Embed Size (px)
Citation preview

INFECTiON AND IMMUNITY, Mar. 1994, p. 961-9670019-9567/94/$04.00+0Copyright ©D 1994, American Society for Microbiology
Extracellular and Cytoplasmic CuZn Superoxide Dismutasesfrom Brugia Lymphatic Filarial Nematode Parasites
LIANG TANG,' XIU OU,' KIMBERLY HENKLE-DUHRSEN,2 AND MURRAY E. SELKIRK"*Wellcome Research Centre for Parasitic Infections, Department of Biochemistry, Imperial College of Science,
Technology and Medicine, London SW7 2AY, United Kingdom,1 and Department of Biochemistry,Bernhard-Nocht Institute for Tropical Medicine, 2000 Hamburg 36, Germany2
Received 14 October 1993/Returned for modification 17 November 1993/Accepted 10 December 1993
We have isolated full-length cDNAs encoding two distinct types ofCuZn superoxide dismutases (SODs) fromthe filarial nematode parasite Brugia pahangi. The derived amino acid sequences suggested that one class ofcDNAs represented a cytoplasmic form of SOD and the second class represented an extracellular (EC) variant.The predicted proteins were highly homologous to each other, but the sequence of the latter contained an
additional 43 residues at the N terminus, the first 16 of which were markedly hydrophobic, and four potentialsites for N-linked glycosylation. Western blotting (immunoblotting) with an antiserum to a partial SODexpressed in Escherichia coli revealed two proteins with estimated molecular masses of 19 and 29 kDa.Digestion with N-glycanase indicated that the latter protein corresponded to the EC form, as it possessedN-linked oligosaccharide chains at three sites, leaving a peptide backbone with an estimated molecular mass
of 22 kDa, which was consistent with the additional 27 amino acids predicted from the cDNA sequence. Gelfiltration indicated that both enzymes were dimeric in their native forms, in contrast to the human EC-SOD,which is tetrameric. Comparison of the primary structure of the parasite EC-SOD with that of the human ECenzyme revealed two major differences: the N-terminal extension of the parasite enzyme was shorter by 25residues, and it also lacked the C-terminal charged extension which mediates binding to cell surface sulfatedproteoglycans. Lavage of Mongolian jirds infected intraperitoneally with Brugia malayi resulted in the recovery
of filarial CuZn SODs, principally the EC form, indicating that this form of SOD is secreted in vivo. This ECenzyme may contribute to parasite persistence by neutralizing superoxide generated by activated leukocytes,thus acting as both an antioxidant and an anti-inflammatory factor.
Superoxide dismutases (SODs) have been found in almostall aerobes examined and are thought to be essential for thesurvival of organisms in the presence of oxygen (13). Asoriginally formulated, the superoxide theory of oxygen toxicityascribed the damaging effects of high oxygen tension to theformation of the superoxide radical (O2 ), although much ofthe cellular damage inflicted now appears to be due to thesubsequent formation of more reactive species. SODs are
presumed to protect cells by catalyzing the dismutation of 02to hydrogen peroxide and ground state oxygen. Three majorclasses of SODs have been described on the basis of the metalcomposition in the active site, i.e., Fe, Mn, and CuZn SODs.Fe SODs are found primarily in prokaryotes, Mn SODs arefound in both prokaryotes and eukaryotes, and CuZn enzymesare mainly restricted to eukaryotes and are present in virtuallyall yeast, plant, and animal cells (13); however, it has recentlybeen demonstrated that a number of bacteria also expressCuZn SODs (5, 21, 43-45, 50).Two subtypes of CuZn SODs have more recently been
distinguished: cellular and extracellular (EC) enzymes (16, 29).Most of the EC-SOD in the vascular systems of mammals isapparently bound to endothelial cell surfaces and can bedissociated by intravenous injection of heparin (19). This hasbeen interpreted as a means of protecting plasma membranesagainst oxidative attack while not interfering with the signallingproperties of superoxide in EC fluids, e.g., an involvement inneutrophil chemotaxis (35). EC-SODs have also been docu-
* Corresponding author. Phone: 4471 225 8289. Fax: 4471 225 0960.Electronic mail address: [email protected].
mented to occur in several bacterial species (3, 5, 21, 44, 46).Virulent forms of Mycobacterium tuberculosis secrete SOD (2,22), and it has been suggested, as for other intracellularpathogenic bacteria (3), that this may confer resistance to theoxidative burst of phagocytic cells. More direct evidence forsuch a protective role has been obtained via administration ofa monoclonal antibody specific for SOD of Nocardia asteroides(4) and construction of SOD deletion mutants of Brucellaabortus (49). In both cases, inactivation of SOD led to en-
hanced clearance of bacteria from infected mice.Helminth parasites are long-lived pathogens which do not
have the ability to undergo antigenic variation and whosepersistence must owe much to down-regulation and neutral-ization of immune effector functions (25). Although primarilyextracellular organisms, they may still be subjected to reactiveoxygen species from leukocytes targeted to the parasite surfaceby specific antibodies. SODs have been detected enzymaticallyin many helminths, in both somatic extracts and tissue culturefluid (6); indeed, one of the first EC-SODs described was fromTrichinella spiralis (36). More recently, a number of helminthCuZn SODs have been cloned, from Schistosoma mansoni (17,42) and Onchocerca volvulus (15), and one of these shows thecharacteristics of an EC enzyme (42). In this communication,we describe the cloning and characterization of both cellularand EC forms of CuZn SOD from the filarial nematodeparasite Brugia pahangi. Both forms are dimeric and theirderived amino acid sequences are highly homologous. TheBrugia EC-SOD is considerably divergent from mammalianEC-SODs, suggesting that it evolved directly from the parasitecytoplasmic enzyme.
961
Vol. 62, No. 3
on July 25, 2019 by guesthttp://iai.asm
.org/D
ownloaded from

962 TANG ET AL.
MATERIALS AND METHODS
Isolation and sequencing of cDNAs. A cDNA library fromadult B. pahangi in Xgtl1 (39) was screened by hybridizationwith cDNA pA3A, which encodes a cytoplasmic CuZn SODfrom 0. volvulus (15). Plaque lifts were hybridized in 5 x SSC(1 x SSC is 0.15 M NaCl plus 0.015 M sodium citrate)-0.1%sodium dodecyl sulfate (SDS)-0.25% milk powder at 55°C andwashed in 0.2x SSC-0.1% SDS at the same temperature.Positive clones were sequenced by dideoxy chain termination(38) after being subcloned into Bluescript SK vectors. Theextreme 5' ends of the cDNAs were generated by PCRamplification from first-strand adult B. pahangi cDNA withinternal primers linked to an XhoI restriction site and anoligonucleotide containing a NotI restriction site linked to thenematode spliced leader sequence SL1 (20) (5'-GGGCGGCCGCGGG7TllAATTACCAAGTFlllGAG-3'; see Fig. 1). Astandard 100-,u PCR mixture containing 30 ng of cDNA and500 ng of each primer was utilized; the reaction consisted of 30cycles at 94°C for 30 s, 65°C for 1 min, and 72°C for 2 min.
Expression and purification of C-SOD 1. The cDNA cloneC-SOD 1, encoding amino acid residues arginine 10 to glycine108 (see Fig. 1), was ligated to pDS78/RBS11 (47) followingthe addition of a 5' BamHI site and a 3' HindlIl site by PCR.The resultant construct was expressed in Escherichia coli TG2cells as previously described (32). Cells were sonicated inphosphate-buffered saline (PBS)-1.5% n-octyl glucoside-pro-tease inhibitors, and the expressed protein was partially puri-fied by gel filtration on Sepharose HP 16/10 (Pharmacia). Apolyclonal antiserum was raised in a rabbit by intramuscularinoculation of 400 pLg of protein emulsified in Freund's incom-plete adjuvant, which was followed by two subcutaneouslyadministered booster injections of 100 ,ug of protein inFreund's incomplete adjuvant, one after 4 weeks and one after6 weeks. The rabbit was bled after 8 weeks, and the antiserumwas designated rabbit anti-C-SOD 1.
Gel filtration chromatography and assay of SOD activity.Adult Brugia malayi parasites were recovered from the perito-neal cavities of Mongolian gerbils (Meriones unguiculatus)obtained from TRS Laboratories, Athens, Ga., infected morethan 3 months previously. Extracts were prepared by homog-enization for 30 min in a mixture containing 50 mM PBS, pH7.5, and a cocktail of protease inhibitors on ice followed bycentrifugation at 15,000 x g for 30 min (24). Fast proteinliquid chromatography was performed on a Superdex 200 60/16column (60 cm by 1.6 cm; Pharmacia) equilibrated with 50 mMPBS (pH 7.5)-0.024% Triton X-100 at a flow rate of 2.0 mlmin-'. Molecular mass standards (Sigma MW-GF-200) wereused to obtain a standard curve, 200 RI of parasite extract (3mg ml- l) was loaded onto the column, and 2-ml fractions werecollected. SOD activity was measured in column fractions bythe inhibition of cytochrome c reduction, essentially as de-scribed by McCord and Fridovich (30). Reduction of cyto-chrome c was assayed in a solution containing 0.1 mM EDTA,11 mM cytochrome c, 50 ,uM xanthine, and 0.15 U of xanthineoxidase ml 1 in 50 mM phosphate buffer, pH 7.5, and thechange in absorbance over 2 min was monitored at 550 nm.One unit of SOD is defined as the amount which inhibits thereduction of cytochrome c by 50% under the assay conditions.The proportion of CuZn SOD in samples was identified byinhibition with 0.1 mM KCN.SDS-PAGE and Western blotting (immunoblotting). Para-
site extracts, bacterial extracts, and purified protein prepara-tions were resolved by SDS-14% polyacrylamide gel electro-phoresis (PAGE) according to standard protocols. Westernblotting was carried out as previously described (24). The
primary antiserum (rabbit anti-C-SOD 1) was used at adilution of 1/400 in PBS-5% milk powder-0.1% Tween 20overnight at 4°C, and horseradish peroxidase-conjugated anti-rabbit immunoglobulin G was used at a dilution of 1:1,000 for2 h at room temperature in PBS alone. Binding was visualizedwith 4-chloro-1-naphthol at a final concentration of 0.5 mgml- '. Parasite extracts were digested with serial dilutions ofpeptide N-glycosidase F (N-glycanase) prior to electrophoresisas previously described (26).
Secretion of parasite SOD in vivo was determined byrecovery of peritoneal fluid from Mongolian jirds (M. unguicu-latus) infected intraperitoneally with 200 infective larvae of B.malayi 4 months previously. Anesthetized animals were la-vaged with 10 ml of PBS for 10 min, and the fluid wasrecovered, centrifuged at 1,000 x g for 10 min, and filteredthrough 0.2-p.m-pore-size syringe filters to remove microfi-lariae and cells. The fluid was then concentrated on Centricon10 microconcentrators to a volume of 500 plI, and 20 plI wasresolved by SDS-PAGE, blotted, and reacted with the rabbitanti-C-SOD 1 as described above. Control (uninfected) jirdswere lavaged and treated in the same manner. Additionalcontrol experiments were performed in which inflammatorycells were elicited with an intraperitoneal injection of 6%sodium caseinate in saline 4 days before lavage.
Nucleotide sequence accession numbers. The nucleotidesequences reported in this paper have been submitted to theGenBank EMBL data library with accession numbers X76283(B. pahangi EC-SOD mRNA) and X76284 (B. pahangi cyto-plasmic SOD mRNA.
RESULTS
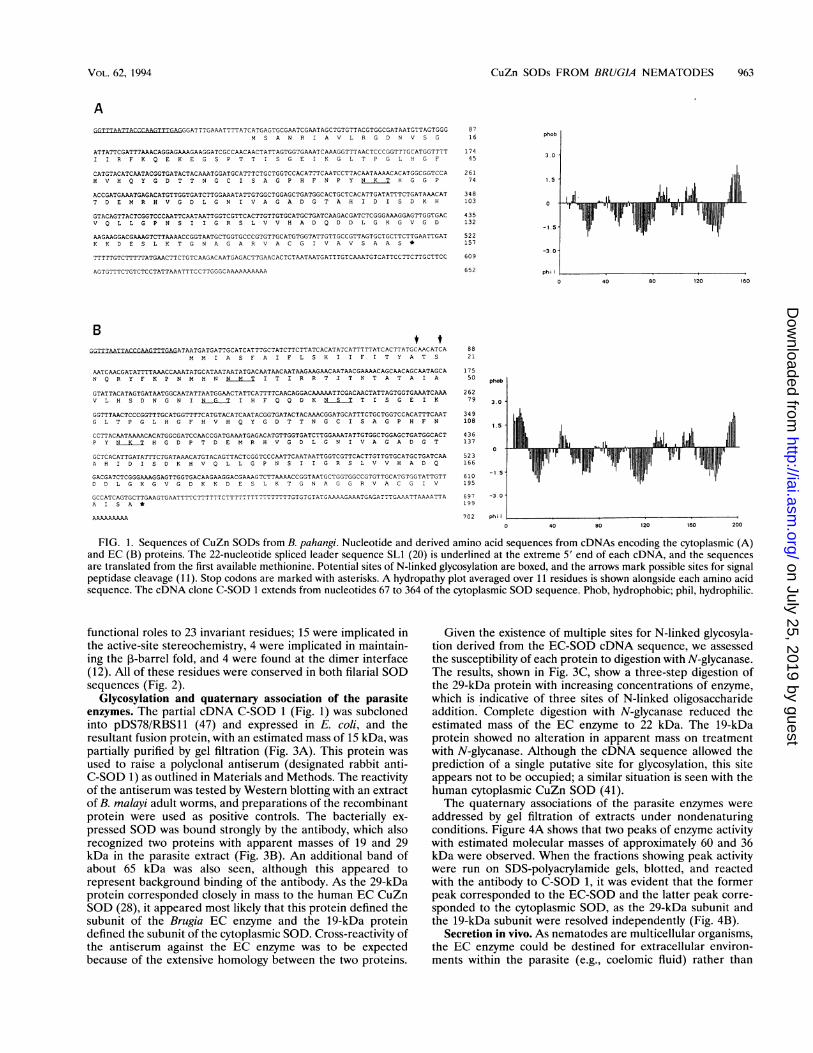
Isolation of cDNAs and primary structure of CuZn SODs.We utilized a cDNA for a CuZn SOD from 0. volvulus (15) asa probe to screen a Xgtl 1 cDNA library constructed from adultB. pahangi RNA. A total of 39 clones were isolated, and onsubsequent characterization they could be grouped into twofamilies by sequence homology. Analysis of the derived proteinsequences from the two cDNA families indicated that oneencoded a cytoplasmic CuZn SOD (Fig. 1A) and the otherencoded an EC form, as the latter contained an N-terminalextension of 43 amino acids, including a markedly hydrophobicputative signal sequence (Fig. 1B). Two good candidates for asignal peptidase cleavage site existed in this protein (11),immediately following alanine 19 and serine 21 and lying 3 and5 residues after the hydrophobic core of the signal sequence,respectively (Fig. 1B). The EC-SOD sequence contained fourpotential sites for N-linked glycosylation, whereas the se-quence of the cytoplasmic SOD contained only one of thesesites (Fig. 1).Homology to mammalian CuZn SODs. The derived amino
acid sequences of the two parasite enzymes were highlyhomologous and showed 88% identity over the core sequence(Fig. 2). Also illustrated is a comparison with the humancytoplasmic and EC CuZn SODs (Fig. 2). The Brugia cytoplas-mic SOD showed 57% identity to its human counterpart,whereas the EC enzymes were only 35% identical over thecores of the mature proteins (i.e., excluding the N-terminalsignal sequences and the C-terminal extension of the humanEC-SOD). In addition, the parasite EC-SOD showed twodistinct differences from the human EC-SOD in (i) lacking aC-terminal extension and (ii) having an N-terminal extensiontruncated by 25 residues.A recent analysis of the structure of CuZn SOD and the
alignment of 15 distinct primary sequences, including one froma bacterial source, has allowed the definition and assignation of
INFEC-F. IMMUN.
on July 25, 2019 by guesthttp://iai.asm
.org/D
ownloaded from
VOL. 62, 1994
AGGTTTAATTACCCAAGTTTGAGGGATTTGAAATTTTATCATGAGTGCGAATCGAATAGCTGTGTTACGTGGCGATAATGTTAGTGGG
M S A N R I A V L R G D N V S G
ATTATTCGATTTAAACAGGAGAAAGAAGGATCGCCAACAACTATTAGTGGTGAAATCAAAGGTTTAACTCCCGGTTTGCATGGTTTTI I R F K Q E K E G S P T T I S G E I K G L T P G L H G F
CATGTACATCAATACGGTGATACTACAAATGGATGCATTTCTGCTGGTCCACATTTCAATCCTTACAATAAAACACATGGCGGTCCAH V H Q Y G D T T N G C I S A G P H F N P Y N K T H G G P
ACCGATGAAATGAGACATGTTGGTGATCTTGGAAATATTGTGGCTGGAGCTGATGGCACTGCTCACATTGATATTTCTGATAAACATT D E M R H V G D L G N I V A G A D G T A H I D I S D K H
GTACAGTTACTCGGTCCCAATTCAATAATTGGTCGTTCACTTGTTGTGCATGCTGATCAAGACGATCTCGGGAAAGGAGTTGGTGACV Q L L G P N S I I G R S L V V H A D Q D D L G K G V G D
AAGAAGGACGAAAGTCTTAAAACCGGTAATGCTGGTGCCCGTGTTGCATGTGGTATTGTTGCCGTTAGTGCTGCTTCTTGAATTGATK K D E S L K T G N A G A R V A C G I V A V S A A S *
TTTTTGTCTTTTTATGAACTTCTGTCAAGACAATGAGACTTGAACACTCTAATAATGATTTGTCAAATGTCATTCCTTCTTGCTTCC
AGTGTTTCTGTCTCCTATTAAATTTCCTTGGGCAAAAAAAAAA
CuZn SODs FROM BRUGL4 NEMATODES 963
8716
17445
26174
348103
435132
522157
609
652
40 8A 120 160
BGGTTTAATTACCCAAGTTTGAGATAATGATGATTGCATCATTTGCTATCTTCTTATCACATATCATTTTTAT CACTTATGCAACATCA
M M I A S F A I F L S H I I F I T Y A T S
AATCAACGATATTTTAAACCAAATATGCATAATAATATGACAATAACAATAAGAAGAACAATAACGAAAACAGCAACAGCAATAGCAN Q R Y F K P N M H N N M.l T I T I R R T .I T K T A T A I A
GTATTACATAGTGATAATGGCAATATTAATGGAACTATTCATTTTCAACAGGACAAAAATTCGACAACTATTAGTGGTGAAATCAAAV L H S D N G N I N GT I H F Q Q D K N S T T I S G E I K
GGTTTAACTCCCGGTTTGCATGGTTTTCATGTACATCAATACGGTGATACTACAAACGGATGCATTTCTGCTGGTCCACATTTCAATG L T P G L H G F H V H Q Y G D T T N G C I S A G P H F N
CCTTACAATAAAACACATGGCGATCCAACCGATGAAATGAGACATGTTGGTGATCTTGGAAATATTGTGGCTGGAGCTGATGGCACTP Y N K T H G D P T D E M R H V G D L G N I V A G A D G T
GCTCACATTGATATTTCTGATAAACATGTACAGTTACTCGGTCCCAATTCAATAATTGGTCGTTCACTTGTTGTGCATGCTGATCAAA H I D I S D K H V Q L L G P N S I I G R S L V V H A D Q
D D L G K G V G D K GD E S L K T G N A DG R V A C G I V
A I S A *
AAAAAAAAA
8821
17550
26279
349108
436137
523166
610195
697199
702
phob
3.0
1.5
-1.51
-3i 0
phi
DI 1h.i1'11 ll\ 1''lRIII 41111lJIIg D .l pIIr 'IIII "'D 11"'l11
21.0 40 80 120 160 200
FIG. 1. Sequences of CuZn SODs from B. pahangi. Nucleotide and derived amino acid sequences from cDNAs encoding the cytoplasmic (A)and EC (B) proteins. The 22-nucleotide spliced leader sequence SLI (20) is underlined at the extreme 5' end of each cDNA, and the sequencesare translated from the first available methionine. Potential sites of N-linked glycosylation are boxed, and the arrows mark possible sites for signalpeptidase cleavage (11). Stop codons are marked with asterisks. A hydropathy plot averaged over 11 residues is shown alongside each amino acidsequence. The cDNA clone C-SOD 1 extends from nucleotides 67 to 364 of the cytoplasmic SOD sequence. Phob, hydrophobic; phil, hydrophilic.
functional roles to 23 invariant residues; 15 were implicated inthe active-site stereochemistry, 4 were implicated in maintain-ing the ,-barrel fold, and 4 were found at the dimer interface(12). All of these residues were conserved in both filarial SODsequences (Fig. 2).
Glycosylation and quaternary association of the parasiteenzymes. The partial cDNA C-SOD 1 (Fig. 1) was subclonedinto pDS78/RBS11 (47) and expressed in E. coli, and theresultant fusion protein, with an estimated mass of 15 kDa, waspartially purified by gel filtration (Fig. 3A). This protein wasused to raise a polyclonal antiserum (designated rabbit anti-C-SOD 1) as outlined in Materials and Methods. The reactivityof the antiserum was tested by Western blotting with an extractof B. malayi adult worms, and preparations of the recombinantprotein were used as positive controls. The bacterially ex-pressed SOD was bound strongly by the antibody, which alsorecognized two proteins with apparent masses of 19 and 29kDa in the parasite extract (Fig. 3B). An additional band ofabout 65 kDa was also seen, although this appeared torepresent background binding of the antibody. As the 29-kDaprotein corresponded closely in mass to the human EC CuZnSOD (28), it appeared most likely that this protein defined thesubunit of the Brugia EC enzyme and the 19-kDa proteindefined the subunit of the cytoplasmic SOD. Cross-reactivity ofthe antiserum against the EC enzyme was to be expectedbecause of the extensive homology between the two proteins.
Given the existence of multiple sites for N-linked glycosyla-tion derived from the EC-SOD cDNA sequence, we assessedthe susceptibility of each protein to digestion with N-glycanase.The results, shown in Fig. 3C, show a three-step digestion ofthe 29-kDa protein with increasing concentrations of enzyme,which is indicative of three sites of N-linked oligosaccharideaddition. Complete digestion with N-glycanase reduced theestimated mass of the EC enzyme to 22 kDa. The 19-kDaprotein showed no alteration in apparent mass on treatmentwith N-glycanase. Although the cDNA sequence allowed theprediction of a single putative site for glycosylation, this siteappears not to be occupied; a similar situation is seen with thehuman cytoplasmic CuZn SOD (41).The quaternary associations of the parasite enzymes were
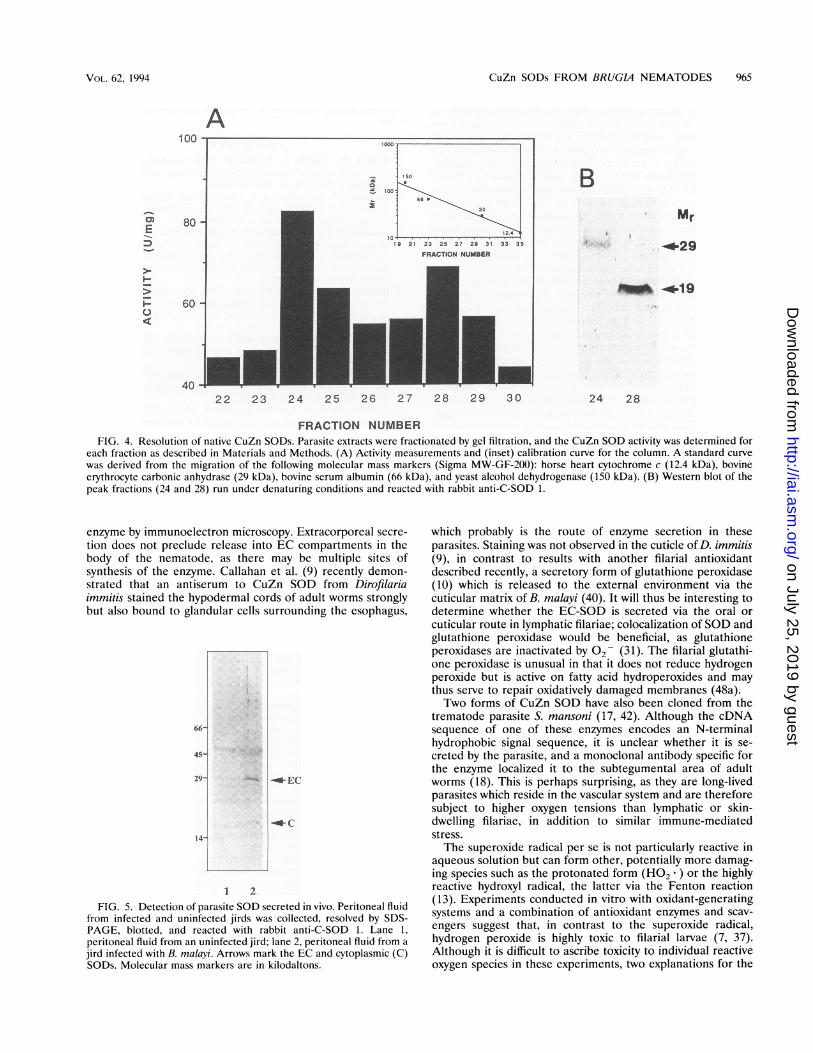
addressed by gel filtration of extracts under nondenaturingconditions. Figure 4A shows that two peaks of enzyme activitywith estimated molecular masses of approximately 60 and 36kDa were observed. When the fractions showing peak activitywere run on SDS-polyacrylamide gels, blotted, and reactedwith the antibody to C-SOD 1, it was evident that the formerpeak corresponded to the EC-SOD and the latter peak corre-sponded to the cytoplasmic SOD, as the 29-kDa subunit andthe 19-kDa subunit were resolved independently (Fig. 4B).
Secretion in vivo. As nematodes are multicellular organisms,the EC enzyme could be destined for extracellular environ-ments within the parasite (e.g., coelomic fluid) rather than
IL .-- 11], hi -11
on July 25, 2019 by guesthttp://iai.asm
.org/D
ownloaded from

964 TANG ET AL.
BE MMIASFAIFLSHIIFITYATSNQRYFKPNMHNNMTIT 37HE MLALLCSCLLLAAGASDAWTGEDSAEPNSDSAEWIR 36
BE IRRTIT-------------------------ETRTA 49BC MS NRk 6BC ATKAV 5HE DMYAKVTEIWQEVMQRRDDDGTLHAACQVQPSATLDA 73
BE DN-GNIN H Q N TTISGE 83BC AVLRGDNI___VSGIIIIIFKQIEpEGPTTISBGE IKGLTP 40HC VG
GQ N EQX ESN G 40
BE L T _ Q APRGLD FFALEIJLPLDANS 108
BE LHGFHVHQYGDTTNGCISAGPHFNPYNKTHG 120BC GLHGF8VHQYGDTTNGCISAGPHFNPYNP;THGGPTDE1 77RC G L H G F
V GD1TAGCTSAGPHFNPLSRKPHGGPEj 77HE FS LAVP_ 140
BE MRBVGDLGNIVAGADGTAHIDISDKHVQLLGPNSIIG 157BC MRHVGDLGNIVAGADGTAHIDISDKHVQLLGPNSIIG 114BC ERBVGDLGB TA EDGVADVIEDSVILBG _BD3IIG 114BE AQnG -F RDGSLWRYRAGLAASLAPiIVG 1 7 5
BE RSLVVHADQDDLGKGVGDKKDESLKTGNAG GGRVACI 194BC RSLVVHADQDDLGKGVGDKKDESLKTGNAG AIRVACGI 151BC RTLVVBEKADDLGKG-G --NEESTETGBAGBSRILACJV 148BE RNAAGEDDLG5G_G--QASVERNG L V 209
BE 199BC AVSAAS 1 5 8BC IGIAQ 1 5 3BE tjjGMCGPGLWERQAREHSERKKRRRESECKAA 2 40
FIG. 2. Amino acid sequence alignment of CuZn SODs. Thesequences are aligned according to the invariant residues defined byGetzoff et al. (12), which are marked with asterisks. Residues arenumbered from the initiator methionine, except for the human cyto-plasmic enzyme, for which the first amino acid of the mature proteinhas been demonstrated to be an acetylated alanine. Residues identicalto those of the Brugia cytoplasmic SOD are boxed. BE, Brugia ECenzyme; BC, Brugia cytoplasmic enzyme; HC, human cytoplasmicenzyme (41); HE, human EC enzyme (16).
extracorporeal secretion. This obviously has implications forinterpretations of biological function, the latter option beingmore consistent with protection against host-derived thanendogenously derived oxidants. We therefore utilized theantiserum to C-SOD 1 to probe for parasite-derived CuZnSOD in peritoneal fluid of Mongolian jirds infected with B.malayi. Following lavage, recovery, filtration, and concentra-tion (see Materials and Methods), fluid was resolved bySDS-PAGE, blotted, and reacted with the antiserum to C-SOD 1. Figure 5 shows that both EC and cytoplasmic CuZnSODs could be detected in peritoneal fluid from infectedanimals, although the EC-SOD was the predominant formobserved, with some indication of breakdown products. No
A
ii_I14 -
I bvo o
reactivity was observed with peritoneal fluid from uninfectedjirds following either direct lavage (Fig. 5) or elicitation ofinflammatory cells with sodium caseinate (data not shown).
DISCUSSIONOn the basis of sequence comparison of CuZn SODs with
the human EC-SOD (the only mammalian EC-SOD whoseprimary structure had been determined at that time), it wasproposed that mammalian EC-SODs evolved from the CuZnSODs after the bacteriocupreins but before the occurrence offungi and plants (16). It is clear that the two filarial CuZnSODs described in this communication are very closely related(88% amino acid identity), show considerable homology tohuman cytoplasmic CuZn SODs (55 to 57% identity), andshow rather less homology to the human EC enzyme (35 to36% identity). The human EC-SOD, however, shows only 38%identity to the human cytoplasmic CuZn SOD. This disparityin primary sequences between the human enzymes is furtherhighlighted by the absence of serological cross-reactivity (29),in contrast to the current observations of strong cross-reactivitybetween the filarial enzymes (Fig. 3). It therefore appears thatthe filarial EC-SOD evolved independently of the mammalianEC-SODs and probably did so relatively recently, presumablydirectly from the cytoplasmic CuZn SOD of the parasite. Asexpected, the B. pahangi enzymes are very similar to the 0.volvulus cytoplasmic CuZn SOD (15), a cDNA coding forwhich was used as a hybridization probe in the present study.The cytoplasmic enzymes from these two species differ at only20 of 158 amino acid residues (i.e., 87% identity). Recently, thesequence for a putative EC-SOD from 0. volvulus has beenentered into the GenBank data base (18a). Perhaps surpris-ingly, the derived amino acid sequence for this protein showsonly 54.5% identity with the B. pahangi EC-SOD.The filarial EC-SOD differs from the human EC enzyme in
that it is dimeric, and this may result from the truncated Nterminus; the N terminus has been proposed to mediateassociation into a tetrameric form for the mammalian EC-SODs (16). Adoption of a tetrameric form is thought to conferstability of the mammalian enzyme in extracellular fluids, andit is possible that the more extensive glycosylation of the filarialEC-SOD compensates for its dimeric form and contributes toa longer half-life. We are currently addressing this issue andinvestigating the localization and route of secretion of the
1 2 3 1 2 3 1 2 3 4
FIG. 3. (A and B) Identification and glycosylation of Brugia CuZn SODs. Lanes 1, E. coli TG2 cell lysate expressing C-SOD 1; lanes 2, purifiedC-SOD 1; lanes 3, lysate of B. malayi adult worms. Panel A was stained with Coomassie blue, and panel B shows the same preparation as in panelA reacted with a rabbit polyclonal antibody to C-SOD 1. The partial SOD expressed in E. coli is indicated by an arrow in panel A, and the parasiteSODs detected with the antibody are indicated by arrows in panel B. (C) Deglycosylation of the parasite EC-SOD. Parasite extracts were treatedwith increasing concentrations of N-glycanase prior to resolution on polyacrylamide gels, blotted, and reacted with the rabbit anti-C-SOD 1. Lane1, no enzyme; lane 2, 10 mU ml- 1; lane 3, 100 mU ml - 1; lane 4, 1 U ml - 1. The fully digested EC-SOD is indicated by an arrow. Molecular massmarkers are in kilodaltons.
INFECT. IMMUN.
66-
45-
29-
I 4
on July 25, 2019 by guesthttp://iai.asm
.org/D
ownloaded from
CuZn SODs FROM BRUGIA NEMATODES 965
aaX
23 25 27 29 31 33 35
FRACTION NUMBER
22 23 24 25 26 27 28 29 30
BMr
!4' 29
4.*19
24 28
FRACTION NUMBERFIG. 4. Resolution of native CuZn SODs. Parasite extracts were fractionated by gel filtration, and the CuZn SOD activity was determined for
each fraction as described in Materials and Methods. (A) Activity measurements and (inset) calibration curve for the column. A standard curve
was derived from the migration of the following molecular mass markers (Sigma MW-GF-200): horse heart cytochrome c (12.4 kDa), bovineerythrocyte carbonic anhydrase (29 kDa), bovine serum albumin (66 kDa), and yeast alcohol dehydrogenase (150 kDa). (B) Western blot of thepeak fractions (24 and 28) run under denaturing conditions and reacted with rabbit anti-C-SOD 1.
enzyme by immunoelectron microscopy. Extracorporeal secre-
tion does not preclude release into EC compartments in thebody of the nematode, as there may be multiple sites ofsynthesis of the enzyme. Callahan et al. (9) recently demon-strated that an antiserum to CuZn SOD from Dirofilariaimmitis stained the hypodermal cords of adult worms stronglybut also bound to glandular cells surrounding the esophagus,
66-
45-
29-
14-
--C
1 2
FIG. 5. Detection of parasite SOD secreted in vivo. Peritoneal fluidfrom infected and uninfected jirds was collected, resolved by SDS-PAGE, blotted, and reacted with rabbit anti-C-SOD 1. Lane 1,
peritoneal fluid from an uninfected jird; lane 2, peritoneal fluid from a
jird infected with B. malayi. Arrows mark the EC and cytoplasmic (C)SODs. Molecular mass markers are in kilodaltons.
which probably is the route of enzyme secretion in theseparasites. Staining was not observed in the cuticle of D. immitis(9), in contrast to results with another filarial antioxidantdescribed recently, a secretory form of glutathione peroxidase(10) which is released to the external environment via thecuticular matrix of B. malayi (40). It will thus be interesting todetermine whether the EC-SOD is secreted via the oral or
cuticular route in lymphatic filariae; colocalization of SOD andglutathione peroxidase would be beneficial, as glutathioneperoxidases are inactivated by o2 (31). The filarial glutathi-one peroxidase is unusual in that it does not reduce hydrogenperoxide but is active on fatty acid hydroperoxides and may
thus serve to repair oxidatively damaged membranes (48a).Two forms of CuZn SOD have also been cloned from the
trematode parasite S. mansoni (17, 42). Although the cDNAsequence of one of these enzymes encodes an N-terminalhydrophobic signal sequence, it is unclear whether it is se-creted by the parasite, and a monoclonal antibody specific forthe enzyme localized it to the subtegumental area of adultworms (18). This is perhaps surprising, as they are long-livedparasites which reside in the vascular system and are thereforesubject to higher oxygen tensions than lymphatic or skin-dwelling filariae, in addition to similar immune-mediatedstress.The superoxide radical per se is not particularly reactive in
aqueous solution but can form other, potentially more damag-ing species such as the protonated form (HO2.) or the highlyreactive hydroxyl radical, the latter via the Fenton reaction(13). Experiments conducted in vitro with oxidant-generatingsystems and a combination of antioxidant enzymes and scav-
engers suggest that, in contrast to the superoxide radical,hydrogen peroxide is highly toxic to filarial larvae (7, 37).Although it is difficult to ascribe toxicity to individual reactiveoxygen species in these experiments, two explanations for the
A100
E
C).4
150
00
30
lo1 ~ ~ 1.4I.80
60
40
VOL. 62, 1994
A...
aI
4.
on July 25, 2019 by guesthttp://iai.asm
.org/D
ownloaded from

966 TANG ET AL.
observations can be proposed. First, as hydrogen peroxide isuncharged, it can freely diffuse across membranes and formhydroxyl radicals with intracellular iron pools (13), whereas thesuperoxide radical appears to require a suitable ion channel or
transport facility to permeate lipid bilayers (23, 48). Second,relatively high levels of SOD have been measured in filarialextracts, but low to negligible levels of catalase and glutathioneperoxidase appear to be present (6-8). This would explain whyexogenous scavengers of hydrogen peroxide are more effectivethan scavengers of superoxide in protecting parasites in theseexperiments, but it raises the question of how filariae metab-olize hydrogen peroxide. Coexpression and colocalization ofSOD and a hydrogen peroxide-metabolizing enzyme reducethe risk of production of hydroxyl radicals. For example, SODis used for the treatment of ischemic reperfusion injury in a
wide variety of tissues, but a bell-shaped dose-response curve
indicating that SOD at high concentrations loses its effective-ness and may enhance the extent of injury has been observed.These observations have recently been attributed to the pro-
duction of hydroxyl radicals due to the increased Fentonreaction resulting from excess hydrogen peroxide (27). Thishas important implications for defense of filarial parasitesagainst antibody-dependent cellular cytotoxicity. The genera-
tion of hypohalous acids from hydrogen peroxide is an impor-tant part of the armory of granulocytes, and a recent study ofeosinophil granule proteins highlighted this pathway as themost potently cytotoxic mechanism against filarial larvae (14).Excess SOD activity would thus be of limited protective valuewithout an efficient means of metabolizing the product.
Recently, a second form of mammalian EC-SOD has beencloned from rat epididymal mRNA (33). The epididymal tissuethus shows a profile of antioxidants which is similar to that offilarial parasites in expressing not only an EC-SOD but also an
unusual selenium-independent secretory glutathione peroxi-dase (34). These enzymes most probably protect spermatozoaduring storage, as they contain a high proportion of membranephospholipids whose fatty acids are polyunsaturated and are
thus highly susceptible to lipid peroxidation. We are currentlyexamining the composition of the epicuticular (surface) mem-
brane of adult B. malayi in order to determine whether itcontains a similar proportion of phospholipids with polyunsat-urated fatty acids.The Brugia EC-SOD lacks the C-terminal extension found in
mammalian counterparts (16, 33). As the positively chargedresidues of this C-terminal extension have been implicated inbinding to sulfated proteoglycans on cell surfaces (1, 19), onewould predict that the filarial EC enzyme would not show thisproperty but would exist free in solution in the EC milieu.Continuous turnover of a soluble EC-SOD might perform one
of two functions: bulk scavenging of 02- or an anti-inflamma-tory activity. Superoxide reacts directly with an undefinedprecursor in plasma to produce a factor that is chemotactic forneutrophils, and the induction of this factor can be inhibited bySOD (35). This is an attractive role for a filarial EC-SOD, asneutrophils are cytotoxic in vitro (37) but are rarely seen
surrounding viable organisms in vivo. The sequences of theputative EC-SODs from S. mansoni (42) and 0. volvulus (18a)also lack this C-terminal extension, and thus this may be an
adaptation common to pathogens which inhabit EC fluids. Weare now examining the role of this enzyme as a potential factorin the promotion of parasite persistence.
ACKNOWLEDGMENTS
This work was supported by the Medical Research Council and theWellcome Trust.
L. Tang and X. Ou should be considered joint first authors of thisarticle.
REFERENCES1. Adachi, T., and S. L. Marklund. 1989. Interactions between
human extracellular superoxide dismutase C and sulfated polysac-charides. J. Biol. Chem. 264:8537-8541.
2. Andersen, P., D. Askgaard, L. Ljungqvist, J. Bennedsen, and I.Heron. 1991. Proteins released from Mycobacterium tuberculosisduring growth. Infect. Immun. 59:1905-1910.
3. Beaman, B. L., C. M. Black, F. Doughty, and L. Beaman. 1985.Role of superoxide dismutase and catalase as determinants ofpathogenicity of Nocardia asteroides: importance in resistance tomicrobicidal activities of human polymorphonuclear neutrophils.Infect. Immun. 47:135-141.
4. Beaman, L., and B. L. Beaman. 1990. Monoclonal antibodiesdemonstrate that superoxide dismutase contributes to protectionof Nocardia asteroides within the intact host. Infect. Immun.58:3122-3128.
5. Beck, B. L., L. B. Tabatabai, and J. E. Mayfield. 1990. A proteinisolated from Brucella abortus is a CuZn superoxide dismutase.Biochemistry 29:372-376.
6. Callahan, H. L., R. K. Crouch, and E. R. James. 1988. Helminthanti-oxidant enzymes: a protective mechanism against host oxi-dants? Parasitol. Today 4:218-225.
7. Callahan, H. L., R. K. Crouch, and E. R. James. 1990. Hydrogenperoxide is the most toxic oxygen species for Onchocerca cervicalismicrofilariae. Parasitology 100:407-415.
8. Callahan, H. L., R. K. Crouch, and E. R. James. 1991. Dirofilariaimmitis superoxide dismutase: purification and characterization.Mol. Biochem. Parasitol. 49:245-252.
9. Callahan, H. L., D. Hazen-Martin, R. K. Crouch, and E. R. James.1993. Immunolocalization of superoxide dismutase in Dirofilariaimmitis adult worms. Infect. Immun. 61:1157-1163.
10. Cookson, E., M. L. Blaxter, and M. E. Selkirk. 1992. Identificationof the major soluble cuticular glycoprotein of lymphatic filarialnematode parasites (gp29) as a secretory homolog of glutathioneperoxidase. Proc. Natl. Acad. Sci. USA 89:5837-5841.
11. Dalbey, R. E., and G. von Heijne. 1992. Signal peptidases inprokaryotes and eukaryotes-a new protease family. Trends Bio-chem. Sci. 17:474-478.
12. Getzoff, E. D., J. A. Tainer, M. M. Stempien, G. I. Bell, and R. A.Hallewell. 1989. Evolution of CuZn superoxide dismutase and theGreek key P-barrel structural motif. Proteins 5:322-336.
13. Halliwell, B., and J. M. C. Gutteridge. 1989. Free radicals inbiology and medicine, 2nd ed. Oxford University Press, Oxford.
14. Hamann, K. J., G. J. Gleich, J. L. Checkel, D. A. Loegering, J. W.McCall, and R. L. Barker. 1990. In vitro killing of Brugia pahangiand Brugia malayi by eosinophil granule proteins. J. Immunol.144:3166-3173.
15. Henkle, K. J., E. Liebau, S. Muller, B. Bergmann, and R. D.Walter. 1991. Characterization and molecular cloning of a Cu/Znsuperoxide dismutase from the human parasite Onchocerca volvu-lus. Infect. Immun. 59:2063-2069.
16. Hjalmarsson, K., S. L. Marklund, A. Engstrom, and T. Edlund.1987. Isolation and sequence of complementary DNA encodinghuman extracellular superoxide dismutase. Proc. Natl. Acad. Sci.USA 84:6340-6344.
17. Hong, Z., P. T. LoVerde, M.-L. Hammarskjold, and D. Rekosh.1992. Schistosoma mansoni: cloning of a complementary DNAencoding a cytosolic Cu/Zn superoxide dismutase and high yieldexpression of the enzymatically active gene product in Escherichiacoli. Exp. Parasitol. 75:308-322.
18. Hong, Z., P. T. LoVerde, A. Thakur, M.-L. Hammarskjold, and D.Rekosh. 1993. Schistosoma mansoni: a Cu/Zn superoxide dis-mutase is glycosylated when expressed in mammalian cells andlocalises to a subtegumental region in adult schistosomes. Exp.Parasitol. 76:101-114.
18a.James, E. R., and D. C. McLean, Jr. Accession number L13778.19. Karlsson, K., and S. L. Marklund. 1987. Heparin-induced release
of extracellular superoxide dismutase to human blood plasma.Biochem. J. 242:55-59.
20. Krause, M., and D. Hirsh. 1987. A trans-spliced leader sequence
INFEC-F. IMMUN.
on July 25, 2019 by guesthttp://iai.asm
.org/D
ownloaded from

CuZn SODs FROM BRUGIA NEMATODES 967
on actin mRNA in C. elegans. Cell 49:753-761.21. Kroll, J. S., P. R. Langford, and B. M. Loynds. 1991. Copper-zinc
superoxide dismutase of Haemophilus infiuenzae and H. parainflu-enzae. J. Bacteriol. 173:7449-7457.
22. Kusunose, E., K. Ichihara, Y. Noda, and M. Kusunose. 1976.Superoxide dismutase from Mycobacterium tuberculosis. J. Bio-chem. 80:1343-1352.
23. Lynch, R. E., and I. Fridovich. 1978. Permeation of the erythrocytestroma by superoxide radical. J. Biol. Chem. 253:4696-4699.
24. Maizels, R. M., M. L. Blaxter, B. R. Robertson, and M. E. Selkirk.1991. Parasite antigens, parasite genes: a laboratory manual formolecular parasitology, p. 224. Cambridge University Press, Cam-bridge.
25. Maizels, R. M., D. A. P. Bundy, M. E. Selkirk, D. F. Smith, andR. M. Anderson. 1993. Immunological modulation and evasion byhelminth parasites in human populations. Nature (London) 365:797-805.
26. Maizels, R. M., W. F. Gregory, G.-E. Kwan-Lim, and M. E.Selkirk. 1989. Filarial surface antigens: the major 29,000 mol.wt.glycoprotein and a novel 17,000-200,000 mol.wt. complex fromadult Brugia malayi parasites. Mol. Biochem. Parasitol. 32:213-227.
27. Mao, G. D., P. D. Thomas, G. D. Lopaschuk, and M. J. Poznansky.1993. Superoxide dismutase (SOD)-catalase conjugates: role ofhydrogen peroxide and the Fenton reaction in SOD toxicity. J.Biol. Chem. 268:416-420.
28. Marklund, S. L. 1982. Human copper-containing superoxide dis-mutase of high molecular weight. Proc. Natl. Acad. Sci. USA79:7634-7638.
29. Marklund, S. L. 1984. Properties of extracellular superoxidedismutase from human lung. Biochem. J. 220:269-272.
30. McCord, J. M., and I. Fridovich. 1969. Superoxide dismutase: anenzymic function of erythrocuprein (hemocuprein). J. Biol. Chem.244:6049-6055.
31. Meister, A., and M. E. Anderson. 1983. Glutathione. Annu. Rev.Biochem. 52:711-760.
32. Paxton, W. A., M. Yazdanbakhsh, A. Kurniawan, F. Partono,R. M. Maizels, and M. E. Selkirk. 1993. Primary structure of andimmunoglobulin E response to the repeat subunit of gplS/400from human lymphatic filarial parasites. Infect. Immun. 61:2827-2833.
33. Perry, A. C. F., R. Jones, and L. Hall. 1993. Isolation andcharacterization of a rat cDNA clone encoding a secreted super-oxide dismutase reveals the epididymis to be a major site of itsexpression. Biochem. J. 293:21-25.
34. Perry, A. C. F., R. Jones, L. S. P. Niang, R. M. Jackson, and L.Hall. 1992. Genetic evidence for an androgen-regulated epididy-mal secretory glutathione peroxidase whose transcript does notcontain a selenocysteine codon. Biochem. J. 285:863-870.
35. Petrone, W. F., D. K. English, K. Wong, and J. M. McCord. 1980.Free radicals and inflammation: superoxide-dependent activationof a neutrophil chemotactic factor in plasma. Proc. Natl. Acad. Sci.USA 77:1159-1163.
36. Rhoads, M. L. 1983. Trichinella spiralis: identification and purifi-cation of superoxide dismutase. Exp. Parasitol. 56:41-54.
37. Rzepczyk, C., and C. J. Bishop. 1984. Immunological and ultra-structural aspects of the cell-mediated killing of Dirofilaria immitismicrofilariae. Parasite Immunol. 6:443-457.
38. Sanger, F., S. Nicklen, and A. R. Coulson. 1977. DNA sequencingwith chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA74:5463-5467.
39. Selkirk, M. E., D. A. Denham, F. Partono, I. Sutanto, and R. M.Maizels. 1986. Molecular characterization of antigens of lymphaticfilarial parasites. Parasitology 91:S15-S38.
40. Selkirk, M. E., W. F. Gregory, R. E. Jenkins, M. Yazdanbakhsh,and R. M. Maizels. 1990. Cuticular localisation and turnover ofthe major surface glycoprotein (gp29) of adult Bruigia malayi. Mol.Biochem. Parasitol. 42:31-44.
41. Sherman, L., N. Dafni, J. Lieman-Hurwitz, and Y. Groner. 1983.Nucleotide sequence and expression of human chromosome 21-encoded superoxide dismutase mRNA. Proc. Natl. Acad. Sci. USA80:5465-5469.
42. Simurda, M. C., H. van Keulen, D. M. Rekosh, and P. T. LoVerde.1988. Schistosoma ntansoni: identification and analysis of anmRNA and a gene encoding superoxide dismutase. Exp. Parasitol.67:73-84.
43. Steinman, H. M. 1982. Copper-zinc superoxide dismutase fromCaulobacter crescentuls CB15. J. Biol. Chem. 257:10283-10293.
44. Steinman, H. M. 1985. Bacteriocuprein superoxide dismutases inpseudomonads. J. Bacteriol. 162:1255-1260.
45. Steinman, H. M. 1987. Bacteriocuprein superoxide dismutase ofPhotobacterium leiognathi. J. Biol. Chem. 262:1882-1887.
46. Steinman, H. M., and B. Ely. 1990. Copper-zinc superoxidedismutase of Caulobacter crescentus: cloning, sequencing, andmapping of the gene and periplasmic location of the enzyme. J.Bacteriol. 172:2901-2910.
47. Stueber, D., H. Matile, and G. Garotta. 1991. System for high levelproduction in E. coli and rapid purification of recombinantproteins: application to epitope mapping, preparation of antibod-ies and structure-function analysis, p. 121-152. In I. Lefkovits andB. Pernis (ed.), Immunological methods, vol. IV. Academic Press,London.
48. Takahashi, M.-A., and K. Asada. 1983. Superoxide anion perme-ability of phospholipid membranes and chloroplast thylakoids.Arch. Biochem. Biophys. 226:558-566.
48a.Tang, L., et al. Unpublished data.49. Tatum, F. M., P. G. Detilleux, J. M. Sacks, and S. M. Hailing.
1992. Construction of Cu-Zn superoxide dismutase deletion mu-tants of Brucella abortus: analysis of survival in vitro in epithelialand phagocytic cells and in vivo in mice. Infect. Immun. 60:2863-2869.
50. Vignais, P. M., A. Terech, C. M. Meyer, and M.-F. Henry. 1982.Isolation and characterisation of a protein with cyanide-sensitivesuperoxide dismutase activity from the prokaryote Paracoccusdenitrificans. Biochim. Biophys. Acta 701:305-317.
VOL. 62? 1994
on July 25, 2019 by guesthttp://iai.asm
.org/D
ownloaded from





























