Embed Size (px)
Citation preview

ゲノム編集技術が与える 医学へのインパクト
―遺伝子改変マウス作製への応用を中心に―
メディカルサイエンス推進研究所 遺伝子実験センター 中 島 修
1
2015.7.22 メディカルサイエンス推進研究所技術セミナー

ゲノム編集はPCR以来のゲームチェンジャー(game changer:世論の動向を大きく変える出来事)
2 20 | NATURE | VOL 522 | 4 JUNE 2015

3
20 | NATURE | VOL 522 | 4 JUNE 2015

4
20 | NATURE | VOL 522 | 4 JUNE 2015

本日のメニュー
1. ゲノム編集の原理 2. ゲノム編集の歴史と展望 3. マウス受精卵でのゲノム編集 4. ゲノム編集の遺伝子改変マウス作製への応
用 5. 山形大学医学部遺伝子実験センターでのこ
れまでのゲノム編集による遺伝子改変マウスの作製の実際
6. 山形大学医学部遺伝子実験センターでの受託作製の手順
5

ゲノム編集技術とは?
ゲノム編集(Genome Editing)とは、人工ヌクレアーゼのZinc Finger Nucleases(ZFNs)やTranscription Activator-Like Effector Nucleases(TALENs)、CRISPR/Casシステムを用いてゲノム上
の標的遺伝子の破壊やレポーター遺伝子のノックインなどを可能にする技術である。(Joung and Sander, Nat Rev Mol Cell Biol, 2012; Barrangou, Nat Biotechnol, 2012)。
6
ゲノム編集コンソーシアムHPより http://www.mls.sci.hiroshima-u.ac.jp/smg/genome_editing/index.html

ゲノム編集で利用される人工ヌクレアーゼ 人工ヌクレアーゼ(ZFNやTALEN)は、DNAに特異的に結合するドメインと、制限酵素FokIのDNA切断ドメインを連
結させたキメラタンパク質である。2つの人工ヌクレアーゼが近接する標的配列に結合するとDNA切断ドメインが2量体となりDNAを切断する。
7
ZFN,TALEN
特異的に認識できる配列をデザインするには、何種類かのDNA結合ドメインを適切に組み合わせた融合タンパク質をつくる必要があり、初心者は手を出しにくい系である
ゲノム編集コンソーシアムHPより http://www.mls.sci.hiroshima-
u.ac.jp/smg/genome_editing/index.html

CRISPR/Casシステムとは?
CRISPR (Clustered Regularly Interspaced Short Palindromic Repeat; クリスパー) には多くのプロトスペーサー配列が含まれており、この配列はウィルス(ファージ)ゲノムなどの種々の外来DNAと相同性がある。このプロトスペーサー配列をもったRNAがCAS(Crispr associated, an RNA-guided DNA endonuclease associated with the CRISPR)に結合して、プラスミドやファージといった外来DNAを識別するプローブとなって、 CRISPR/Casが外来DNAのみを切断させるため、細菌の獲得免疫として機能している。
8

9 http://www.addgene.org/crispr/reference/history/
細菌ゲノム内CRISPRが存在
CRISPRが転写され、pre-crRNAが出来る
pre-crRNAとtracrRNAがCas9に結合し、さらにRNaseIIIが結合する
RNaseIIIがpre-crRNAを切断して、crRNA:tracrRNA:Cas9複合体を形成させる
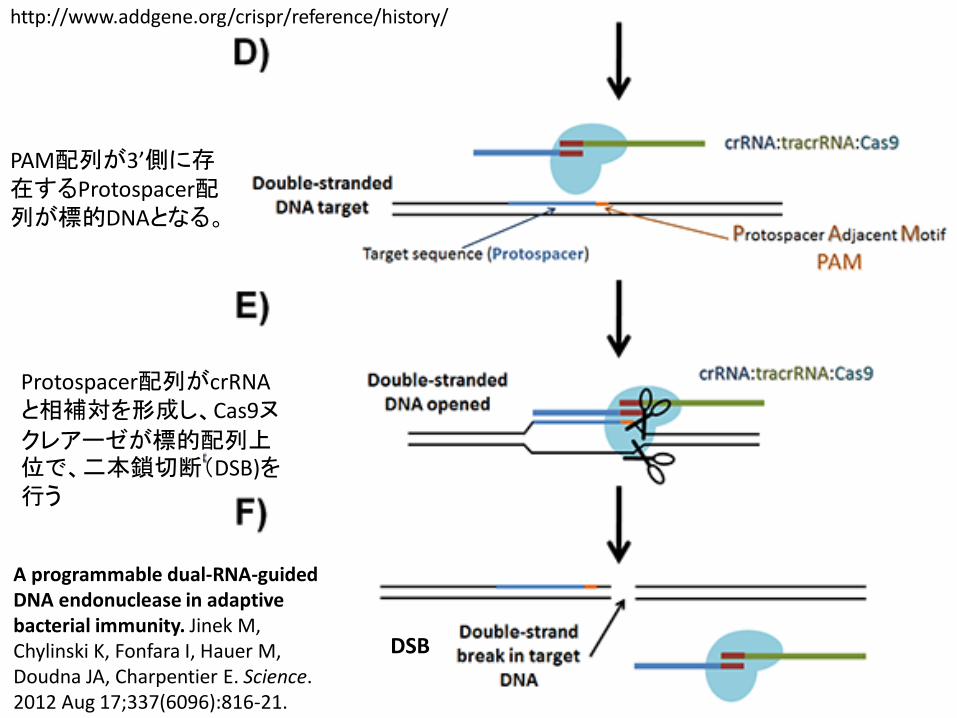
10
http://www.addgene.org/crispr/reference/history/
PAM配列が3’側に存在するProtospacer配列が標的DNAとなる。
Protospacer配列がcrRNAと相補対を形成し、Cas9ヌクレアーゼが標的配列上位で、二本鎖切断(DSB)を行う
DSB
A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Jinek M, Chylinski K, Fonfara I, Hauer M, Doudna JA, Charpentier E. Science. 2012 Aug 17;337(6096):816-21.

Figure 6-27 Essential Cell Biology (© Garland Science 2010)
非相同末端連結nonhomologous end-joiningによる二本鎖切断の修復
偶発的な二本鎖切断
切断部位の末端をヌクレアーゼにより加工
DNAリガーゼにより末端を連結
二本鎖切断は修復されるが、修復部位でヌクレオチドの欠失が生じる
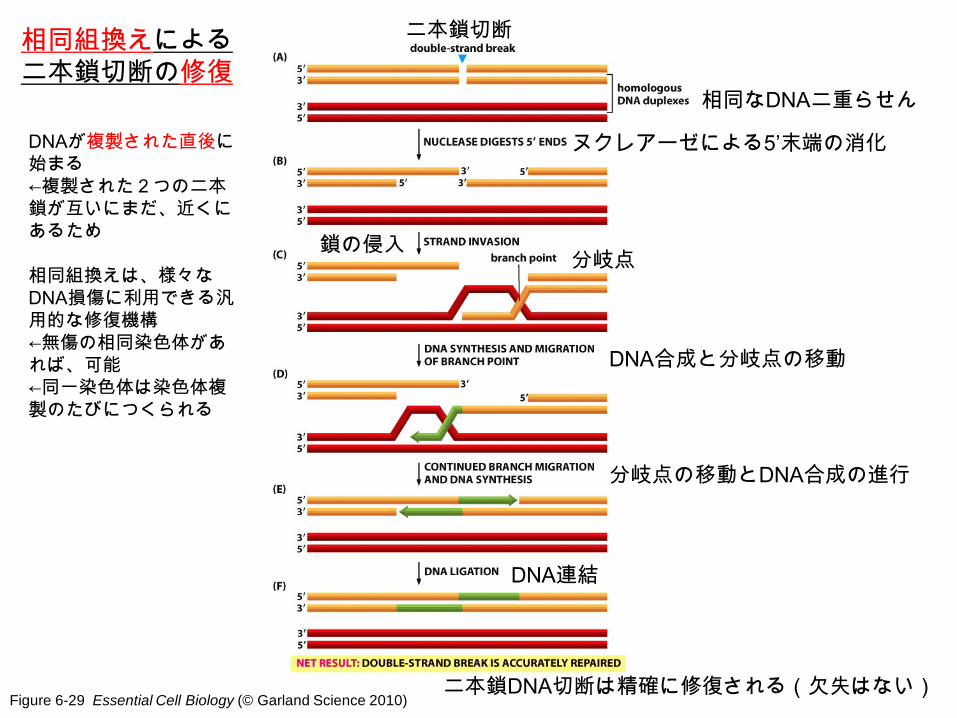
Figure 6-29 Essential Cell Biology (© Garland Science 2010)
相同組換えによる二本鎖切断の修復
相同なDNA二重らせん
二本鎖切断
ヌクレアーゼによる5’末端の消化
鎖の侵入 分岐点
DNA合成と分岐点の移動
分岐点の移動とDNA合成の進行
DNA連結
二本鎖DNA切断は精確に修復される(欠失はない)
DNAが複製された直後に始まる ←複製された2つの二本鎖が互いにまだ、近くにあるため 相同組換えは、様々なDNA損傷に利用できる汎用的な修復機構 ←無傷の相同染色体があれば、可能 ←同一染色体は染色体複製のたびにつくられる

(NHEJ)
小さい欠失等が生じる
(HR)
二本鎖DNA切断(DSB) 二本鎖DNA切断(DSB)
相同染色体を使って欠失を起こさず修復
相同染色体
細胞の中で起こったDNA二本鎖切断の2通りの修復
京都大HPより 13
相同なDNAが存在する場合のみ起こる
ヌクレアーゼにより5’末端の一部が消化され削られる
相同染色体(DNA)を鋳型にして修復される
ヌクレアーゼにより5’末端の一部が消化され削られる

14 http://www.addgene.org/crispr/guide/
Non Homologous End Joining Repair

15 http://www.addgene.org/crispr/guide/
Single or double strand Donor DNA
Homology Directed Recombinant Repair
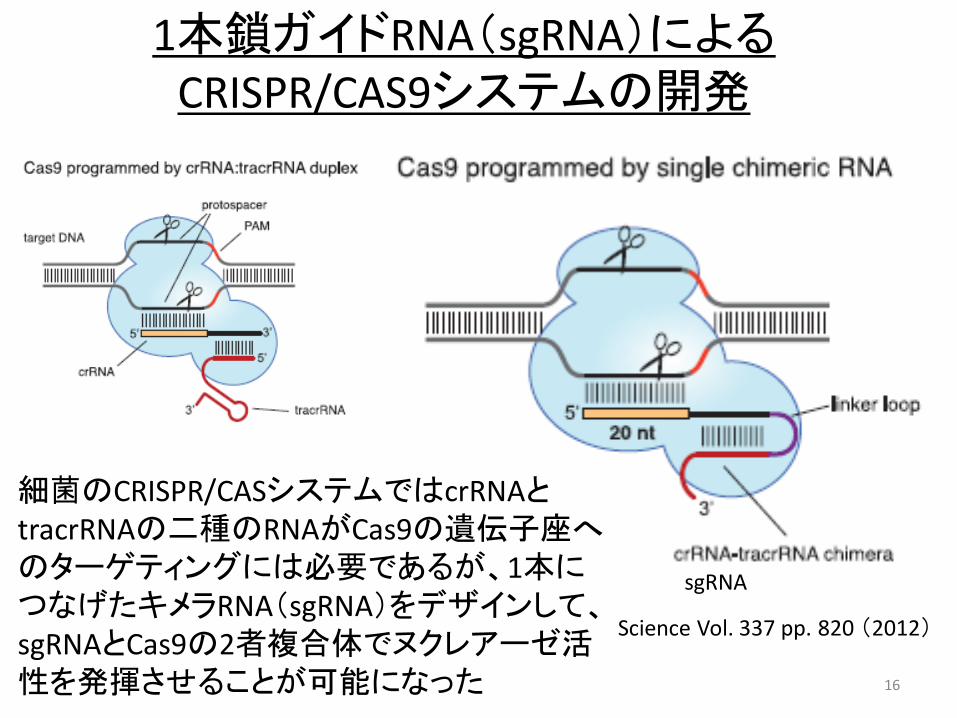
16
1本鎖ガイドRNA(sgRNA)による CRISPR/CAS9システムの開発
sgRNA
細菌のCRISPR/CASシステムではcrRNAとtracrRNAの二種のRNAがCas9の遺伝子座へのターゲティングには必要であるが、1本につなげたキメラRNA(sgRNA)をデザインして、sgRNAとCas9の2者複合体でヌクレアーゼ活性を発揮させることが可能になった
Science Vol. 337 pp. 820 (2012)

CRISPR/CAS9システム用オールインワンベクターpX330
17
U6:全身性U6プロモーター。U6遺伝子はスプライソソームを構成する5つの核内低分子リボ核タンパク質(snRNP)の一つ CBh:全身性トリβアクチンハイブリッドプロモーター
標的遺伝子座に対するガイドシークエンス20bpをクローニングすると、目的のsgRNAとCas9 mRNAを同時に
発現させることが可能な、哺乳類細胞での発現ベクターとなる。これを細胞の中に入れれば、二本鎖切断が起こせる http://www.addgene.org/

CRISPR/CASによるゲノム編集とは ゲノムの任意の遺伝子座で、遺伝子破壊と任意の
変異の導入(編集)を行う技術
1.非相同末端連結 Non Homologous End Joining (NHEJ)による修復が起こる場合 ⇒当該遺伝子で欠失が生じ、exon上で起こった場合はフレームシフト変異となる可能性が高いため、遺伝子破壊ができる。 2.人工的に任意の変異を導入した相同遺伝子断片(ドナーDNA)を修復時に同時に存在させた時に、相同組換え修復 Homologous Recombinant Repair (HRR)による修復が起こる場合 ⇒当該遺伝子には、ドナーDNAに由来する変異が導入される(編集される)
18
CRISPR/CASにより生じた二本鎖切断に対して

本日のメニュー
1. ゲノム編集の原理 2. ゲノム編集の歴史と展望 3. マウス受精卵でのゲノム編集 4. ゲノム編集の遺伝子改変マウス作製への応
用 5. 山形大学医学部遺伝子実験センターでのこ
れまでのゲノム編集による遺伝子改変マウスの作製の実際
6. 山形大学医学部遺伝子実験センターでの受託作製の手順
19

20
Ishino, Y., Shinagawa, H., Makino, K., Amemura, M. & Nakata, A. J. Bacteriol. 169,5429–5433 (1987).
20 | NATURE | VOL 522 | 4 JUNE 2015

21
Jinek, M. et al. Science 337, 816–821 (2012).
20 | NATURE | VOL 522 | 4 JUNE 2015

点変異や短い配列の挿入を起こす場合は1本鎖DNAでも可能
欠失が多いが小さな置換なども起こりうる
点変異導入や元々ない配列の挿入が可能
タカラバイオHPより 22

23
ヌクレアーゼ活性を持たないDead Cas
転写制御 エピゲノム修飾 イメージング
染色体レベルでの遺伝子改変
CRISPR/Cas9の応用

24
https://en.wikipedia.org/wiki/Epigenome_editing

25 20 | NATURE | VOL 522 | 4 JUNE 2015

26
Oye, K. A. et al. Science 345, 626–628 (2014).
“CRISPR Gene drive”

27 20 | NATURE | VOL 522 | 4 JUNE 2015

28
ヒト胚(ただし、前核が3つある異常受精卵)に対するCRISPR/Cas9を介したゲノム編集

29

Cell Cycle Ahead of Print DOI: 10.1080/15384101.2015.1046791
ヒト幹細胞でのゲノム編集によるHIVに対する遺伝子治療
CCR5 found on the surface of white blood cells acts as a co-receptor for HIV entry to the host cells.
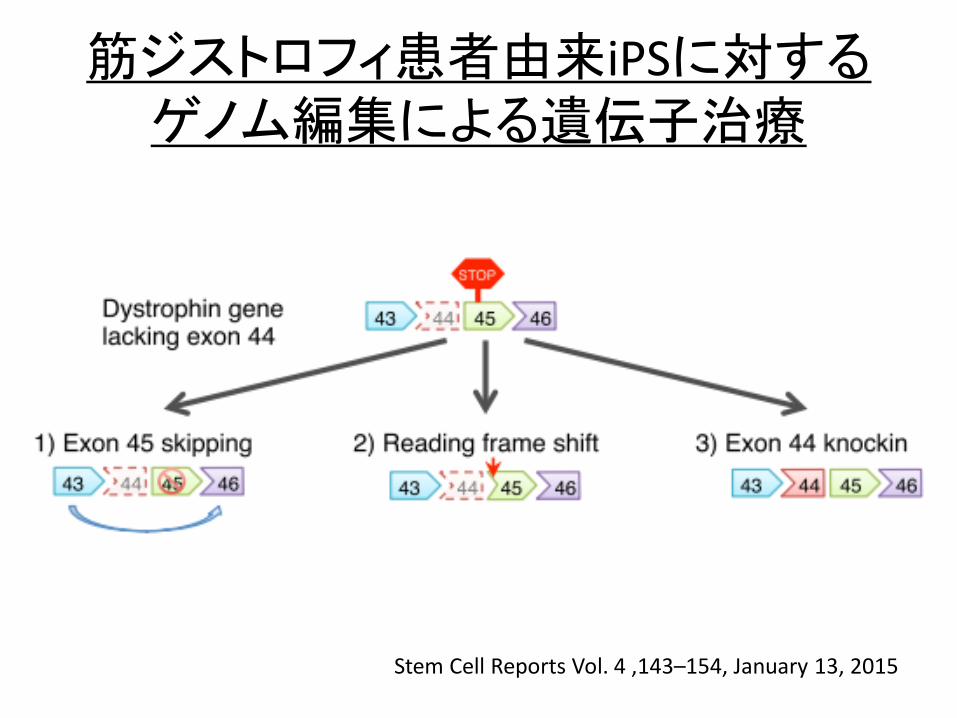
筋ジストロフィ患者由来iPSに対する ゲノム編集による遺伝子治療
Stem Cell Reports Vol. 4 ,143–154, January 13, 2015

Stem Cell Reports Vol. 4 , 143–154, 2015

ゲノム編集によるヒト胚に対する遺伝子治療の論点
• 精確さ inaccurate editing and off-target(二本鎖切断が起こる目的外遺伝子座) mutations
• Monogenic disease such as Huntington’s disease • 胚に対するゲノム編集は現実的に困難 • IVF embryos using preimplantation genetic diagnosis (PGD) • PGDによる胚の選別による代替可能なケースが多い • ホモ接合体の胚(例:同一遺伝子変異の聾者同士の胚)の場合はPGD代替不可
• Reducing the risk of common diseases like APOE ε4 variant (Alzheimer’s disease and cardiovascular disease), PCSK9 (myocardial infraction)
33 From Eric S. Lander. "Brave New Genome." N. Engl. J. Med. Published online 2015 June 3

本日のメニュー
1. ゲノム編集の原理 2. ゲノム編集の歴史と展望 3. マウス受精卵でのゲノム編集 4. ゲノム編集の遺伝子改変マウス作製への応用 5. 山形大学医学部遺伝子実験センターでのこれ
までのゲノム編集による遺伝子改変マウスの作製の実際
6. 山形大学医学部遺伝子実験センターでの受託作製の手順
34

マウス受精卵でのゲノム編集の報告
35 Cell. 2013 May 9; 153(4): 910–918.

36
A ES細胞での5重遺
伝子破壊クローンの作成 B マウス受精卵での二重遺伝子破壊マウスの作製 マウス受精卵での二重相同組み換え体の作製
Cell. 2013 May 9; 153(4): 910–918.

37 Cell. 2013 May 9; 153(4): 910–918.
Ten-eleven translocation family members Tet1, Tet2, Tet3での標的配列
エキソン内にある制限酵素サイトを切断するデザイン
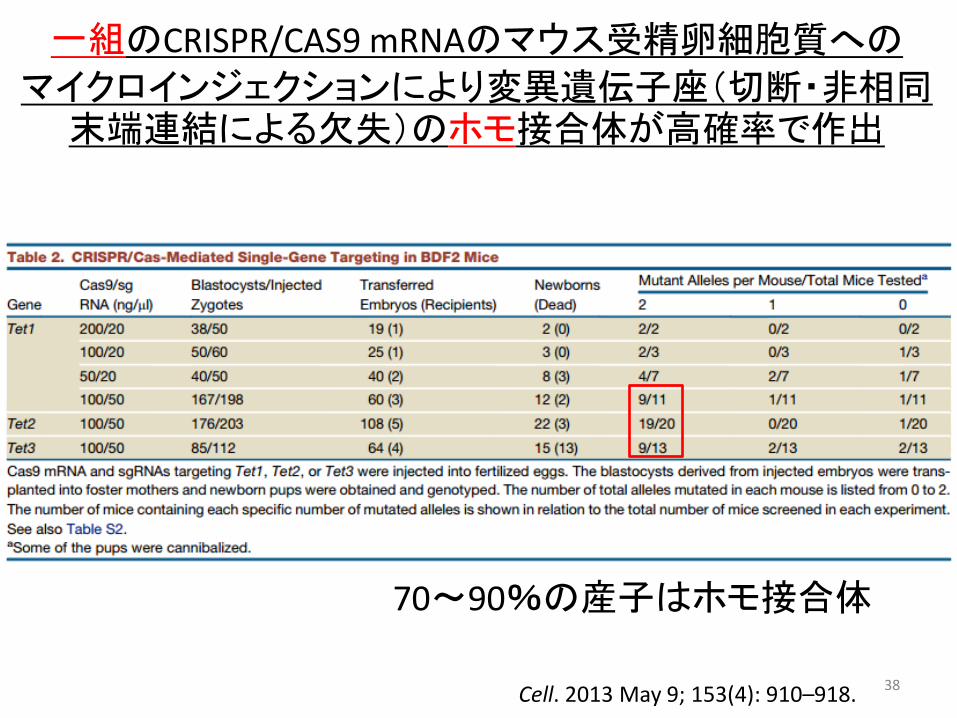
38
一組のCRISPR/CAS9 mRNAのマウス受精卵細胞質への マイクロインジェクションにより変異遺伝子座(切断・非相同
末端連結による欠失)のホモ接合体が高確率で作出
70~90%の産子はホモ接合体
Cell. 2013 May 9; 153(4): 910–918.

39 実際に出来る変異遺伝子座は数bp~数十bpの欠失が多いが、挿入の場合もある
制限酵素サイトが無くなるので、検出される断片が長くなる バンドが1本なのでホモが多い

40
二種のCRISPR/CAS9 mRNAのマウス受精卵前核への マイクロインジェクションによる 二重遺伝子破壊マウスの作出
Cell. 2013 May 9; 153(4): 910–918.

41
二種のCRISPR/CAS9 mRNA・1本鎖ドナーDNAのマウス受精卵
前核へのマイクロインジェクションによる 二重2塩基変異導入マウスの作出
相同組換えのホモ
相同組換えと欠失のヘテロ
相同組換えと欠失のヘテロ
相同組換えと欠失のヘテロ
Cell. 2013 May 9; 153(4): 910–918.

42 http://www.egr.biken.osaka-u.ac.jp/CRISPRデザイン法20131219.pdf
大阪大学微生物病研究所 伊川正人先生のプロトコール

43 http://www.egr.biken.osaka-u.ac.jp/CRISPRデザイン法20131219.pdf

44 http://www.egr.biken.osaka-u.ac.jp/CRISPRデザイン法20131219.pdf
Single Strand Annealing
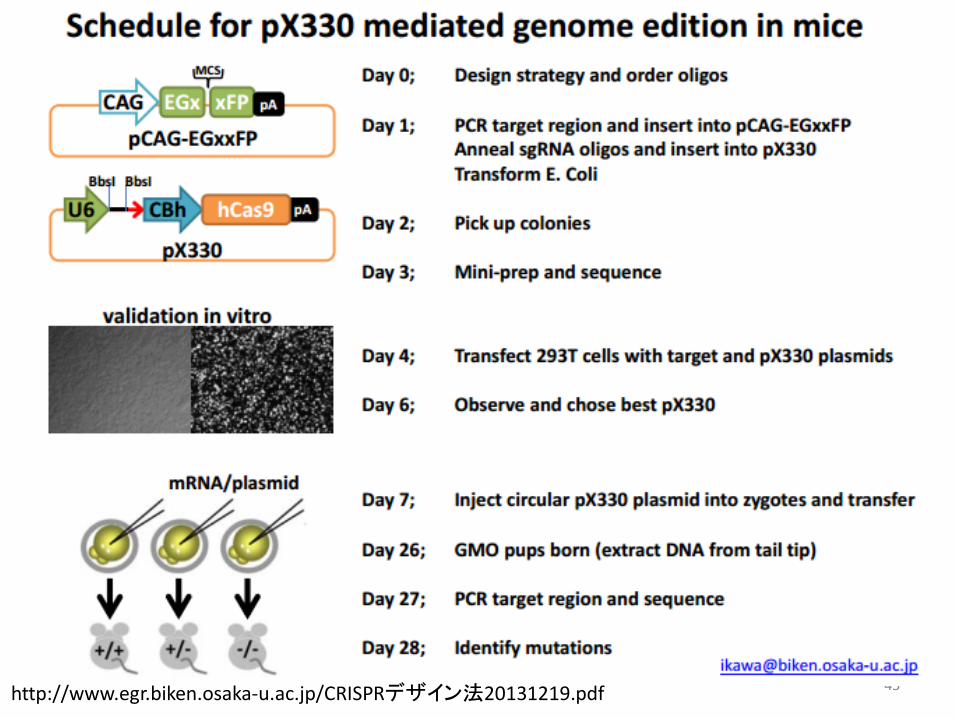
45 http://www.egr.biken.osaka-u.ac.jp/CRISPRデザイン法20131219.pdf

CRISPR/Cas9 vector
環状のCRISPR/Cas9 vectorとdonor oligo DNAを導入 問題点:ベクターのゲノム挿入が起こりうる。RNAインジェクションではこの点は回避できる。
CRISPR/Cas9プラスミドベクターの C57BL/6Jマウス受精卵への導入
Donor oligo
pX330
CBh Flag-NLS-hSpCas9-NLS pA U6 target gRNA
筑波大学基礎医学系 高橋智教授 H26山形大学大学院講義資料より一部改変

NHEJ: 非相同末端連結 HR: 相同組換え修復
熊本大HPより

CRISPR/CASによる個体でのゲノム編集の利点
• 汎用性が高く、簡便で低コスト(↔ ZFN・TALEN , ES細胞)
• 受精卵が得られればどんな生物種でも可能(実施例:ラット、ウサギ、豚、ゼブラフィッシュ、アクソロートル、サル)
• もちろん、マウス系統もC57BL/6に限らない • 遺伝子改変マウスが比較的短期間で作出可能 • 多重遺伝子破壊が簡便にできる • 1回の実験でホモ変異体の作出が期待できる(交配不要)
• 単純遺伝子破壊のみならず、ドナーDNA次第で様々な遺伝子改変が可能
48

Off-target問題
• Few studies using whole genome analysis already showed that the mutation rates induced with Cas9 endonucleases are not systematically higher than spontaneous (non-induced) natural mutations
• Off-target remains ultimately much less critical for mice as for human genome engineering, as it is easily diluted out amongst generations
49
Delerue and Ittner, Clon Transgen 2015, 4:1 http://dx.doi.org/10.4172/2168-9849.1000135

本日のメニュー
1. ゲノム編集の原理 2. ゲノム編集の歴史と展望 3. マウス受精卵でのゲノム編集 4. ゲノム編集の遺伝子改変マウス作製への応
用 5. 山形大学医学部遺伝子実験センターでのこ
れまでのゲノム編集による遺伝子改変マウスの作製の実際
6. 山形大学医学部遺伝子実験センターでの受託作製の手順
50

51 Science 5 SEPTEMBER 2014 • VOL 345, 1184-1188
筋ジストロフィーマウス受精卵に対するCRISPR/Cas9を介したゲノム編集による 産子における筋ジストロフィ―発症予防
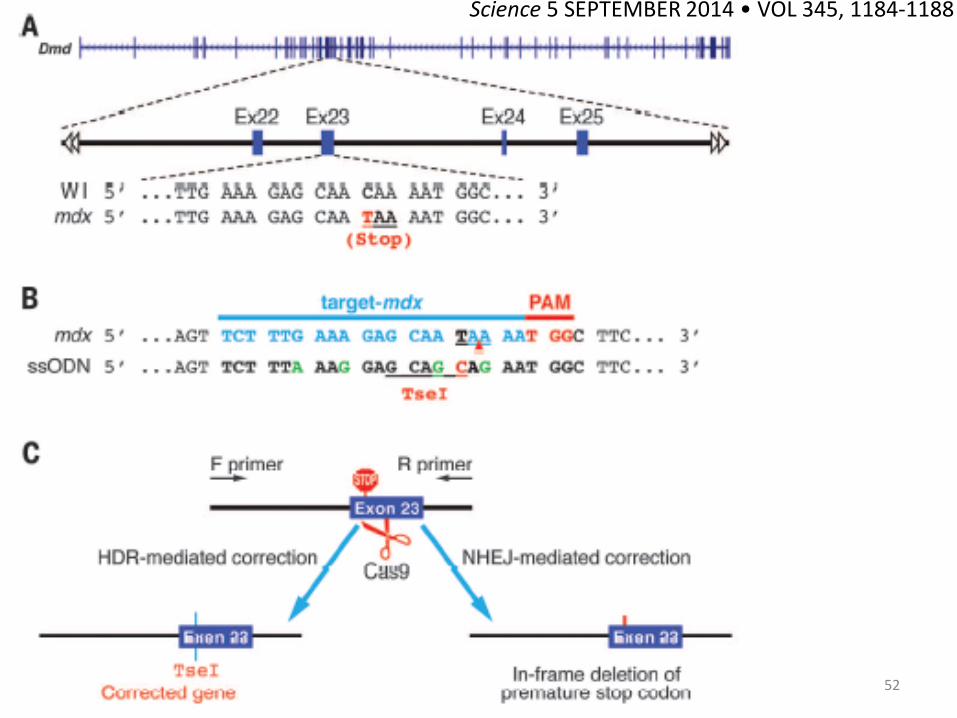
52
Science 5 SEPTEMBER 2014 • VOL 345, 1184-1188

53
Science 5 SEPTEMBER 2014 • VOL 345, 1184-1188

54
ゲノム編集マウスでのジストロフィンタンパク質発現の回復
Science 2014 345 1184-1188

55
ゲノム編集マウスでの血清クレアチンキナーゼの低下と前足握力の回復
Science 5 SEPTEMBER 2014 • VOL 345, 1184-1188

56
成体遺伝性チロシン血症マウスに対する sgRNA/Cas9mRNA/donor DNA尾静注による治療
(肝機能の回復)
Nat Biotechnol. 2014 Jun;32(6):551-3.
fumarylacetoacetate hydrolase (FAH)
Fah: フマリルアセト酢酸ヒドラーゼ遺伝子

57
成体遺伝性チロシン血症マウスに対する sgRNA/Cas9mRNA/donor DNA尾静注による治療
(肝機能の回復)
Nat Biotechnol. 2014 Jun;32(6):551-3.

58
Cell 159, 440–455, October 9, 2014
Rosa locus(すべての組織で発現)へのCas9 Knock-inマウスの確立とがん抑制遺伝子を標的とするsgRNAのウィルスベクターによる導入

59
Rosa26領域 Rosa26領域にβガラクトシダーゼ遺伝子が挿入されたマウスの系統は全身すべての細胞がX-gal染色陽性となるため,移植実験のコントロールとしてきわめて有用であった.さらにβガラクトシダーゼ遺伝子を他のマーカー遺伝子と置換しても全身にマーカーを発現されることが可能で,レポーターコンストラクトの挿入に頻用されている すなわち、どの組織でも発現するということは、組織特異的なエピジェネティックな抑制制御を受けない遺伝子座といえる。
https://www.yodosha.co.jp/jikkenigaku/keyword/474.html
Cell 159, 440–455, October 9, 2014
Schematic of the Cre-dependent Cas9 Rosa26 targeting vector Creにより、この領域が欠失するとCas9が発現する

60
In Vivo Genome Editing in the Brain of Cre-Dependent Cas9 Mice
Cell 159, 440–455, October 9, 2014

本日のメニュー
1. ゲノム編集の原理 2. ゲノム編集の歴史と展望 3. マウス受精卵でのゲノム編集 4. ゲノム編集の遺伝子改変マウス作製への応
用 5. 山形大学医学部遺伝子実験センターでのこ
れまでのゲノム編集による遺伝子改変マウスの作製の実際
6. 山形大学医学部遺伝子実験センターでの受託作製の手順
61

62
1本につなげたキメラRNA(sgRNA)をオリゴハウスでRNA合成するには長すぎるため、組換え実験が必要となる Science Vol. 337 pp. 820 (2012)
遺伝子実験センターでは組換え実験を省略するためcrRNA/tracrRNA/Cas9のシステムを利用
( crRNA/tracrRNA/Cas9 mRNAをco-injection)
75nt 共通で使える sgRNA 101nt
42nt 個別の実験でデザインする

遺伝子実験センターでのゲノム編集マウス受託作製の手順
1. 依頼者:ガイドRNA(crRNA)の配列のデザイン 2. 依頼者:オリゴRNA/ドナーDNAの注文 3. センター:ガイドRNA(crRNA)を介したDNA切断活
性のin vitroでの確認
4. センター:マウス受精卵前核へのCRISPR/CAS9 mRNA, ss donorDNAの導入
5. 依頼者:ゲノム解析による変異個体の確認
63
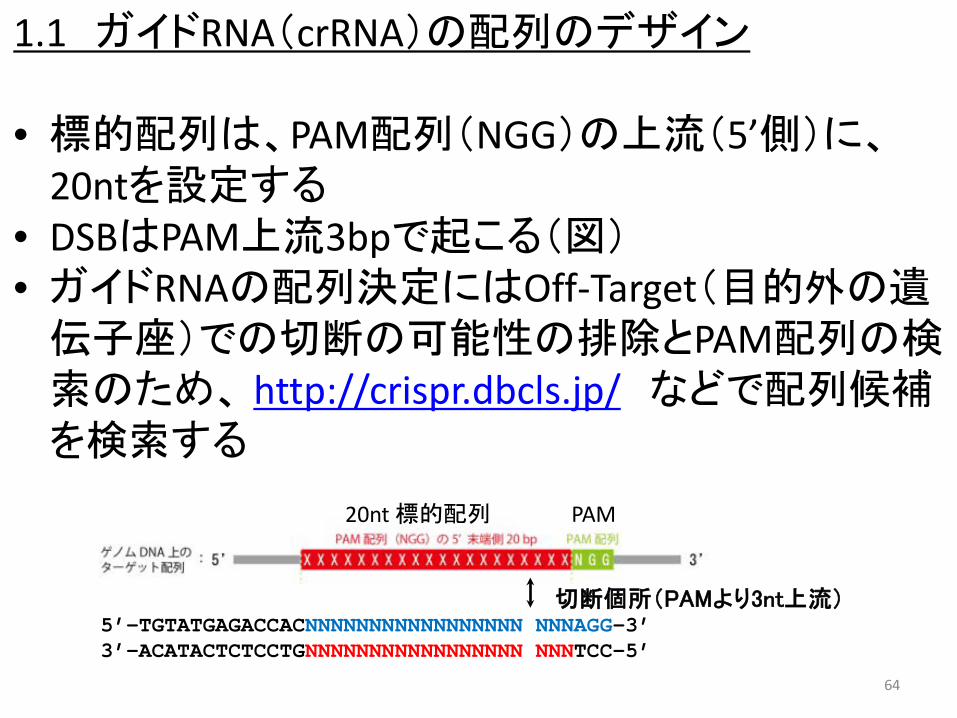
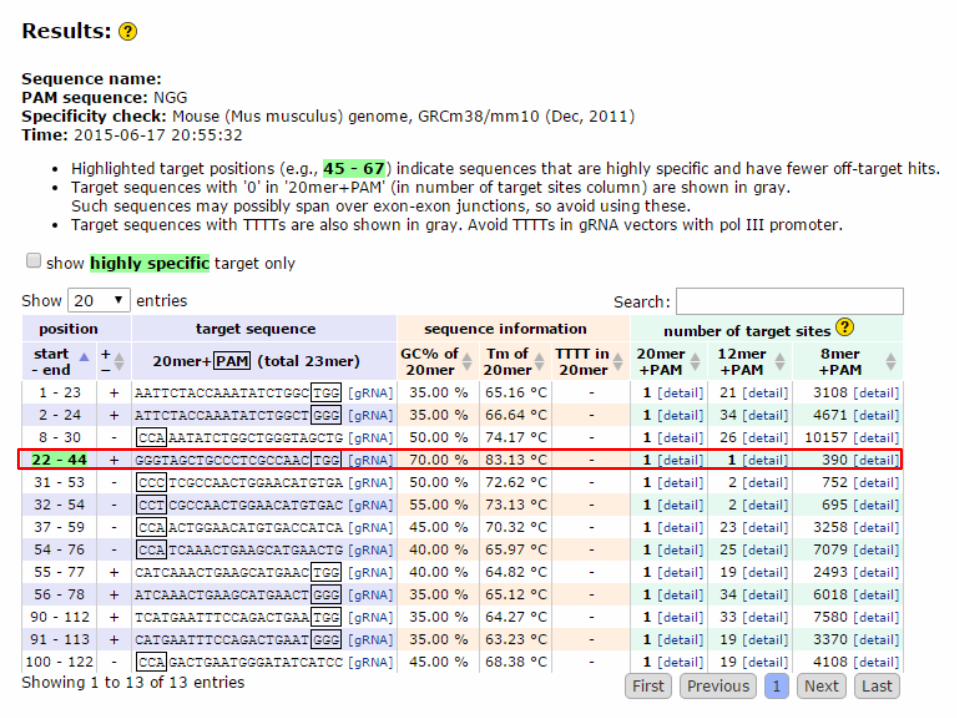
1.1 ガイドRNA(crRNA)の配列のデザイン
• 標的配列は、PAM配列(NGG)の上流(5’側)に、20ntを設定する
• DSBはPAM上流3bpで起こる(図) • ガイドRNAの配列決定にはOff-Target(目的外の遺伝子座)での切断の可能性の排除とPAM配列の検索のため、 http://crispr.dbcls.jp/ などで配列候補を検索する
5’-TGTATGAGACCACNNNNNNNNNNNNNNNNN NNNAGG-3’ 3’-ACATACTCTCCTGNNNNNNNNNNNNNNNNN NNNTCC-5’
切断個所(PAMより3nt上流)
64
PAM 20nt 標的配列

65
http://crispr.dbcls.jp/
66

67

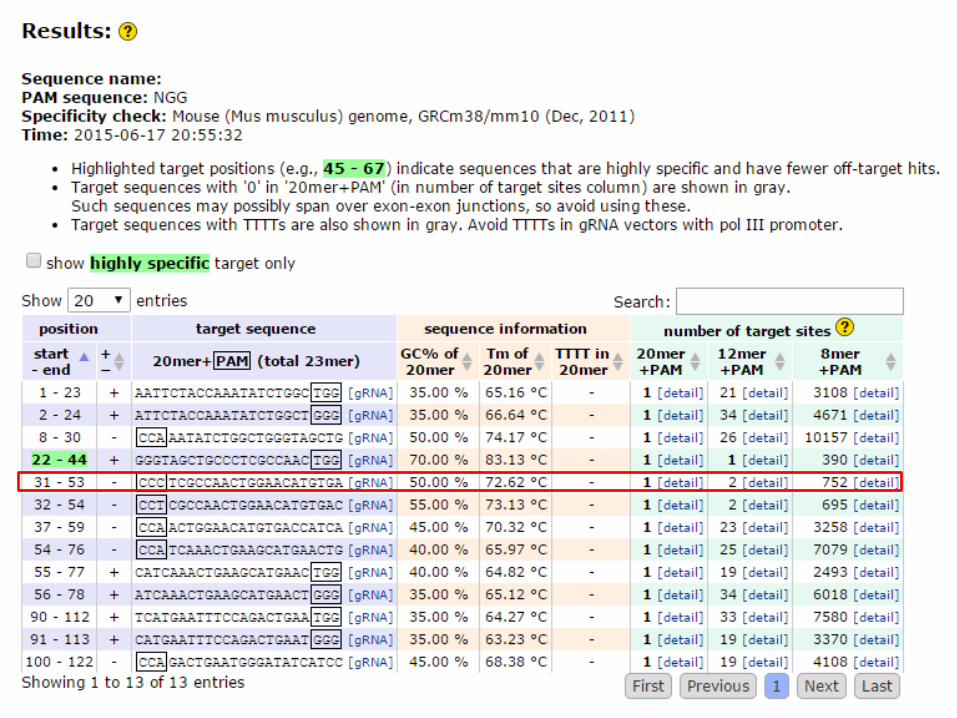
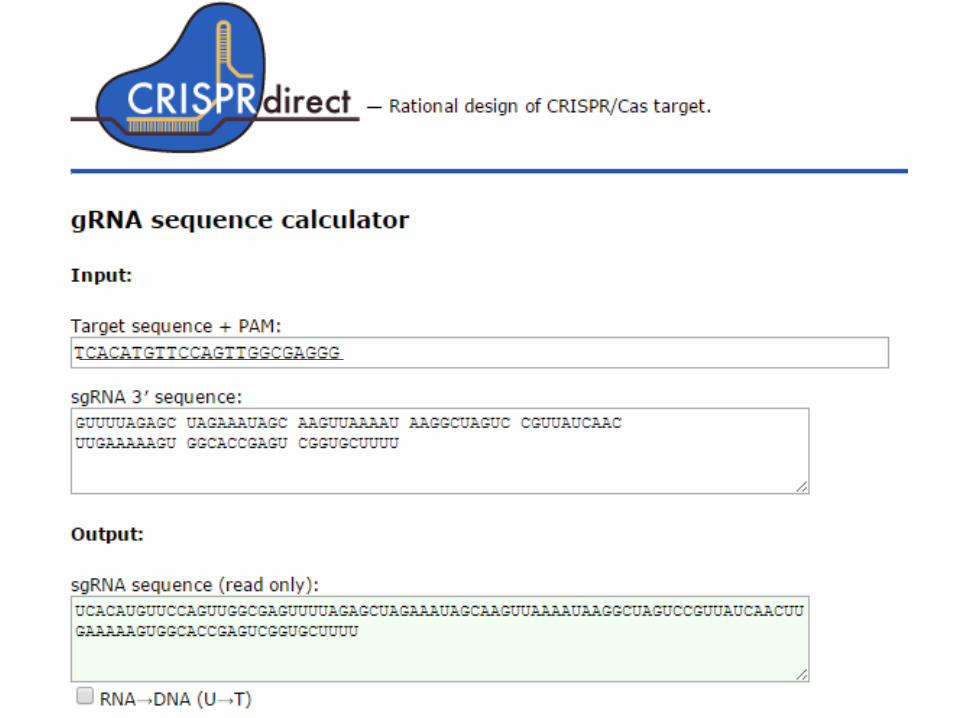
1.2 ガイドRNA(crRNA)の配列のデザイン
• crRNA配列は検索した20nt(PAM配列は除く)の3’側にGuuuuaGAgcuaugcuguuuUGを付加した42nt RNA(図)となる。
• ただし、点変異を導入する場合、切断個所は点変異導入箇所により近い方が好ましいため、候補が限られる。
• tracrRNAはセンター側で用意したものを実験に用いる。
68
crRNA配列 42nt RNA 5’-XXXXXXXXXXXXXXXXXGuuuuaGAgcuaugcuguuuUG PAM配列を除いた20ntを22ntにつなげる tracrRNA 75nt RNA 5’-aaacagcauagcAAGUuaaaaUaaggcuaguccguuaucaacUUGAAaaaGUGgcaccgaGucggu gcUUUuuuU

69
70
71

72

2. オリゴRNA/DNAの注文 • 1.でデザインしたcrRNA(42mer)を依頼者が注文する。 • ドナーDNAを利用する場合、変異導入塩基の両側45ntずつの相同
領域を含む1本鎖オリゴDNA(~90mer)を設計し(図)、センターでの切断活性試験の結果をみてから注文する。
(切断活性が無い場合はcrRNAデザインを変更した方が良い場合があるため) • 現時点で、crRNA(HPLC精製グレード)は株式会社ジーンデザイン
URL: http://www.genedesign.co.jp/で、ドナーDNA(PAGE精製グレード)はIDT URL https://sg.idtdna.com/site (日本代理店MBL)で合成されたもので実験可能なことを確認している。実績があれば、オリゴハウスはどこでも可
73
1本鎖ドナーDNA 45nt+Mutation+45nt 5’-XXXXX XXXXX XXXXX XXXXX XXXXX XXXXX XXXXX XXXXX XXXXX M XXXXX XXXXX XXXXX XXXXX XXXXX XXXXX XXXXX XXXXX XXXXX loxPなど短い配列(40~50nt)を挿入する場合、相同領域として、前後に>60ntずつ繋げる

• 依頼者は、2.のcrRNAと増幅した標的遺伝子座のPCR産物を遺伝子実験センターに送付する
• 遺伝子実験センターにおいて、これらに、CAS9タンパク質, tracrRNAを加え、37℃でインキュベート後、電気泳動により二本鎖切断反応がin vitroで起こることを確認する(図)
• 切断が確認されない場合は、crRNAの塩基配列の再検討を行う。
(-)
Int3
/int5
Int3
Int5
19.3kb 7.7kb 6.2kb 4.3kb 3.5kb
1.9kb 2.7kb
0.9kb
0.4kb
3. ガイドRNA(crRNA)を介したDNA切断活性のin vitroでの確認

5. ゲノム解析による変異個体の確認 • 産子の尾を依頼者に送付する。 • ゲノムDNAを抽出し、PCRによる標的遺伝子領域の増幅後、制限酵素処理や塩
基配列解析、電気泳動などにより、標的遺伝子座での変異を、依頼者が確認する。変異のある個体を、遺伝子実験センターから依頼者へ譲渡する。
伊川 正人 領域融合レビュー, 3, e008 (2014)より
75
4. マウス受精卵前核へのCRISPR/CASベクターの導入 • 依頼者から送られたcrRNA,ドナーDNAに、tracrRNA, Cas9 mRNAを
加えた混合液を、遺伝子実験センターにおいて、C57BL/6受精卵前核へ導入する。
• C57BL/6以外の受精卵で行う場合は要相談

76
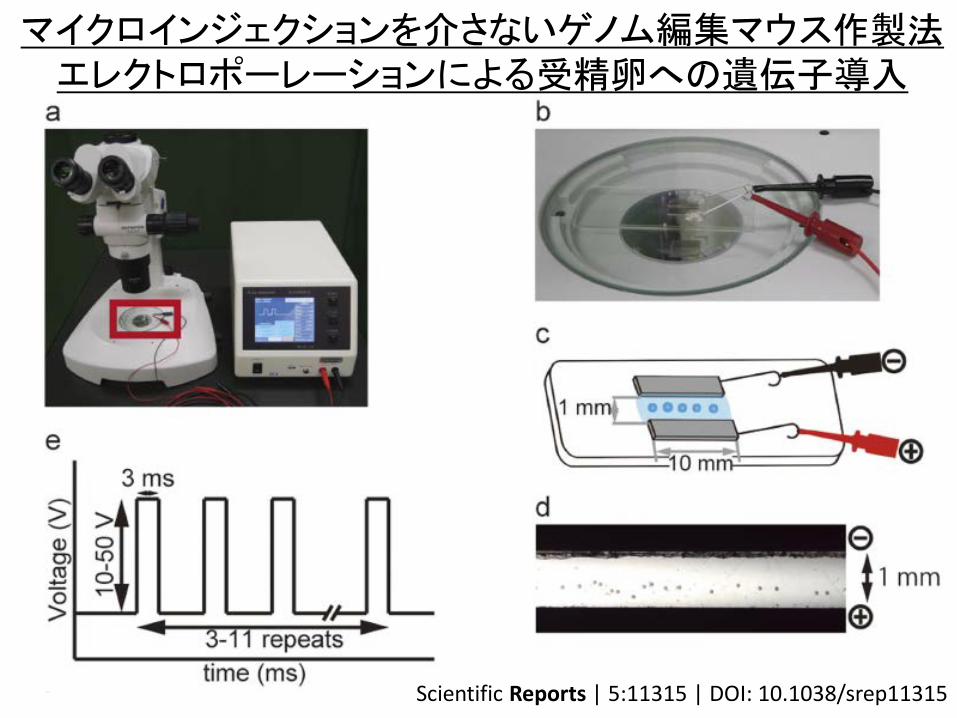
マイクロインジェクションを介さないゲノム編集マウス作製法 エレクトロポーレーションによる受精卵への遺伝子導入
Scientific Reports | 5:11315 | DOI: 10.1038/srep11315
77
マイクロインジェクションを介さないゲノム編集マウス作製法 エレクトロポーレーションによる受精卵への遺伝子導入
Scientific Reports | 5:11315 | DOI: 10.1038/srep11315

78
マイクロインジェクションを介さないゲノム編集マウス作製法 卵管でのエレクトロポーレーションによる遺伝子導入
Scientific Reports | 5:11406 | DOI: 10.1038/srep11406

79 Scientific Reports | 5:11406 | DOI: 10.1038/srep11406
マイクロインジェクションを介さないゲノム編集マウス作製法 卵管でのエレクトロポーレーションによる遺伝子導入

80

81
ゲノム編集受託作製負担金は1系統40万円(ただし、学内は35万円)を予定して、学内規則を策定しました





























