the olfactory epithelium (OE). The staining patterns of L1 (A,C,E) and OMP (B,D,F) are compared on alternate sections at E l 6 tA,B), E21 (C,D), and P30 (E,F). At E16, L1-positive cells ( A ) are present a t all depths of the OE. OMP-positive cells (B) are located in the superficial half to one-third of the OE. At E21, L1-positive
cells (C) are restricted to the deep half of the OE, whereas OMP-positive cells iD) are present in the superficial two-thirds of the OE. At P30, L1-positive cells are present in the deep half of the OE, where- as OMP-positive cells are present in the middle third of the OE. Arrowheads indicate the OMP-positive cells. LP, laminar propria. Bar = 25 pm.
NCAM is a sialoglycoprotein that is involved in cell-cell interactions in the central and peripheral nervous systems (Walsh and Doherty, 1991). NCAM is ubiquitously ex- pressed during development. Its role in axonal guidance and neurite outgrowth is supported by the results of many studies. Evidence that NCAM promotes neurite outgrowth comes from in vivo and in vitro antibody blocking studies (Bixby et al., 1987; Chang et al., 1987; Chuah et al., 1991; Drazba and Lemmon, 1990) and through transfection of the gene for NCAM into heterologous cells (Doherty and Walsh. 1989; Doherty et al., 1990a). NCAM expression
patterns and perturbation experiments in vivo provide evidence that this molecule is involved in neuronal pattern formation (Landmesser et al., 1988, 1990; Rutishauser et al., 1983).
The expression of NCAM in the developing rat olfactory system is similar to that reported for the mouse (Chung et al., 1991; Miragall et al., 1989; Schwanzel-Fukuda et al., 1992; Terkelsen et al., 1989). NCAM is present on the surface of the olfactory receptor neurons and their axons. The cranial mesenchyme around the olfactory nerve trajec- tory is negative for NCAM immunoreactivity. Anew finding

ECMS AND CSMS IN EARLY OLFACTORY DEVELOPMENT 11
in this study is that most of the deep penetrating (pioneer) olfactory axons in the ventricular zone of the telencephalon also possess NCAM on their surface although the level of expression appears to be lower compared to the more proximal portion of the axons. This expression pattern suggests that NCAM might be very important in axon-axon interaction within the olfactory nerve but is less prominent in promoting neurite extension into the ventricular zone of the telencephalon. The fact that there is some NCAM expression in the olfactory bulb primordium suggests that NCAM on telencephalic cells could interact with olfactory axons. In vitro studies suggest the importance of NCAM to neurite extension in the olfactory system. Antibodies block- ing NCAM inhibit neurite outgrowth of olfactory receptor neurons (Chuah et al., 1991).
Like previous investigators of the mouse olfactory sys- tem, we have also observed NCAM-positive cells migrating out of the rat olfactory placode along the olfactory nerve pathway (Schwanzel-Fukuda et al., 1992). Schwanzel- Fukuda et a1 (1992) reported that NCAM is selectively expressed on cells in the olfactory nerve pathway associated with migrating LHRH-containing neurons and suggested that NCAM might function as a “scaffold” for these migrating cells. However, these investigators did not show NCAM immunoreactivity in the olfactory epithelium or olfactory axons. With our immunocytochemical techniques, we observed that the NCAM is strongly expressed by both olfactory axons and neurons in the olfactory epithelium as well as migrating cells along the olfactory nerve. It is not surprising then that NCAM is present on all neuronal elements in the developing pathway, including migrating cells. Thus, although NCAM may have some functional role in the migration of LHRH-containing cells, its ubiquitous expression on all neuronal elements suggests that this molecule alone cannot explain the directed migration of LHRH neurons from the epithelium to the brain.
NCAM has been reported to be expressed on olfactory Schwann cell surfaces later in development and in vitro (Chuah and Au, 1993; Miragall et al., 1989). Chuah et al. suggested that NCAM on olfactory Schwann cells might play a role in axon outgrowth. In support, they reported that antibodies against NCAM inhibited olfactory neurite extension in vitro (Chuah et al., 1991). In the present study, however, we did not detect NCAM on the olfactory Schwann cells during early axon outgrowth. The low-affinity nerve growth factor receptor (LNGFr) and S-100 are present on olfactory nerve Schwann cells at this time; however the cells are located not within, but at the periphery of, the olfactory nerve pathway. Schwann cells are not present among the olfactory axons until later in development (E14) after olfactory axons have already contacted the telencepha- lon (Gong et al., 1994). Thus, although NCAM on olfactory nerve Schwann cells might be important for olfactory axon outgrowth in later development or in adults, it does not appear to be essential to the outgrowth of the earliest olfactory axons.
Axon fasciculation is an important mechanism in guiding axons to their targets. L1 promotes neurite outgrowth and fasciculation in vitro (Chang et al., 1987; Drazba and Lemmon, 1990; Lagenauer and Lemmon, 1987). The devel- opmental expression pattern of L1 and perturbation experi- ments using L1 antibodies have suggested that this mol- ecule plays important role(s) in neurite outgrowth and fasciculation. The role of L1 in PNS-Schwann cell interac- tions has also been investigated (Bixby et al., 1988; Kleit-
man et al., 1988; Seilheimer and Schachner, 1988). Neurite outgrowth from dorsal root (Seilheimer and Schachner, 1988) and ciliary ganglion neurons (Bixby et al., 1988) on Schwann cells is sensitive to anti-Ll antibodies, as are neurites from retinal ganglion cells on Schwann cells (Kleitman et al., 1988). L1 is expressed in the developing mouse olfactory system (Miragall et al., 19891, but the expression of L1 in the initial stages of olfactory nerve development has not been characterized. As olfactory nerve Schwann cells do not appear to be present in the olfactory nerve pathway during the early stages (E12-13) of the olfactory axons outgrowth, L1 on olfactory axons probably does not interact with the Schwann cells. However, L1 immunoreactivity is strong in the olfactory nerve, suggest- ing that L1 may be important in axonal extension and/or fasciculation.
Preliminary observations suggested that L1 is expressed in immature, but not mature olfactory receptor neurons. If these observations are correct, then the L1 staining should be 1) expressed on ORNs, before OMP expression, 2) L1 expressed in immature ORNs which are in the deep layer of the OE, and 3 ) present on a subpopulation of ORNs in the adult OE, since there is constant ORN turnover. In this study, we observed L1-positive cells in the olfactory placode a t E12. The first OMP-positive cells were seen a t E16. Therefore, L1 appears in the OE before OMP expression. We also observed that L1- and OMP-positive olfactory receptor neurons are separate populations. L1 cells are located on the lower portion of the olfactory epithelium, whereas OMP-positive cells are located in the superficial portion of the epithelium just below the nuclei of the supporting cells. This observation is consistent with the fact that olfactory receptor neurons are generated from basal cells in the bottom layer of the olfactory epithelium, from where they migrate superficially to become mature olfactory receptor neurons. L1-positive cells were also observed in the deep parts of the adult olfactory epithelium. Therefore, L1 appears to be expressed in differentiating ORNs.
Laminin is a glycoprotein present in basement mem- branes where it provides an adhesive substrate for many cell types (Timpl et al., 1983; Timpl and Rohde, 1979). It is present in the CNS and PNS (Letourneau et al., 1988; Rogers et al., 1986; Yip and Yip, 1992; Zhou, 1990). Laminin promotes neurite outgrowth of peripheral and central axons in vitro (Evercooren et al., 1982; Liesi et al., 1984; Manthorpe et al., 1983; Rogers et al., 1983). I t has also been suggested to guide fiber outgrowth in vivo (Ham- marback et al., 1985). Punctate laminin staining, such as seen here in the olfactory nerve, has been described in the CNS and in the developing optic pathway. One report located punctiform laminin along the trajectory of future nerve tracts tZhou, 1990), e.g., in medial forebrain bundle, fascicularis retroflexus, and cortical subplate prior to, or at the time of, arrival of growing axons. Letourneau et al. ( 1988) observed that laminin puncta accumulate in the ventral longitudinal pathway of the spinal cord during development. Liesi and Silver (1988) reported that punc- tate laminin is present on immature glial cells in the developing optic nerve. In the present study, punctate laminin was observed within the developing olfactory path- way. This pattern of laminin expression was already pre- sent at E l 2 when the first olfactory axons grow out of the olfactory placode. The laminin puncta are highly restricted to the olfactory nerve pathway. This finding, taken with

12 Q. GONG AND M.T. SHIPLEY
were recently shown to define borders between rhombo- meres in the chick hindbrain (Layer and Alber, 1990). Davies et al. (1990) extracted PNA binding glycoprotein and showed that it inhibits growth cone motility. PNA also binds to future chondrogenic tissues (Layer et al., 1988; Liu and Layer, 1984; Oakley and Tosney, 1991). PNA binding sites are present in the developing olfactory nerve, but the staining patterns are not immediately consistent with a barrier function. The levels of expression appeared to correlate with the degree of fasciculation of the olfactory nerve. PNA staining is low in the olfactory nerve at E l 2 and E l 3 when the olfactory nerve is not tightly bundled but increases a t E l 4 and E l 5 when the axons become more tightly fasciculated. PNA staining is low where the olfac- tory nerve contacts the surface of the telencephalon. As the molecules recognized by PNA are still poorly characterized, it is premature to conjecture what these staining patterns might mean in functional terms other than to speculate that this lectin might bind to molecules associated with fasciculated axons in the developing olfactory nerve. I t would be interesting to determine if PNA binding is altered during the degeneration or regeneration of the olfactory nerve following lesions.
In summary, NCAM and L1 are present along the olfactory nerve during early olfactory axon outgrowth and pathfinding, indicating that these molecules may be in- volved in promoting olfactory axon outgrowth and fascicula- tion. Laminin has a unique punctate staining pattern within the olfactory nerve pathway and may function in olfactory axon guidance and growth. When the earliest olfactory axons contact the telencephalon, the meninges break down to allow the axons to enter the brain. The mechanisms that underlie this breakdown deserve further investigation as this phenomenon may play a role in Kallmann's syndrome.
previous reports, supports the suggestion of Liesi and Silver (1988) that the punctate form of laminin plays a role in guiding and promoting axons to their target.
The meninges surrounding the developing telencephalon are laminin and fibronectin positive. At E14, the laminin- fibronectin-positive meninges break down precisely at the point where the olfactory nerve axons enter the telencepha- lon. This breakdown has the appearance of gaps in an otherwise solid wall. If the part of the telencephalon that differentiates into olfactory primordium is intrinsically different from other parts of the telencephalon, then the meninges in this location may break down to allow the olfactory axons to enter the brain. On the other hand, it is possible that the olfactory primordium is not different but, rather, the pioneer olfactory axons have the ability to break the meningeal barrier in order to enter the telencephalon. Doucette (1991) reported that the meninges are intact on the surface of the adult olfactory bulb. Our observations indicate that the meninges completely break down at the point where the earliest olfactory axons enter the telen- cephalon. Twenty-four to 48 hours later in development, the laminin-positive meninges begin to reform, but gaps always remain where bundles of axons enter the olfactory bulb. Together, these observations suggest that interac- tions between early olfactory axons and the surface of the olfactory bulb primordium are important in olfactory bulb development. If, for example, the meninges remain intact, or open and then close again, the olfactory axons and/or migrating LHRH cells might be excluded from the brain. Kallmann's syndrome is characterized by the failure of olfactory axons and LHRH neurons to enter the CNS. The possibility that the meningeal barrier could be abnormal in sufferers of this disease has not been previously suggested but warrants consideration.
Fibronectin is also an axon growth-promoting factor, but it may not provide guidance for growing axons (Rogers et al., 1989). During initial olfactory axon outgrowth (E12- E13), fibronectin is ubiquitously distributed throughout the cranial mesenchyme, but there is no specific pattern of this molecule associated with the olfactory nerve.
The earliest olfactory axons initially grow directly toward the telencephalon. When they reach a point approximately 50 pm from the telencephalic surface, they turn 90" and extend rostrally. One possible explanation for this sharp change in the direction of axon growth is that inhibitory molecules may be present at the point where olfactory axons turn. We investigated three molecules which have been shown to have barrier functions, tenascin, chondroitin sulfate proteoglycan and PNA-binding sites (Brittis et al., 1992; Gonzalez and Silver, 1992; Snow et al., 1990). However, we were unable to demonstrate any specific staining pattern at the point where the axons turn. How- ever, these are negative findings, and it is possible that other molecules, not investigated, play a role in deflecting the trajectory of growing olfactory axons toward the rostral telencephalon. I t is also possible that the axons turn toward a diffusable attractive factor, or that they are reacting to mechanical forces caused by the rostral growth of the snout.
Peanut agglutinin (PNA) binds specifically to D-Gal-1,3- b-NAcGal disaccharide residues of glycoconjugates (Lotan et al., 1975). PNA binding sites have been associated with barrier functions (Tosney and Oakley, 1990). The chick spinal nerve pathway through the scleratome is delineated by the absence of staining with PNA. PNA binding sites
ACKNOWLEDGMENTS This work was supported by NIH DC 00347 and NS
29218.
LITERATURE CITED Appel, F., J. Holm, J. Conscience. and M. Schachner (19931 Several
extracellular dumains of the neural cell adhesion molecule L1 are involved in neurite outgrowth and cell body adhesion. J. Neurosci. 13.4764-4775.
Bixby, J .L . (1991, Molecular mechanisms of axon growth and guidance. Annu. Rev. Cell Uiol. 7:117-159.
Bixby. J.L., R.L. Pratt, J. Lilien, and L.F. Reichardt (1987, Neurite outkvowth on muscle cell surfaces involves extracellular matrix recep- tors as well as Ca' '-dependent and independent cell adhesion molecules. Proc. Natl. Acad. Sci. [J.S.A. 84.2555-2569.
Bixby, J .L., J. Lilien. and L.F. Reichardt (1988) Identification o f the major proteins that promote neuronal process outgrowth on Schwann cells in vitro. J. Cell Biol. 107:353-361.
Brittis, P.A., D.R. Canning. and J. Silver (1992) Chondroitin sulfate as a rekwlatnr of ncuronal patterning in the retina. Science 255733-736.
Chang, S.. F.G. Kath.jen, and J.A. Raper (1987, Extension of neurites on axons is impaired by antibodies against specific neural cell surface glycoproteins. J. Cell Biol. 104:355-362.
Chuah, M.I., and I . Au (1993) Cultures of ensheathing cells from neonatal rat olfactory bulbs. Brain Kes. 601:213-220.
Chuah, M.I. , S. David, and 0. Blaschuk (1991, Differentiation and survival of rat olfactory epithelial neurons in dissociated cell culture. Dev. Brain Kes. 60:123-132.
Chung. W., C.F. Lagenaur. Y. Yan, and J.S. Lund 119911 Developmental expression uf neural cell adhesion molecules in the mouse neocortex and olfactory bulb. ,I. Comp. Neurol. 314.290-305.

ECMS AND CSMS IN EARLY OLFACTORY DEVELOPMENT 13
Cunningham, B.A., J . J . Hemperly, B.A. Murray, E.A. Prediger. R. Uracken- bury, and R. Edelman 11987) Neural cell adhesion molecule: structure, immunoglobulin-like domains, cell surface modulation, and alternative RNA splicing. Science 236:799-806.
Davies. 5 .4 . . G.M. Conk. C.D. Stern, and R.J. Keynes (1990) Isolation from chick somites of a glycoprotein fraction causes collapse of dorsal root ganglion growth cones. Neuron 2:11-20.
Doherty. P., and F.S. Wakh I19891 Neurite guidance molecules. Curr. Opin. Cell Biol. 1:1102.
Doherty. P., and F.S. Walsh I19921 Cell adhesion molecules, second messen- gers and axonal growth. Curr, Opin. Neurobiol. 2.595-601.
D0hert.v. P.. J. Cohen, and F.S. Walsh l1990aJ Neurite outgrowth in response ti1 transfected N-CAM changes during development and is modulated by polysialic acid. Neuron 5209-219.
Doherty, l’., M. Fruns, P. Seaton, G. Dickson, C.H. Barton, T.A. Sears, and F.S. Walsh (1990bl A threshold effect of the major isoforms of NCAM on neurite outgrowth. Nature 343:464-466.
Doucette. K. i 1991) PNS-CNS transitional z m e of the first cranial nerve. J . Comp. Neurol. 312:415-466.
Drazha. J . . and V. Lemmon 11990) The role of cell adhesion molecules in neuritr, outgrowth on Muller cells. Dev. Bid . I,?X:82-93.
Evercooren, A.B. , H.K. Kleinman, S. Ohno, P. Marangos, J.P. Schwartz. and M.E. Dubois-Dalcq ( 1982) Nerve growth factor. laminin, and fibronectin promote neurite growth in human fetal sensory ganglia cultures. J. Neurosci. Res. 8:179-193.
Gong, Q.. and M.T. Shipley 119951 Evidence that pioneer olfactory axons regulate telencephalnn cell cycle kinetics to induce the formation of the olfactory bulb. Neuron 14:91-101.
Gong. Q.. M.S. Bailey. S.K. Pixley, M. Ennis, W. Liu, and M.T. Shipley 119941 Localization and regeneration of low affinity nerve growth factor recep- tor expression in the ra t olfactory system during development and regeneration. J. Comp. Neurol. 344.336-348.
Gonzalez, M.D.L., and J . Silver 11992) Role of inhihitory ECM molecules in the formation of olfactory bulb glomeruli. Soc. Neurosci. Abstr. I8:620.
Grumet. M., U. Rutishauser, and G.M. Edelman 119821 N-CAM mediates adhesion between embryonic nerve and muscle cell in vitro. Nature 295:699-695.
Hammarback, I.A., S.L. Palm, L.T. Furcht, and P.C. Letourneau ll985l Guidance of neurite o u t g o w t h by pathways of substratum-adsorbed laminin. J . Neurosci. Res. 13213-220.
Jacobson. M., and U. Rutishauser 119861 Induction of neural cell adhesion molecule (NCAMl in Xenopus embryos. Dev. Biol. 116:524-531.
Jousimaa, J.. J. Merinmies, and H. Rauvala 11984) Neurite outgrowth of neuroblastoma cells induced by proteins covalently coupled to glass coverslips. Eur. J . Cell Biol. 35:51-6l.
Kapfhammer, J.P., and J.A. Raper t1987al Collapse ofgrowth cone structure on contact with specific neurites in culture. J . Neurosci. 7301-212.
Kapfhammer, J .P. , and J.A. Raper 11987b! Interactions between F o w t h cones and neurites growing from different neural tissues in culture. J. Neurosci. 7.1595-1600.
Kleitm;in. N., D. Simon, M. Schachner, and R. Runge 11988) Growth of emhryonic retinal neurites elicited by contact with Schwann cell surfaces is blocked by antibodies to L I . Exp. Neurol. 102.298-306.
Laaenauer. C.. and V. Lemmon 119871 A LI-like molecule. the 8D9 anticen. c’is a po ten t substrate for neurite extension. Proc. Natl.’ Acad. Sci. U.S.A. 84.7753-7757.
Lander, A.D.. D.K. Fujii, and L.F. Reichardt 119851 Laminin is associated with the neurite outgrowth-promoting factors found in conditioned media. Proc. Natl. Acad. Sci. U.S.A. 82.2183-2187.
Landmesser, L., L. Dahm. K. Schultz, and U. Kutishauser 11988) Distinct roles for adhesion molecules during innervation embryonic chick muscle. Dev. Biol. 103.645-670.
Landmtwer, L., L. Dahm, J . Tang, and U. Rutishauser 11990) Polysialic acid a s ;I regulator of intramuscular nerve branching during embryonic development. Neuron 4.655-667.
Layer. P.G, and R. Alber (1990) Patterning of chick hrain vesicles a s revealed hy peanut agglutinin and cholinesterases. Development 109613- 624
Layer, P.G , R. Alber, and F.G. Rath.jen 11988) Sequential activation of but~rulcholinesterase in rostra1 half somites and acetylcholinesterase in motorneuri)ns and myotomes preceding growth of motor awlins. Develop- ment 102:387-396.
Lemmon, V.. K.L. Farr, and C. 1:agenaur 11989) L1-mediated axon out- growth occurs via a homophilic binding mechanism. Neuron 2.1597- 1603
Lemmon. V.. S.M. Burden, H.R. I’ayne, G.J. Elmslie,and M.L. Hlavin 119921 Neurite growth on different substrates: permissive versus instructive influences and the role of adhesive s t r e n a h . J. Neurosci. 12.818-826.
Letm~rneau , P.C., A.M. Madsen, S.1,. Palm, and L.T. Furcht (1988) Immuno- reactivity for laminin in the developing ventral longkudinal pathway of the b r a i n I)ev. B i d 12SI35-144.
Liesi, P., and J . Silver 11988) Is astrocyte laminin involved in axon guidance in thr: mamm;ili;in CNS? Dev. Bid . 130:774-785.
Liesi, P., D. Dahl, and A. Vaheri (1984, Neurons cultured from developing rat hrain attach and spread preferentially to laminin. J. Neurosci. 11:241-251.
Liu. I,., and P.G l a y e r I 19841 Spatio-temporal patterns of differentiation of whole heads of the embryonic chick as revealed by binding of a
pled peanut agglutinin IFITC-PNA). Dev. Brain Res. 12173-
Skutelsky, D. Danon, and N. Sharon 11975) The purification. on and specificity of the anti-T lectin from peanut larachis
hypogaeai. J . Uiol. Chem. 250.8518-8523. Manthorpe. M., I!!. E n p d l , E!. Ruoslahti, F.M. Longo. G.E. Davis, and S.
Varon 119831 Laminin promotes neuritic regeneration from cultured peripheral and central neurons. J . Cell Biol. 97.1882.
Miragall, F.. G . Kadmon, and M. Schachner 119891 Expression (if LI and N-CAM cell adhesion molecules during development of the mouse olfactm-y systetn. Dev. Biol. 1.27272-286.
Moos, M.. K. l‘acke. H. Scherer, D. ‘replow, K. Fruh, and M. Schachner I 19881 Neural cell adhesion molecule L1 as a member of the immuno- globulin superfamily with binding domains similar to fibronectin. Na- ture :1:14:701-703.
Neugebauer. K.M., K.J. Tomaselli, J. Lilien, and L.F. Reichardt (1988) N-cadherin, NCAM, and integrins promote retinal neurite outgrowth on astrocytrs in vitro. J Cell B i d 107:1177-1187.
Oakley, K.A.. and K.W. Tosney i 19911 Peanut agglutinin and chondroitin-6- sulfate a re nii)lecular markers for tissues tha t act a s barriers to axon advance in the avian embryo. Dev. Biol. 147:187-206.
Patterson, P .H. 11988) On the importance of being inhibited, o r saying no to growth cones. Neuron 1.263-267.
Pixley. S.K.K.. and C.W. Cotman i 19861 Laminin suppots short-term survival of rat septa1 neurons in low-density, serum-free cultures. J . Neurosci. 15: 1-1 7.
Reichardt. L.F.. and K.J. Tomaselli 119911 Extracellular matrix molecules and their roccptors: functions in neural development. Annu. Rev. Neurnsci. 14:531-570.
Rogers, S.L., P .C Letourneau, S.L. Palm, J. McCarthy, and L.T. Furcht ( 19831 Neurite extension by peripheral and central nervous system neurons in response to substratum-bound fibronectin and laminin. Dev. 13iol. 9H-2 12-220.
Rogers, S.L., K.J. Edson. C. Letourneau, and S.C. McLoon 11986) Distribu- tion of laminin in the developingperipheral nervous system o f t h e chick. Ilev. Diol. I I :1:4 29-435.
Rogers. S.L., P .C Letourneau. and I.V. Pech (1989) The role of fihronectin in neural development. Dev. Neurosci. 11248-265.
Rutishauser, LJ., and L. Landmesser (1991) Polysialic acid on the surface of axons regulates pa t t r rns of normal and activity-dependent innervation. TINS 14.528-532.
Rutishauser, U., M. Grumct. and G.M. Edelman 11983) N-CAM mediate initial interactions hetween spinal cord neurons and muscle cells in culturc:. J . Cell B i d 97.145-1.52.
Sanes. J .R. I 1989) Extracellular matrix molecules tha t influence neural develiipment. Annu. Rev. Neurosci. 12;491-516.
Schwanzel-Fukuda, M., S. Abraham. K.L. Crossin, G.M. Edelman. and D.W. PfafT( 1992, Inimunocytochemical demonstration of neural cell adhesion mnlecule NCAMl along the migmtion route of luteinizing hormone- releasing hormone ILHKHJ neurons in mice. J. Comp. Neural. 321:l-18.
Seilheimer. R., and M. Schachner 11988) Studies of adhesion molecules mediating interactions between cells of peripheral nervous system indicate a major r o k f i r L1 in mediating sensory neuron growth (in Schwann cells in culture. J. Cell Uiol. 107.341-351.
Silver, J. , and U. Rutishauser 119841 Guidance of optic axons in vivo by a perfbrmed adhesive pathway on neuroepithelial endfeet. Dev. Biol. IOfi.485499.
Silver, J., M. Postiin, and U. Kutishauser (1987) Axon pathway boundaries in the developing brain. I. Cellular and molecular determinants tha t separate the optic and olfactory projections. J . Neurnsci. 7.2264-2272.

14
Snow, 1I.M.. V. Lemmon, D.A. Carrino, A.I. Caplan, and J . Silver (19901 Sulfatcd proteoglycans in astroglial barriers inhibit neurite outgrowth in c i f m Exp. Neurol. 109:ll l-130.
Terkelhen. O.B.F., E. Bock, and K. Mollgard (19891 NCAM and Thy-I in special sense organs of the developing mouse. Anat. Embryol. iBerl.! 279:311-318.
Thiery. J.P.. J .L . Duband, U. Rutishauser, and G.M. Edelman (1982, Cell adhesicin molecules in early chicken embryogenesis. Proc. Natl. Acad. Sci. C.S.A. 79:6737-6741.
Timpl, K.. and H. Rohde (1979) Laminin-a glycoprotein from basement membranes. J . Biol. Chem. 251;9933-9937.
Timpl, K.. J. Engel, and G.R. Martin (1983) Laminin-a multifunctional protein of' basement membranes. Trends Bid. Sci. Junrt207-209.
Q. GONG AND M.T. SHIPLEY
'I'osney, K.W., and K.A. Oakley i 1990) The perinotochordal mesenchyme acts as a barrier to axon advance in the chick embryo: implications for a general mechanism ofaxonal guidance. Exp. Neurobiol. 109:75-89.
Walsh, F.S., and P. I h h e r t y (1991 1 Glycosylphosphatidylinositol anchored recognition molecules tha t function in axonal fasciculation, growth and guidance in the nervous system. Cell Biol. Int . Kep. I5:1151-1166.
Williams, A.F., and A.N. Barclay i 19881 The immunoglobulin superfamily- domains for cell surfice recognition. Annu. Rev. Immunol. 6381-405.
Yip, J .W., and Y.P.L. Yip (1992) 1,aminin-developmental expression and role in axnnal nutgrnwth in the peripheral nervous system of the chick. Dev. I3rain Kes. 6833-33.
Zhou, F.C. (1990) Four patterns of laminin-immunoreactive structure in developing ra t brain. Dev. Brain Res. 55:191-201.





























