Embed Size (px)
Citation preview

JUNE 1990 A scoeasc tH Nt rAI w ANEN sls rn rsn USA 235
FIRST RECORD OF ASCOGREGARINA TAIWANENSIS (APICOMPLEXA:LECUDINIDAE) IN NORTH AMERICAN AEDES ALBOPICTUS
LEONARD E. MUNSTERMANN eNo DAWN M. WESSON
Department of Binlogiral Sciences, Uniuersity of Notre Dame, Notre Darne, IN 46556
ABSTRACT. Aedes albopictus collected in the East St. Louis, Illinois, area were found infected withthe gregarine protozoan, Aicogregarina taiwanensis. Infection rates varied from 67 to 95% at 4 sites and0 t;10% at 2 others. Lower infection levels were found in Ae. epactius (42Vo) and. Culer restuans (onelarva). Four mosquito species were cross-infected in the laboratory with gregarines isolated from field-collected hosts.. edes atropalpus was 90% susceptible to A. taiwanensis (I00Vo in Ae. ahopictus), withabnormal development and some melanization of trophozoites and gametocysts. In Ae. aegypti and Cx.restuans, the experimental infection was much lower (12-56%) and exhibited abnormalities similar tothe Ae. atropalpus infections. Ascogregarina oocysts recovered from both Ae. aegypti and. Ae. atropalpttshosts were iubsequently infective to Ae. albopictus.ln Ae. triseriattn, A. taiwanensls infection was verylow (25Vo,1-2 trophozoites per larva); gametocysts were not observed nor were infectious oocystsobtained. We conclude lhat A. taiwanensiq newly introduced to the USA with Ae. albopictus, can developin 4 in_digenous mosquito species and can produce deleterious effects in at least 2, Ae aegypti and Ae.atropal,pus,
INTRODUCTION
The recent introduction of the Asian tigermosquito, Aedes albopictus (Skuse), into theWestern Hemisphere (Sprenger and Wuithir-anyagool 1986, Hawley et al. 1987) is viewedwith concern because of the proven capacity ofthis mosquito as a disease vector in Asia. It hasbecome widespread in the central and easternUnited States. with concentrations in severalmajor cites-particularly Houston, New Orle-ans, Kansas City and St. Louis. In spite ofsubstantial research interest in this mosquitoinvolving both ecological and genetic studies(Hawley 1988), there have been no North Amer-ican reports of the parasites that are normallyassociated with Ae. albopictus in its native range.
Gregarine protozoan parasites of Ae. albopic-fus were described from field collections of lar-vae in Taiwan as Ascogregarina (:!sst.rtslt1taiwanensis (Lien and Levine 1980). Althoughthese protozoans have been found in other AsianAe. albopictus populations, notably in India (Ray1933), China (Feng in Vavra 1969) and Malaysia(EIse and Dangsupa 1974), descriptions of theirrange or infection intensity have not been pub-lished. In citations previous to 1980, the Ae.albopictus gegarines were identified as Ascogre-garina (:Gregarina, :lqnpesteria) culicis, fnstdescribed by Ross from Aedes aegypti (Linn.).However, the inability to cross-infect Ae. aegyptiwith the gregarine obtained from Ae. albopictus(Else and Dangsupa 1974) clearly indicated thisgregarine as a distinct species (Lien and Levine1980).
Where larval guts of field populations of othercontainer-breeding Aedes species have been ex-amined, host-specific gregarines have beenfound to be widespread and in high frequency.Ascogregarina cuLicis is common throughout the
range of Ae. aegypti in the United States; A.clarki is found in California tree holes with itshost Ae. sierrensis (Ludlow), and A. barretti hasbeen isolated from many populations of. Ae' tris-eriatus (Say) (Beier and Craig 1985).
In the following, we: 1) document the exist-ence of A. taiwanensis in the United States, 2)describe its apparent pathogenicity in exotichosts, 3) note a disjunct distribution of infectedand uninfected hosts, 4) report how cross-infec-tion experiments have supported our identifi-cation of this gregarine and associated it withits natural host and 5) indicate the potential ofexotic hosts for its perpetuation.
MATERIALS AND METIIODS
Collection sites: All samples (larvae, pupae andadult stages) of Aedes albopictus and associatedgregarine were collected from Madison and St.Clair counties of Illinois in the East St. Louisarea (Fig. 1). This was accomplished as part ofan intensive regional suwey for Ae. albopictusin the summer of 1988. Table 1 lists habitatparameters of sites (all with used tires) fromwhich larvae were collected and inspected forgregarines. Well-shaded tire piles tended to havemore water-containing tires and larger popula-tions of mosquito larvae, similar to the descrip-tions of Beier et al. (1983); consequently, thesesites were sampled more intensively. Adultswere sampled both by landing-biting collectionsand with a battery-powered suction aspirator(Nasci 1981). Efforts were concentrated at Io-cations where the vegetation was thick and adultmosquito density was high.
Dissection: Dissections of larvae and pupaeproceeded as follows: 1) Thorax and head wereseparated from the abdomen on a microscopeslide with a pair of minuten pins in orangewood

236 JounNer, oF THE AvsRrceN Moseurro CorrRor, Assocrarrorv Vor,. 6, No. 2
GRANINTE
ia ' f ';N,
Fig. 1. Location of Aedes albopictus sites in the East St. Louis area, showingthose positive for Ascogregarinataiwanensis. Sites unexamined for Ascogregarina had few larval habitats or had very low larval populations.(The Ae. albopictus sites were numbered consecutively within each county as they were discovered.)
sticks, while viewing through a dissecting micro-scope. 2) Contents of the abdomen were gentlyteased out through the anterior of the abdomenby placing pressure at the posterior abdomenand gradually pressing anteriorly. 3) The Sutcontents enclosed in the peritrophic membranewere pulled from inside the isolated gut. 4) Head,thorax and abdominal remains were pressed un-der one coverslip; gut and gut contents werecovered with second coverslip. 5) Larvae wereidentified at 160x with a Leitz Dialux(e micro-scope. 6) Guts and contents were scanned forgregarines at 40x, then checked at 160x withboth bright-field and polarized light optics. Thehighly birefringent amylopectin granules of theprotozoan were easily seen against a dark field.Even early (globular) stages of the gregarinewere distinguishable from tissue obstructionsand other granulated gut cells.
Dissection fluid under both coverslips wasexamined. Normally, few gregarines were foundoutside the isolated gut. Displacement of tro-phozoites and gamonts was monitored throughthe dissecting microscope during dissection.Movement from original positions was minimal,although increased coverslip pressure provedmore disruptive.
Source of mosquito and gregarine strains: Theorigin and dates of collection of the laboratory
strains used in the cross-infection experimentswere as follows: l) Ae. albopictus-3 strains,STCLAIR-2 and MADISON-I from East St.Louis, Illinois, area (Table 1, Fig. 1), summer of1988; NEW-ORLEANS-87 from Jefferson Par-ish, New Orleans, Louisiana, June 1987. 2) Ae.aegypti-2 strains, WACO from Waco, Texas,1987; ROCK originally from the Rockefeller In-stitute, colonized for more than 50 years. 3) Ae.atropalpus (Coquillett)-SG strain, from St. Jo-seph Co., Indiana, June 1985. 4) Ae. triseriatus-WALTON strain, from St. Joseph Co., Indiana,June 1969. 5) Culex restuans Theobald-field-collected as egg rafts from tires in St. JosephCo., Indiana, September 1988.
Two strains of Ascogregarina taiw anensis wercused in the experimental infections. ESL wasisolated from East St. Louis Ae. albopictus col-Iected in the summer of 1988. The second, MA-LAY, originated from Ae. albopictus collected inKuala Lampur, Malaysia, by W. Hawley, De-cember 1988. A second gregarine species, A.barretti from Ae. triseriatus, was isolated fromcollections in St. Joseph Co., Indiana, 1987.
Oocyst isolation and mosquito infection: Fol-Iowing the dissection of field collected material,infected adult mosquito carcasses were pooledaccording to mosquito species. The method ofoocyst isolation has been previously described

JUNE 1990 ASCOGREGABINA TAIWANENSTS IN THE USA 237
Table 1. Location and qualitative parameters oftire sites in the East St. Louis (Illinois) area from whichlarvae were dissected fot Ascogregarina.
General Mosquito speciesSite code*
Latitude/Iongitude Locality env[onment identified**
Mad,i.sonCountyMADISON-1
MADISON.5
St. Clair CountySTCLAIR-2
STCLAIR.4
STCLAIR-5
STCLAIR-6
38'37'�42.8"N90"08'4r.5"w
38'39'�48.4"N90"08'�47.5"w
38'38'�08.0"N90"08'�13.3"w
38"38'�16.9"N90'09'11.3"w
38'38'�47.8'�N90"09'�35.3"w
38'37'�58.9"N90"08'27.6"W
Eagle Park Rd.(Watson's Tire)
Eagle Park Rd. N.
9th and LynchAve.
National stock-yards (1st andLast Chance)
National City(roadside ditch)
9th and BowmanAve.
Partially shaded toopen sun, thou-sands of tires in 2Iong, 250-m par-allel heaps.
5 0 x 1 2 x 2 m p a r -tially shaded pile,several hundredtires, containingmuch organic de-bris.
Shaded by largetrees along free-way; 75-100 tires,weedy.
Shaded by largetrees, severalhundred tires,with highly or-ganic contents.
Ten tires, discardedcontainers, weedyarea.
Open lot, severaltire piles of 10-40; weedy, over-grown with vines.
ALB, EPA, TRI,PIP, SIG, RUT
ALB, EPA, TER,PUN
ALB, EPA, PIP,PUN
ALB, EPA, TRI,PIP
ALB
ALB, EPA, PIP,RES, TER, PUN
* The numbers for the sites in each county match those shown in Fig' 1'** The 3-letter acronyms correspond to the first 3 letters of the species name: ALB : Aed'es albopictw;E�PA
: Ae. epactiusi PIP : Culex prpiens; PUN : Anophelcs punctipennis; R,F,S : C.r. restttansi RUT : Toxorhyn-
chites iutihu; SIG : Orthopodomyia signifero; TER : Cx. territans; TRI : Ae. triseriatus.
by Beier and Craig (1985); this was effective forconcentrating oocysts but not for quantificationof oocysts per infected individual. Mosquitostrains were infected by administering doses ofapproximately 1,000 oocysts per first instarlarva in each experimental container. As con-trols, synchronous cohorts of uninfected larvaeof each mosquito strain and species were rearedand dissected. The ESL strain of A. taiwanensiswas introduced to each ofthe species and strainslisted above. The MALAY strain of A. taiwanen-sis was provided to the STCLAIR-2 strain of Ae.albopictus as an additional comparison of host-parasite specificity. Finally, infection of Cx. res-tuans was attempted with 2 gregarine species,A. taiwanensis ESL and A. barcetti.
A second set of infections was designed toverify the identification of field-collected A. tai-wanensis and to estimate oocyst production andviability in both natural and exotic hosts.Oocysts were isolated from 3 field-collected mos-quito species and one laboratory-infected spe-cies. The infected Ae. albopictus, Ae. epactius
Dyar and Knab andAe. triseriatuswere collectedfrom site STCLAIR-4 (Fig. 1, Table 1). Thegregarines in Ae. triseriallus were identified mor-phologically as A. barretti, while those from theother 2 species corresponded to A. taiwanensis'"Ihe Ae. aegypti WACO strain was infected withoocysts isolated from field samples of adult Ae.albopictus from the STCLAIR-4 site. Oocystsfrom each of the four mosquito species werepresented separately to 50 first instar larvaeeach of Ae. albopictus STCLAIR-2 strain andAe. triseriatus WALTON strain. Viability of theoocysts was determined by dissecting 8 fourthinstar larvae from each host-parasite combina-tion. Presence of trophozoites in a larva waspositive evidence that the source mosquito hadpassed viable oocysts.
RESULTS
Ascogregarina identification: The morphologyof the Ascogregarina found in East St. LouisAedes albopictus corresponded well to the de-

238 JounNel oF THE AunRrceN Moseurro ColrrRor, Assocrnrrox V o L . 6 , N o . 2
Table 2. Occurrence of Ascogregarino in mosquito species collected from tires in the East St. Louis area.
Mosquito species No. dissectedPercent positive
(Ascogre gar ina species)
Aedes albopictusAe. epactiusAe. triseriatusCulex restuansCx. pipiensCx. tenitansO r thopo do my ia s ignif e r aA nophe le s p unc tipennis
36667t 21
J D
3I
7l (A. taiwanensis)42 (A. taiwanensis)58 (A. barretti)l+l (A. taiwanensis)
0 -0 -0 -0 -
scription of r,he protozoan by Lien and Levine(1980). Those authors identified the parasite inTaiwan Ae. albopictu.s as different from A. cuLi-cls, based on morphology and host specificity.Our designation of the gregarine as A. taiwanen-sis was based both on the morphological char-acterization of Lien and Levine and the cross-infection experiments reported below.
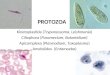
Distribution of gregarines by species and bysite: A summary of all gut dissections of field-collected mosquitoes from the East St. Louissites is shown in Table 2. The mosquito mostcommonly infected was Ae. albopictus with A.taiwanensis (Fig. 2 A-C), followed by Ae. tris-eriatus and A. barretti (Fig. 2D) and Ae. epactiuswith A. taiwanensis (Fig. 3 E-F). A single Culexrestuans larva was found with a trophozoite withthe morphologio of A. taiwanensis.
In the survey of Ae. albopictrzs in the East St.Louis area, A. taiwanensis parasites were foundin 5 of 6 sites (Fig. 1, Table 3). In St. Clair Co.,4 sites were positive, with 67-95% of the mos-quitoes infected. In larvae, the mean infectionintensity among the 4 sites ranged from 22-98trophozoites per larva, with a maximum of 299noted in one larva. Of 10 larvae dissected fromthe MADISON-5 site (in Madison Co.), onelarva was found with a single trophozoite. None-theless, MADISON-5 was denoted in Fig. 1 as a"negative site" because of this unusually lowrate. All other mosquitoes examined from Mad-ison Co. were negative for A. taiwanensis. Asseen in Fig. 1, the infected and uninfected siteswere in close proximity, with the closest pairseparated by no more than 2 km.
Pupae and adults ofAe. epactius from 2 sites,STCLAIR-4 and MADISON-1, also yielded dis-tinctly different results. While 15 of 16 dissec-tions were positive for A. taiwanensis atSTCLAIR-4, none of 23 were infected at MAD-ISON-I. Three of those infected at STCLAIR-4 were dead or moribund pupae with very heavyinfections (Fig. 3E). The adults had varyingmanifestations and intensities of infections.Some had very heavy infections, usually withlarge numbers of gamonts or early gametocysts
melanized in the Malpighian tubes (Fig. 3F).Others had light infections with low numbers o{stunted trophozoites in the midgut, and st.others presented only a few abnormally largitrophozoites. Although the larvae of Ae. epactiuwere quite common in tires, the adults werecollected more rarely. Compared with Ae. albo-pictus,the Ae. epactius adults made up only 4.6%(6511414) at STCLAIR-4 and0.3% (3/1,158) atMADISON-1 of the respective samples.
E xpe rimental cros s - inf ections; The referencestrains of Ae. albopicfus showed a 100% infectionand very large numbers of trophozoites (up toI,2I0), although 2 individuals carried as few as2 (Table 4). In the 2 strains of Ae. aegyptj, boththe infection rate and number of trophozoiteswere lower. The pattern of infection was quiteaberrant, including oversized trophozoites (Fig.3C) and, rarely, gametocysts in the midgut. Ga-monts (the motile stage in pupae) were fre-quently melanized in the Malpighian tubes, es-pecially in the dead pupae. In many, a Iargebolus of both melanized and normal gamontswas present in the posterior midgut near theopening of the Malpighian tubes. Mortality inA. taiwanensls-infected Ae. aegypti seemed.quitehigh, although unfortunately these trials werenot designed for quantitative comparisons.
Variation in infectivity in Ae. albopiclius wasfound with respect to both host strain origin andgregarine strain. The ESL strain of. A. taiwanen-sis from East St. Louis produced high infectionsin the 3 USA strains of Ae. albopictus (Table 4).The MADISON-I and NEW-ORLEANS-87strains showed much higher mean trophozoiteload (409 and 499, respectively) than did theSTCLAIR-2 strain (86), although the rangesoverlapped substantially. The MALAY (fromMalaysia) strain ofA. taiwanensis was also quiteinfective to the STCLAIR-2 mosquitoes. In 4larval dissections, the number of trophozoitesranged from 126 to 218. Pupal dissectionsshowed 3 of 3 infected; 2 pupae had all 5 Mal-pighian tubes infected with 134 and 144 totalgametocysts, respectively. These gregarines ap-peared normal in all developmental stages.



JUNE 1990 AscocnEGARINA TAIqANENSIS ru'rHp USA
Table 3. Ascogregarina infection intensities in Aed,es albopictus as distributed among collecting sites in theEast St. Louis area.
Trophozoites in larvae
247
County-srte
No.dissected*
Infected% No. larvae Mean Range
STCLAIR-2STCLAIR.4STCLAIR-5STCLAIR-6MADISON-1MADISON.2
899567950
l + l
70135
q
197610
2629470I
525498z z
I
4-2995-230t q q n
1-48
* Includes larvae, pupae and adults.
Table 4. Experimental infections by Ascogregarina taiwanensis from East St. Louis in strains of Aedes
itiipirti,An. aegypti and Ae. atripalpui. The Ae. triseriatus bioassay data (Table 5) are included for
comParison.
Larvae dissected No. of trophozoites
Strain No. % positive Mean* Range
Aedes aeglptiROCKwAco
Aedes albopictusSTCLAIR.2MADISON-1NEW.ORLEANS.ST
Aedes atropalpusSG
Aedes triseriatusWALTON
16
855
56.3t2.0
r00.0100.0100.0
90.1
25.0
5-1105-24
,_r1'l72-t,210
q n o q
t4-225
l-2
40l7
86409449
85
1.5
1 1**
16* Calculations include only infected larvae.*+ Eight ofthe 10 infected larvae were moribund'
Table 5. Bioassay for viability of oocysts using Aedes albopicttts STCLAIR-2 strain and Ae. triseriatusWALTON strain. A. Ascogregarina oocysts isolated from East St. Louis field samples of Aedes albopictus(ALB), Ae. epactius (EPAj and ae. biseriatus (TRI). B. Ascogregarina taiwanen'sis oocysts from East St'
Louis passaged through Ae. aegypti WACO (AEG)'
Oocyst origin Bioassay hostNo. Percent Range
dissected infected (trophs/larva)
A. Eost St. Louis field oocystsALB*ALB*EPA*EPA*TRI**TRI+*
B. Laboratory passagedoocysts
AEG*AEG*
Ae. albopictusAe, triseriatusAe. albopictusAe. triserintusAe. albopictusAe. triseriatus
Ae. albopictusAe. triseriatus
100
10000
88
250
8168888
88
35-3111-2
10-12100
8-175
2-60
* Infections were identified as Ascogregarina taiwanensis from morphology of trophozoites in larval dissec-tions.
*+ Infections identified as A. barretti.
to Ae. albopictus. Aedes aegypti also produced alow level of viable A. taiwanensis oocysts capableof infecting Ae. albopictus. Although Ae. triser-iofus was slightly susceptible to A. taiwanensisftom Ae. albopictus, oocysts from neither Ae.epactius nor Ae. aegypti resulted in discernibleinfection in Ae. triseriatlus larvae.
DISCUSSION
Ascogregarina taiwanensis is probably presentin many North American populations of Ae.albopictus. Subsequent to its discovery in EastSt. Louis, one of us (D.M.W.) identified thisparasite/mosquito combination at 3 additionalsites-Chicago, Illinois, Polk County, Florida,

242 JouRr)rel, oF THE AlrnnrcnN Mosourro CoNrRor, Assocrarror\ V o r . 6 , N o . 2
and Potosi, Missouri. Nonetheless, the gregarinein East St. Louis is not uniformly distributed inspite of the close proximity of the sites (Fie. 1,Table 3). This is probably due to "founder ef-fect," where the populations at each site werefounded by mosquitoes of a different origin andwith very different levels of gregarine infection.Subsequently, the infected mosquitoes did notdisperse into the Madison Co. sites from thesites in St. Clair Co., only 2 to 3 km distant.
Low dispersal capability of Ae. albopicfr.m hasbeen measured in several field release studies(Hawley 1988). Electrophoretic data from sam-ples within and between cities in the UnitedStates have indicated both founder effectsamong cities and lack of genetic similarity be-tween adjacent tire sites (Black et al. 1988).Origin of tires (local or purposely transported)and reasons for tire placement (discards on rub-bish h6ap, tires for resale, tires for processing)seem to be more important for dispersal thanmovement of the mosquito through the environ-ment. If this is the case, then finding of disjunctdistributions of the Ascogregarina is not surpris-ing.
Although the gregarine-positive and negativepopulations of Aedes albopictus are probably ofdifferent geographic origins, how can the gregar-ine be absent from any site, given its high rateof infection in the St. Clair Co. sites and theintense level of induced infection in mosquitoesfrom the MADISON-1 site? Two scenarios canbe constructed. The first is as follows: If unin-fected individuals are more reproductively fitthan infected, uninfected individuals can ex-pand throughout a site at a greater rate than theinfected individuals. In new colonizations thegregarine may not be capable of spreadingquickly through a rapidly expanding host popu-Iation. If sampling of the population occurs dur-ing the expansion phase, the gregarine may es-cape detection. Data from other mosquito-gre-garine systems do not support this interpre-tation-generally, the gregarine has little effecton its natural host (Beier and Craig 1985). Forthe A. taiwanensis-Ae. albopictus association,however, some larval and pupal mortality seemsto occur in laboratory infections (D. M. Wesson,unpublished observations). The relevance of thislaboratory mortality to effects on reproductivefitness in the field remains to be investigated.
Alternatively, the uninfected population mayconsist of individuals genetically resistant toparasitization. As the gregarine oocysts are in-troduced to resistant mosquitoes, they are neu-tralized in some manner. However, for the Mad-ison Co. populations, this explanation is un-likely. The MADISON-I strain, derived fromthe uninfected site, proved very receptive tolaboratory infections.
In the field populations where it was found,A. taiwanensls was highly infectious. Infectionrates ranged from 67 to 95%, compared to nat-ural infection levels of 60% fot A. barcetti (Beierand Harris, 1983),70% for A. culicis (Barrett etal. 1971) and 28% for A. clarhi (Sanders andPoinar 1973). In addition, A. taiwanensls has ademonstrated capacity to pass through at leastone generation in non-Ae. aLbopictus mosquitohosts. This has not been demonstrated in othermosquito gregarines and lends a new dimensionto its survival potential. The A. taiwanensisdescribed from Taiwan apparently did not showthis capability. Even though 48% of Ae. aegyptilarvae carried trophozoite stages in laboratoryinfections, no infectious oocysts were produced(Lien and Levine 1980). Since the U.S. Ae. al-bopictus are thought to have been introducedfrom mainland Japan (Hawley et al. 1987), theassociated gregarine may be quite different fromthe Taiwan gregarine with respect to its devel-opmental capabilities in exotic hosts.
The extent to which A. taiwanensis has dele-terious effects on Ae. albopictus and sympatriccontainer-breeding mosquitoes is still unknown.Our initial observations indicate that (at leastin the exotic mosquito hosts Ae. aegypti and Ae.epactius) A. taiwanensls may have a significanteffect on mosouito survival.
ACKNOWLEDGMENTS
We thank J. F. Cully, Jr., and G. B. Craig, Jr.,for reading the manuscript and offering sugges-tions for improvement. We also thank othermembers of the Vector Biology Laboratory-W.Hawley, C. Pumpuni, S. Hanson, T. Streit andM. Niebylski-for assisting in confirmatoryfield collections. This work was supported byNIH grant AI -02753 and NIH Training Program2 T32 AI-07030.
REFERENCES CITED
Barrett, Jr., W. L., F. M. Miller and J. W. Kliewer.1971. Distribution in Texas of Lanhesteria culicis(Ross), a parasite of Aedes aeeypti (L.). Mosq. News3l:23-27.
Beier, J. C. and G. B. Craig, Jr. 1985 Gregarine para-sites of mosquitoes, pp. 167-184. /n: M. Laird (ed.),Integrated mosquito control methodologies, vol. 2.Academic Press, London.
Beier, J. C. and C. Harris. 1983. A scogregarina barretti(Sporozoa: Diplocystidae) infections in natural pop-ulations of Aedes triseriatus (Diptera: Culicidae). J.Parasitol. 69:430-431.
Beier. J. C.. M. Travis. C. Patricoski and J. Kranz-

AscoensceruNt rerwewovsrs in the USA 243
felder. 1983. Habitat segregation among larval mos-quitoes (Diptera: Culicidae) in tire yards in Indiana,USA. J. Med. Entomol. 20:76-80.
Black IV, W. C., J. A. Ferrari, K. S. Rai and D.Sprenger. 1988. Breeding structure of a colonizingspecies: Aedes ahopictus (Skuse) in the UnitedStates. Heredity 60:173-181.
Else, J. G. and P. Dangsupa. 1974. Lanhesterin, agregarine protozoan, previously unreported in mos-quitoes of Malaysia. Southeast Asian J. Trop. Med.Public Health 5:454.
Hawley, W. A. 1988. The biology of Aed'es ahopictw.J. Am. Mosq. Control Assoc. 4(Suppl. 1): 1-40.
Hawley, W. A., P. Reiter, R. S. Copeland, C' B. Pum-puni and G. B. Craig, Jr. 1987. Aedes albopictus inNorth America: probable introduction in used tiresfrom northern Asia. Science 236:1114-1116.
Lien, S. M. and N. D. Levine. 1980. Three new speciesof Ascocystis (Apicomplexa, Lecudinidae) from mos-
quitoes. J. Protozool. 27 :147 -751.Nasci, R. S. 1981. A lightweight battery-powered as-
pirator for collecting resting mosquitoes in the field.Mosq. News 41:808-811.
Ray, H. 1933. On the gregarine Lanhesteria culicis(Ross) in the mosquito A edes (Stegomyia) albopictusSkuse. Parasitology 25:392 396.
Sanders. R. D.. and G. O. Poinar, Jr. 19?3. Finestructure and life cycle of Lanhesteria clarhi sp. n-(Sporozoa: Eugregarinida) parasitic in the mosquitoAedes sierrensis (Ludlow). J. Protozool. 20:594-602-
Sprenger, D. and T. Wuithiranyagool. 1986. The dis-covery and distribution of Aedes albopictusinHatrisCounty, Texas. J. Am. Mosq. Control Assoc. 2:217-279.
Vavra, J. 1969. Lanhesteria barretti n. sp. (Eugregar-inida, Diplocystidae), a parasite of the mosquitoAedes triseriatus (Say), and a review of the genusLanhesteria Mingazzini. J. Protozool. 16:546-570.