Embed Size (px)
Citation preview

Proc. Natl. Acad. Sci. USAVol. 84, pp. 975-979, February 1987Biophysics
Force generation by muscle fibers in rigor: A lasertemperature-jump study
(rabbit psoas muscle/force from melting in subfragment 2 hinge/musele contraction/helix-coil model)
JULIEN S. DAVIS AND WILLIAM F. HARRINGTONDepartment of Biology, The Johns Hopkins University, Baltimore, MD 21218
Contributed by William F. Harrington, October 27, 1986
ABSTRACT A clear prediction of the helix-coil model forforce generation in muscle is that force should be producedwhen the equilibrium (helix-coil) of a rigor (or activated) fiberis perturbed by a temperature jump near the melting temper-ature of the light meromyosin/heavy meromyosin hinge. Aninfrared, iodine-photodissociation laser was used to heat thefibers by ==50C in under 1 ps. Under ionic conditions whererigor bridges are predominantly associated with the thickfilament backbone, an abrupt drop in tension typical ofnormalthermoelastic expansion was seen. A similar response wasobserved below 41'C for thick rilament-released rigor bridges.Above this temperature, a rubber-like thermoelastic responsewas obtained typical of a helix-coil transition. At temperaturesnear 50TC, the amount of force generated by a rigor fiber waslarge and comparable to that seen for an activated fiber at 50C.The relaxation spectra of force generation obtained for bothsystems (rigor *and activated) show a step change followed by abiexponential kinetic process. The reciprocal relaxation timesand amplitudes for these individual processes in activated andrigor fibers differ only by factors of24. Force generation in therigor muscle appears to arise from melting in the subfragment2 hinge region of the myosin molecule since binding ofsubfragment 2 to the thick filament backbone inhibits forceproduction. No significant force generation was observedfollowing temperature jumps of relaxed fibers.
It is likely that force generation in skeletal muscle resultsfrom a structural change in myosin crossbridge [the heavymeromyosin (HMM) region of myosin] while it is attached toactin in an active bridge cycle. One suggested mechanism (1,2) is that the structural transition is confined to the myosinhead [subfragment 1 (S-1) subunit], resulting in a change inthe effective angle of this moiety while it is bound to actin.Another proposal (3, 4) is that a part of the a-helicalsubfragment 2 (S-2) segment of the crossbridge [the lightmeromyosin (LMM)/HMM hinge domain] undergoes melt-ing to random-coil when the actin-attached bridge swingsaway from the stabilizing environment of the thick filamentsurface. This process induces shortening in S-2 and producesthe force required to slide the thin filaments relative to thethick filaments during active contraction.Force generation by polymers, like the a-helical coiled-coil
of the hinge, that undergo a transition from an extendedsemirigid form (helical in structure) to a highly flexiblerandom-coil form, buffeted by thermal (Brownian) motion, isa well-documented phenomenon. The heating ofa rubber-likethermoelastic polymer (as the above is described) results incontraction; expansion results when a substance with normalthermoelasticity is heated. To date, only normal thermoelas-ticity has been observed in muscle and not the rubber-likethermoelasticity expected of the helix-coil transition (see ref.
5). The maximum force expected from a helix-coil transitionin an a-helical (coiled-coil) structure the size of the hingeregion is -3 pN (4), a force similar in magnitude to thatdeveloped by a cycling crossbridge (6).The original reason for localizing force generation in the
hinge region is that it is potentially the least stable segmentof the rod-like tail of the myosin molecule. The size of thehinge domain in vertebrate muscle (-150 residues per poly-peptide chain) has been determined from its amino acidsequence (7, 8). The replacement of some of the hydrophobicresidues at the interface between the two a-helices by polarresidues appears to be the physical basis for the reducedstability of this region of the myosin rod. Electron micros-copy has shown (9, 10) that only the intact molecule and itshinge-containing fragments, rod and long S-2, shorten (by10-15 nm) when heated from 250C to 50'C under conditionsof zero load. The extension of the enzyme probe technique(proteolytic digestibility as a measure of helix-coil stability)to organized muscle structure has provided a powerful assayfor sites of local melting in the helical structure in situationswhere conventional solution-based physical techniques arenot easily applied. The method relies on random-coil struc-ture serving as a good substrate and a-helical structure not(11, 12). Evidence for the direct participation of the hingeregion during contraction derives from the observation thatthe rate of proteolytic cleavage in the activated state, atseveral widely distributed sites within the hinge domain, iselevated some 100-fold over that in rigor or relaxed fibers(13).Two major questions can now be asked. First, does melting
within the hinge segment in the environment of the musclefiber actually produce force of comparable magnitude to thatin activated muscle? Second, how does ATP hydrolysis resultin hinge melting and force development? This paper address-es the first of these questions. The approach is a simple oneand was conceived in the light of the clear prediction of thehelix-coil model that force should be generated when a rigor(or activated) fiber is subjected to a temperature jump (Tjump) to perturb the helix-coil equilibrium near the meltingtemperature of the hinge. Similar T-jump experiments tothose described in this paper have failed to detect rubber-likethermoelasticity in rigor fibers (14). In these studies rigorfibers exhibited an abrupt drop in tension on heating, con-sistent with normal thermoelastic expansion.
Since the entire, glycerinated fiber is heated in the laserT-jump experiments described in this paper, it is important toemploy a strategy that allows a thermally induced force to beassigned to a particular structure within the sarcomere. Anoteworthy feature of the S-2 segment is that its attachmentto and release from the helix-stabilizing environment of thebackbone of the thick filament can be modulated by divalentmetal ions (17-21). The solvents employed to do this in the
Abbreviations: S-2, subfragment 2; S-i, subfragment 1; [M2],divalent metal ion concentration; Tes, N-[tris(hydroxymethyl)meth-yllaminoethanesulfonic acid; T jump, temperature jump.
975
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.
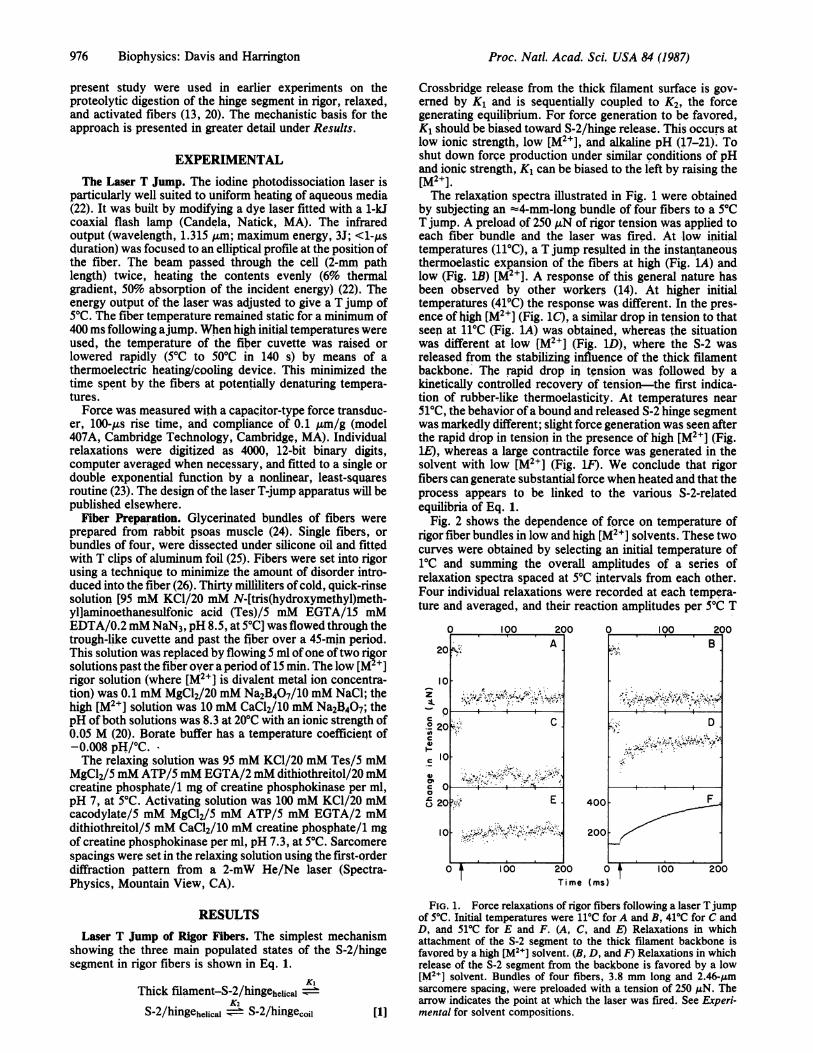
976 Biophysics: Davis and Harrington
present study were used in earlier experiments on theproteolytic digestion of the hinge segment in rigor, relaxed,and activated fibers (13, 20). The mechanistic basis for theapproach is presented in greater detail under Results.
EXPERIMENTALThe Laser T Jump. The iodine photodissociation laser is
particularly well suited to uniform heating of aqueous media(22). It was built by modifying a dye laser fitted with a 1-kJcoaxial flash lamp (Candela, Natick, MA). The infraredoutput (wavelength, 1.315 ,um; maximum energy, 3J; <1-psduration) was focused to an elliptical profile at the position ofthe fiber. The beam passed through the cell (2-mm pathlength) twice, heating the contents evenly (6% thermalgradient, 50% absorption of the incident energy) (22). Theenergy output of the laser was adjusted to give a T jump of50C. The fiber temperature remained static for a minimum of400 ms following ajump. When high initial temperatures wereused, the temperature of the fiber cuvette was raised orlowered rapidly (5'C to 500C in 140 s) by means of athermoelectric heating/cooling device. This minimized thetime spent by the fibers at potentially denaturing tempera-tures.
Force was measured with a capacitor-type force transduc-er, 100-As rise time, and compliance of 0.1 Am/g (model407A, Cambridge Technology, Cambridge, MA). Individualrelaxations were digitized as 4000, 12-bit binary digits,computer averaged when necessary, and fitted to a single ordouble exponential function by a nonlinear, least-squaresroutine (23). The design of the laser T-jump apparatus will bepublished elsewhere.
Fiber Preparation. Glycerinated bundles of fibers wereprepared from rabbit psoas muscle (24). Single fibers, orbundles of four, were dissected under silicone oil and fittedwith T clips of aluminum foil (25). Fibers were set into rigorusing a technique to minimize the amount of disorder intro-duced into the fiber (26). Thirty milliliters of cold, quick-rinsesolution [95 mM KCl/20 mM N-[tris(hydroxymethyl)meth-yl]aminoethanesulfonic acid (Tes)/5 mM EGTA/15 mMEDTA/0.2 mM NaN3, pH 8.5, at 5'C] was flowed through thetrough-like cuvette and past the fiber over a 45-min period.This solution was replaced by flowing S ml ofone oftwo rigorsolutions past the fiber over a period of 15 min. The low [M2L]rigor solution (where [M2+] is divalent metal ion concentra-tion) was 0.1 mM MgCl2/20 mM Na2B407/10 mM NaCl; thehigh [M2+] solution was 10 mM CaCl2/10 mM Na2B4O7; thepH of both solutions was 8.3 at 20'C with an ionic strength of0.05 M (20). Borate buffer has a temperature coefficient of-0.008 pH/0C.The relaxing solution was 95 mM KCl/2Q mM Tes/5 mM
MgCl2/5 mM ATP/5 mM EGTA/2mM dithiothreitol/20 mMcreatine phosphate/i mg of creatine phosphokinase per ml,pH 7, at 50C. Activating solution was 100 mM KCl/20 mMcacodylate/5 mM MgCl2/5 mM ATP/5 mM EGTA/2 mMdithiothreitol/5 mM CaCl2/10 mM creatine phosphate/i mgof creatine phosphokinase per ml, pH 7.3, at 50C. Sarcomerespacings were set in the relaxing solution using the first-orderdiffraction pattern from a 2-mW He/Ne laser (Spectra-Physics, Mountain View, CA).
RESULTSLaser T Jump of Rigor Fibers. The simplest mechanism
showing the three main populated states of the S-2/hingesegment in rigor fibers is shown in Eq. 1.
KiThick filament-S-2/hingehefical =
K2
Crossbridge release from the thick filament surface is gov-erned by K1 and is sequentially coupled to K2, the forcegenerating equilibrium. For force generation to be favored,K1 should be biased toward S-2/hinge release. This occurs atlow ionic strength, low [M2"], and alkaline pH (17-21). Toshut down force production under similar conditions of pHand ionic strength, K1 can be biased to the left by raising the[M2+].The relaxation spectra illustrated in Fig. 1 were obtained
by subjecting an 4-mm-long bundle of four fibers to a 5TCT jump. A preload of 250 AN of rigor tension was applied toeach fiber bundle and the laser was fired. At low initialtemperatures (11TC), a T jump resulted in the instantaneousthermoelastic expansion of the fibers at high (Fig. LA) andlow (Fig. 1B) [M2+]. A response of this general nature hasbeen observed by other workers (14). At higher initialtemperatures (419C) the response was different. In the pres-ence of high [M2+] (Fig. 1C), a similar drop in tension to thatseen at 11TC (Fig. LA) was obtained, whereas the situationwas different at low [M2+] (Fig. 1D), where the S-2 wasreleased from the stabilizing influence of the thick filamentbackbone. The rapid drop in tension was followed by akinetically controlled recovery of tension-the first indica-tion of rubber-like thermoelasticity. At temperatures near510C, the behavior ofa bound and released S-2 hinge segmentwas markedly different; slight force generation was seen afterthe rapid drop in tension in the presence of high [M2"] (Fig.lE), whereas a large contractile force was generated in thesolvent with low [M2+] (Fig. lF). We conclude that rigorfibers can generate substantial force when heated and that theprocess appears to be linked to the various S-2-relatedequilibria of Eq. 1.
Fig. 2 shows the dependence of force on temperature ofrigor fiber bundles in low and high [M2+] solvents. These twocurves were obtained by selecting an initial temperature of1MC and summing the overall amplitudes of a series ofrelaxation spectra spaced at 5YC intervals from each other.Four individual relaxations were recorded at each tempera-ture and averaged, and their reaction amplitudes per 5YC T
Time (ms)
FIG. 1. Force relaxations of rigor fibers following a laser Tjumpof 50C. Initial temperatures were 110C for A and B, 41'C for C andD, and 51'C for E and F. (A, C, and E) Relaxations in whichattachment of the S-2 segment to the thick filament backbone isfavored by a high [M2I] solvent. (B, D, and F) Relaxations in whichrelease of the S-2 segment from the backbone is favored by a low[M2+] solvent. Bundles of four fibers, 3.8 mm long and 2.46-,umsarcomere spacing, were preloaded with a tension of 250 MAN. Thearrow indicates the point at which the laser was fired. See Experi-mental for solvent compositions.
Proc. Natl. Acad. Sci. USA 84 (1987)
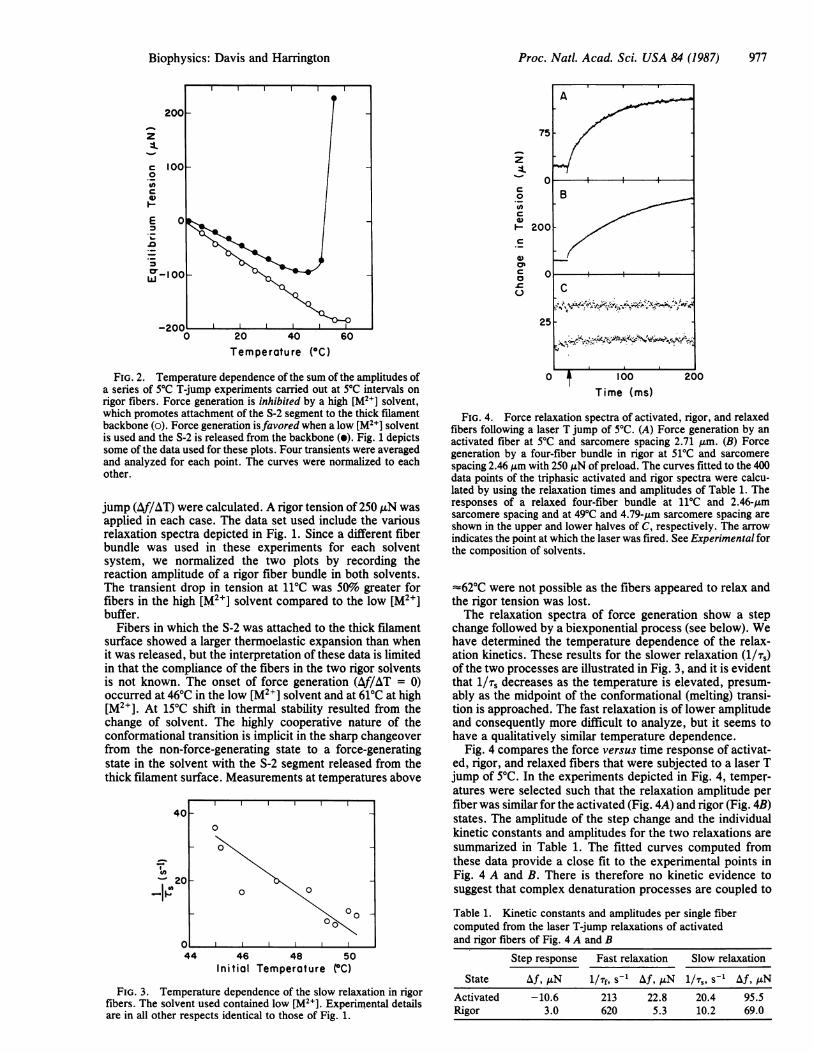
Proc. Natl. Acad. Sci. USA 84 (1987) 977
7
z
0
0)
C7
0'
20
Temperature (IC)
FIG. 2. Temperature dependence of the sum of the amplitudes ofa series of 5PC T-jump experiments carried out at 50C intervals onrigor fibers. Force generation is inhibited by a high [M2+] solvent,which promotes attachment of the S-2 segment to the thick filamentbackbone (0). Force generation isfavored when a low [M2+] solventis used and the S-2 is released from the backbone (0). Fig. 1 depictssome of the data used for these plots. Four transients were averagedand analyzed for each point. The curves were normalized to eachother.
jump (Af/AT) were calculated. A rigor tension of 250 AtN wasapplied in each case. The data set used include the variousrelaxation spectra depicted in Fig. 1. Since a different fiberbundle was used in these experiments for each solventsystem, we normalized the two plots by recording thereaction amplitude of a rigor fiber bundle in both solvents.The transient drop in tension at 110C was 50o greater forfibers in the high [M2+] solvent compared to the low [M2+]buffer.
Fibers in which the S-2 was attached to the thick filamentsurface showed a larger thermoelastic expansion than whenit was released, but the interpretation of these data is limitedin that the compliance of the fibers in the two rigor solventsis not known. The onset of force generation (Af/AT = 0)occurred at 460C in the low [M2+] solvent and at 61'C at high[M2+]. At 150C shift in thermal stability resulted from thechange of solvent. The highly cooperative nature of theconformational transition is implicit in the sharp changeoverfrom the non-force-generating state to a force-generatingstate in the solvent with the S-2 segment released from thethick filament surface. Measurements at temperatures above
40
0
20 X
r0,0 . . .00
44 46 48 50Initial Temperature (IC)
FIG. 3. Temperature dependence of the slow relaxation in rigorfibers. The solvent used contained low [M2+]. Experimental detailsare in all other respects identical to those of Fig. 1.
0 t 100 200Time (ms)
FIG. 4. Force relaxation spectra of activated, rigor, and relaxedfibers following a laser T jump of 50C. (A) Force generation by anactivated fiber at 50C and sarcomere spacing 2.71 Aum. (B) Forcegeneration by a four-fiber bundle in rigor at 51'C and sarcomerespacing 2.46 Am with 250 .iN of preload. The curves fitted to the 400data points of the triphasic activated and rigor spectra were calcu-lated by using the relaxation times and amplitudes of Table 1. Theresponses of a relaxed four-fiber bundle at 110C and 2.46-Lmsarcomere spacing and at 490C and 4.79-pm sarcomere spacing areshown in the upper and lower halves of C, respectively. The arrowindicates the point at which the laser was fired. See Experimental forthe composition of solvents.
-620C were not possible as the fibers appeared to relax andthe rigor tension was lost.The relaxation spectra of force generation show a step
change followed by a biexponential process (see below). Wehave determined the temperature dependence of the relax-ation kinetics. These results for the slower relaxation (1/;)of the two processes are illustrated in Fig. 3, and it is evidentthat 1/Ts decreases as the temperature is elevated, presum-ably as the midpoint of the conformational (melting) transi-tion is approached. The fast relaxation is of lower amplitudeand consequently more difficult to analyze, but it seems tohave a qualitatively similar temperature dependence.
Fig. 4 compares the force versus time response of activat-ed, rigor, and relaxed fibers that were subjected to a laser Tjump of 50C. In the experiments depicted in Fig. 4, temper-atures were selected such that the relaxation amplitude perfiber was similar for the activated (Fig. 4A) and rigor (Fig. 4B)states. The amplitude of the step change and the individualkinetic constants and amplitudes for the two relaxations aresummarized in Table 1. The fitted curves computed fromthese data provide a close fit to the experimental points inFig. 4 A and B. There is therefore no kinetic evidence tosuggest that complex denaturation processes are coupled to
Table 1. Kinetic constants and amplitudes per single fibercomputed from the laser T-jump relaxations of activatedand rigor fibers of Fig. 4 A and B
Step response Fast relaxation Slow relaxation
State Af, ,N l/Tf, s-' Af, AN 1/;,, s-1 Af, ANActivated -10.6 213 22.8 20.4 95.5Rigor 3.0 620 5.3 10.2 69.0
A
F5toB10
!5-
I,>c a bC. 1 -~'lI..-~I-4 . ?
Biophysics: Davis and Harrington
2

978 Biophysics: Davis and Harrington
force generation in rigor fibers at high temperatures. As canbe seen from Table 1, the reciprocal relaxation times andamplitudes for activated and rigor fibers differ only by factorsof2-4. It will also be noted that the initial step change in forceis negative for activated fibers and positive for rigor fibers athigh temperature.Relaxed fibers do not appear to generate force when
subjected to a laser Tjump since similar responses were seenat 110C and a normal sarcomere spacing and at 51TC whenstretched beyond overlap (Fig. 4C). These controls excludethe possibility of a significant thermoelastic response arisingfrom the structures responsible for passive tension.
DISCUSSIONThere can be little question that rigor muscle generates forcein response to a change in temperature, thereby fulfilling amajor prediction of the helix-coil model of muscle contrac-tion. The site of the conformational transition, its physicalorigin, and its relationship to the contraction of activatedmuscle are considered in what follows.
Crossbridge-actin interaction appears to be a prerequisitefor force generation in rigor fibers. Force does not appear toarise from a temperature-induced change within the struc-tural components responsible for the passive tension ofrelaxed fibers because relaxed fibers and, in particular,relaxed fibers stretched beyond overlap at high temperaturesat significant passive tensions do not generate force whensubjected to a T jump.A good case can be made for locating the force-generating
process in rigor fibers within the S-2 segment of the myosinmolecule. The strategy used was to switch S-2 between statesin which it was released from or bound to the shaft ofthe thickfilament (17-21). Biasing the first equilibrium in Eq. 1 fromthe attached to the released state by changing solvents fromhigh to low [M2+] resulted in a downward'displacement in theonset of force generation by 15'C. The midpoint of the localmelting transition within the hinge region of myofibrils asdetermined by proteolytic digestion (20) is also lowered by anidentical alteration in solvent composition. We thereforeconclude that force production originates somewhere in theS-2 region of the myosin molecule, since these two differentphenomena (one of which is definitely located in the hingesegment) respond equivalently to crossbridge release fromthe surface of the thick filament. If the linkage betweentemperature and the two phenomena described above hadbeen nonequivalent, then the coupling of crossbridge releaseto force generation elsewhere in the myosin molecule mighthave been a possibility. Additionally, it seems unlikely thatthe S-1 head could generate force when its structure isostensibly at the energy minimum of the ATPase cycle.
Localization of the force-generating event in the hingesegment is supported by other evidence. Much of thisinformation was presented in the Introduction and includesdata on the low stability of the hinge a-helical segment, asevidenced by in vitro melting data (27, 28), the distinctiveamino acid sequence (7, 8), physical shortening of the hingein myosin and hinge-containing rod fragments under zeroload when heated (9, 10), and the marked proteolytic sus-ceptibility of the hinge of activated, compared to rigor orrelaxed, fibers (13, 20). In the laser T-jump experiments, themidpoint of the force producing transition appears to liebetween 550C and 60'C in low [M2+] solvents. This value ishigh compared to the melting temperature of 350C obtainedfor the hinge segment in solution (28). Nevertheless, itappears that an increasing level of structural complexity(segments of the myosin molecule containing the hinge, tomyofibrils, to glycerinated fibers) leads to progressivelyhigher melt temperatures (13, 20). It should also be noted thatthe preload rigor tension used in the laser T-jump experi-
ments would elevate the transition temperature by favoringhelix formation.Force generation in rigor fibers almost certainly has its
origin in a helix-coil-type transition. The high cooperativityand the kinetic form of the T-jump data are typical of ahelix-coil transition in a linear polymer. Conventional T-jumpexperiments on isolated segments of myosin a-helical coiled-coil structures reveal a typical biexponential response inwhich the reciprocal relaxation times decline with increasingtemperature toward the midpoint ofthe transition (28, 29). Infiber experiments, the reciprocal relaxation time of the slowprocess 1/; and probably the fast 1/rf also decrease inmagnitude as the temperature is elevated. The other obviouspoint is that the S-2 is an a-helical coiled-coil structure (15)and it does appear to be the myosin segment in which force,originates.The question that remains, is whether the force seen in
rigor and activated fibers is of similar origin. There are anumber of lines of evidence that support this view. (i) Inactivated fibers of rabbit psoas muscle, local melting with amelting temperature of 2Q0C ± 50C occurs at various sitesthroughout the hinge region (13). Rigor and relaxed fibershave significantly higher melting temperature values =(13, 19).Therefore, the hydrolysis of ATP during a contractile cycleappears to be coupled to the destabilization of the hingestructure. (it) The amount of force generated in response toa T jump is of similar magnitude in a rigor fiber at 510C andan activated fiber at 50C. (iii) The kinetic responses ofthe twosystems are in many ways similar. A rapid but small transientchange in force is observed within the dead time of theinstrument. This is followed by a fast relaxation of moderateamplitude and a slow relaxation of larger amplitude. Theindividual kinetic constants and amplitudes for both systemsdiffer by no more than factors of2-4 from each other. (iv) Thecloseness of fit of the data to a biexponential function in theactivated and rigor systems points to a physically similarorigin in a mechanism with a minimum of three states.The remaining point is that force is being produced in the
present experiments by a muscle ostensibly at the forceenergy minimum of the ATPase cycle. If the scheme asoutlined above is correct, then it is the indirect coupling ofATP hydrolysis (16) to the helix-coil melt that destabilizes thea-helical coiled-coil. In rabbit muscle operating at 390C, itseems logical to produce force by switching from a stable(high melting temperature) state to a less-stable (low meltingtemperature) force-generating state.
We thank Drs. John Gormally, Frederick Davidson, and HoraceFurumoto for help with the design and construction of the laser; Mr.Zygmunt Schroeder for constructing the force measurement cell; Dr.Michael Rodgers for comments on the manuscript, interfacing thecomputer to the instrument, and the data handling and analysisroutines; and Dr. Yale Goldman for generously allowing access tounpublished material. The work was supported by Grant AM-04349from the National Institutes of Health (to W.F.H.). The paper iscontribution no. 1344 from the Biology Department (Johns HopkinsUniv.).
1. Huxley, H. E. (1969) Science 164, 1356-1366.2. Huxley, H. E. & Kress, M. (1985) J. Muscle Res. Cell Motil.
6, 153-161.3. Harrington, W. F. (1971) Proc. Natl. Acad. Sci. USA 68,
685-689.4. Harrington, W. F. (1979) Proc. Natl. Acad. Sci. USA 76,
5066-5070.5. Woledge, R. C., Curtin, N. A. & Homsher, E. (1985) Ener-
getic Aspects of Muscle Contraction. Monographs of thePhysiological Society No. 41 (Academic, London).
6. Huxley, A. F. & Simmons, R. M. (1971) Nature (London) 233,533-538.
7. Lu, R. C. & Wong, A. (1985) J. Biol. Chem. 260, 3456-3461.8. Strehler, E. E., Strehler-Page, M. A., Perriard, J. C.,
Proc. Natl. Acad Sci. USA 84 (1987)

Biophysics: Davis and Harrington
Periasamy, M. & Nadal-Ginard, B. (1986) J. Mol. Biol. 190,291-317.
9. Walzthony, D., Eppenberger, H. M., Ueno, H., Harrington,W. F. & Wallimann, T. (1986) Eur. J. Cell Biol. 41, 38-43.
10. Walker, M. & Trinick, J. A. (1986) J. Mol. Biol. 192, 661-667.11. Ueno, H. & Harrington, W. F. (1984) J. Mol. Biol. 173, 35-61.12. Ueno, H. & Harrington, W. F. (1984) J. Mol. Biol. 180,
667-701.13. Ueno, H. & Harrington, W. F. (1986) J. Mol. Biol. 190, 69-82.14. Smith, J. J., Goldman, Y. E., Hibberd, M. G., Ligouri, G. A.,
Luttmann, M. A. & McCray, J. A. (1984) Proceedings of theEighth International Biophysics Congress [Int. Union PureAppl. Phys., Bristol, UK and Royal Soc. (London), London,UK], p. 204.
15. McLachlan, A. D. & Karn, J. (1983) J. Mol. Biol. 164, 605-626.
16. Geeves, M. A., Goody, R. S. & Gutfreund, H. (1984) J.Muscle Res. Cell Motil. 5, 351-361.
17. Ueno, H., Rodgers, M. E. & Harrington, W. F. (1983) J. Mol.Biol. 160, 207-228.
18. Reisler, E., Liu, J. & Cheung, P. (1983) Biochemistry 22,4954-4960.
Proc. Natl. Acad. Sci. USA 84 (1987) 979
19. Persechini, A. & Rowe, A. J. (1984) J. Mol. Biol. 172, 23-39.20. Ueno, H. & Harrington, W. F. (1986) J. Mol. Biol. 190, 59-68.21. Harrington, W. F., Ueno, H. & Davis, J. S. (1986) in Sympo-
sium on Molecular Mechanisms of Muscle Contraction, eds.Sugi, H. & Pollack, G. H. (Plenum, New York), in press.
22. Holzwarth, J. F. (1979) in Techniques and Applications ofFastReactions in Solution, eds. Gettins, L. J. & Wyn-Jones, E.(Reidel, Dordrecht, Netherlands), pp. 47-59.
23. McCalla, T. R. (1967) in Introduction to Numerical Methodsand Fortran Programming (Wiley, New York), pp. 255-260.
24. Brenner, B. (1983) Biophys. J. 41, 99-102.25. Goldman, Y. E. & Simmons, R. M. (1984) J. Physiol. 350,
497-518.26. Schoenberg, M. & Eisenberg, E. (1985) Biophys. J. 48, 863-
871.27. Swenson, C. A. & Ritchie, P. A. (1980) Biochemistry 19,
5371-5375.28. Tsong, T. Y., Himmelfarb, S. & Harrington, W. F. (1983) J.
Mol. Biol. 164, 431-450.29. Tsong, T. Y., Karr, T. & Harrington, W. F. (1979) Proc. Natl.
Acad. Sci. USA 76, 1109-1113.





























