Embed Size (px)
Citation preview

2241Journal of Cell Science 108, 2241-2251 (1995)Printed in Great Britain © The Company of Biologists Limited 1995
Expression of the β6 integrin subunit in development, neoplasia and tissue
repair suggests a role in epithelial remodeling
J. M. Breuss1, J. Gallo2, H. M. DeLisser9, I. V. Klimanskaya1, H. G. Folkesson8, J. F. Pittet4, S. L. Nishimura1,5, K. Aldape6, D. V. Landers7, W. Carpenter11, N. Gillett10, D. Sheppard1,8, M. A. Matthay8, S. M. Albelda9, R. H. Kramer2,8 and R. Pytela1,3,8,*1Lung Biology Center, Department of Medicine, Departments of 2Stomatology, 3Pharmacology, 4Anesthesiology, 5Pathology,6Neuropathology, 7Obstetrics and Gynecology and the 8Cardiovascular Research Institute, University of California at SanFrancisco, San Francisco, CA 94143, USA9Department of Medicine, University of Pennsylvania, Philadelphia, PA 19104, USA10Genentech Inc., South San Francisco, CA 94080, USA11University of the Pacific School of Dentistry, San Francisco, CA 94115, USA
*Author for correspondence at address 1
The αvβ6 integrin was identified in cultured epithelial cellsand functions as a fibronectin receptor. We have now usedmonoclonal antibodies to determine in vivo expressionpatterns of the β6 subunit in normal and pathologicalhuman or primate tissues, and during experimental woundhealing or induced lung injury. The results indicate that β6expression is restricted to epithelia and is up-regulated inparallel with morphogenetic events, tumorigenesis, andepithelial repair. During development of the kidney, lung,and skin, we found that β6 is expressed by specific types ofepithelial cells, whereas it is mostly undetectable in normaladult kidney, lung and skin. In contrast, we detected high-level expression in several types of carcinoma. For example,β6 is almost invariably neo-expressed in squamous cell car-cinomas derived from the oral mucosa, often focallylocalized at the infiltrating edges of tumor islands.
Expression of β6 is also upregulated in migrating ker-atinocytes at the wound edge during experimentalepidermal wound healing. Similarly, β6 expression isinduced in type II alveolar epithelial cells during lunginjury caused by injection of live bacteria. We alsoobserved β6 expression in adult lungs and kidneys at focalsites of subclinical inflammation, as well as in a variety ofclinical specimens from patients with chronic or acuteinflammation of the lungs or kidneys. From these findingsand earlier results, we hypothesize that αvβ6 affects cellspreading, migration and growth during reorganization ofepithelia in development, tissue repair, and neoplasia.
Key words: carcinoma, kidney, lung, wound healing, inflammation,fetal development
SUMMARY
INTRODUCTION
Integrins are a family of cell surface receptors that mediatecell adhesion to each other or to extracellular matrix substrata(Hynes, 1992; Giancotti and Mainiero, 1994). In addition toproviding anchorage for stationary cells and traction duringcell movement, integrins are believed to modulate cell differ-entiation, gene expression, proliferation, and survival.Integrins are composed of one α subunit and one β subunit,both of which are transmembrane glycoproteins consisting oflarge extracellular domains and, in most cases, short cyto-plasmic domains. Fifteen α subunits and eight β subunitshave been defined by the corresponding cDNA sequences,and by their antigenic and biochemical properties. Some βsubunits can associate with more than one α subunit, andsome α subunits have multiple β subunit partners. Two largesubfamilies of integrins are defined by the β1 and αvsubunits, respectively. The β1 subunit can associate with tendifferent α subunits (α1 through α9, αv), while the αv
subunit forms five different heterodimers (αvβ1, αvβ3,αvβ5, αvβ6, and αvβ8).
The β6 subunit was first identified in cultured epithelialcells as part of the αvβ6 heterodimer, and the αvβ6 complexwas shown to interact with fibronectin (Sheppard et al., 1990;Busk et al., 1992). In addition to fibronectin, αvβ6 may alsorecognize tenascin (Tn-C), based on experiments usingrecombinant fragments containing the third fibronectin-likerepeat of human tenascin (Prieto et al., 1993). In cells platedon fibronectin substrata, αvβ6 localizes in focal contacts(Cone et al., 1994; I. V. Klimanskaya and R. Pytela, unpub-lished). The αvβ6 integrin mediates adhesion and spreadingof colon carcinoma cells on a fibronectin substratum, in thepresence of blocking antibodies to β1-containing fibronectinreceptors (Weinacker et al., 1994). In cultured airway epi-thelial cells, the expression of β6 is up-regulated by TGF-β(Sheppard et al., 1992). Colon carcinoma cells transfectedwith the β6 cDNA attain increased potential to proliferate andsurvive inside a collagen gel (Agrez et al., 1994).

2242 J. M. Breuss and others
Expression patterns of different β1 integrins have beenstudied in some detail, while only very limited information isavailable on the tissue distribution and regulation of most αvintegrins. The distribution of β6 mRNA in adult primate tissueshas been studied by in situ hybridization (Breuss et al., 1993).The β6 mRNA was only detected in epithelial cells and wasvery low or undetectable in most normal tissues. High-levelexpression was observed only in secretory endometrial glands.Low-level expression was detected in the ductal epithelia ofsalivary gland, mammary gland and epididymis, in gall- andurinary bladder, and in the digestive tract.
We have now used monoclonal antibodies to human β6 inorder to examine its expression pattern in normal adult andfetal tissues and in response to injury, inflammation andmalignant transformation. We have found that β6 is a devel-opmentally regulated integrin subunit that is downregulated indifferentiated adult epithelia, and re-expressed in injured andinflamed epithelia, and in at least some types of epithelial-derived tumors.
MATERIALS AND METHODS
TissuesHuman fetal tissues were collected following elective pregnancy ter-minations, with informed consent using consent forms approved bythe UCSF Committee on Human Research (IRB). Rhesus monkeytissues were obtained either two days after birth (newborn) or from a4 year old animal. These tissues were either part of a Rhesus monkeytissue bank maintained at Genentech, Inc., or leftover tissues fromanimals sacrificed for other purposes. Adult human lung and kidneytissues were obtained from normal margins of tumor resections.Specimens from 5 different individuals were analyzed, with essen-tially identical results. Pathological kidney tissues used in this studyinclude one specimen each of arterial nephrosclerosis, acute tubularnecrosis, chronic pyelonephritis, acute diffuse thrombotic glomeru-lopathy, and 10 rejected renal transplants. Human lung tissues withinflammatory changes examined for β6 expression include specimensof organizing pneumonia, chronic interstitial pneumonitis, bronchi-olitis obliterans and extrinsic allergic alveolitis and immune complexmediated pulmonary hemorrhage (one to three specimens each). Allof these clinical specimens were leftover archival samples obtainedthrough the UCSF Department of Pathology. Normal oral mucosabiopsies, and 30 head and neck squamous cell carcinoma specimenswere obtained from a tissue bank maintained at the UCSF Departmentof Stomatology. All tissues were either directly embedded in OCT(optimal cutting temperature compound; Miles Laboratories)embedding medium and snap-frozen or first immersed in 10% (w/v)sucrose at 4°C for 4 hours and then embedded (fetal tissues) andfrozen.
AntibodiesTwo monoclonal antibodies recognizing αvβ6 were used for immuno-histochemical staining. The monoclonal antibody 9G6 was raised byimmunizing mice with human recombinant secreted αvβ6 (I. V. Kli-manskaya and R. Pytela, unpublished; and Weinacker et al., 1994).The anti-β6 monoclonal antibody R6G9 was raised to transfectedNIH3T3 cells expressing β6, as previously described (Weinacker etal., 1994). Hybridoma culture supernatants were used for staining atdilutions ranging from 1:5 to 1:25, corresponding to antibody con-centrations of approx. 1-5 µg/ml. Unless indicated otherwise, theresults shown were obtained by employing monoclonal antibody 9G6.
Immunohistochemistry Cryostat sections (5 µm) were air dried for 30 minutes, fixed in −18°C
acetone for 5 minutes, and air dried again. Endogenous peroxidaseactivity was blocked with Peroxoblock Solution (Zymed Laborato-ries, South San Francisco, CA.) for 45 seconds at room temperature.After rinsing in PBS for 15 minutes to remove the OCT, sections wereincubated with 0.5% casein/0.05% thimerosal/PBS for 30 minutes atroom temperature and then incubated overnight at 4°C, or for 2 hoursat room temperature, with either the 9G6 or the R6G9 monoclonalantibody (diluted in Biomeda primary antibody dilution buffer). Afterrinsing in automation buffer (Biomeda, Foster City, CA), sectionswere incubated in peroxidase-conjugated horse anti-mouse secondaryantibody (Vector Laboratories, Burlingame, CA) at a dilution of 1:200in PBS with 1% horse serum for 1 hour at room temperature. Tissuesections were rinsed and then incubated with diaminobenzidine (DABPlus Kit, Zymed Laboratories) for 8 minutes at room temperature.Counterstaining was performed with hematoxylin. Sections were sub-sequently dehydrated in graded ethanol solutions, transferred intoHemo-De (Fisher Scientific, Pittsburgh, PA) and then into xylene, andfinally mounted with Permount (Fisher Scientific, Pittsburgh, PA).Negative controls were performed by using an irrelevant primaryantibody, the mouse monoclonal anti-trinitrophenyl antibodyproduced by the hybridoma 1B7.11 (obtained from the AmericanType Culture Collection).
Skin grafting and wounding A detailed description has been published previously (Juhasz et al.,1993). Briefly, severe combined immunodeficient (SCID) mice weretransplanted with human full-thickness neonatal foreskin. After 4weeks, the grafts were completely healed. Longitudinal dermo-epi-thelial excisional wounds, approximately 8 mm long and 2 mm wide,were made and the depth of the wound was adjusted so as not toextend past the human dermis. At the designated time points afterwounding, the mice were euthanized and the wounded skin graft, withthe surrounding murine skin, was removed, snap-frozen in OCT, andstored at −70°C.
Model for acute lung injuryIn order to study β6 expression in the adult lung in the presence ofacute inflammation, we used a well established model of acute lunginjury that develops within a few hours following the intravenousinjection of live bacteria (Brigham et al., 1974; Wiener-Kronish et al.,1991; Pittet et al., 1994). In this model, the injection of live bacteriaresults in a marked increase in lung endothelial permeability withaccumulation of protein-rich edema fluid in the interstitium of thelung along with sequestration of neutrophils in the microcirculationof the lung. This is a standard model of human septicemia that hasbeen used by us and other investigators to study mechanisms of lungendothelial and epithelial injury. In these experiments, 1 ml of 1011
colony forming units (c.f.u.) of live Gram negative bacteria(Pseudomonas aeruginosa, PAO1, Fisher immunotype 7) wereinjected intravenously over 5 minutes in 2 anesthetized, ventilatedsheep. Control studies were done with 2 anesthetized, ventilated sheepthat were given 1 ml of saline intravenously over 5 minutes. Duringthis interval, the animals were supported with positive pressure ven-tilation and supplemental oxygen. At the end of 6 hours, the sheepwere exsanguinated and several samples from both lungs wereremoved and frozen in liquid nitrogen to be analyzed morphologicallyfor β6 mRNA expression. Similar studies were carried out in rabbitsexcept that the intravenous dose of bacteria was 109 c.f.u. of P. aerug-inosa. As in the sheep studies, the rabbits were anesthetized and ven-tilated for 6 hours before they were exsanguinated and the lungsremoved for analysis of β6 mRNA expression. There were 2 rabbitsgiven live bacteria and 2 control rabbits given intravenous saline.
In situ hybridization A 35S-labeled human β6 antisense cRNA probe (868 nucleotides long)was used as previously described (Breuss et al., 1993). As a control,we used a cRNA probe prepared under identical conditions, but cor-

2243Integrin β6 in epithelial remodeling
responding to the sense strand of the β6 cDNA. Rabbit or sheeptissues were fixed in 4% paraformaldehyde at 4°C for 2 hours,embedded, frozen, and thaw-mounted onto glass slides. In situ hybrid-ization was performed as described before (Breuss et al., 1993) withthe exception of using 52°C for the final wash instead of 55°C.Hybridizations were performed in 50% formamide, 0.3 M NaCl, 20mM Tris, pH 8.0, 5 mM EDTA, 1× Denhardt’s solution, 10% dextransulfate, 10 mM dithiothreitol, and 0.05% yeast tRNA, for 16 hours at55°C. Slides were treated with RNase A solution (20 µg/ml) for 30minutes at room temperature, and the final wash was performed with0.1× SSC for 2 hours at 52°C.
RESULTS
Antibody specificity, and expression of the β6integrin subunit in normal adult primate tissuesTwo different anti-β6 monoclonal antibodies, which are likelyto recognize distinct epitopes, were used throughout this study.The monoclonal antibody 9G6 was raised by immunizing micewith human recombinant secreted αvβ6, and was shown tospecifically recognize the β6 subunit and/or the αvβ6 het-erodimer by flow cytometry, indirect immunofluorescencestaining of cultured cells, and immunoprecipitation (I. V. Kli-manskaya and R. Pytela, unpublished). The anti-β6 mono-clonal antibody R6G9 was raised to transfected cells express-ing β6, as previously described (Weinacker et al., 1994).Epitopes recognized by these two antibodies are likely to bedistinct because R6G9 cross-reacts with Rhesus monkeytissues, while 9G6 does not. Most of the human tissue stainingexperiments were performed with both 9G6 and R6G9 andproduced identical staining patterns. This strongly suggeststhat the positive signals are indeed due to the presence of β6,and not to coincidental cross-reaction of the antibodies withother molecules.
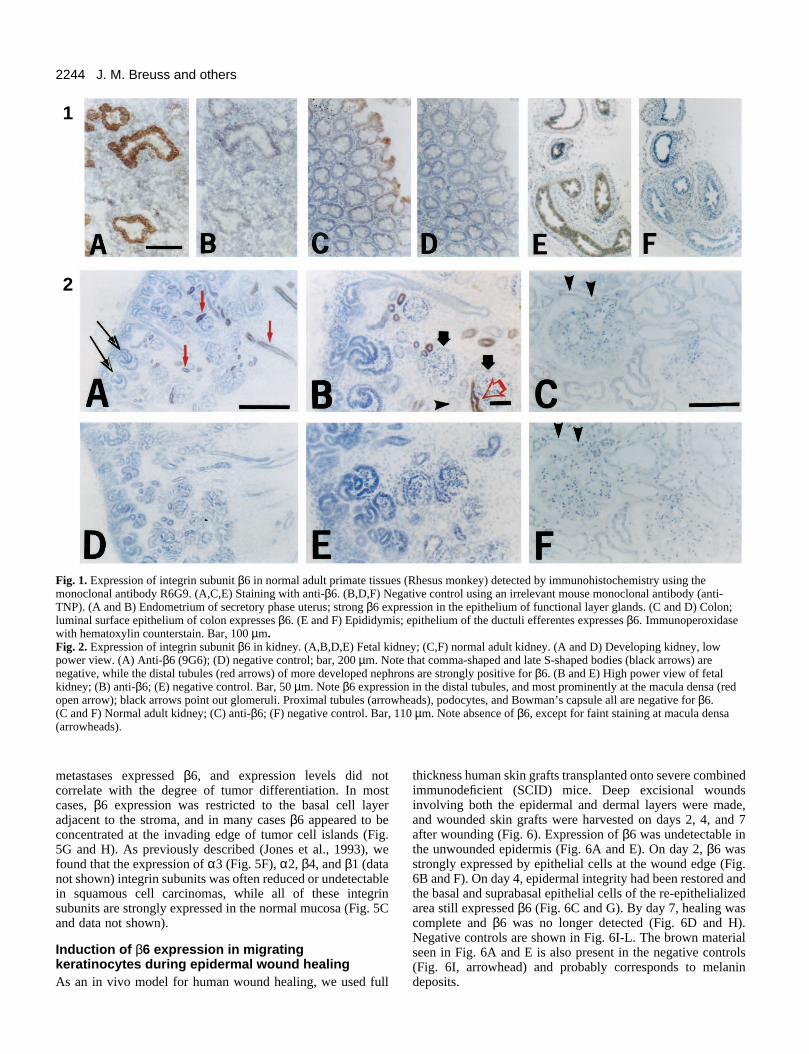
Immunohistochemical staining of normal adult Rhesusmonkey or human tissues revealed that the distribution of β6protein parallels the previously described distribution of β6mRNA (Breuss et al., 1993). Only epithelial cells werepositive, and the strongest expression was seen in secretoryendometrial glands (Fig. 1A). As previously described for β6mRNA, β6 protein expression was very low in the basal layerof the endometrium, and gradually increased in the functionallayer, being highest in glands close to the luminal surface ofthe uterus. The luminal surface epithelium was also positive.Fig. 1 also shows low-level β6 expression in colon (Fig. 1C),and epididymis (Fig. 1E). Epithelial cells in salivary andmammary gland ducts, in gall bladder, urinary bladder, andureter were also weakly positive (data not shown). Negativecontrol staining with an irrelevant antibody produced no signalin any of these tissues (Fig. 1B, D, and F).
The matching distribution patterns obtained by antibodystaining and in situ hybridization further confirm that thesignals obtained in the antibody staining experiments arespecific.
Expression of β6 during epithelial development offetal kidney, lung, and skinThe development of nephrons provides a convenient model forstudying epithelial differentiation. It begins with a mesenchy-mal-epithelial transition leading to formation of primary
epithelial vesicles, which further differentiate and expandgoing through successive stages termed comma-shaped bodiesand S-shaped bodies (Fig. 2A, black arrows), eventuallyforming nephrons that consist of the differentiated epithelia ofdistal and proximal tubules, glomerular podocytes andBowman’s capsule. Since new nephrons are continuouslyformed throughout fetal development and even after birth, allof these stages can be observed simultaneously at any timepoint throughout mid- or late gestation.
Fig. 2A and B show staining for β6 in human fetal kidneyat approx. 18 weeks of gestation. Negative controls are shownin Fig. 2D and E. At early stages of nephrogenesis (vesicles,comma- and S-shaped bodies), β6 is not detected (black arrowsin Fig. 2A). In more developed fetal nephrons (characterizedby capillary loop stage glomeruli), intense β6 staining is foundin distal tubules (red arrows in Fig. 2A), especially at themacula densa (red open arrow in Fig. 2B) and at the loopbetween distal and proximal tubules (early loop of Henle).Proximal tubules (arrowhead in Fig. 2B) and glomeruli (blackarrows in Fig. 2B) do not express detectable amounts of β6. Innormal kidney of the adult, β6 staining is barely detectableeven at the macula densa (arrowheads in Fig. 2C). Thus, itappears that high level expression of β6 in distal tubules istransient during nephrogenesis, and is down-regulated in adultkidney. We also detected β6 expression in medullary collect-ing ducts (not shown) of fetal, but not adult, kidney.
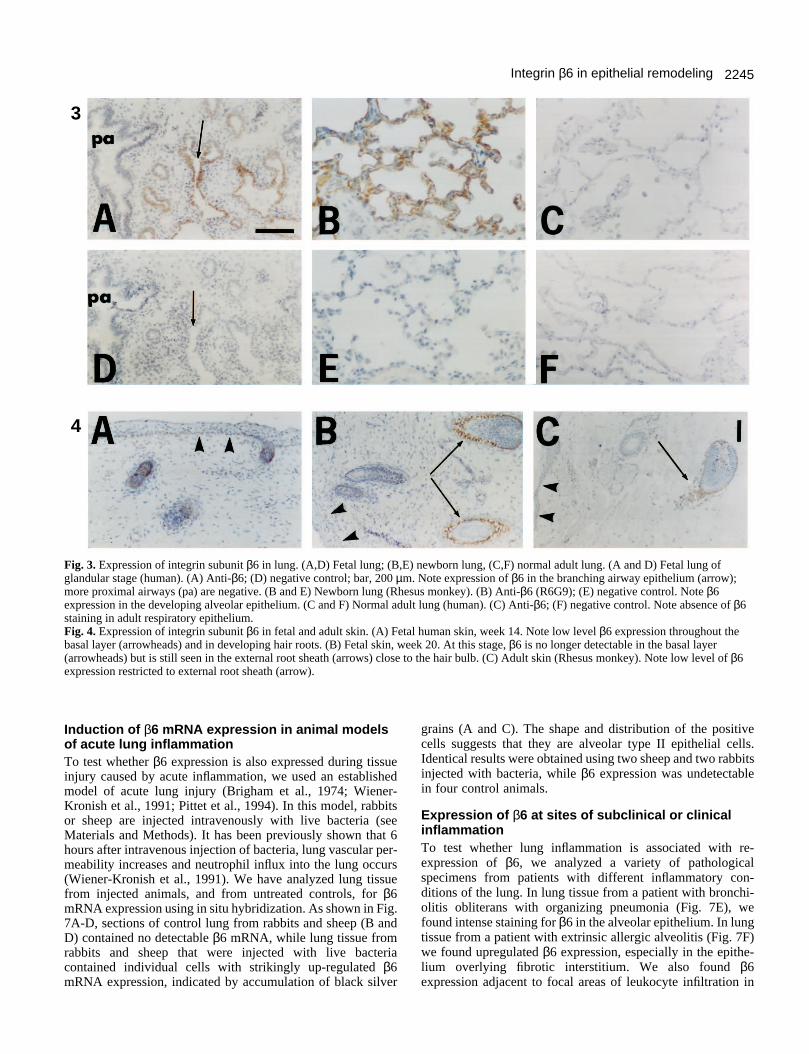
In human fetal lung at 14-24 weeks of gestation, moderateβ6 expression was observed in epithelial cells of branchingdistal airways (Fig. 3A, arrow). The proximal airways (pa),which are larger and more differentiated, were mostly negative.In the lung of a newborn Rhesus monkey, β6 expression wasrarely detected in airways (not shown). In contrast, strong β6expression was detected in the alveolar epithelium, whichforms and expands mostly after birth (Fig. 3B). This expressionof β6 was seen in the type I epithelial cells lining the alveoli,and also in type II cells and in cells lining respiratory bron-chioles. In normal adult lung tissue, β6 was absent from bothairways and alveoli (Fig. 3C).
In human fetal skin at 14 weeks of gestation, β6 was weaklyexpressed throughout the basal layer of the epidermis (Fig.4A). Expression was more pronounced in the primordia of hairroots. By week 21, and also in the newborn and adult,expression in the epidermis was no longer detectable.However, expression in the external root sheath of hair rootswas high at week 21 (Fig. 4B) and could still be seen in adultskin (Fig. 4C).
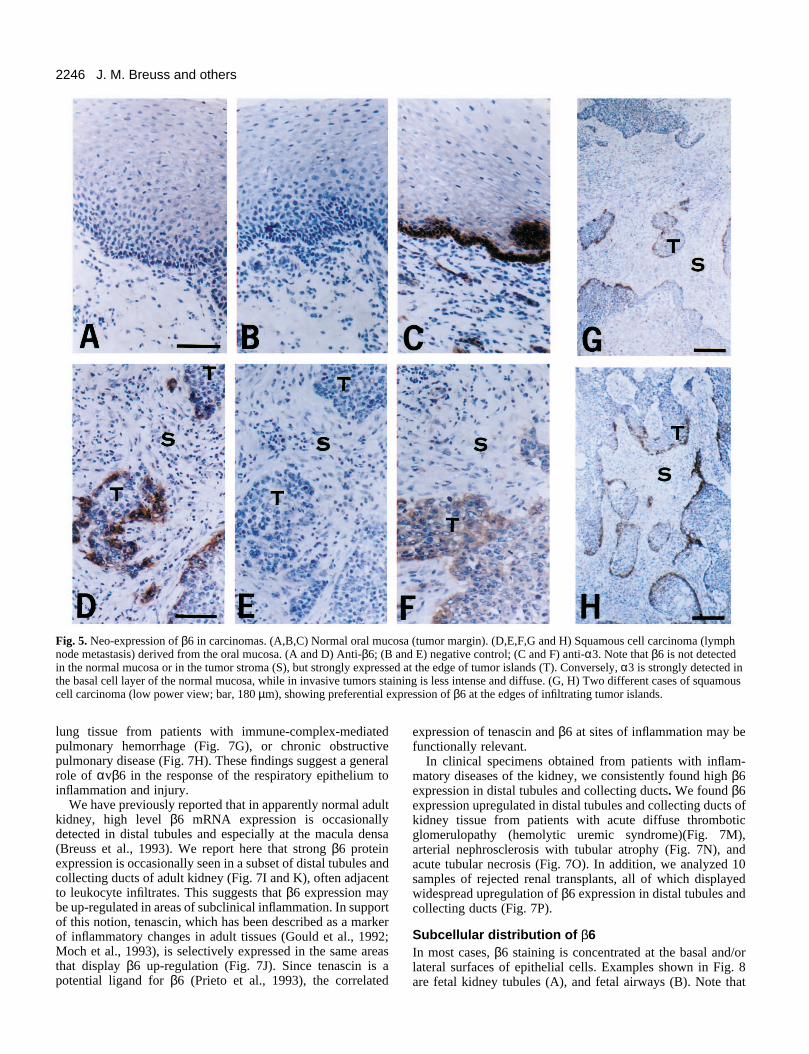
Neo-expression of β6 in squamous cell carcinomasarising from the oral mucosaThe widespread expression of β6 in carcinoma cell linessuggests that carcinomas in vivo may also express this integrinsubunit. Our initial survey of a variety of human carcinomatissues showed that β6 expression was frequently detectable incarcinomas of the lung, breast, pancreas, ovary, colon, and skin(data not shown). To obtain systematic data on a specific typeof carcinoma, we have analyzed 30 specimens of squamouscell carcinomas derived from the oral cavity, and adjacent areasof normal oral mucosa. In 27 of the tumor specimens, β6 wasstrongly expressed in a subset of the carcinoma cells (Fig. 5D,G, and H). Expression of β6 was undetectable in the normaloral mucosa (Fig. 5A). Both primary tumors and lymph node
2244 J. M. Breuss and others
1
2
Fig. 1. Expression of integrin subunit β6 in normal adult primate tissues (Rhesus monkey) detected by immunohistochemistry using themonoclonal antibody R6G9. (A,C,E) Staining with anti-β6. (B,D,F) Negative control using an irrelevant mouse monoclonal antibody (anti-TNP). (A and B) Endometrium of secretory phase uterus; strong β6 expression in the epithelium of functional layer glands. (C and D) Colon;luminal surface epithelium of colon expresses β6. (E and F) Epididymis; epithelium of the ductuli efferentes expresses β6. Immunoperoxidasewith hematoxylin counterstain. Bar, 100 µm.Fig. 2. Expression of integrin subunit β6 in kidney. (A,B,D,E) Fetal kidney; (C,F) normal adult kidney. (A and D) Developing kidney, lowpower view. (A) Anti-β6 (9G6); (D) negative control; bar, 200 µm. Note that comma-shaped and late S-shaped bodies (black arrows) arenegative, while the distal tubules (red arrows) of more developed nephrons are strongly positive for β6. (B and E) High power view of fetalkidney; (B) anti-β6; (E) negative control. Bar, 50 µm. Note β6 expression in the distal tubules, and most prominently at the macula densa (redopen arrow); black arrows point out glomeruli. Proximal tubules (arrowheads), podocytes, and Bowman’s capsule all are negative for β6. (C and F) Normal adult kidney; (C) anti-β6; (F) negative control. Bar, 110 µm. Note absence of β6, except for faint staining at macula densa(arrowheads).
metastases expressed β6, and expression levels did notcorrelate with the degree of tumor differentiation. In mostcases, β6 expression was restricted to the basal cell layeradjacent to the stroma, and in many cases β6 appeared to beconcentrated at the invading edge of tumor cell islands (Fig.5G and H). As previously described (Jones et al., 1993), wefound that the expression of α3 (Fig. 5F), α2, β4, and β1 (datanot shown) integrin subunits was often reduced or undetectablein squamous cell carcinomas, while all of these integrinsubunits are strongly expressed in the normal mucosa (Fig. 5Cand data not shown).
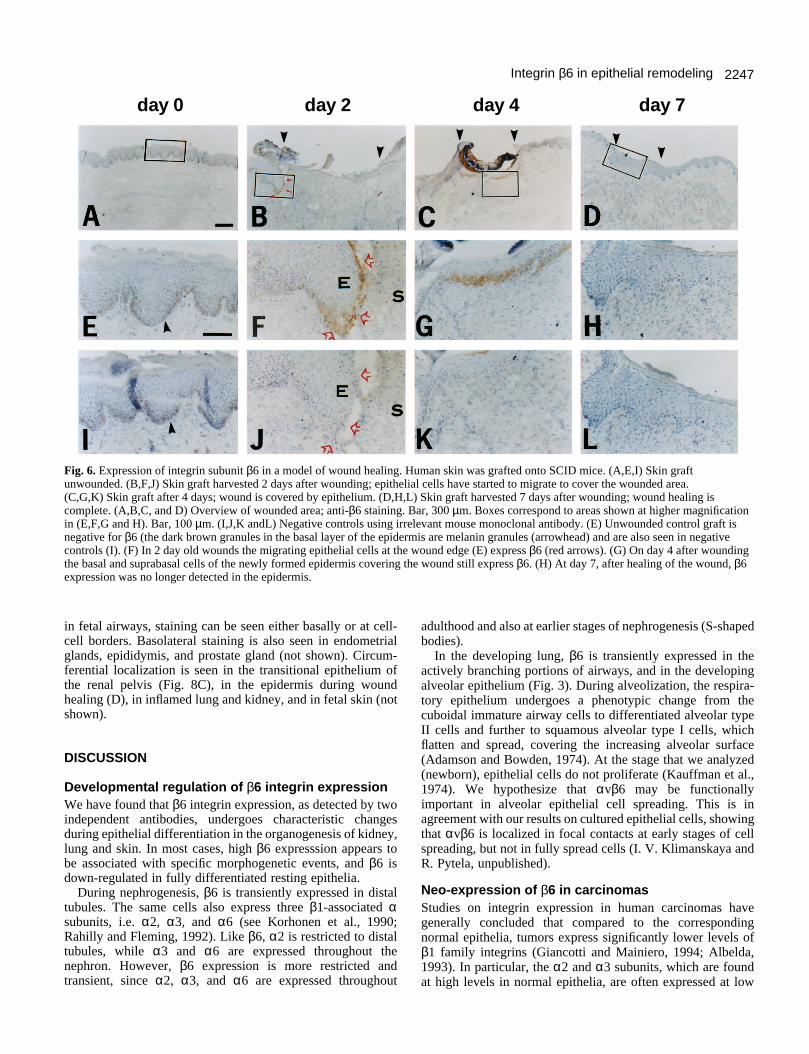
Induction of β6 expression in migratingkeratinocytes during epidermal wound healingAs an in vivo model for human wound healing, we used full
thickness human skin grafts transplanted onto severe combinedimmunodeficient (SCID) mice. Deep excisional woundsinvolving both the epidermal and dermal layers were made,and wounded skin grafts were harvested on days 2, 4, and 7after wounding (Fig. 6). Expression of β6 was undetectable inthe unwounded epidermis (Fig. 6A and E). On day 2, β6 wasstrongly expressed by epithelial cells at the wound edge (Fig.6B and F). On day 4, epidermal integrity had been restored andthe basal and suprabasal epithelial cells of the re-epithelializedarea still expressed β6 (Fig. 6C and G). By day 7, healing wascomplete and β6 was no longer detected (Fig. 6D and H).Negative controls are shown in Fig. 6I-L. The brown materialseen in Fig. 6A and E is also present in the negative controls(Fig. 6I, arrowhead) and probably corresponds to melanindeposits.
2245Integrin β6 in epithelial remodeling
3
4
Fig. 3. Expression of integrin subunit β6 in lung. (A,D) Fetal lung; (B,E) newborn lung, (C,F) normal adult lung. (A and D) Fetal lung ofglandular stage (human). (A) Anti-β6; (D) negative control; bar, 200 µm. Note expression of β6 in the branching airway epithelium (arrow);more proximal airways (pa) are negative. (B and E) Newborn lung (Rhesus monkey). (B) Anti-β6 (R6G9); (E) negative control. Note β6expression in the developing alveolar epithelium. (C and F) Normal adult lung (human). (C) Anti-β6; (F) negative control. Note absence of β6staining in adult respiratory epithelium. Fig. 4. Expression of integrin subunit β6 in fetal and adult skin. (A) Fetal human skin, week 14. Note low level β6 expression throughout thebasal layer (arrowheads) and in developing hair roots. (B) Fetal skin, week 20. At this stage, β6 is no longer detectable in the basal layer(arrowheads) but is still seen in the external root sheath (arrows) close to the hair bulb. (C) Adult skin (Rhesus monkey). Note low level of β6expression restricted to external root sheath (arrow).
Induction of β6 mRNA expression in animal modelsof acute lung inflammationTo test whether β6 expression is also expressed during tissueinjury caused by acute inflammation, we used an establishedmodel of acute lung injury (Brigham et al., 1974; Wiener-Kronish et al., 1991; Pittet et al., 1994). In this model, rabbitsor sheep are injected intravenously with live bacteria (seeMaterials and Methods). It has been previously shown that 6hours after intravenous injection of bacteria, lung vascular per-meability increases and neutrophil influx into the lung occurs(Wiener-Kronish et al., 1991). We have analyzed lung tissuefrom injected animals, and from untreated controls, for β6mRNA expression using in situ hybridization. As shown in Fig.7A-D, sections of control lung from rabbits and sheep (B andD) contained no detectable β6 mRNA, while lung tissue fromrabbits and sheep that were injected with live bacteriacontained individual cells with strikingly up-regulated β6mRNA expression, indicated by accumulation of black silver
grains (A and C). The shape and distribution of the positivecells suggests that they are alveolar type II epithelial cells.Identical results were obtained using two sheep and two rabbitsinjected with bacteria, while β6 expression was undetectablein four control animals.
Expression of β6 at sites of subclinical or clinicalinflammationTo test whether lung inflammation is associated with re-expression of β6, we analyzed a variety of pathologicalspecimens from patients with different inflammatory con-ditions of the lung. In lung tissue from a patient with bronchi-olitis obliterans with organizing pneumonia (Fig. 7E), wefound intense staining for β6 in the alveolar epithelium. In lungtissue from a patient with extrinsic allergic alveolitis (Fig. 7F)we found upregulated β6 expression, especially in the epithe-lium overlying fibrotic interstitium. We also found β6expression adjacent to focal areas of leukocyte infiltration in
2246 J. M. Breuss and others
Fig. 5. Neo-expression of β6 in carcinomas. (A,B,C) Normal oral mucosa (tumor margin). (D,E,F,G and H) Squamous cell carcinoma (lymphnode metastasis) derived from the oral mucosa. (A and D) Anti-β6; (B and E) negative control; (C and F) anti-α3. Note that β6 is not detectedin the normal mucosa or in the tumor stroma (S), but strongly expressed at the edge of tumor islands (T). Conversely, α3 is strongly detected inthe basal cell layer of the normal mucosa, while in invasive tumors staining is less intense and diffuse. (G, H) Two different cases of squamouscell carcinoma (low power view; bar, 180 µm), showing preferential expression of β6 at the edges of infiltrating tumor islands.
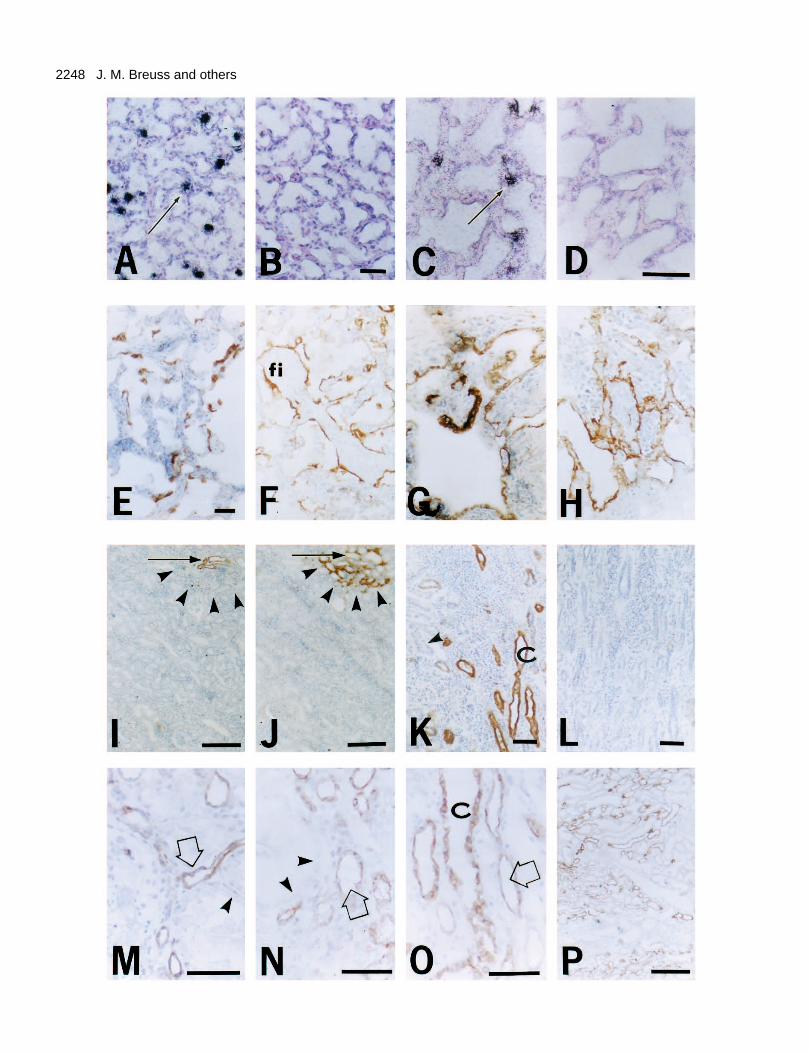
lung tissue from patients with immune-complex-mediatedpulmonary hemorrhage (Fig. 7G), or chronic obstructivepulmonary disease (Fig. 7H). These findings suggest a generalrole of αvβ6 in the response of the respiratory epithelium toinflammation and injury.
We have previously reported that in apparently normal adultkidney, high level β6 mRNA expression is occasionallydetected in distal tubules and especially at the macula densa(Breuss et al., 1993). We report here that strong β6 proteinexpression is occasionally seen in a subset of distal tubules andcollecting ducts of adult kidney (Fig. 7I and K), often adjacentto leukocyte infiltrates. This suggests that β6 expression maybe up-regulated in areas of subclinical inflammation. In supportof this notion, tenascin, which has been described as a markerof inflammatory changes in adult tissues (Gould et al., 1992;Moch et al., 1993), is selectively expressed in the same areasthat display β6 up-regulation (Fig. 7J). Since tenascin is apotential ligand for β6 (Prieto et al., 1993), the correlated
expression of tenascin and β6 at sites of inflammation may befunctionally relevant.
In clinical specimens obtained from patients with inflam-matory diseases of the kidney, we consistently found high β6expression in distal tubules and collecting ducts. We found β6expression upregulated in distal tubules and collecting ducts ofkidney tissue from patients with acute diffuse thromboticglomerulopathy (hemolytic uremic syndrome)(Fig. 7M),arterial nephrosclerosis with tubular atrophy (Fig. 7N), andacute tubular necrosis (Fig. 7O). In addition, we analyzed 10samples of rejected renal transplants, all of which displayedwidespread upregulation of β6 expression in distal tubules andcollecting ducts (Fig. 7P).
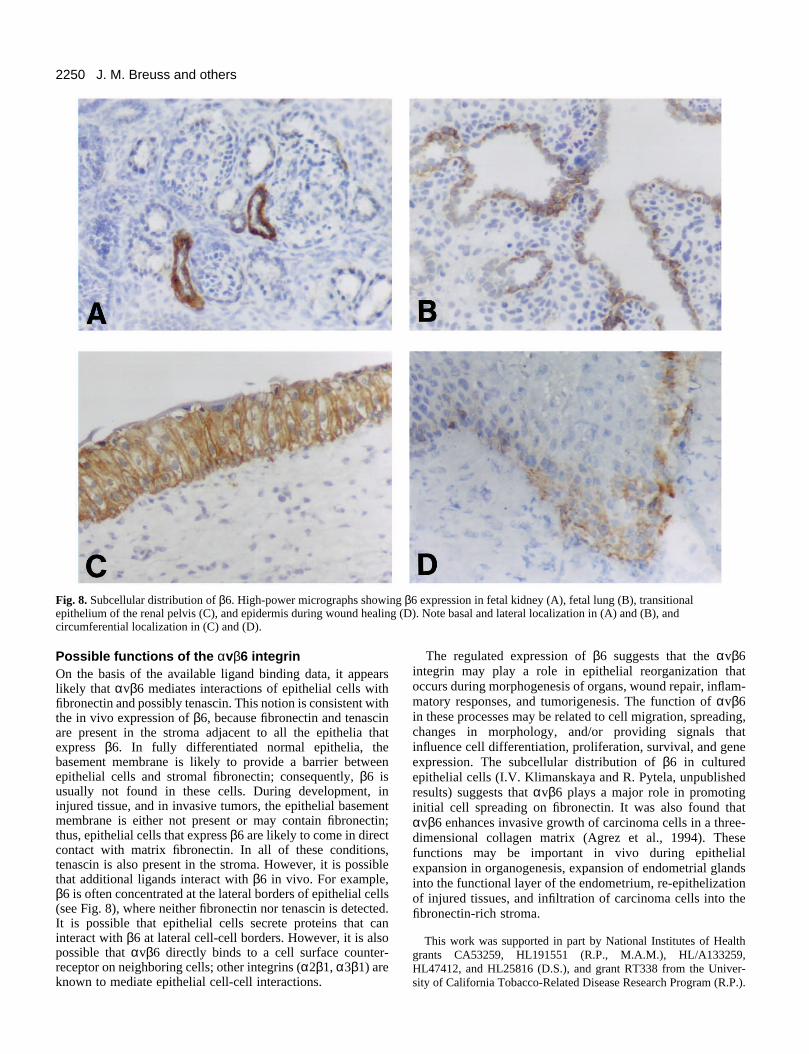
Subcellular distribution of β6In most cases, β6 staining is concentrated at the basal and/orlateral surfaces of epithelial cells. Examples shown in Fig. 8are fetal kidney tubules (A), and fetal airways (B). Note that
2247Integrin β6 in epithelial remodeling
day 0 day 2 day 4 day 7
Fig. 6. Expression of integrin subunit β6 in a model of wound healing. Human skin was grafted onto SCID mice. (A,E,I) Skin graftunwounded. (B,F,J) Skin graft harvested 2 days after wounding; epithelial cells have started to migrate to cover the wounded area. (C,G,K) Skin graft after 4 days; wound is covered by epithelium. (D,H,L) Skin graft harvested 7 days after wounding; wound healing iscomplete. (A,B,C, and D) Overview of wounded area; anti-β6 staining. Bar, 300 µm. Boxes correspond to areas shown at higher magnificationin (E,F,G and H). Bar, 100 µm. (I,J,K andL) Negative controls using irrelevant mouse monoclonal antibody. (E) Unwounded control graft isnegative for β6 (the dark brown granules in the basal layer of the epidermis are melanin granules (arrowhead) and are also seen in negativecontrols (I). (F) In 2 day old wounds the migrating epithelial cells at the wound edge (E) express β6 (red arrows). (G) On day 4 after woundingthe basal and suprabasal cells of the newly formed epidermis covering the wound still express β6. (H) At day 7, after healing of the wound, β6expression was no longer detected in the epidermis.
in fetal airways, staining can be seen either basally or at cell-cell borders. Basolateral staining is also seen in endometrialglands, epididymis, and prostate gland (not shown). Circum-ferential localization is seen in the transitional epithelium ofthe renal pelvis (Fig. 8C), in the epidermis during woundhealing (D), in inflamed lung and kidney, and in fetal skin (notshown).
DISCUSSION
Developmental regulation of β6 integrin expressionWe have found that β6 integrin expression, as detected by twoindependent antibodies, undergoes characteristic changesduring epithelial differentiation in the organogenesis of kidney,lung and skin. In most cases, high β6 expresssion appears tobe associated with specific morphogenetic events, and β6 isdown-regulated in fully differentiated resting epithelia.
During nephrogenesis, β6 is transiently expressed in distaltubules. The same cells also express three β1-associated αsubunits, i.e. α2, α3, and α6 (see Korhonen et al., 1990;Rahilly and Fleming, 1992). Like β6, α2 is restricted to distaltubules, while α3 and α6 are expressed throughout thenephron. However, β6 expression is more restricted andtransient, since α2, α3, and α6 are expressed throughout
adulthood and also at earlier stages of nephrogenesis (S-shapedbodies).
In the developing lung, β6 is transiently expressed in theactively branching portions of airways, and in the developingalveolar epithelium (Fig. 3). During alveolization, the respira-tory epithelium undergoes a phenotypic change from thecuboidal immature airway cells to differentiated alveolar typeII cells and further to squamous alveolar type I cells, whichflatten and spread, covering the increasing alveolar surface(Adamson and Bowden, 1974). At the stage that we analyzed(newborn), epithelial cells do not proliferate (Kauffman et al.,1974). We hypothesize that αvβ6 may be functionallyimportant in alveolar epithelial cell spreading. This is inagreement with our results on cultured epithelial cells, showingthat αvβ6 is localized in focal contacts at early stages of cellspreading, but not in fully spread cells (I. V. Klimanskaya andR. Pytela, unpublished).
Neo-expression of β6 in carcinomasStudies on integrin expression in human carcinomas havegenerally concluded that compared to the correspondingnormal epithelia, tumors express significantly lower levels ofβ1 family integrins (Giancotti and Mainiero, 1994; Albelda,1993). In particular, the α2 and α3 subunits, which are foundat high levels in normal epithelia, are often expressed at low
2248 J. M. Breuss and others

2249Integrin β6 in epithelial remodeling
or undetectable levels in carcinomas of the colon, breast, andskin (Nigam et al., 1993; Pignatelli et al., 1992; Jones et al.,1993). Also, the basally polarized expression of α6β4 is oftenlost in carcinomas.
A few studies have reported up-regulation or neo-expressionof the αv subunit in carcinomas, but the corresponding βsubunit has not been identified (Pignatelli et al., 1992;Korhonen et al., 1992). We now report that β6 is commonlyexpressed in squamous cell carcinomas derived from the oralmucosa, and may be widespread in carcinomas of variousorigins. This suggests that the αv integrin identified in earlierstudies may be αvβ6. These results raise the possibility thatcarcinoma progression is associated not only with a loss ofcertain integrins, but also an increase in the levels of otherintegrins, such as αvβ6. Another member of the αv integrinsubfamily, αvβ3, was found to be neo-expressed during theinvasive growth phase of melanoma and glioblastoma (Albeldaet al., 1990; Gladson and Cheresh, 1991). Thus, it appears thatβ1 integrins involved in maintaining normal tissue organiz-ation are often lost during invasive tumor growth, while at leastsome αv integrins are upregulated. The notion that αvβ6 mayplay a role in carcinoma invasion is strengthened by our obser-
Fig. 7. Re-expression of β6 during inflammation. (A-D) Expressionof β6 mRNA analyzed by in situ hybridization in rabbit and sheepmodels of lung injury. (E-H) Protein expression of β6 in inflamedadult lung and (I-P) in inflamed adult kidney. Rabbits (A) or sheep(C) were injected intravenously with Pseudomonas aeruginosa orsham-treated (B, D), sacrificed after 5 hours, and the lung tissue wasprocessed for in situ hybridization. The positive hybridization signalis indicated by accumulation of black silver grains. Bar, 50 µm. (Aand C) In lung of rabbits (A) or sheep (C) injected with P.aeruginosa, the β6 probe detects β6 mRNA expression in putativetype II alveolar epithelial cells (black silver grains). (B and D)Negative controls using lung sections of sham-treated animals; no β6mRNA was detected. Using sense strand β6 probe as negativecontrol no signal was detected (data not shown). (E-H) Pathologicaladult lungs, anti-β6; bars, 50 µm. (E) Bronchiolitis obliterans withorganizing pneumonia. Note strong focal β6 expression. (F) Extrinsicallergic alveolitis and (G) immune-complex-mediated pulmonaryhemorrhage. Note strong β6 expression in the epithelium. (H)Chronic obstructive pulmonary disease. Focal β6 expression in aregion of lung parenchyma associated with a focal inflammatoryinfiltrate. (I and J) Adult kidney with focal leukocyte infiltration. (I) Anti-β6; (J) anti-tenascin. Bar, 200 µm. Note β6 expression indistal tubules (arrow) next to inflammatory infiltrate with focaltenascin expression in the same region (arrowheads). Staining of anadjacent section with anti-tenascin antibody is shown (J). Note thattenascin is detected in a larger area (arrowheads) centered around thedistal tubule that expresses β6. (K and L) A case of a more extensiveleucocytic infiltrate in adult kidney. (K) Anti-β6; (L) negativecontrol. Bar, 50 µm. Note high-level expression of β6 in distaltubules and collecting ducts (C). Proximal tubules are negative forβ6 (arrowhead). (M) Kidney sections from a patient with acutediffuse thrombotic glomerulopathy; stained with anti-β6. Noteupregulated expression of β6 in distal tubules (arrow). Kidney from apatient with acute tubular necrosis (N) and kidney with arterialnephrosclerosis and tubular atrophy (O) stained with anti-β6; noteexpression of β6 in distal tubules (arrows) and collecting ducts (C).Proximal tubules (arrowheads) are negative. (P) Rejected renaltransplant stained with anti-β6; bar, 200 µm; expression of β6 is seenin distal tubules and collecting ducts throughout the medulla andmany areas in the cortex.
vation that β6 expression often appears highest at the invadingedge of tumor cell islands (Fig. 5G and H).
We have not detected the α5 subunit in squamous cell car-cinomas (data not shown). Thus, this type of tumor is charac-terized by high β6, low α3, and no α5 expression. In contrast,high levels of all three of these fibronectin receptor subunitsβ6, α3, and α5 are expressed during epidermal wound repair.Further studies will be necessary to determine whether thesedifferences are relevant to the invasive behavior of epithelialtumors.
Wound healingIt has been shown that both α5 and αv are up-regulated at themigrating edge of the healing epidermis in a human skin graftmodel of epidermal wound healing (Larjava et al., 1993;Juhasz et al., 1993). The nature of the β subunit associated withαv has been unclear. We now show that β6 is neo-expressedin the migrating basal epithelial cells during wound healing(Fig. 6), in a localization that is precisely coincident with thearea where αv is up-regulated. This suggests that the αvβ6 het-erodimer is present in these cells, and that the expression ofboth αv and β6 is enhanced by factors, such as TGF-β, that arepresent during wound healing. It has been previously shownthat TGF-β upregulates β6 expression in cultured trachealepithelial cells (Sheppard et al., 1992).
Inflammation and β6 expression Experimental evidence for inflammation-induced β6expression in adult lung is provided by our data showing neo-expression of β6 mRNA in the lung of rabbits or sheep thatwere made septic by injection of live bacteria. Striking up-reg-ulation of β6 mRNA expression observed only 6 hours afterinjection of bacteria (Fig. 7A-D) supports the hypothesis thatβ6 expression is induced at an early stage of inflammation. Theresponding cells appear to be type II pneumocytes, whichagrees with the generally held notion that this cell type is mostimportant in alveolar epithelial repair (Agrez et al., 1994).Type II cells are believed to proliferate, spread and migrate inresponse to injury. During this process, type II cells apparentlylose some of their differentiated properties, such as thepresence of lamellar bodies involved in surfactant production,and eventually are converted into type I cells, which replacethe damaged lining of airspaces. In our experimental model ofacute inflammation, only type II cells express β6; in contrast,in clinical specimens of chronic inflammation and in the devel-oping lung, both type I and type II cells are positive. Thus, β6mRNA expression may initially be induced in type II cells, butprotein expression may persist as type II cells spread out andconvert to type I cells during the alveolar repair process. In thissystem, β6 expression may be induced by factors such as TGF-α, which enhances alveolar epithelial repair in vitro (Kherad-mand et al., 1994), and increases β6 expression in culturedepithelial cells (F. Kheradmand, M. A. Matthay and R. Pytela,unpublished).
Our data support the notion that induction of β6 expressionis part of an early epithelial response to injury and/or inflam-mation. It is interesting to note that in inflammatory tissues, β6is up-regulated only in epithelia that express β6 during devel-opment (distal tubules, collecting ducts, airways, alveolarepithelium).
2250 J. M. Breuss and others
Fig. 8. Subcellular distribution of β6. High-power micrographs showing β6 expression in fetal kidney (A), fetal lung (B), transitionalepithelium of the renal pelvis (C), and epidermis during wound healing (D). Note basal and lateral localization in (A) and (B), andcircumferential localization in (C) and (D).
Possible functions of the αvβ6 integrinOn the basis of the available ligand binding data, it appearslikely that αvβ6 mediates interactions of epithelial cells withfibronectin and possibly tenascin. This notion is consistent withthe in vivo expression of β6, because fibronectin and tenascinare present in the stroma adjacent to all the epithelia thatexpress β6. In fully differentiated normal epithelia, thebasement membrane is likely to provide a barrier betweenepithelial cells and stromal fibronectin; consequently, β6 isusually not found in these cells. During development, ininjured tissue, and in invasive tumors, the epithelial basementmembrane is either not present or may contain fibronectin;thus, epithelial cells that express β6 are likely to come in directcontact with matrix fibronectin. In all of these conditions,tenascin is also present in the stroma. However, it is possiblethat additional ligands interact with β6 in vivo. For example,β6 is often concentrated at the lateral borders of epithelial cells(see Fig. 8), where neither fibronectin nor tenascin is detected.It is possible that epithelial cells secrete proteins that caninteract with β6 at lateral cell-cell borders. However, it is alsopossible that αvβ6 directly binds to a cell surface counter-receptor on neighboring cells; other integrins (α2β1, α3β1) areknown to mediate epithelial cell-cell interactions.
The regulated expression of β6 suggests that the αvβ6integrin may play a role in epithelial reorganization thatoccurs during morphogenesis of organs, wound repair, inflam-matory responses, and tumorigenesis. The function of αvβ6in these processes may be related to cell migration, spreading,changes in morphology, and/or providing signals thatinfluence cell differentiation, proliferation, survival, and geneexpression. The subcellular distribution of β6 in culturedepithelial cells (I.V. Klimanskaya and R. Pytela, unpublishedresults) suggests that αvβ6 plays a major role in promotinginitial cell spreading on fibronectin. It was also found thatαvβ6 enhances invasive growth of carcinoma cells in a three-dimensional collagen matrix (Agrez et al., 1994). Thesefunctions may be important in vivo during epithelialexpansion in organogenesis, expansion of endometrial glandsinto the functional layer of the endometrium, re-epithelizationof injured tissues, and infiltration of carcinoma cells into thefibronectin-rich stroma.
This work was supported in part by National Institutes of Healthgrants CA53259, HL191551 (R.P., M.A.M.), HL/A133259,HL47412, and HL25816 (D.S.), and grant RT338 from the Univer-sity of California Tobacco-Related Disease Research Program (R.P.).

2251Integrin β6 in epithelial remodeling
S.N. was supported by a National Research Service Award(CA09335) from the National Institutes of Health. H.M.D. wassupported by the American Heart Association.
REFERENCES
Adamson, I. Y. R. and Bowden, D. H. (1974). The type II cell as progenitor ofalveolar epithelial regeneration. Lab. Invest. 30, 35-42.
Agrez, M., Chen, R. I., Cone, R. I., Pytela, R. and Sheppard, D. (1994). Thealpha-v beta-6 integrin promotes proliferation of colon carcinoma cellsthrough a unique region of the beta-6 cytoplasmic domain. J. Cell Biol. 127,547-556.
Albelda, S. M. (1993). Role of integrins and other cell adhesion molecules intumor progression and metastasis. Lab. Invest. 68, 4-17.
Albelda, S. M., Mette, S. A., Elder, D. E., Stewart, R., Damjanovich, L.,Herlyn, M. and Buck, C. A. (1990). Integrin distribution in malignantmelanoma: association of the beta 3 subunit with tumor progression. CancerRes. 50, 6757-64.
Breuss, J. M., Gillett, N., Lu, L., Sheppard, D. and Pytela, R. (1993).Restricted distribution of integrin beta 6 mRNA in primate epithelial tissues.J. Histochem. Cytochem. 41, 1521-7.
Brigham, K. L., Woolverton, W. C., Blake, C. H. and Staub, N. C. (1974).Increased sheep lung vascular permeability caused by Pseudomonasbacteremia. J. Clin. Invest. 54, 792-804.
Busk, M., Pytela, R. and Sheppard, D. (1992). Characterization of theintegrin alpha v beta 6 as a fibronectin-binding protein. J. Biol. Chem. 267,5790-6.
Cone, R. I., Weinacker, A., Chen, A. and Sheppard, D. (1994). Effects ofbeta subunit cytoplasmic domain deletions on the recruitment of the integrinalpha v beta 6 to focal contacts. Cell Adhes. Commun. 2, 101-13.
Giancotti, F. G. and Mainiero, F. (1994). Integrin-mediated adhesion andsignaling in tumorigenesis. Biochim. Biophys. Acta 1198, 47-64.
Gladson, C. L. and Cheresh, D. A. (1991). Glioblastoma expression ofvitronectin and the alpha v beta 3 integrin. Adhesion mechanism fortransformed glial cells. J. Clin. Invest. 88, 1924-32.
Gould, V. E., Martinez-Lacabe, V., Virtanen, I., Sahlin, K. M. andSchwartz, M. M. (1992). Differential distribution of tenascin and cellularfibronectins in acute and chronic allograft rejection. Lab. Invest. 67, 71-79.
Hynes, R. O. (1992). Integrins: versatility, modulation, and signaling in celladhesion. Cell 69, 11-25.
Jones, J., Sugiyama, M., Watt, F. M. and Speight, P. M. (1993). Integrinexpression in normal, hyperplastic, dysplastic, and malignant oralepithelium. J. Pathol. 169, 235-43.
Juhasz, I., Murphy, G. F., Yan, H. C., Herlyn, M. and Albelda, S. M.(1993). Regulation of extracellular matrix proteins and integrin cellsubstratum adhesion receptors on epithelium during cutaneous humanwound healing in vivo. Am. J. Pathol. 143, 1458-69.
Kauffman, S. L., Burri, P. H. and Weibel, E. R. (1974). The postnatal growthof the rat lung. II. Autoradiography. Anat. Rec. 180, 63-76.
Kheradmand, F., Folkesson, H. G., Shum, L., Derynck, R., Pytela, R. andMatthay, M. A. (1994). Transforming growth factor-alpha enhancesalveolar epithelial cell repair in a new in vitro model. Am. J. Physiol.: LungCell Mol. Physiol. 267, 728-39.
Korhonen, M., Ylanne, J., Laitinen, L. and Virtanen, I. (1990). The alpha 1-alpha 6 subunits of integrins are characteristically expressed in distinctsegments of developing and adult human nephron. J. Cell Biol. 111, 1245-54.
Korhonen, M., Laitinen, L., Ylanne, J., Koukoulis, G. K., Quaranta, V.,Juusela, H., Gould, V. E. and Virtanen, I. (1992). Integrin distributions inrenal cell carcinomas of various grades of malignancy. Am. J. Pathol. 141,1161-71.
Larjava, H., Salo, T., Haapasalmi, K., Kramer, R. H. and Heino, J. (1993).Expression of integrins and basement membrane components by woundkeratinocytes. J. Clin. Invest. 92, 1425-35.
Moch, H., Torhorst, J., Durmuller, U., Feichter, G. E., Sauter, G. andGudat, F. (1993). Comparative analysis of the expression of tenascin andestablished prognostic factors in human breast cancer. Pathol. Res. Pract.189, 510-14.
Nigam, A. K., Savage, F. J., Boulos, P. B., Stamp, G. W., Liu, D. andPignatelli, M. (1993). Loss of cell-cell and cell-matrix adhesion moleculesin colorectal cancer. Br. J. Cancer 68, 507-14.
Pignatelli, M., Cardillo, M. R., Hanby, A. and Stamp, G. W. (1992).Integrins and their accessory adhesion molecules in mammary carcinomas:loss of polarization in poorly differentiated tumors. Hum. Pathol. 23, 1159-66.
Pittet, J. F., Wiener-Kronish, J. F., McElroy, M. C., Folkesson, H. G. andMatthay, M. A. (1994). Stimulation of lung epithelial liquid clearance byendogenous release of catecholamines in septic shock in anesthetized rats. J.Clin. Invest. 94, 663-71.
Prieto, A. L., Edelman, G. M. and Crossin, K. L. (1993). Multiple integrinsmediate cell attachment to cytotactin/tenascin. Proc. Nat. Acad. Sci. USA 90,10154-8.
Rahilly, M. A. and Fleming, S. (1992). Differential expression of integrinalpha chains by renal epithelial cells. J. Pathol. 167, 327-34.
Sheppard, D., Rozzo, C., Starr, L., Quaranta, V., Erle, D. J. and Pytela, R.(1990). Complete amino acid sequence of a novel integrin beta subunit (beta6) identified in epithelial cells using the polymerase chain reaction. J. Biol.Chem. 265, 11502-7.
Sheppard, D., Cohen, D. S., Wang, A. and Busk, M. (1992). Transforminggrowth factor beta differentially regulates expression of integrin subunits inguinea pig airway epithelial cells. J. Biol. Chem. 267, 17409-14.
Weinacker, A., Chen, A., Agrez, M., Cone, R. I., Nishimura, S., Wayner, E.,Pytela, R. and Sheppard, D. (1994). Role of the integrin alpha v beta 6 incell attachment to fibronectin. Heterologous expression of intact and secretedforms of the receptor. J. Biol. Chem. 269, 6940-8.
Wiener-Kronish, J. P., Albertine, K. H. and Matthay, M. A. (1991).Differential responses of the endothelial aqnd epithelial barriers of the lungin sheep to Escherichia coli endotoxin. J. Clin. Invest. 88, 864-75.
(Received 23 January 1995 - Accepted 23 March 1995)





























