Embed Size (px)
Citation preview
CORRESPONDENCE
Follow-up and Extension of a Prior Genome-wideAssociation Study of Posttraumatic Stress Disorder:Gene � Environment Associations and StructuralMagnetic Resonance Imaging in a Highly TraumatizedAfrican-American Civilian PopulationTo the Editor:
Agenome-wide significant single nucleotide polymorphism(SNP) was recently reported (p ¼ 3.97 � 10�8) on chromo-some 7p12, rs406001, in a genome-wide association
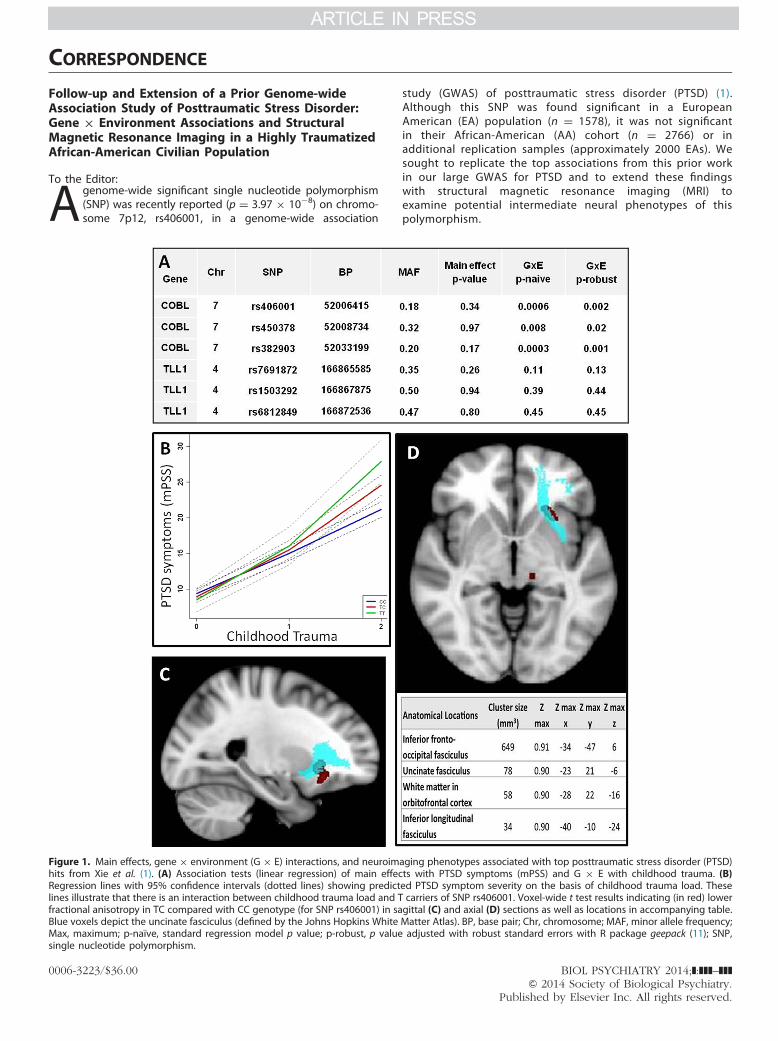
Figure 1. Main effects, gene � environment (G � E) interactions, and neuroimhits from Xie et al. (1). (A) Association tests (linear regression) of main effecRegression lines with 95% confidence intervals (dotted lines) showing predictlines illustrate that there is an interaction between childhood trauma load and Tfractional anisotropy in TC compared with CC genotype (for SNP rs406001) in saBlue voxels depict the uncinate fasciculus (defined by the Johns Hopkins WhiteMax, maximum; p-naïve, standard regression model p value; p-robust, p valuesingle nucleotide polymorphism.
0006-3223/$36.00
study (GWAS) of posttraumatic stress disorder (PTSD) (1).Although this SNP was found significant in a EuropeanAmerican (EA) population (n ¼ 1578), it was not significantin their African-American (AA) cohort (n ¼ 2766) or inadditional replication samples (approximately 2000 EAs). Wesought to replicate the top associations from this prior workin our large GWAS for PTSD and to extend these findingswith structural magnetic resonance imaging (MRI) toexamine potential intermediate neural phenotypes of thispolymorphism.
aging phenotypes associated with top posttraumatic stress disorder (PTSD)ts with PTSD symptoms (mPSS) and G � E with childhood trauma. (B)ed PTSD symptom severity on the basis of childhood trauma load. Thesecarriers of SNP rs406001. Voxel-wide t test results indicating (in red) lowergittal (C) and axial (D) sections as well as locations in accompanying table.Matter Atlas). BP, base pair; Chr, chromosome; MAF, minor allele frequency;adjusted with robust standard errors with R package geepack (11); SNP,
BIOL PSYCHIATRY 2014;]:]]]–]]]& 2014 Society of Biological Psychiatry.
Published by Elsevier Inc. All rights reserved.

e2 BIOL PSYCHIATRY 2014;]:]]]–]]] Correspondence
Our sample consisted of study participants from GradyMemorial Hospital (Atlanta, Georgia) as part of the Grady TraumaProject. As previously shown (2), these participants were adult,primarily female, highly traumatized, impoverished, primarily AA,with high rates of PTSD. We assessed current PTSD symptoms inover 3000 AA subjects with GWAS data with the PTSD symptomscale (mPSS) (3). Other phenotype measurements included in thedata collection were Childhood Trauma Questionnaire (4) asa measure of childhood trauma; and Beck Depression Inventory(5) as a measure of depressive symptoms.
The DNA was extracted from saliva in Oragene collection vials(DNA Genotek, Ontario, Canada) with the DNAdvance kit (Beck-man Coulter Genomics, Danvers, Massachusetts). The top SNPsfrom Xie et al. (1) were genotyped on the Illumina Omni1-Quadmicroarray, and statistical genetic analyses were performed inPLINK (6) and R statistical software (http://www.r-project.org/).The genotyping call rate for the SNPs was �.99, and thedistribution did not depart from Hardy-Weinberg equilibrium(p � .1). To reduce potential bias by population stratification,we restricted the analysis to 3215 AA subjects who clusteredtogether on principal component analysis. With linear regression,we found that the top SNPs reported in Xie et al. (1) did notreplicate as main effects in associations with PTSD symptoms inour study (Figure 1A); however, we found a significant genotype� environment interaction (G � E) with childhood trauma in theirtop SNP rs406001 (n ¼ 3076, t ¼ 14.98, p ¼ .0006) (Figure 1B).In fact, all three tested SNPs near the gene COBL had significantG � E interactions with childhood trauma in our AA population.
To examine the potential effects of this genotype on brainstructure, we used MRI data from a subset of subjects on whomwe had collected imaging data. Diffusion-weighted images (iso-tropically distributed along 60 directions; 39 � 2.5-mm-thick axialslices, matrix ¼ 128 � 128, field of view ¼ 220 � 220 mm, voxelsize ¼ 1.72 � 1.72 � 2.5 mm) were acquired from AA womenbetween 18 and 62 years of age (n ¼ 66, mean age ¼ 38.5, SD ¼12.7) with parameters previously described (7). All image proces-sing and analysis were conducted with FMRIB Software Library(FSL version 4.1; http://fsl.fmrib.ox.ac.uk/fsl/fslwiki/) (8), similar to aprevious study (7). Tract-based spatial statistics (version 1.2) wereused to examine associations between rs406001 genotype, child-hood trauma, and fractional anisotropy (FA) across the entirebrain. There were no significant main effects of childhood traumaexposure or G � E interactions with regard to FA values (p � .1,family-wise error corrected threshold). However, we observed amain effect of genotype. Compared with the CC genotype, carriersof the risk (T) allele demonstrated significantly lower FA in thefollowing areas: left inferior-fronto occipital fasciculus; left inferiorlongitudinal fasciculus; white matter in the left frontal orbitalcortex; and the left uncinate fasciculus (p � .1, family-wiseerrorcorrected) (Figure 1C,D). Univariate analysis of covariance resultsrevealed that, after statistically controlling for age, current PTSD(mPSS), and depression symptoms (Beck Depression Inventory),the relationships between rs406001 genotype and FA within theseclusters remained statistically significant (all p values � .01).
In summary, we found a significant effect of rs406001 (andother SNPs in linkage disequilibrium) interacting with childhoodtrauma exposure in a large AA cohort. These data suggest thatexamining trauma exposure in combination with genetic riskmight allow more robust integration across population cohorts,because the original study found no main effect in AA subjects.
The rs406001 SNP is intergenic with no known function;however, the closest gene is COBL, which might be relatedto actin polymerization and neuronal development and function.
www.sobp.org/journal
Our brain imaging findings indicate that risk allele carriersdemonstrate alterations in white matter integrity in brain regionsassociated with emotion processing; in particular, the uncinatefasciculus serves as a primary connection between the amygdalaand ventral aspects of the prefrontal cortex, which is thought toplay a role in extinction of learned fear (9,10). Thus, it is possiblethat risk allele carriers are more vulnerable to the development ofanxious psychopathology via decrements in these white matterpathways. These data serve as a partial replication and extensionof the first large GWAS for PTSD and suggest a putative whitematter intermediate phenotype that might underlie PTSD risk.
Lynn M. Almlia
Amita Srivastavac
Negar Fania
Kimberly Kerleye
Kristina B. Mercera
Hao Fengb
Bekh Bradleya,d
Kerry J. Resslera,e*
aDepartment of Psychiatry and Behavioral Sciences; bDepartment of HumanGenetics, Emory University School of Medicine; cMorehouse School of Medicine;dAtlanta VA Medical Center, Atlanta, Georgia; and the eHoward Hughes MedicalInstitute, Chevy Chase, Maryland..
*Corresponding author E-mail: [email protected].
Authors LMA and AS contributed equally to this work.This work was primarily supported by the National Institutes of Mental Health
(MH071537 and MH096764 to KJR). Support was also received from Emory and GradyMemorial Hospital General Clinical Research Center, National Institutes of HealthNational Centers for Research Resources (M01RR00039), and Howard Hughes MedicalInstitute (KJR). We would like to thank staff and participants from the Grady TraumaProject for their time and effort in supporting this research.
The authors report no biomedical financial interests or potential conflicts of interest.
1. Xie P, Kranzler HR, Yang C, Zhao H, Farrer LA, Gelernter J (2013):Genome-wide association study identifies new susceptibility loci forposttraumatic stress disorder. Biol Psychiatry 74:656–663.
2. Gillespie CF, Bradley B, Mercer K, Smith AK, Conneely K, Gapen M, et al.(2009): Trauma exposure and stress-related disorders in inner cityprimary care patients. Gen Hosp Psychiatry 31:505–514.
3. Falsetti SA, Resnick HS, Resick PA, Kilpatrick D (1993): The ModifiedPTSD Symptom Scale: A brief self-report measure of posttraumaticstress disorder. The Behavior Therapist 16:161–162.
4. Bernstein DP, Fink L (1998): Childhood Trauma Questionnaire Manual.San Antonio, TX: Psychological Corporation.
5. Beck AT, Ward CH, Mendelson M, Mock J, Erbaugh J (1961): Aninventory for measuring depression. Arch Gen Psychiatry 4:561–571.
6. Purcell S, Neale B, Todd-Brown K, Thomas L, Ferreira MA, Bender D,et al. (2007): PLINK: A tool set for whole-genome association andpopulation-based linkage analyses. Am J Hum Genet 81:559–575.
7. Fani N, King TZ, Jovanovic T, Glover EM, Bradley B, Choi K, et al. (2012):White matter integrity in highly traumatized adults with and withoutpost-traumatic stress disorder. Neuropsychopharmacology 37:2740–2746.
8. Smith SM, Jenkinson M, Woolrich MW, Beckmann CF, Behrens TE,Johansen-Berg H, et al. (2004): Advances in functional and structuralMR image analysis and implementation as FSL. Neuroimage 23(suppl1):S208–S219.
9. Phelps EA, Delgado MR, Nearing KI, LeDoux JE (2004): Extinction learningin humans: Role of the amygdala and vmPFC. Neuron 43:897–905.
10. Maren S, Phan KL, Liberzon I (2013): The contextual brain: implicationsfor fear conditioning, extinction and psychopathology. Nat RevNeurosci 14:417–428.
11. Halekoh U, Højsgaard S, Yan J (2006): The R package geepack forgeneralized estimating equations. J Stat Softw 15:1–11.
http://dx.doi.org/10.1016/j.biopsych.2014.01.017







![OEF/OIF/OND Veterans Components of Self-Reported Sleep ... · While the associations between psychological distress (e.g., posttraumatic stress disorder [PTSD], depression) and sleep](https://img.pdfslide.net/doc/110x75/5c01681a09d3f2fa038c99a8/oefoifond-veterans-components-of-self-reported-sleep-while-the-associations.jpg)





















