Embed Size (px)
Citation preview

J Sci Food Agric 1997, 75, 341È351
Factors Limiting the Biodegradation of spUlvaCell-Wall PolysaccharidesChristine Bobin-Dubigeon,1,2 Marc Lahaye,1* Fabienne Guillon,2 Jean-Luc Barry2and Daniel J Gallant21 INRA, Laboratoire de Biochimie et Technologie des Glucides, BP 71627, F-44316 Nantes Cedex 03,France2 INRA, Laboratoire de Technologie Applique� e a la Nutrition, BP 71627, F-44316 Nantes Cedex 03,France
(Received 24 September 1996 ; revised version received 20 January 1997 ; accepted 18 April 1997)
Abstract : The dietary Ðbres of the seaweed Ulva sp (sea-lettuce) consist of water-soluble ulvan, alkali-soluble b(1,4)-D-glucuronan and b(1,4)-D-glucoxylan, and aninsoluble a-cellulose containing xylose residues. They are poorly degraded byhuman colonic bacteria particularly when associated within the intact plant cellwall. In order to better understand this resistance to microbial attack, theirorganisation in the cell-wall has been investigated by light and electron micros-copy after sequential chemical extractions. Their susceptibility to enzymatic deg-radation and their accessibility to bacteria and enzyme were also studied.Microscopic localisation in native and sequentially extracted Ulva sp demon-strated that ulvan is in all the cell-walls of the algae and particularly between thetwo cell layers constituting the thallus. Glucuronan is close to the cytoplasmicmembrane facing the outside of the seaweed and between adjacent cells. Thexylose and glucose containing polysaccharides form packed layers surroundingthe cells. A model of the spatial distribution of the di†erent polysaccharideswithin the algae is proposed.
Ulvan and glucuronan did not limit the xyloglucan and a-cellulose degrada-tion by an endo-xylanase in the whole seaweed and its insoluble dietary Ðbre butthe a-cellulose was not a†ected by a cellulase. The cell-wall of Ulva sp was acces-sible to enzymes but poorly to bacteria as assessed from porosity measurements.These results established that the poor fermentation of sea-lettuce by humancolonic Ñora is primarily due to the ubiquitous presence of the degradation-resistant ulvan in the cell wall of Ulva sp.
J Sci Food Agric 75, 341È351 (1997)No. of Figures : 5. No. of Tables : 3. No. of References : 48
Key words : Ulva, chlorophyta, seaweed, dietary Ðbres, ulvan, biodegradation.
INTRODUCTION
Ulva sp is one of the seaweeds consumed as a vegetablein many countries (Arasaki and Arasaki 1985 ; Inder-gaard and Minsaas 1991) and among its nutritionalbeneÐts is its richness in dietary Ðbre (Lahaye 1991 ;Lahaye and Jegou 1993 ; Lahaye et al 1995). As indigest-ible food polysaccharides can modulate metabolic andbiological processes of the host through their biodegra-
* To whom correspondence should be addressed.Contract grant sponsor : EC.Contract grant number : AIR 1CT 92-0518.
dation products in the colon, we recently studied invitro the fate of this sea-vegetable and its isolatedsoluble and insoluble Ðbres and some of their com-ponent sugars and disaccharide in presence of humanfaecal Ñora (Bobin-Dubigeon et al 1997). This demon-strated that the whole algae and its soluble Ðbre, ulvan,were poorly degraded (32 and D26% of the initial sugarcontent, respectively) whereas the insoluble Ðbre wasmoderately a†ected (D51% disappearance of initialsugar). The refractory behaviour of ulvan to biodegra-dation was related to the absence of colonic bacteriaproducing enzymes able to cleave its peculiar chemicalstructure since its component sugars (glucuronic acid,
3411997 SCI. J Sci Food Agric 0022-5142/97/$17.50. Printed in Great Britain(
342 C Bobin-Dubigeon et al
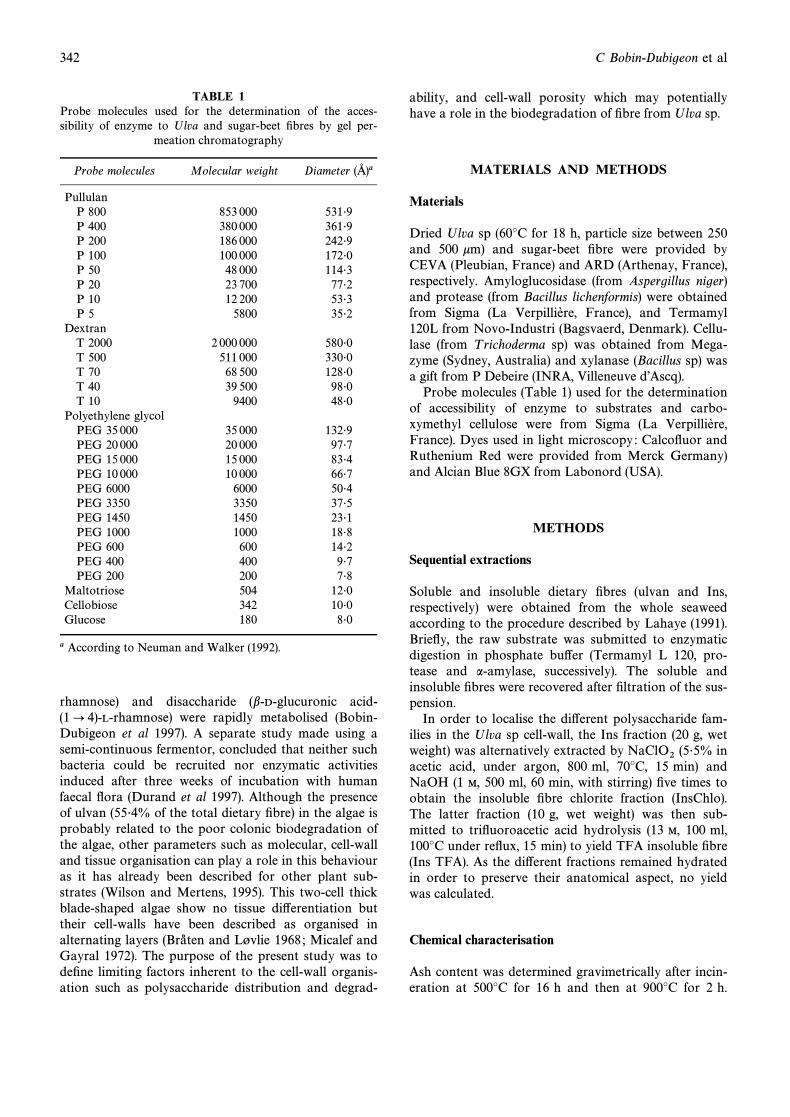
TABLE 1Probe molecules used for the determination of the acces-sibility of enzyme to Ulva and sugar-beet Ðbres by gel per-
meation chromatography
Probe molecules Molecular weight Diameter (Ó)a
PullulanP 800 853 000 531É9P 400 380 000 361É9P 200 186 000 242É9P 100 100 000 172É0P 50 48 000 114É3P 20 23 700 77É2P 10 12 200 53É3P 5 5800 35É2
DextranT 2000 2 000 000 580É0T 500 511 000 330É0T 70 68 500 128É0T 40 39 500 98É0T 10 9400 48É0
Polyethylene glycolPEG 35 000 35 000 132É9PEG 20 000 20 000 97É7PEG 15 000 15 000 83É4PEG 10 000 10 000 66É7PEG 6000 6000 50É4PEG 3350 3350 37É5PEG 1450 1450 23É1PEG 1000 1000 18É8PEG 600 600 14É2PEG 400 400 9É7PEG 200 200 7É8
Maltotriose 504 12É0Cellobiose 342 10É0Glucose 180 8É0
a According to Neuman and Walker (1992).
rhamnose) and disaccharide (b-D-glucuronic acid-(1 ] 4)-L-rhamnose) were rapidly metabolised (Bobin-Dubigeon et al 1997). A separate study made using asemi-continuous fermentor, concluded that neither suchbacteria could be recruited nor enzymatic activitiesinduced after three weeks of incubation with humanfaecal Ñora (Durand et al 1997). Although the presenceof ulvan (55É4% of the total dietary Ðbre) in the algae isprobably related to the poor colonic biodegradation ofthe algae, other parameters such as molecular, cell-walland tissue organisation can play a role in this behaviouras it has already been described for other plant sub-strates (Wilson and Mertens, 1995). This two-cell thickblade-shaped algae show no tissue di†erentiation buttheir cell-walls have been described as organised inalternating layers (Bra- ten and 1968 ; Micalef andLÔvlieGayral 1972). The purpose of the present study was todeÐne limiting factors inherent to the cell-wall organis-ation such as polysaccharide distribution and degrad-
ability, and cell-wall porosity which may potentiallyhave a role in the biodegradation of Ðbre from Ulva sp.
MATERIALS AND METHODS
Materials
Dried Ulva sp (60¡C for 18 h, particle size between 250and 500 km) and sugar-beet Ðbre were provided byCEVA (Pleubian, France) and ARD (Arthenay, France),respectively. Amyloglucosidase (from Aspergillus niger)and protease (from Bacillus lichenformis) were obtainedfrom Sigma (La Verpilliere, France), and Termamyl120L from Novo-Industri (Bagsvaerd, Denmark). Cellu-lase (from T richoderma sp) was obtained from Mega-zyme (Sydney, Australia) and xylanase (Bacillus sp) wasa gift from P Debeire (INRA, Villeneuve dÏAscq).
Probe molecules (Table 1) used for the determinationof accessibility of enzyme to substrates and carbo-xymethyl cellulose were from Sigma (La Verpilliere,France). Dyes used in light microscopy : CalcoÑuor andRuthenium Red were provided from Merck Germany)and Alcian Blue 8GX from Labonord (USA).
METHODS
Sequential extractions
Soluble and insoluble dietary Ðbres (ulvan and Ins,respectively) were obtained from the whole seaweedaccording to the procedure described by Lahaye (1991).BrieÑy, the raw substrate was submitted to enzymaticdigestion in phosphate bu†er (Termamyl L 120, pro-tease and a-amylase, successively). The soluble andinsoluble Ðbres were recovered after Ðltration of the sus-pension.
In order to localise the di†erent polysaccharide fam-ilies in the Ulva sp cell-wall, the Ins fraction (20 g, wetweight) was alternatively extracted by (5É5% inNaClO2acetic acid, under argon, 800 ml, 70¡C, 15 min) andNaOH (1 M, 500 ml, 60 min, with stirring) Ðve times toobtain the insoluble Ðbre chlorite fraction (InsChlo).The latter fraction (10 g, wet weight) was then sub-mitted to triÑuoroacetic acid hydrolysis (13 M, 100 ml,100¡C under reÑux, 15 min) to yield TFA insoluble Ðbre(Ins TFA). As the di†erent fractions remained hydratedin order to preserve their anatomical aspect, no yieldwas calculated.
Chemical characterisation
Ash content was determined gravimetrically after incin-eration at 500¡C for 16 h and then at 900¡C for 2 h.

Factors limiting the biodegradation of Ulva sp cell-wall polysaccharides 343
Protein content was determined by the micro-Kjedahlmethod or, for ulvan, by colorimetry (Lowry et al 1951).The di†erent fractions were hydrolysed (Hoebler et al1989) and the neutral sugar content was determined byGC following the procedure of Blakeney et al (1983).Uronic acid content was quantiÐed according to Thi-bault (1979), and sulphate content according to Lahayeand Axelos (1993). Enzymatic degradation of Ulva spand its insoluble Ðbre by cellulase and xylanase was fol-lowed by measuring the increase in reducing sugar inthe supernatant of the reaction mixture (Nelson 1944).
Microscopic observations
L ight microscopyFrozen samples ([20¡C) were cut using the cryotomeMicrom HM 500 OM and sections (5È10 km thick)were dried at 60¡C overnight then stained as follows.
CalcoÑuorSections were stained with 0É01% CalcoÑuor in waterfor 1 min according to the procedure described byWood et al (1983). Excess stain was removed by a 1 minwash in running deionised water.
Alcian Blue (AB)This stain was used according to the proceduredescribed by Parker and Diboll (1966). For the observ-ation of both sulphate and carboxylic groups, sectionswere treated with AB (0É1% AB in acetic acid 3%) atpH 2É4 for 20 min. The sections were then washed threetimes with deionised water for 1 min each. For thedetection of sulphate groups only, the sections werestained for 10 min with AB at pH 0É5 (0É1% AB in 0É5 N
HCl) followed by three deionised water washes asabove. AB staining was also performed on gel of ulvan(3% added with 50 mM and 400 mMH3BO3 CaCl2 ;Lahaye et al 1996).
Ruthenium Red (RR)Sections were stained with RR in water for0É025&
90 min according to the procedure of Luft (1971).Stained sections were observed on a Leica DMRB
microscope equipped with the appropriate ÑuorescenceÐlters (UV excitation 340È380 nm, emission [430 nm)for calcoÑuor Ñuorescence observation.
T ransmission electron microscopyFixation of pre-hydrated samples was performed usingglutaraldehyde (Bra- ten and 1968) followed byLÔvlie,post-Ðxation in and embedding in Epon. The pol-OsO4ymerisation was done at 40¡C for 24 h then 60¡C for 4days. The sections (80 nm) were achieved with Ultra-microtome Microm RMC MT-7000.
Section contrast was enhanced with uranyl acetateand bismuth subnitrate (Riva 1974). Sections collectedon copper grids (200 mesh) were stained by Ñoating on
a drop of uranyl acetate (2É8%) in methanol (50%) for30 min at 48¡C. After extensive washes in deionizedwater (5 min, 6 times), the grids were treated withbismuth subnitrate (0É04%) in sodium tartrate (0É08%)for 3 min before extensive washing with deionisedwater. The periodic acidÈthiosemicarbazideÈbismuth(PATBi) staining method was the PATAg reactionaccording to Thie� ry (1967) modiÐed at the laboratoryby replacing silver proteinate by bismuth subnitrate.Sections were collected on gold grids and Ñoated on adrop of periodic acid solution in deionised water (1%,30 min). After extensive washes with deionised water(5 min, 6 times), the grids were treated with thiosemi-carbazide (0É2%) in aqueous acetic acid solution (20%)for 24 h at room temperature. The material was rinsedin a series of acetic acid solutions (20, 15, 10, 5 and2É5%), for 5 min each. After extensive washes in deion-ised water, the grids were treated with bismuth sub-nitrate as described above.
Uranyl acetate/bismuth subnitrate contrast andPATBi were performed on raw substrates, on fermenta-tion residues of Ulva sp and Ins, and on the enzymaticresidues of Ulva sp and Ins after xylanase degradation.
In situ degradations of cell-wall polysaccharides ofUlva sp of cellulase and endo-b-1,4-xylanase werestudied according to the procedure of Joseleau and Ruel(1985), here again replacing silver proteinate by bismuthsubnitrate (ETBi).
The fractions were observed with a JEOL transmis-sion electron microscope operating at 80 keV.
Enzymatic degradation
The cell-wall polysaccharide degradation of Ulva sp andits insoluble dietary Ðbre (10 mg ml~1 in water, 10 ml)by an endo-b-1,4-xylanase and an endo-b-1,4-cellulase(100 nkat ml~1 in water, 10 ml) was followed for 5, 10,20, 30 and 60 min by the release of reducing sugars inthe supernatant (centrifugation : 4000 ] g for 5 min).
In a second procedure, at time 0, an aliquot of xyla-nase (10 ml, 20 nkat ml~1 in water) was added to Ulvasp suspension (10 ml, 10 mg ml~1 in water). After 3, 6,18 and 24 h of incubation, the above suspension wascentrifuged (4000] g for 5 min) in order to remove theextracted sugars. The pellet was re-suspended in water(10 ml) and xylanase (10 ml, 20 nkat ml~1) and after afurther 28 h, the medium was centrifuged (4000] g for5 min). The supernatant was removed and the pelletwas kept for sugar content analysis and transmissionelectron microscopy (TEM) observations. Blanks wereperformed without enzyme.
Inhibition of cellulase activity by ulvan was expressas the percentage decrease of reducing power measuredbetween an aqueous solution of carboxymethyl cellulose(1%, w/v) with or without ulvan (1%, w/v) after 5 to30 min of incubation at 25¡C and 40¡C.
344 C Bobin-Dubigeon et al
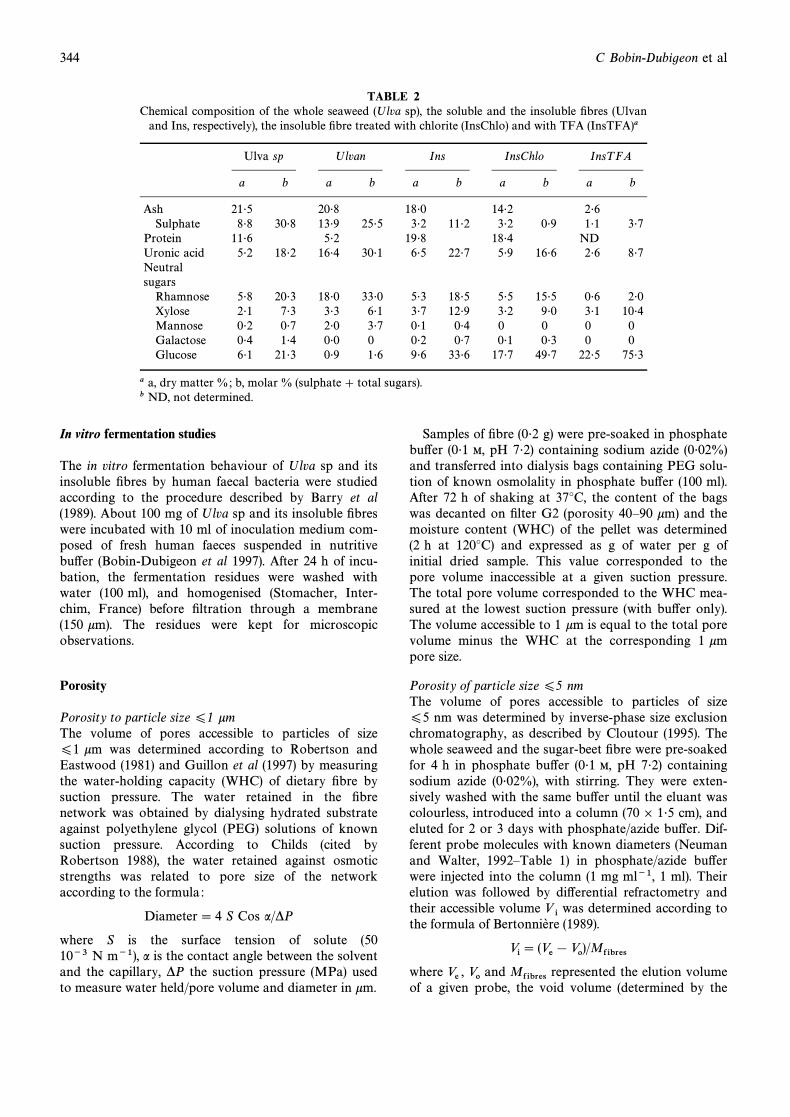
TABLE 2Chemical composition of the whole seaweed (Ulva sp), the soluble and the insoluble Ðbres (Ulvan
and Ins, respectively), the insoluble Ðbre treated with chlorite (InsChlo) and with TFA (InsTFA)a
Ulva sp Ulvan Ins InsChlo InsT FA
a b a b a b a b a b
Ash 21É5 20É8 18É0 14É2 2É6Sulphate 8É8 30É8 13É9 25É5 3É2 11É2 3É2 0É9 1É1 3É7
Protein 11É6 5É2 19É8 18É4 NDUronic acid 5É2 18É2 16É4 30É1 6É5 22É7 5É9 16É6 2É6 8É7Neutralsugars
Rhamnose 5É8 20É3 18É0 33É0 5É3 18É5 5É5 15É5 0É6 2É0Xylose 2É1 7É3 3É3 6É1 3É7 12É9 3É2 9É0 3É1 10É4Mannose 0É2 0É7 2É0 3É7 0É1 0É4 0 0 0 0Galactose 0É4 1É4 0É0 0 0É2 0É7 0É1 0É3 0 0Glucose 6É1 21É3 0É9 1É6 9É6 33É6 17É7 49É7 22É5 75É3
a a, dry matter %; b, molar % (sulphate ] total sugars).b ND, not determined.
In vitro fermentation studies
The in vitro fermentation behaviour of Ulva sp and itsinsoluble Ðbres by human faecal bacteria were studiedaccording to the procedure described by Barry et al(1989). About 100 mg of Ulva sp and its insoluble Ðbreswere incubated with 10 ml of inoculation medium com-posed of fresh human faeces suspended in nutritivebu†er (Bobin-Dubigeon et al 1997). After 24 h of incu-bation, the fermentation residues were washed withwater (100 ml), and homogenised (Stomacher, Inter-chim, France) before Ðltration through a membrane(150 km). The residues were kept for microscopicobservations.
Porosity
Porosity to particle size O1 kmThe volume of pores accessible to particles of sizeO1 km was determined according to Robertson andEastwood (1981) and Guillon et al (1997) by measuringthe water-holding capacity (WHC) of dietary Ðbre bysuction pressure. The water retained in the Ðbrenetwork was obtained by dialysing hydrated substrateagainst polyethylene glycol (PEG) solutions of knownsuction pressure. According to Childs (cited byRobertson 1988), the water retained against osmoticstrengths was related to pore size of the networkaccording to the formula :
Diameter\ 4 S Cos a/*P
where S is the surface tension of solute (5010~3 N m~1), a is the contact angle between the solventand the capillary, *P the suction pressure (MPa) usedto measure water held/pore volume and diameter in km.
Samples of Ðbre (0É2 g) were pre-soaked in phosphatebu†er (0É1 M, pH 7É2) containing sodium azide (0É02%)and transferred into dialysis bags containing PEG solu-tion of known osmolality in phosphate bu†er (100 ml).After 72 h of shaking at 37¡C, the content of the bagswas decanted on Ðlter G2 (porosity 40È90 km) and themoisture content (WHC) of the pellet was determined(2 h at 120¡C) and expressed as g of water per g ofinitial dried sample. This value corresponded to thepore volume inaccessible at a given suction pressure.The total pore volume corresponded to the WHC mea-sured at the lowest suction pressure (with bu†er only).The volume accessible to 1 km is equal to the total porevolume minus the WHC at the corresponding 1 kmpore size.
Porosity of particle size O5 nmThe volume of pores accessible to particles of sizeO5 nm was determined by inverse-phase size exclusionchromatography, as described by Cloutour (1995). Thewhole seaweed and the sugar-beet Ðbre were pre-soakedfor 4 h in phosphate bu†er (0É1 M, pH 7É2) containingsodium azide (0É02%), with stirring. They were exten-sively washed with the same bu†er until the eluant wascolourless, introduced into a column (70] 1É5 cm), andeluted for 2 or 3 days with phosphate/azide bu†er. Dif-ferent probe molecules with known diameters (Neumanand Walter, 1992ÈTable 1) in phosphate/azide bu†erwere injected into the column (1 mg ml~1, 1 ml). Theirelution was followed by di†erential refractometry andtheir accessible volume was determined according toV ithe formula of Bertonniere (1989).
Vi \ (Ve [ Vo)/Mfibreswhere and represented the elution volumeVe , Vo Mfibresof a given probe, the void volume (determined by the

Factors limiting the biodegradation of Ulva sp cell-wall polysaccharides 345
Fig 1. Schematic representation of a cross-section of Ulva spthallus showing the di†erent cell wall zones discussed in the
text.
elution volume of Pullulan 800), and the dry weight ofthe Ðbres, respectively.
RESULTS
Chemical composition of Ulva sp fractions (Table 2)
The main sugars in Ulva sp were glucose, rhamnose,xylose and uronic acids. The algae also contained 8É8%of sulphate on its dry weight basis. Ulvan, the solubleÐbre, was composed mainly of rhamnose, uronic acidand sulphate, whereas the insoluble Ðbre fraction (Ins)was richer in glucose, xylose and uronic acids andpoorer in the other components. The molar proportionsof rhamnose, xylose uronic acid and sulphate decreasedand that of glucose increased after alkali and sodiumchlorite treatments of the insoluble Ðbre (InsChlo). After
hot acid treatment (InsTFA), the main residual sugarswere glucose, xylose and uronic acids. The materialunaccounted for in these analyses may be ascribed tothe difficulty in quantitatively hydrolysing uronic acidÈrhamnose linkages (BeMiller 1967), to the presence oflipids associated with membranes and/or to particularlyacid resistant polymers, especially in the TFA treatedÐbres for which the protein content was not measured.
Microscopic observations
In order to facilitate the description of Ulva sp cell wallorganisation, the terminology used in the text is pre-sented on a schematic representation of a cross-sectionof the algae (Fig 1). Three cell-wall zones are distin-guished according to their localisation : median, lateraland external cell-walls and a cuticle covering both sidesof the algae.
Light microscopic observations of the di†erent Ulvasp and insoluble Ðbre fractions conÐrmed the bi-seriatenature of the algae (Fig 2(A)). Extraction of ulvanyielded insoluble Ðbre (Ins) apparently free of intracellu-lar material and of organisation similar to that of thewhole algae except that the two cell layers were eitherstarting to dissociate, looking, in cross-section, like anopening zipper, or existed as separate strands (Fig 2(B)).A similar aspect was observed after alkali and sodiumchlorite treatment of the insoluble Ðbre (InsChlo).However, after acid treatment (InsTFA), only separatestrands of external cell-walls associated with intact
Fig 2. Light microscopy. (A), (B), (C) : Cross-section of Ulva sp, Ins and InsTFA, respectively, stained with Alcian Blue 8GX at pH2É4. (D) : Primary Ñuorescence of a cross section of Ulva sp. (E) : Cross-section of Ulva sp stained with calcoÑuor (barÈ50 km, cu :
cuticle).
346 C Bobin-Dubigeon et al
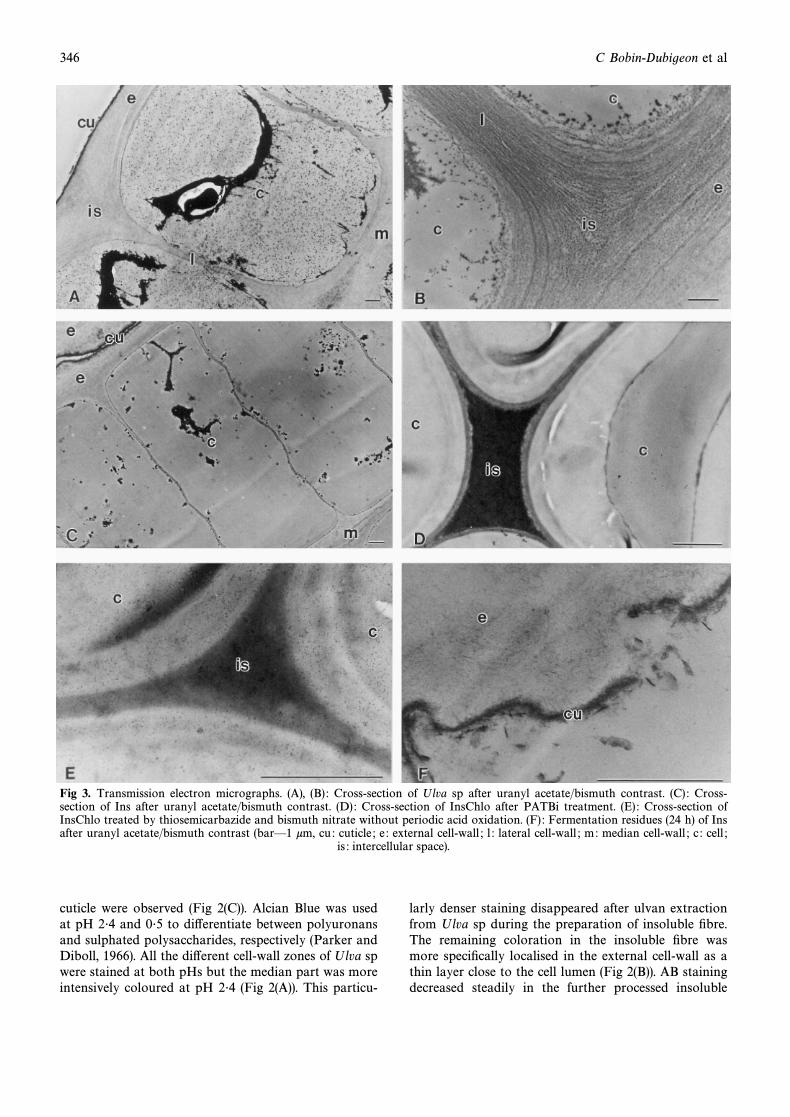
Fig 3. Transmission electron micrographs. (A), (B) : Cross-section of Ulva sp after uranyl acetate/bismuth contrast. (C) : Cross-section of Ins after uranyl acetate/bismuth contrast. (D) : Cross-section of InsChlo after PATBi treatment. (E) : Cross-section ofInsChlo treated by thiosemicarbazide and bismuth nitrate without periodic acid oxidation. (F) : Fermentation residues (24 h) of Insafter uranyl acetate/bismuth contrast (barÈ1 km, cu : cuticle ; e : external cell-wall ; l : lateral cell-wall ; m: median cell-wall ; c : cell ;
is : intercellular space).
cuticle were observed (Fig 2(C)). Alcian Blue was usedat pH 2É4 and 0É5 to di†erentiate between polyuronansand sulphated polysaccharides, respectively (Parker andDiboll, 1966). All the di†erent cell-wall zones of Ulva spwere stained at both pHs but the median part was moreintensively coloured at pH 2É4 (Fig 2(A)). This particu-
larly denser staining disappeared after ulvan extractionfrom Ulva sp during the preparation of insoluble Ðbre.The remaining coloration in the insoluble Ðbre wasmore speciÐcally localised in the external cell-wall as athin layer close to the cell lumen (Fig 2(B)). AB stainingdecreased steadily in the further processed insoluble
Factors limiting the biodegradation of Ulva sp cell-wall polysaccharides 347
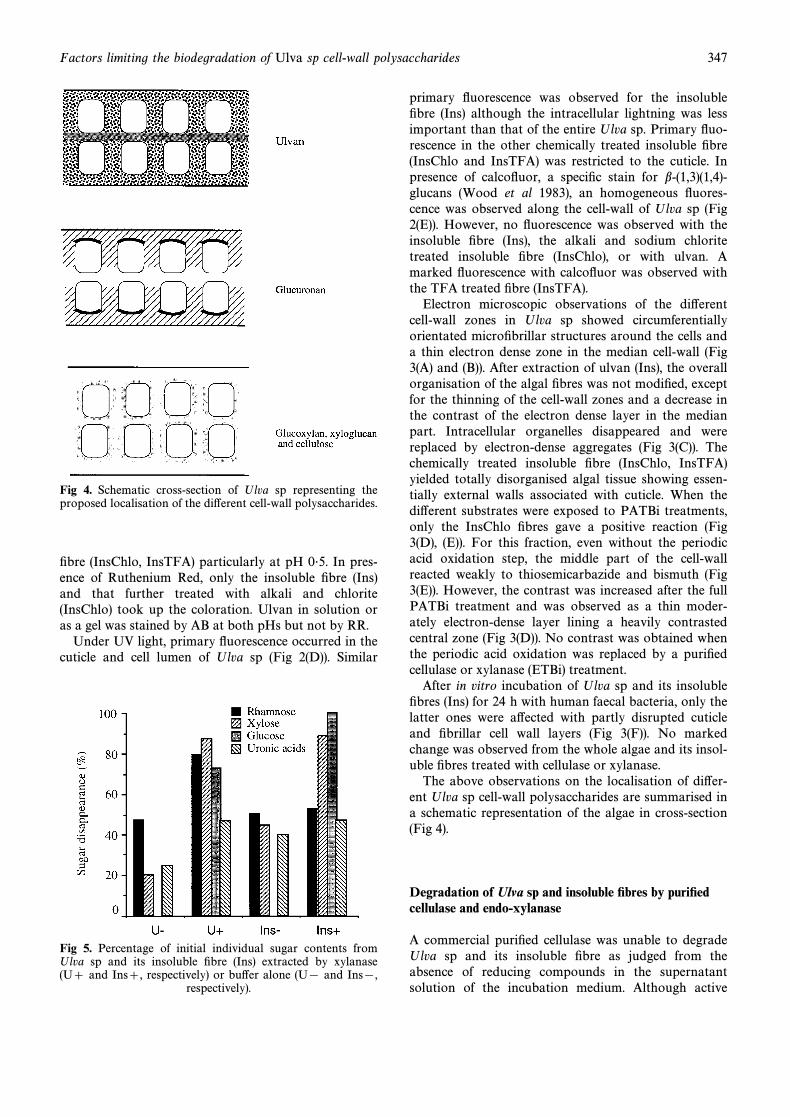
Fig 4. Schematic cross-section of Ulva sp representing theproposed localisation of the di†erent cell-wall polysaccharides.
Ðbre (InsChlo, InsTFA) particularly at pH 0É5. In pres-ence of Ruthenium Red, only the insoluble Ðbre (Ins)and that further treated with alkali and chlorite(InsChlo) took up the coloration. Ulvan in solution oras a gel was stained by AB at both pHs but not by RR.
Under UV light, primary Ñuorescence occurred in thecuticle and cell lumen of Ulva sp (Fig 2(D)). Similar
Fig 5. Percentage of initial individual sugar contents fromUlva sp and its insoluble Ðbre (Ins) extracted by xylanase(U] and Ins], respectively) or bu†er alone (U[ and Ins[,
respectively).
primary Ñuorescence was observed for the insolubleÐbre (Ins) although the intracellular lightning was lessimportant than that of the entire Ulva sp. Primary Ñuo-rescence in the other chemically treated insoluble Ðbre(InsChlo and InsTFA) was restricted to the cuticle. Inpresence of calcoÑuor, a speciÐc stain for b-(1,3)(1,4)-glucans (Wood et al 1983), an homogeneous Ñuores-cence was observed along the cell-wall of Ulva sp (Fig2(E)). However, no Ñuorescence was observed with theinsoluble Ðbre (Ins), the alkali and sodium chloritetreated insoluble Ðbre (InsChlo), or with ulvan. Amarked Ñuorescence with calcoÑuor was observed withthe TFA treated Ðbre (InsTFA).
Electron microscopic observations of the di†erentcell-wall zones in Ulva sp showed circumferentiallyorientated microÐbrillar structures around the cells anda thin electron dense zone in the median cell-wall (Fig3(A) and (B)). After extraction of ulvan (Ins), the overallorganisation of the algal Ðbres was not modiÐed, exceptfor the thinning of the cell-wall zones and a decrease inthe contrast of the electron dense layer in the medianpart. Intracellular organelles disappeared and werereplaced by electron-dense aggregates (Fig 3(C)). Thechemically treated insoluble Ðbre (InsChlo, InsTFA)yielded totally disorganised algal tissue showing essen-tially external walls associated with cuticle. When thedi†erent substrates were exposed to PATBi treatments,only the InsChlo Ðbres gave a positive reaction (Fig3(D), (E)). For this fraction, even without the periodicacid oxidation step, the middle part of the cell-wallreacted weakly to thiosemicarbazide and bismuth (Fig3(E)). However, the contrast was increased after the fullPATBi treatment and was observed as a thin moder-ately electron-dense layer lining a heavily contrastedcentral zone (Fig 3(D)). No contrast was obtained whenthe periodic acid oxidation was replaced by a puriÐedcellulase or xylanase (ETBi) treatment.
After in vitro incubation of Ulva sp and its insolubleÐbres (Ins) for 24 h with human faecal bacteria, only thelatter ones were a†ected with partly disrupted cuticleand Ðbrillar cell wall layers (Fig 3(F)). No markedchange was observed from the whole algae and its insol-uble Ðbres treated with cellulase or xylanase.
The above observations on the localisation of di†er-ent Ulva sp cell-wall polysaccharides are summarised ina schematic representation of the algae in cross-section(Fig 4).
Degradation of Ulva sp and insoluble Ðbres by puriÐedcellulase and endo-xylanase
A commercial puriÐed cellulase was unable to degradeUlva sp and its insoluble Ðbre as judged from theabsence of reducing compounds in the supernatantsolution of the incubation medium. Although active

348 C Bobin-Dubigeon et al
TABLE 3Total volume of pore P1 km (macroporosity) and P5 nm
(microporosity) in Ulva sp and its insoluble Ðbre
Ulva Insolublesp Ðbres
MacroporosityTotal volume (ml g~1) 12É3 18É9Pore volume P1 km (ml g~1) 3É4 9É2% Pore volume P1 km 28É0 48É8
MicroporosityPore volume P5 nm (ml g~1) 8É9%Pore volume P5 km 72É3 NDa
a ND, not determined.
against carboxymethyl cellulose, the cellulase activitywas partially inhibited when ulvan was present in thereaction medium and the extent of inhibition decreasedwith increasing temperature (37É5 ^ 2É5% at 25¡C and19É0 ^ 2É9% at 40¡C) and this inhibition was constantbetween 5 and 30 min of incubation. In contrast, about69% of the initial sugar content of Ulva sp (expressed asdry weight of total sugar content) and its insolubleÐbres were extracted by a puriÐed endo-xylanase (Fig5). Although rhamnose, xylose and uronic acids wereextracted by bu†er alone (47É7, 20É0 and 24É0% of theinitial sugar content, respectively), the main sugarsa†ected by the enzymatic treatment were glucose andxylose since most of them were removed from the Ðbresby the action of the xylanase (Fig 5).
Porosity
The total volume of pores and those of mean diameteraccessible to particles O1 km (macroporosity) andO5 nm (microporosity) are given in Table 3. The insol-uble Ðbres showed the highest total and macroporevolumes among the di†erent substrates studied. Due totechnical difficulties with the insoluble Ðbre, the micro-porosity was only determined for the whole algae.
DISCUSSION
Ulva sp (sea-lettuce) consists of two layers of palisade-like cells (Micalef and Gayral 1972 ; Fig 2) which shapeand cell-wall thickness change according to their local-isation within the thallus, with the algal species andwith the ecophysiological conditions of the plant(Brynhi 1978 ; Phillips 1988 ; Kulhenkamp and Le
Bozec, pers comm). The cell-wall polysaccharidesconsist of water-soluble xyloglucuronorhamnan sul-phate (ulvan), alkali-soluble glucoxylan and glucuronanand amorphous cellulose (a-cellulose) containing xyloseresidues (Lahaye et al 1994, 1996). According to theknown solubility behaviours of these di†erent poly-saccharides and the chemical composition of the Ðbrefractions isolated in the present study (Table 2), ulvancorresponded to the soluble dietary Ðbre whereas thealkali-soluble and insoluble polysaccharides composedmost of the water-insoluble Ðbre. The distribution ofthese polysaccharides within the algal cell-wall wasstudied by relating chemical compositions with micro-scopic observations of Ulva sp and insoluble Ðbre frac-tions obtained after sequential extractions.
Light microscopy conÐrmed that ulvan which wasstained by Alcian Blue at pH 0É5 due to its sulphategroups, was in the three cell-wall zones depicted in Fig2 AB (Parker and Diboll, 1966) and particularly in themedian area. The dense staining in this area disap-peared in the insoluble Ðbre (Ins) which also containedlower content in ulvan as judged from the decrease inrhamnose and sulphate (chemical markers of ulvan)compared to that of the whole algae (Table 2). In con-trast to Micalef and Gayral (1972) observations, ulvanlocalisation in Ulva sp by Ruthenium Red stainingfailed and, although the polysaccharide containinguronic acids are known to react with the stain (Luft1971), no coloration was observed with ulvan alone.However, staining was observed with the insoluble Ðbre(Ins) and those treated by alkali and chlorite (InsChlo).Of the formers, the thin, densely stained layer observedin the external wall close to the cellular lumen was par-ticularly strongly coloured when AB was used at pH 2É4and not at pH 0É5, demonstrating that uronic acidswere involved in the staining reactions. Since the ulvancontent was low in both Ins and InsChlo Ðbres, thestainings observed were attributed to the b-(1,4)-D-glu-curonan. This polysaccharide would mainly be distrib-uted in the external and lateral walls and most likelywas closely associated with the “ÐbrillarÏ part of the cell-wall constituted by the glucose and xylose containingpolysaccharides (Fig 4).
The homogeneous CalcoÑuor Ñuorescence in the cell-wall of the whole Ulva sp was not due to an even dis-tribution of cellulosic material since it disappeared inIns after extraction of ulvan. Instead, the stain wasprobably associated with the glucose and protein-richcompounds that formed CalcoÑuor Ñuorescent particleson re-dissolution of a freeze-dried clear ulvan solution(data not shown). CalcoÑuor positive staining wasobserved only after severe de-encrustation of the cellu-losic material by hot TFA suggesting that the celluloseÐbrils were deeply embedded within a matrix which pre-vented the access of the stain to the glucans.
Electron microscopic observations of Ulva sp and ofits insoluble Ðbres conÐrmed the multilayer array of the

Factors limiting the biodegradation of Ulva sp cell-wall polysaccharides 349
cell-wall of ulvales (Fig 3(A) and (B) ; Bra- ten and LÔvlie1968 ; and Bra- ten 1968 ; Micalef and GayralLÔvlie1972 ; Pierre et al 1974 ; Callow and Evans 1977). Thelocalisation of ulvan and glucuronan at the ultrastruc-tural level was not possible due to the lack of speciÐcstains. In particular, the periodic acid oxidation ofsugars containing contiguous hydroxyl groups andstaining of the resulting aldehyde groups by bismuthatoms (PATBi) was not applicable because of theknown resistance of these acidic polysaccharides to suchtreatment (Percival and McDowell 1967 ; Painter 1983).In contrast, the glucoxylan and a-cellulose should havebeen detected by the PATBi and ETBi stainingmethods. The absence of staining in Ins, InsTFA and inthe whole Ulva sp conÐrmed the results obtained withthe closely related algae E intestinalis (Callow andEvans, 1977) and may be related to the presence of pro-teins and/or glucuronan protecting the polysaccharidesfrom oxidation. Only when Ins was exposed to an oxi-dative agent (sodium chlorite), a positivethiosemicarbazide-bismuth staining was observed (Fig3(E)). The increase in contrast observed after applica-tion of the full PATBi treatment (Fig 3(D)) indicatedthat the embedding resin (Epon) did not limit the di†u-sion of the reagent through the thin section. Furtherevidence for this di†usion was given by the PATBistaining of starch granules in sections from the wholeUlva sp (data not shown). However, the embeddingseverely a†ected the tissue organisation of InsChlo Ðbreas the cell strands observed under light or UV micros-copy appeared totally collapsed under EM. Therefore,the localisation of glucoxylan and a-cellulose close tothe cell lumen and spanning through the entire cell-wallmay be biased by artefacts due to the embedding treat-ment. Instead, these polysaccharides may form packedlayers surrounding the cell lumen with the peripheralones being more amenable to oxidation and thus stain-ing. The disappearance of the PATBi staining in theInsTFA Ðbre can be explained by the degradation ofthe oxidised products by the acid treatment in a reac-tion similar to that of a Smith degradation (Pazur 1986)and by the protection of the remaining oxidisable poly-saccharides by encrusting polymers.
Considering the low or moderate fermentations ofulvan, Ulva sp and its insoluble Ðbre by human faecalbacteria (Bobin-Dubigeon et al 1997), ulvan and glucu-ronan are most likely responsible for the resistance tofeacal bacteria and enzymatic degradation of Ulva spand its insoluble Ðbre observed by electron microscopy.Because the in vitro fermentation studies showed thatglucose and xylose were the sugars most extensivelyused by the bacteria (Bobin-Dubigeon et al 1997), thedegradation of polysaccharides containing such sugarsin Ulva sp and its insoluble Ðbre was studied using apuriÐed cellulase and xylanase. The partial inhibition ofthe cellulase activity by ulvan demonstrated that thepresence of associated acidic polysaccharides can modu-
late the enzymatic degradation of Ulva sp Ðbre.However, further work on the cellulase kinetics arerequired to explain this inhibition. The particular a-cellulose structure together with the presence of glu-coxylan in the algae and its insoluble Ðbre wouldexplain the important release of glucose and xyloseduring the incubation of these substrates with the xyla-nase (Fig 5). The complete (or nearly complete) releaseof the initial xylose and glucose contents from thesesubstrate by the enzyme was in good agreement withthe availability of pores of mean diameters P5 nmallowing for the di†usion of enzymes through the wholealgae (Table 3). These results conÐrmed that the lack ofappropriate enzymes (particularly ulvanolytic activities)was one of the limiting factor to Ulva sp biodegradation(Bobin-Dubigeon et al 1997) rather than a barrier totheir di†usion into the substrate. However, dietary Ðbredegradation within the large bowel is essentially due toenzymes associated with bacterial cell wall (Salyers et al1977). Therefore, another factor determining the rateand extent of degradation of Ðbres is the volume ofpores of mean diameter P1 km allowing the di†usionof the Ñora into the Ðbre network and the presentationof a large area of substrate to the bacteria associatedenzymes. In this respect, the presence of ulvan in thewhole algae limited the porosity of the plant to bacteriasince its solubilisation liberated pores and/or contrib-uted to the reorganisation of the cell wall so that alarger macropore volume was present in Ins (Table 3).
Considering the content of soluble (16É5%) and insol-uble Ðbres (13É3%) with the total algal Ðbre (29É8%) andthe extent of their degradation as the percentage of totalsugar disappeared after 24 h incubation with faecal bac-teria (D26%, 51% and 32% for the soluble, insolubleÐbres and whole algae, respectively, Bobin-Dubigeon etal 1997), the isolated Ðbre was more degraded thanwhen they were associated in the starting plant material.Indeed, on 100 g of Ðbres, about 11É1 g of isolatedsoluble and insoluble Ðbres were removed by the faecalÑora compared to 9É5 g when these polysaccharideswere in the whole Ulva sp. Furthermore, this lattervalue was probably an overestimate due to the presenceof a small amount of starch in the whole algae that wasmost likely used by the Ñora. In the light of the presentresults, this behaviour, which has already beendescribed for cereal Ðbres (Bourquin et al 1992), wasattributed to the presence of the degradation resistantacid polysaccharides (particularly ulvan) which limit theaccess of bacteria to their substrate, encrust a-celluloseand can potentially partially inhibit enzymatic activity.Similar limitations were probably a†ecting the conver-sion of Ulva sp to methane during fermentation trialsfor biogas production (Brouard 1983 ; Morand et al1991) as ulvan appeared to be the most resistant poly-mers. The role of ulvan and glucuronan in limiting Ulvasp fermentation is similar to that observed for otheralgal water-soluble polysaccharides such as alginate and

350 C Bobin-Dubigeon et al
carrageenans in brown and red seaweeds fermentations(King et al 1985 ; et al 1993). To improve fer-^stgaardmentation of these biomasses or food ingredients, pro-cesses increasing the macroporosity of the substratesuch as the enzymatic degradation of the celluloseassociated polysaccharides or the extraction of the latterby cooking in water, for example, are required.
ACKNOWLEDGEMENTS
The authors thank Dr P Debeire from INRA, Ville-neuve dÏAscq, for the gift of the puriÐed endo-xylanaseand T Chevalier and M Rival for their excellent techni-cal assistance. This work was supported in part by theEU contract AIR 1CT 92-0518.
REFERENCES
Arasaki S, Arasaki T 1985 L es de Mer. CommentL e� gumes eü treen Forme. Guy Tredaniel Edition de la Maisnie, Paris,France, pp 1È220.
Au†ret A, Barry J-L, Thibault J-F 1991 In vitro degradationof chemically treated sugar-beet Ðbres by human faecal bac-teria. Food Hydrocoll 5 41È44.
Barry J-L, Chourot J-M, Bonnet C, Kozlowski F, David A1989 In vitro fermentation of neutral monosaccharides byruminal and human faecal microÑora. Acta V et Scand S693È95.
BeMiller J N 1967 Acid-catalyzed hydrolysis of glycosides.Adv Carbohydr Chem 22 25È108.
Bertonniere N R 1989 Pore structure analysis of cotton cellu-lose via gel permeation chromatography. In : Cellulose,Structural and Functional Aspects, eds Kennedy J F, PhillipsG O & Williams P A. Ellis Horwood Limited, Chichester,UK, pp 99È104.
Blakeney A B, Harris P J, Henry R J, Stone B A 1983 Asimple and rapid preparation of alditol acetates for mono-saccharide analysis. Carbohydr Res 113 291È299.
Bobin-Dubigeon C, Lahaye M, Barry J-L 1997 Humancolonic bacterial degradability of dietary Ðbres from sea-lettuce (Ulva sp). J Sci Food Agric 73 149È159.
Bourquin L D, Titgemeyer E C, Garleb K A, Fahey G C 1992Short-chain fatty acid production and Ðber degradation byhuman colonic bacteria : e†ects of substrate and cell wallfractionation procedures. J Nutr 122 1508È1520.
Bra- ten T, A 1968 On the ultrastructure of vegetativeLÔvlieand sporulating cells of the multicellular green alga Ulvamutabilis. Fyn Nytt Mag Bot 15 209È219.
Brouard F 1983 Digestion de la biomasseanae� robie ve� ge� taleaquatique. These Docteur Inge� nieur, Institut National desSciences Applique� es de Toulouse, France, p 120.
Bryhni E 1978 Quantitative di†erences between poly-saccharide compositions in normal di†erentiated Ulvamutabilis and the undi†erentiated mutant lumpy. Phycologia17 119È124.
Callow M E, Evans L V 1977 Studies on the ship-fouling algaEnteromorpha (Chlorophycease, Ulvales) I. V. poly-saccharide and nucleoside diphosphatase localisation. Phy-cologia 163 313È320.
Cloutour F 1995 Caracte� ristiques des Ðbres alimentaires :inÑuence sur leur fermenation in vitro par la Ñore digestivede lÏhomme. These de Doctorat de lÏUniversite de Nantes,France, p 123.
Cronshaw J, Myers A, Preston R D 1958 Chemical and physi-cal investigation of the cell walls of some marine algae.Biochim Biophys Acta 27, 89È103.
Durand M, Beaumatin P, Bulman B, Bernalier A, Grivet J-P,Serezat M, Gramet G, Lahaye M 1997 Fermentation of thegreen alga sea-lettuce (Ulva sp) and metabolism of its sul-phate by human colonic microbiota in a semi-continuousculture system. Reprod Nutr Dev 37 267È283.
Guillon F, Au†ret A, Robertson J A, Thibault J-F, Barry J-L1996 E†ects of treatments on the physical characteristicsand fermentability in sugar-beet Ðbres. Carbohydr Polym.(in press).
Hoebler C, Barry J-L, David A, Delort-Laval J 1989 Rapidacid hydrolysis of plant cell wall polysaccharides and sim-pliÐed quantitative determination of their neutral monosac-charides by gas-liquid-chromatography. J Agric Food Chem37 360È367.
Indergaard M, Minsaas K 1991 Polysaccharides for food andpharmaceutical uses. In : Seaweed Resources in Europe. Usesand Potential, eds Guiry M D & Blunden G. John Wiley,Chichester, UK, pp 169È184.
Joseleau J-P, Ruel K 1985 A new cytochemical method forultrastructure localisation of polysaccharides. Biol Cell 5361È66.
King G M, Guist G G, Lauterbach G E 1985 Anaerobic deg-radation of carrageenan from red macroalga Eucheuma cot-tonii. Appl Environ Microbiol 49 588È592.
Lahaye M 1991 Marine algae as sources of Ðbres : determi-nation of soluble and insoluble dietary Ðbre content in some“sea vegetablesÏ. J Sci Food Agric 54 587È594.
Lahaye M, Axelos M 1993 Gelling properties of water-solublepolysaccharides from proliferating marine green seaweeds(Ulva spp.) Carbohydr Polym 22 261È265.
Lahaye M, Jegou D 1993 Chemical and physico-chemicalcharacteristics of dietary Ðbres from Ulva lactuca (L) Thuretand Enteromorpha compressa (L) Grev J Appl Phycol 5 195È200.
Lahaye M, Jegou D, Bule� on A 1994 Chemical characteristicsof insoluble glucans from the cell wall of the marine greenalgae Ulva lactuca (L.) Thuret Carbohydr Res 262 115È125.
Lahaye M, Gomez-Pinchetti J-L, Jimenez Del Rio M, Garcia-Reina G (1995) Natural decoloration, composition andincrease in dietary Ðbre content of an edible marine algae,Ulva rigida (Chlorophyta), grown under di†erent nitrogenconditions. J Sci Food Agric 68 99È104.
Lahaye M, Ray B, Baumberger S, Quemener B, Axelos M AV 1996 Chemical characterisation and gelling properties ofcell-wall polysaccharides from species of Ulva (Ulvales,Chlorophyta). Hydrobiologia 326/327 473È480.
A, Bra- ten T 1968 On the division of cytoplasm andLÔvliechloroplast in the multicellular green alga Ulva mutabilis
Exper Cell Res 51 211È220.FÔyn.Lowry O H, Rosebrough N J, Farr A L, Randall R J 1951
Protein measurement with the Folin-phenol reagent. J BiolChem 93 265È275.
Luft J H 1971 Ruthenium red and violet. IÈChemistry, puriÐ-cation, methods of use for electron microscopy. Mechanismof action. Anat Rec 171 347È368.
Micalef H, Gayral P 1972 Quelques aspects de lÏinfrastructuredes cellules ve� ge� tatives et des cellules reproductrices dÏUlvalactuca L. (Chlorophyce� es). J Microscopie 13 417È427.
Morand P, Carpentier B, Charlier R H, Maze� J, Orlandini M,Plunkett B A, De Waart J 1991 Bioconversion of seaweeds.In : Seaweed Resources in Europe. Uses and Potential, eds

Factors limiting the biodegradation of Ulva sp cell-wall polysaccharides 351
Guiry M D & Blunden G. John Wiley, Chichester, UK pp95È148.
Nelson N 1944 A photometric adaptation of the Somogyimethod for the determination of glucose. J Biol Chem 153375È380.
Neuman R P, Walker L P 1992 Solute exclusion from cellu-lose in packed columns : experimental investigation andpore volume measurement. Biotechnol Bioengng 40 218È225.
K, Indergaard M, Markussen S, Knutsen S H,^stgaardJensen A 1993 Carbohydrate degradation and methane pro-duction during fermentation of L aminaria saccharina(Laminariales, Phaeophyceae). J Appl Phycol 5 333È342.
Painter T J 1983 Algal polysaccharides. In : T he Poly-saccharides, ed Aspinall G O. Academic Press, London,UK, pp 195È285.
Parker B C, Diboll A G 1966 Alcian stains for histochemicallocalisation of acid and sulphated polysaccharides in algae.Phycologia 6 37È46.
Pazur J H 1986 Neutral polysaccharides. In : CarbohydrateAnalysis, A Practical Approach, eds Chaplin M F &Kennedy J F. IRL Press, Oxford, UK, pp 73È124.
Percival E, McDowell R H 1967 Chemistry and Enzymology ofMarine Algal Polysaccharides. Academic Press, London,UK.
Phillips J A 1988 Field, anatomical and developmental studieson southern australian species of Ulva (Ulvaceae,Chlorophyta). Aust Syst Bot 1 411È438.
Pierre J-F, Kilbertus G, Reisinger O 1974 Observations ultra-structurales de la biode� gradation dÏune algue dans un
e� cosysteme aquatique. Bulletin et L or-Acade� mie Socie� te�raines des Sciences XIII(2) 137È153.
Riva A 1974 A simple and rapid staining method for enhanc-ing the contrasts in tissues previously treated with uranylacetate. J Microscopie 19 105È106.
Robertson J A 1988 Physicochemical characteristics of foodand the digestion of starch and dietary Ðbre during guttransit. Proc Nutr Soc 47 143È152.
Robertson J A, Eastwood M A 1981 A method to measure thewater-holding properties of dietary Ðbre using suction pres-sure. Br J Nutr 46 247È255.
Salyers A A, Vercellotti J R, West S E H, Wilkins T D 1977Fermentation of mucin and plant polysaccharides by strainsof Bacteroides from the human colon. Appl Environ Micro-biol 33 319È322.
Thibault J-F 1979 Automatisation du dosage des substancespectiques par la me� thode au me� tahydroxydiphe� nyl.L ebensm W iss U T echnol 12 247È251.
Thie� ry J-P 1967 Mise en e� vidence des polysaccharides surcoupes Ðnes en microscopie e� lectronique. J Microscopie 6987È1018.
Wilson J R, Mertens D R 1995 Crop quality and utilisation :cell wall accessibility and cell structure limitations to micro-bial digestion of forage. Crop Sci 35 251È259.
Wood P J, Fulcher R G, Stone B A 1983 Studies on the speci-Ðcity of the interaction of cereal cell wall components withCongo Red and CalcoÑuor. SpeciÐc detection and histo-chemistry of (1-3)(1-4)-b-D-glucan. J Cereal Sci 1 95È110.





























