Embed Size (px)
Citation preview

Forelimb Anatomy of the Microsyopidae (Mammalia: Primatomorpha):
implications for primate origins
By DANIEL L. GEBO1 and K. CHRISTOPHER BEARD2
1Department of Anthropology, Northern Illinois University, 2Department of Ecology & Evolutionary Biology, University of Kansas
Introduction
Although historical attempts to reconstruct the
phylogenetic relationships of the extinct family
Microsyopidae have yielded variable and often
conflicting results, all recent analyses have interpreted
these animals as stem Primates or members of the
crown clade Primatomorpha. Here we describe the
forelimb anatomy of Microsyops and Niptomomys, two
microsyopids from the Eocene of North America. In
both cases the distal humeral anatomy of these
microsyopids mirrors that of crown clade primates
from the early Eocene in having a spherical capitulum
associated with a wide and distinct spatial gutter
separating the capitulum from the trochlea. This
primate-like distal humeral anatomy contrasts with
that of other known plesiadapiforms, which have a
subspherical capitulum and lack the distinct
separation between the capitulum and trochlea found
in microsyopids. Microsyopid elbow morphology
allows for an extensive range of forearm pronation
and supination, movements that are often utilized
during locomotion on small diameter arboreal
supports or during single-handed manual
manipulation of food items. In this poster, we consider
the best phyletic position for the Microsyopidae
Acknowledgments
We thank Dr. Luo Zhe-Xi and April Neander at the
University of Chicago for microCT scanning these
fossils. Rendering and images for this poster was
done by Joshua Schwartz (Geology Department,
Northern Illinois University). Financial support was
provided by the David B. Jones Foundation.
Figure 2. Forelimb elements associated
with the maxilla of Microsyops
latidens, USGS 16647
References
Beard KC. 1993a Origin and evolution of gliding in early Cenozoic
Dermoptera (Mammalia, Primatomorpha). In Primates and their relatives in
phylogenetic perspective (ed RDE MacPhee), pp. 63-90. New York: Plenum
Press.
Beard KC. 1993b Phylogenetic systematics of the Primatomorpha, with
special reference to Dermoptera. In Mammal phylogeny, volume 2,
placentals (eds FS Szalay, MJ Novacek, MC McKenna), pp. 129-150. New
York: Springer-Verlag.
Beard KC, Houde P. 1989 An unusual assemblage of diminutive
plesiadapiforms (Mammalia, ?Primates) from the early Eocene of the Clark’s
Fork Basin, Wyoming. J. Vert. Paleontol. 9, 388-399.
Bloch JI, Silcox MT, Boyer DM, Sargis EJ. 2007 New Paleocene skeletons
and the relationship of plesiadapiforms to crown-clade primates. Proc. Natl.
Acad. Sci. USA 104, 1159-1164.
Bloch JI, Chester, SG, Silcox MT. 2016 Cranial anatomy of Paleogene
Micromomyidae and implications for early primate evolution. J. Hum. Evol.
96, 58-81.
Bown TM, Rose KD, Simons EL, Wing SL. 1994 Distribution and
stratigraphic correlation of upper Paleocene and lower Eocene fossil
mammal and plant localities of the Fort Union, Willwood, and Tatman
formations, southern Bighorn Basin, Wyoming. U.S. Geol. Surv. Prof. Pap.
1540, 1-103.
Gunnell GF. 1989 Evolutionary history of Microsyopoidea (Mammalia,
?Primates) and the relationship between Plesiadapiformes and Primates.
Univ. Michigan Pap. Paleontol. 27, 1-157.
MacPhee, RDE, Cartmill M, Gingerich PD 1983. New Paleogene primate
basicrania and the definition of the order Primates. Nature 301:309-311.
Silcox, M.T., Benham, A.E., and Bloch, J.I. 2010. Endocasts of Microsyops
(Microsyopidae, Primates) and the evolution of the brain in primitive
primates. J Hum Evol 58:505-521.
Silcox MT, Bloch JI, Boyer DM, Chester SGB, López-Torres S. 2017 The
evolutionary radiation of plesiadapiforms. Evol. Anthropol. 26, 74-94.
Szalay, F.S., Tattersall, I., and Decker, R.L. 1975. Phylogenetic Relationships
of Plesiadapis – Postcranial Evidence. In F.S. Szalay, ed., Approaches to
Primate Paleontology, Contrib. Primates, Karger, Basel, Vol. 5:136-166.
Discussion
The phylogenetic position of microsyopids has been
controversial for decades. Recent phylogenetic
analyses of euarchontan relationships reconstructs
microsyopids as relatively basal members of the
plesiadapiform radiation, all of which are interpreted as
comprising a paraphyletic stem group of primates
(Bloch et al., 2007, 2016; Silcox et al., 2017). However,
disagreement persists with regard to the precise
phylogenetic placement of microsyopids in these
analyses, with some tree topologies reconstructing
microsyopids as the sister group of micromomyids
(Bloch et al., 2016), while others reconstruct
microsyopids as being more closely related to
paromomyids, plesiadapoids, and crown clade primates
(Bloch et al., 2007; Silcox et al., 2017). In contrast, the
large-scale phylogenetic analysis of morphological and
molecular data reported by Ni et al. (2016)
reconstructed microsyopids and other plesiadapiforms
either as stem dermopterans or stem primatomorphs.
One approach advocated before has been Beard’s
(1993a,b) Primatomorpha, a phyletic grouping that links
both Orders, Dermoptera and Primates, with several
clades of plesiadapiforms contained within Dermoptera.
Microsyopid elbow anatomy adds a new anatomical
complex to this phyletic debate and this character
complex supports a closer evolutionary connection to
primates for the Microsyopidae -- in contrast to an
interpretation as a basal member among the clades of
plesiadapiforms. When we assess additional
anatomical features that might also signal a phyletic link
between the Microsyopidae with primates or
“euprimates” as sister taxa, the list of features is short.
The arterial supply to the brain (MacPhee et al., 1983)
or the larger encephalization quotient for Microsyops
(Silcox et al., 2010) are additional features worthy of
consideration. In contrast, the dental evidence for
microsyopids has largely been interpreted as “primitive”
among comparisons with other plesiadapiforms. If
microsyopids are a sister taxon to primates than a
sister link with Plesiadapodiea is not correct In the
end, the phyletic position of the Microsyopidae
continues to be controversial.
Conclusion
The forelimb anatomy of the
Microsyopidae closely resembles crown
clade primates, especially relative to any
of the distal humeri known for the other
plesiadapiforms. If the elbow
morphology among microsyopids makes
a sister taxon link to early Eocene
primates, a position we advocate here,
then the Plesiadapoidea are not. The
plesiadapoid clades, the Saxonellidae,
the Carpolestidae, and the
Plesiadapidae, must be re-evaluated as
a close phyletic link to “euprimates” in
the Bloch et al. (2007) or Silcox et al.
(2007) sense. A formal cladistic analysis
will be needed to support this claim.
Materials
Two specimens from the early Eocene Willwood
Formation, Bighorn Basin, Wyoming, provide
information on the forelimb anatomy of Microsyopidae.
USGS 16647, consisting of associated maxillary
fragments (Fig. 1) and left forelimb elements (Fig. 2),
was collected by Dr. Chris Beard at locality D-1647,
which occurs stratigraphically approximately 591 m
above the base of the Willwood Formation (Bown et
al., 1994). Based on the size and morphology of its
dentition, USGS 16647 pertains to Microsyops
latidens, which is the only microsyopine known to
occur at this stratigraphic level in the Willwood
Formation (Gunnell, 1989).
USNM 639759 is a tiny right distal humerus (Fig.
3) from the same freshwater limestone that yielded the
sample of micromomyid plesiadapiforms reported by
Beard and Houde (1989). The provenance of this
fossil is very near University of Michigan locality SC-4,
which occurs stratigraphically in the early Wasatchian
Wa-1 faunal zone. Based on its size, morphology, and
provenance, we allocate this isolated distal humerus
to the uintasoricine microsyopid Niptomomys
doreenae.
Figure 1. USGS 16647,
maxilla of Microsyops
latidens
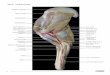
Figure 3. Microsyopid elbow anatomy. Left: USNM 639759,
Niptomomys doreenae; Right: USGS 16647, Microsyops
latidens. At the top, (a) and (e) compare distal humeri at scale.
Below, (b) through (d) are enlarged and reversed humeral comparisons of USNM 639759 at relatively the same bicondylar
width of USGS 16647 (anterior (b) and (f), distal edge (c) and (g),
and posterior (d) and (h) views.
Figure 4. Distal humeral comparisons. A through d are early Eocene fossil primates: a) Cantius, USGS 6759; b)
asiadapine, GU 713, reversed; c) Teilhardina, IRSNB M2160; d) Archicebus, IVPP V18618. e) Plesiadapis,
MNH BR 3L and B4L, redrawn from Szalay et al., 1975) and f ) Niptomomys, USNM 639759) are
plesiadapiforms. Note the less rounded capitulum and the capitulaum-trochlear connection (shaded joints) in
the elbow joint of Plesiadapis (e) relative to early Eocene fossil primates (a-d). In contrast to Plesiadapis, the
microsyopid elbow of Niptomomys (f) is similar to that of early Eocene fossil primates (a-d).
Enlarged & Reversed
Niptomomys Microsyops