Embed Size (px)
Citation preview

JOURNAL OF MASS SPECTROMETRYJ. Mass Spectrom. 2005; 40: 1173–1186Published online 22 July 2005 in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/jms.891
Fragmentation reactions of deprotonated peptidescontaining proline. The proline effect
Alex G. Harrison∗ and Alex B. Young
Department of Chemistry, University of Toronto, Toronto, Canada
Received 3 January 2005; Accepted 10 May 2005
The collision-induced dissociation (CID) fragmentation reactions of a variety of deprotonated peptidescontaining proline have been studied in detail using MS2 and MS3 experiments, deuterium labelling andaccurate mass measurements when necessary. The [M–H–CO2]− (a2) ion derived from H-Pro-Xxx-OHdipeptides shows an unusual fragmentation involving loss of C2H4; this fragmentation reaction is notobserved for larger peptides. The primary fragmentation reactions of deprotonated tripeptides with anN-terminal proline are formation of a3 and y1 ions. When proline is in the central position of tripeptides,a3, y2 and y1 ions are the primary fragmentation products of [M–H]−, while when the proline is in theC-terminal position, a3and y1 ions are the major primary products. In the latter case, the a3 ion fragmentsprimarily to the ′′b2 ion; further evidence is presented that the ′′b2 ions have a deprotonated oxazolonestructure. Larger deprotonated peptides having at least two amino acid residues N-terminal to prolineshow a distinct preference for cleavage of the amide bond N-terminal to proline to form, mainly, theappropriate y ion. This proline effect is compared and contrasted with the similar proline effect observedin the fragmentation of protonated peptides containing proline. Copyright 2005 John Wiley & Sons, Ltd.
KEYWORDS: proline effect; peptide fragmentation; negative ions; CID
INTRODUCTION
Tandem mass spectrometry of protonated peptides has beenclearly established as a powerful technique for determiningthe amino acid identities and sequence for the peptide.1 – 4 Asa result of many studies over the past decade, considerablemechanistic information concerning fragmentation modesand fragment-ion structures has been elucidated to aid in theinterpretation of the tandem mass spectra; these mechanisticstudies have been reviewed recently in several excellentarticles.5 – 9 Rather less is known concerning the detailedfragmentation modes of deprotonated peptides. A numberof early studies reported the high-energy collision-induceddissociation (CID) mass spectra of the [M–H]� ions of mainlydi- and tripeptides. The mechanistic information obtainedfrom these studies has been summarized by Bowie andcoworkers.10,11 More recently Bowie and coworkers havecarried out extensive studies of the low-energy CID of largerdeprotonated peptides and have shown that useful sequenceinformation can be obtained which often is complementary tothat obtained by CID of the protonated peptide; these studieshave recently been reviewed.12 Cassady and coworkers alsohave explored the low-energy fragmentation of a varietyof larger deprotonated peptides,13 – 16 while Marzluff et al.17
ŁCorrespondence to: Alex G. Harrison, Department of Chemistry,University of Toronto, 80 St. George Street, Toronto, Ontario, M5S3H6, Canada. E-mail: [email protected]/grant sponsor: Natural Sciences and EngineeringResearch Council (Canada).
have explored the lowest energy fragmentation pathways ofa variety of small deprotonated peptides.
The studies to date have shown that the fragmentions observed on collisional activation of deprotonatedpeptides can be categorized as shown in Scheme 1 usingnomenclature15,18 adapted from that used for protonatedpeptide fragmentation.19,20 However, the studies of Bowieand coworkers10 – 12 have shown that there often are frag-mentation reactions that are specific to particular side-chaingroups in the peptide. In our initial study18 of deproto-nated peptides containing only H or alkyl side chains, weobserved that there was more extensive sequence-specificfragmentation under low-energy CID conditions than underhigh-energy CID conditions.21 We have extended these stud-ies to deprotonated peptides containing phenylalanine22 andto deprotonated peptides containing glutamic acid;23 in bothcases fragmentation reactions specific to the side chain func-tionality were observed. In the present work, we explorethe fragmentation reactions observed under low-energycollisional activation for deprotonated peptides containingproline and compare the results obtained with those reported
Scheme 1
Copyright 2005 John Wiley & Sons, Ltd.

1174 A. G. Harrison and A. B. Young
by Bowie and coworkers24 using high-energy collisional acti-vation. A substantial proline effect, i.e. cleavage of the amidebond N-terminal to proline is observed. This ‘proline effect’ iscompared and contrasted with the ‘proline effect’ observed inthe fragmentation of protonated peptides containing proline.
EXPERIMENTAL
Initial CID studies were carried out using an elec-trospray/quadrupole mass spectrometer (VG Platform,Micromass, Manchester, UK) with CID in the interfaceregion between the atmospheric pressure source and thequadrupole mass analyzer, so-called cone-voltage CID. Ithas been clearly established25 – 29 that CID in this fashion pro-duces mass spectra similar to those obtained by low-energyCID in quadrupole cells. MS2 and MS3 experiments alsowere carried out using an electrospray/quadrupole/time-of-flight (Qq-ToF) mass spectrometer (QStar, MDS Sciex,Concord, Canada). In the MS3 experiments, CID in the inter-face region produced fragment ions, those of interest beingmass-selected by the first quadrupole mass analyzer (Q) toundergo collisional activation in the quadrupole collision cell(q), with the ionic fragmentation products being analyzedby the time-of-flight analyzer.30,31 The Qq-ToF instrumentwas operated under conditions which permitted accuratemass measurements to confirm elemental compositions ofthe fragment ions where necessary.
Ionization was by electrospray in both instruments. Forthe single quadrupole instrument, the peptide was dissolvedin 1 : 1 CH3CN:1% aqueous NH3 and introduced into thesource at a flow rate of 30 µl min�1 with nitrogen used as bothnebulizing and drying gas. The use of 1 : 1 CD3CN:1% ND3 inD2O as solvent resulted in exchange of all labile hydrogensand formation of the [M–D]� ion of the deuterated peptide.For the Qq-ToF instrument, the peptide was dissolved in 1 : 1CH3OH:0.1% aqueous NH3 and introduced into the sourceat a flow rate of 80 µl min�1. Nitrogen was used as nebulizingand drying gas and as collision gas in the quadrupole cell.
All peptide samples were obtained from Bachem Bio-sciences (King of Prussia, PA) and were used as received.
RESULTS AND DISCUSSION
DipeptidesThe low-energy CID mass spectra of the [M–H]� ions of H-Pro-Val-OH and H-Ala-Pro-OH have been reported earlier.32
In agreement with the results of Bowie and coworkers,24 themajor primary fragmentation reactions involve eliminationof CO2 to form the a2 ion and formation of the y1 ion(Scheme 1). In the present work, we have studied the tandemmass spectra of deprotonated H-Pro-Ala-OH and H-Pro-Val-OH using the Q-ToF instrument. Bowie and coworkers24
reported unexpected ion signals at m/z 113 and m/z 72in the high-energy CID of the former while there is anunexpected ion signal at m/z 141 for the latter.32 Table 1records the MS2 mass spectrum of the [M–H]� ion of H-Pro-Ala-OH as well as the MS3 mass spectrum of the a2 ionderived therefrom, while Table 2 records similar data forH-Pro-Val-OH. Clearly, the primary fragmentation reactions
Scheme 2
Scheme 3
Table 1. CID mass spectra of anions derived fromH-Pro-Ala-OH (20 eV collision energy)
m/z (ion) [M–H]� (m/z 185) a2 (m/z 141)
167 5.0141 (a2) 52.3139 16.9 47.4113 27.7 100
88 (y1) 10072 6.6 15.2
of the [M–H]� anions involve loss of CO2 to form the a2 ionand nominal elimination of the proline residue to form they1 ion. The other fragment ions observed arise by furtherfragmentation of the a2 ions. The m/z 72 ion of Table 1and the m/z 100 ion of Table 2 originate by the reactionsequence of Scheme 2, first elucidated by Styles and O’Hair33
and elaborated upon by Chass et al.,32 although under thepresent conditions the deprotonated amine was not formedto a significant extent. Accurate mass measurement on them/z 113 ion signal from H-Pro-Ala-OH gave the elementalcomposition [C5H9N2O]� corresponding to elimination ofC2H4 from the a2 ion. Similarly, the m/z 141 ion fromH-Pro-Val-OH gave the composition [C7H13N2O]� againcorresponding to C2H4 elimination from the a2 ion. Aplausible mechanism for ethylene elimination from the a2
ion is shown in Scheme 3.
TripeptidesTable 3 records the CID mass spectra for deprotonated H-Pro-Gly-Gly-OH and two fragment ions derived therefrom asobtained on the Q-ToF instrument at 20 eV collision energy.Five product ions are observed on fragmentation of the
Copyright 2005 John Wiley & Sons, Ltd. J. Mass Spectrom. 2005; 40: 1173–1186

Fragmentation of deprotonated peptides containing proline 1175
Table 2. CID mass spectra of anions derived fromH-Pro-Val-OH (20 eV collision energy)
m/z (ion) [M–H]� (m/z 213) a2 (m/z 169)
169 (a2) 100167 12.7 59.1141 14.1 100116 (y1) 45.9100 5.1 13.9
96 9.4
Table 3. CID mass spectra of anions derived fromH-Pro-Gly-Gly-OH (20 eV collision energy)
m/z (ion) [M–H]� (m/z 228) a3 (m/z 184) 00b2 (m/z 153)
184 (a3) 100153 (00b2) 60.0 62.0138 25.9127 (a2) 19.2 56.5125 100123 29.6
87 (a3-P) 31.9 10074 (y1) 24.8
Scheme 4
[M–H]� ion, as identified in the Table, with the a3 ion[M–H–CO2]� being the base peak. Bowie and coworkers,24
in high-energy CID studies, reported observation of the a3,00b2 and y1 ion, although the signal for the last product wasquite weak. MS3 studies on the a3 ion (m/z 184) shows thatit fragments by loss of CH3NH2 to yield the 00b2 ion, by lossof 57 Da to give the a2 ion and by loss of the proline residueto give m/z 87, with the latter being the base peak. The lossof the N-terminal residue from the a3 ion has been discussedearlier18 as has the formation of 00b2 ions by loss of a neutralamine upon fragmentation of a3 ions.34 The loss of a glycineresidue from the a3 ion is a reaction type which has not beenidentified previously; this reaction presumably involves theelimination of an imine plus CO as illustrated in Scheme 4.The 00b2 ion (m/z 153) fragments by loss of NH3, by loss ofCO and by loss of [H2,CO], the composition of the neutral(s)being confirmed by accurate mass measurements.
Figure 1 compares the cone-voltage CID mass spectrumof deprotonated H-Pro-Gly-Gly-OH with that of the [M–D]�
ion of the peptide in which the labile hydrogens wereexchanged for deuterium. The labelling results show that
Scheme 5
the 00b2 ion incorporates largely one labile hydrogen, thea2 ion largely two labile hydrogens and the a3-P ion threelabile hydrogens to a significant extent, although there isa substantial signal at m/z 89 indicating incorporation ofonly two labile hydrogens. The mechanism(s) of formationand structure(s) of 00b2 ions will be discussed below. Thelabelling results for the a2 ion are largely in accord withthe mechanism of Scheme 4. The labelling results for thea3-P ion indicate that a significant pathway is as shownin Scheme 5, although this pathway will not rationalizethe incorporation of only two labile hydrogens. The y1 ionincorporates, in part, two labile hydrogens, but to a greaterextent one labile hydrogen. Bowie and coworkers24 havereported that the (weak) y1 ion signal showed incorporationof only one labile hydrogen in high-energy CID. Thisdifference in incorporation of labile hydrogens in y1 ionshas been noted previously18 and pathways to y1 ions whichincorporate one or two labile hydrogens have been proposed,as shown in Scheme 6; note, in particular, that in Pathway1 a hydrogen initially bonded to carbon is incorporated inthe final product ion. The differences between the presentresults and the results of Bowie probably reflect differencesin the structure(s) of the [M–H]� ions formed by electrospray(present work) and fast atom bombardment (FAB) as used byBowie and coworkers. In this respect, it should be noted thationization of peptide methyl esters by electrospray involvesdeprotonation at an amide nitrogen18 while ionization ofpeptide esters by FAB involves only deprotonation at carbons˛ to carbonyl groups.35 Thus, it is likely that ionization offree peptides by electrospray will involve, in part, amidenitrogen deprotonation while ionization by FAB will notinvolve amide deprotonation. In Pathway 2 of Scheme 6, wehave proposed that at least some of the [M–H]� ions formedby electrospray ionization originate by proton abstractionfrom an amide nitrogen.
Copyright 2005 John Wiley & Sons, Ltd. J. Mass Spectrom. 2005; 40: 1173–1186

1176 A. G. Harrison and A. B. Young
Figure 1. Comparison of CID mass spectra of [M–H]� of PGG and [M–D]� of PGG-d4. 50 V cone voltage.
Scheme 6
Table 4 records the CID mass spectrum of the [M–H]�
ion of H-Pro-Val-Gly-OH as well as that of the a3 ionderived therefrom. A variety of products are observed, butit appears that the primary fragmentation of the [M–H]� ionyields the a3 ion and the y1 ion with the other productsoriginating by further fragmentation of the a3 ion. Ofparticular note is the ion signal at m/z 183 correspondingto loss of the C3H7 side chain from the a3 ion and the
m/z 155 ion corresponding to further loss of CO fromm/z 183. This is our first observation of the loss of sidechain alkyl groups in low-energy CID, although such losseshave been observed in high-energy CID experiments.12
The major primary fragmentation reactions of the a3 ionremain formation of the 00b2 ion, formation of the a2 ionand loss of the N-terminal proline residue to form m/z129.
Copyright 2005 John Wiley & Sons, Ltd. J. Mass Spectrom. 2005; 40: 1173–1186

Fragmentation of deprotonated peptides containing proline 1177
Table 4. CID mass spectra of anions derived fromH-Pro-Val-Gly-OH (25 eV collision energy)
m/z (ion) [M–H]� (m/z 270) a3 (m/z 226)
226 (a3) 100195 (’’b2) 56.3 48.0183 9.0 21.4169 (a2) 9.0 15.8167 22.8 39.4155 9.2 27.0139 10.7 18.0129 (a3-P) 37.6 100113 (c1) 5.4 15.8
74 (y1) 51.8
Table 5. CID mass spectra of anions derived fromH-Ala-Pro-Gly-OH (20 eV collision energy)
m/z (ion) [M–H]� (m/z 242) a3 (m/z 198) y2 (m/z 171)
198 (a3) 39.2171 (y2) 51.6167 (00b2) 35.2 14.2153 (y2-H2O) 24.0141 (a2) 23.3 12.6139 9.1127 (a2-A),(y2-CO2)
65.7 100 100
125 32.0113 17.4 21.1101 8.1 8.199 36.287 (c1) 7.374 (y1) 100 36.5
Table 5 records the CID mass spectra of deprotonated H-Ala-Pro-Gly-OH and the a3 and y2 ions derived therefrom.The primary fragmentation products of the [M–H]� ionare the a3, y2 and y1 ions. The substantial signal for they2 ion indicates significant cleavage of the amide bond N-terminal to the proline residue, with charge retention onthe C-terminal fragment. The observation of the y2 ion is incontrast to the observation18 that deprotonated tripeptidescontaining only H or alkyl side chains do not fragment toform the y2 ion. The abundant m/z 127 ion signal is shown tohave two origins, loss of the alanine residue from the a3 ionand loss of CO2 from the y2 ion. The CID mass spectrum of they2 ion (deprotonated H-Pro-Gly-OH) has many similaritieswith the spectra of deprotonated H-Pro-Ala-OH (Table 1)and deprotonated H-Pro-Val-OH (Table 2), including loss ofC2H4 from the m/z 127 (a2) ion to give the fragment at m/z99. Fragmentation of deprotonated H-Gly-Pro-Gly-OH andits a3 and y2 ions showed very similar behavior.
Figure 2 compares the CID mass spectrum of the [M–H]�
ion of H-Gly-Pro-Ala-OH with that of the [M–D]� ion of thepeptide in which all the labile hydrogens were exchanged fordeuterium. Three major products are observed, the y1 ion,the y2 ion and m/z 141, which corresponds largely to loss
Scheme 7
of CO2 from the y2 ion with a possible minor contributionfrom loss of the glycyl residue from the a3 ion (which is notobserved with significant intensity). Bowie and coworkers24
reported y2 as the major fragment ion in the high-energyCID of deprotonated H-Gly-Pro-Ala-OH with more minorformation of the y1 ion and m/z 141; they also reportedobservation of the a3 ion. When the labile hydrogens wereexchanged for deuterium and the [M–D]� ion subjected tocollisional activation, they reported that the y1 ion retainedone labile hydrogen and that the y2 ion retained two labilehydrogens; however, one should bear in mind the limitedmass resolution of mass-analyzed ion kinetic spectroscopy(MIKES) experiments.36 Our results (Fig. 2) show that they1 ion retains in part one labile hydrogen and, in part, twolabile hydrogens, similar to the results observed for the y1
ion derived from H-Pro-Gly-Gly-OH (Fig. 1 and Scheme 6).Rather surprisingly, under our experimental conditions, they2 ion incorporates, to a significant extent, only one labilehydrogen. A plausible pathway that leads to incorporationof only one labile hydrogen is outlined in Scheme 7 wherea hydrogen initially bonded to carbon is incorporated inthe product ion; although this pathway suggests protonmigration in the carboxyl-deprotonated species, it couldequally well occur for the species in which the C-terminalamide hydrogen is abstracted in the ionization process. Thedifference in results in our study and that of Bowie andcoworkers probably reflects differences in the deprotonatedspecies formed by FAB ionization (Bowie) and electrosprayionization (present work) as discussed above.
Table 6 records the CID mass spectra of deprotonated H-Ala-Ala-Pro-OH and the a3 and 00b2 ions derived therefrom,obtained on the Q-ToF instrument at 20 eV collision energy.There are two primary fragmentation reactions of the[M–H]� ion, loss of CO2 to form the a3 ion and formation ofthe y1 ion, deprotonated proline. The major, if not the only,route to the 00b2 ion is by loss of neutral pyrrolidine (C4H9N)from the a3 ion as discussed previously.34 The a3 ion alsofragments to a minor extent to form the c1 ion at m/z 87.Collisional activation of the 00b2 ion results in formation of
Copyright 2005 John Wiley & Sons, Ltd. J. Mass Spectrom. 2005; 40: 1173–1186

1178 A. G. Harrison and A. B. Young
Figure 2. Comparison of CID mass spectra of [M–H]� of GPA and [M–D]� of GPA-d4. 55 V cone voltage.
Table 6. CID mass spectra of anions derived fromH-Ala-Ala-Pro-OH (20 eV collision energy)
m/z (ion) [M–H]� (m/z 256) a3 (m/z 212) 00b2 (m/z 141)
212 (a3) 69.0141 (’’b2) 53.6 100124 5.2 21.4114 (y1) 100113 26.0 32.6111 16.3 100
97 6.9 93.787 (c1) 4.8
four products, m/z 124 (NH3 loss), m/z 113 (CO loss), m/z111 (H2,C,O loss) and m/z 97 (C2,H4,O loss) as confirmedby accurate mass measurement. The implications of theseobservations in terms of the structure(s) of 00b2 ions will bediscussed below.
Table 7 summarizes the CID mass spectra of deproto-nated H-Leu-Ala-Pro-OH and the a3 and 00b2 ions derivedtherefrom. The two primary fragmentation reactions of the[M–H]� ion involve loss of CO2 to form the a3 ion and forma-tion of the y1 ion, while the dominant fragmentation reactionof the a3 ion involves loss of pyrrolidine to form the 00b2 ion.The latter ion, in turn, fragments by loss of NH3 (m/z 166),by loss of CO (m/z 155), by loss of [H2,C,O] (m/z 153) and by
Table 7. CID mass spectra of anions derived fromH-Leu-Ala-Pro-OH (20 eV collision energy)
m/z (ion) [M–H]� (m/z 298) a3 (m/z 254) ’’b2 (m/z 183)
254 (a3) 49.6183 (’’b2) 37.4 100166 5.3 22.2155 3.7 26.6 54.5153 2.2 14.4 100139 2.9 35.7129 (c1) 1.4122 7.8114 (y1) 100
loss of [C2,H4,O] (m/z 139) all neutral losses being confirmedby accurate mass measurement. In addition, the m/z 166([00b2 –NH3]�) ion fragments by loss of CO2 (confirmed byaccurate mass measurement) to give m/z 122.
Figure 3 compares the CID mass spectrum of deproto-nated H-Leu-Ala-Pro-OH with that of the [M–D]� ion of thepeptide in which the labile hydrogens have been exchangedfor deuterium. Three major ions are observed, the a3 ion (m/z254), the 00b2 ion (m/z 183) and the y1 ion (m/z 114). Thedeuterium labelling shows that the 00b2 ion retains two labilehydrogens and the y1 ion retains one labile hydrogen whilethe a3 ion incorporates three labile hydrogens as expected.
Copyright 2005 John Wiley & Sons, Ltd. J. Mass Spectrom. 2005; 40: 1173–1186

Fragmentation of deprotonated peptides containing proline 1179
Figure 3. Comparison of CID mass spectra of [M–H]� of LAP and [M–D]� of LAP-d4. 40 V cone voltage.
These results are in agreement with an earlier study34 ofthe low-energy fragmentation of deprotonated H-Leu-Gly-Pro-OH and its deuterium-labelled analogue. Pathway 1of Scheme 6 would lead to no labile hydrogens incorpo-rated in y1 when proline is the C-terminal residue whilePathway 2 would lead to incorporation of one labile hydro-gen, as observed. However, it should be noted that Bowieand coworkers24 have reported that the y1 ion formed byhigh-energy CID of deprotonated H-Leu-Gly-Pro-OH retainsno labile hydrogens indicating dominant formation of thecarboxyl-deprotonated species in their case. Again, this prob-ably reflects different ionization mechanisms in FAB used byBowie and electrospray ionization used in the present work.
Table 8 summarizes the CID mass spectra of deproto-nated H-Ala-Phe-Pro-OH and the a3 and 00b2 ions derivedtherefrom. The two primary fragmentation reactions of the[M–H]� ion involve formation of the a3 ion and the y1 ion.The major fragmentation of the a3 ion involves loss of pyrro-lidine to form the 00b2 ion although there is also significantloss of the benzyl radical to form the fragment ion of m/z197, another example of side chain radical loss. The 00b2 ionshows minor loss of ammonia (formation of m/z 200) andformation of the benzyl anion (m/z 91), however, the majorfragment ion is observed at m/z 97 corresponding to loss ofa neutral or neutrals of 120 Da from the 00b2 ion; accuratemass measurement showed that the neutral(s) lost had the
Table 8. CID mass spectra of anions derived fromH-Ala-Phe-Pro-OH (25 eV collision energy)
m/z (ion) [M–H]� (m/z 332) a3 (m/z 288) ’’b2 (m/z 217)
288 (a3) 32.8217 (’’b2) 100 100200 2.1 5.2197 6.6 21.5189 3.7114 (y1) 85.5
97 5.3 26.7 10091 9.9 18.887 (c1) 6.0 9.0
elemental composition C8H8O (most likely C7H8 C CO).
Structure and fragmentation of ′′b2 ionsThe present study provides substantial information concern-ing the structure(s) and fragmentation of 00b2 ions. Theseobservations can be summarized as follows:
1. When proline (a secondary amine) is at the N-terminus,one labile hydrogen is retained in the 00b2 ion (Fig. 1).However, when there is a primary amino group at theN-terminus the 00b2 ion largely incorporates two labile
Copyright 2005 John Wiley & Sons, Ltd. J. Mass Spectrom. 2005; 40: 1173–1186

1180 A. G. Harrison and A. B. Young
hydrogens (Fig. 3 and Fig. 2 of Ref. 34). These observa-tions very strongly suggest that the labile hydrogensincorporated in the 00b2 ions are those of the N-terminalamine function.
2. On CID, the 00b2 ions show a significant peak correspond-ing to loss of NH3 (Tables 6, 7 and 8 and Ref. 34). Thisimplies the presence of a free amino group in the 00b2 ion.
3. A major fragmentation reaction of 00b2 ions is loss ofCO. In most cases, this is followed by elimination of H2.In addition, loss of R2H from [00b2 – CO]� frequently isobserved where R2 is the side chain that was originallyon the central amino acid residue of the tripeptide. In thecase of H-Ala-Phe-Pro-OH (Table 8), loss of C7H8 fromthe [00b2 – CO]� ion to produce m/z 97 is the dominantfragment ion observed and is accompanied by observationof the complementary [C7H7]� ion. The loss of R2H fromthe [00b2 – CO]� ion appears to be largely a specific reactionin that the 00b2 ion derived from H-Leu-Ala-Pro-OH shows(Table 7) loss of CH4 from the [00b2 – CO]� ion but notloss of C4H10 involving the butyl side chain of leucine.In the same vein, the [00b2 – CO]� ion derived from H-Gly-Leu-Sar-OH shows an abundant ion signal at m/z83 ([00b2 – CO–C4H10]�) while the 00b2 ion from H-Leu-Gly-Pro-OH shows only very minor formation of m/z83.34
The structure(s) of the 00b2 ions is (are) not clearlyestablished. Bowie and coworkers10,12,21 proposed the acyclicstructure a (Scheme 8), assumed to be formed directly fromthe [M–H]� ion. However, recent ab initio calculations37 haveshown that this structure rearranges over a low barrier to thedeprotonated oxazolone structure b. These computationalstudies on the simplest 00b2 ion, that derived from H-Gly-Gly-Xxx-OH, have shown that b is a stable structure but thatthe N-deprotonated diketopiperazine c is more stable by ca.9.5 kcal mol�1 and the C-deprotonated diketopiperazine d issimilar in energy to b. The computations also showed thatthe acyclic species f was similar in energy to the ion c. Theobservation of retention of two labile hydrogens in the 00b2 ion
Scheme 8
rules out f (one labile hydrogen retained, Scheme 8, Ref. 37)and c (one labile hydrogen retained) as likely structures for00b2 ions.
For tripeptides, the 00b2 ion is formed by loss of a neutralamine from the a3 ion.34 A plausible mechanism leading to the00b2 ion as a deprotonated oxazolone is presented in Scheme 9.This pathway leads to preservation of the N-terminal aminogroup and the hydrogens attached thereto, in agreementwith experimental observations. A related pathway leadingto elimination of a neutral amine from the a3 ion involvingformation of a deprotonated diketopiperazine is outlinedin Scheme 10. Such a pathway should lead to significantabstraction of a N-bonded hydrogen (the most acidic37)which would result in a much more pronounced ion signalcorresponding to retention of only one labile hydrogen. Thus,we conclude that the most likely structure for the 00b2 ion is adeprotonated oxazolone. It might be noted that the simplest‘‘b ion formed from deprotonated N-benzoyl peptides has
Scheme 9
Copyright 2005 John Wiley & Sons, Ltd. J. Mass Spectrom. 2005; 40: 1173–1186

Fragmentation of deprotonated peptides containing proline 1181
Scheme 10
been suggested38 to be a deprotonated oxazolone because ofthe observation of CO2 loss upon CID of the 00b ion. Such CO2
loss is not observed for the 00b2 ions studied in the presentwork, the loss of 44 Da from the 00b2 ions in Tables 6 and7 being shown by accurate mass measurement to be resultfrom loss of CO C CH4 not loss of CO2. Speculative pathwaysleading to the loss of R2H and H2 from the [00b2 – CO]� ionare presented in Scheme 11.
Fragmentation of larger proline-containingpeptidesTable 9 presents the CID mass spectrum of deprotonatedH-Ala-Ala-Pro-Ala-OH as well as that of the y2 ion derivedtherefrom. The dominant fragment ion derived from the[M–H]� ion is the y2 ion involving cleavage of theamide bond N-terminal to the proline residue. By contrast,tetraalanine, as well as other tetrapeptides containing onlyH or alkyl side chains, show only very minor formationof y2 ions on fragmentation of the [M–H]� ion.18 The y2
ion formed produces a CID spectrum very similar to thatobtained for deprotonated H-Pro-Ala-OH (Table 1). Figure 4compares the CID mass spectrum of deprotonated H-Gly-Gly-Pro-Ala-OH with that of the [M–D]� ion of the peptidein which the labile hydrogens have been exchanged fordeuterium. Again the y2 ion is very prominent and is seento incorporate, with almost equal probability, one or twolabile hydrogens, suggesting two different pathways to they2 product. A pathway analogous to that shown in Scheme 7would lead to incorporation of only one labile hydrogenin the y2 ion, while initial proton abstraction from the N-terminal amide nitrogen followed by elimination of a neutraloxazolone, analogous to Pathway 2 of Scheme 6, would leadto incorporation of two labile hydrogens in the y2 ion.
Table 10 summarizes the CID mass spectrum of depro-tonated H-Ala-Ala-Pro-Ala-Ala-OH. The y3 ion, as well as
Scheme 11
Table 9. CID mass spectra of anions derived fromH-Ala-Ala-Pro-Ala-OH (20 eV collision energy)
m/z (ion) [M–H]� (m/z 327) y2 (m/z 185)
283 (a4) 3.0238 (00b3) 3.4185 (y2) 100167 15.7141 (y2-CO2) 3.1 55.7139 24.3113 42.988(y1) 7.0 10072 7.1
[y3 – CO2]�, are prominent ions in the CID mass spectrumand involve cleavage N-terminal to proline. Again, by con-trast, CID of deprotonated pentaalanine produces only a veryminor signal for y3, the y1 ion being the base peak.18 In addi-tion, for AAPAA, there is an abundant c2 ion, also involvingcleavage N-terminal to proline. On the other hand, the CIDmass spectrum of H-Gly-Pro-Gly-Gly-OH shows (Table 11)
Copyright 2005 John Wiley & Sons, Ltd. J. Mass Spectrom. 2005; 40: 1173–1186

1182 A. G. Harrison and A. B. Young
Figure 4. Comparison of CID mass spectra of [M–H]� of GGPA and [M–D]� of GGPA-d5. 60 V cone voltage.
Table 10. CID mass spectrum of deprotonated (m/z 398)H-Ala-Ala-Pro-Ala-Ala-OH (25 eV collision energy)
m/z (ion) Rel intensity
309 (00b4) 45.1256 (y3) 100212 (y3-CO2) 49.8159 (y2) 7.9158 (c2) 19.888 (y1) 62.1
Table 11. CID mass spectra of deprotonated (m/z 285)H-Gly-Pro-Gly-Gly-OH
Relative intensity
m/z (ion) 20 eV 25 eV
241 (a4) 17.4 5.4228 (y3) 10.1 5.3210 (00b3) 46.4 20.8184 (a4-G), (y3-CO2) 100 100153 (00b2) 10.2 9.6131 (y2) 8.4 3.9113 6.8 7.687 (a4-G-P) 6.3 8.374 (y1) 8.2 6.3
only a relatively weak y3 ion signal, the main fragmentationchannel being formation of the a4 ion and further fragmenta-tion of this ion. It appears that one needs at least two aminoacid residues N-terminal to proline to achieve dominantcleavage of the amide bond N-terminal to proline possiblybecause a stable oxazolone or diketopiperazine neutral canbe formed in such cases. This also is evident in comparingthe CID mass spectra of deprotonated H-Xxx-Pro-Yyy-OHand H-Xxx-Yyy-Pro-OH as discussed above.
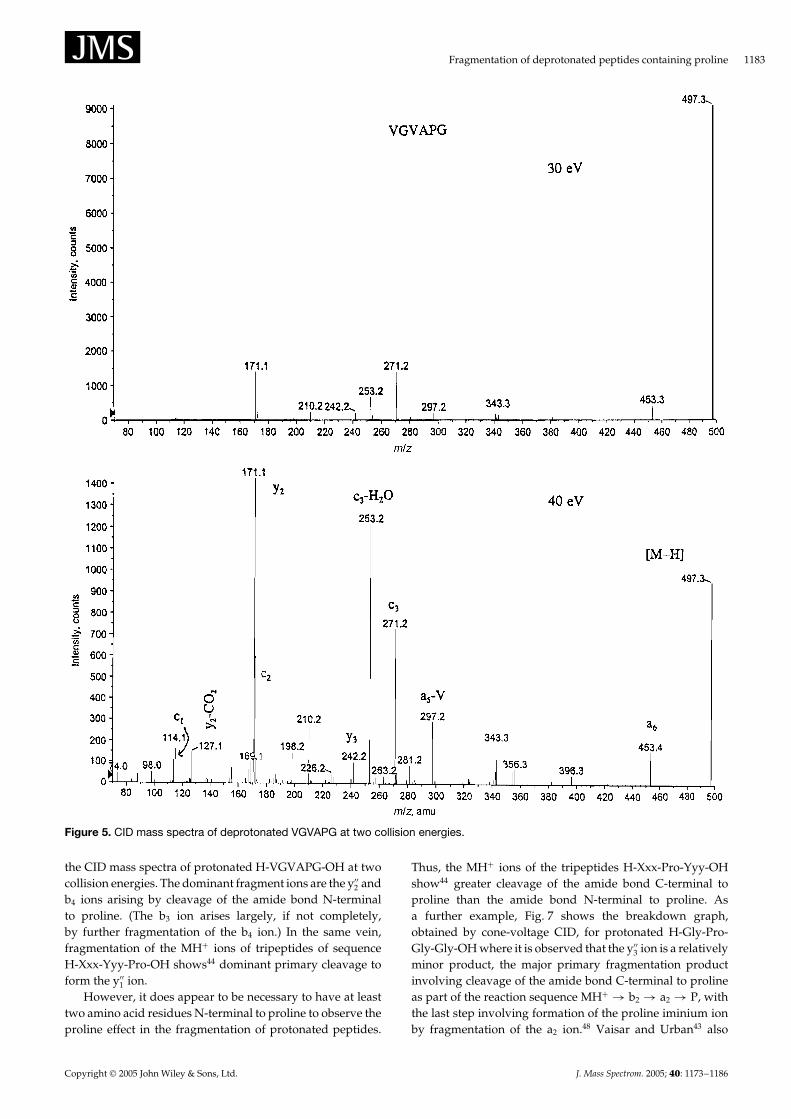
Figure 5 shows the CID mass spectrum of the [M–H]� ionof an even larger peptide, H-Val-Gly-Val-Ala-Pro-Gly-OH(H-VGVAPG-OH). A complex spectrum results, however,the y2 ion (m/z 171) remains the base peak. Of particularnote is the series of c ions at m/z 115 (c1), m/z 172 (c2), m/z271 (c3) and m/z 342 (c4, very weak) which provide sequenceinformation, although for a unknown peptide interpretationof the spectrum might be difficult.
The proline effectThe ‘proline effect’, i.e. the propensity for cleavage of theamide bond N-terminal to proline is well-established bothin the fragmentation of singly protonated peptides39 – 44 andin the fragmentation of multiply protonated peptides.45 – 47 Inthe present study, we observed that singly protonated H-Gly-Gly-Pro-Ala-OH, H-Ala-Ala-Pro-Ala-OH and H-Ala-Ala-Pro-Ala-Ala-OH fragmented almost exclusively by cleavageof the amide bond N-terminal to proline to give theappropriate yn’’ ion. As a further example, Fig. 6 shows
Copyright 2005 John Wiley & Sons, Ltd. J. Mass Spectrom. 2005; 40: 1173–1186
Fragmentation of deprotonated peptides containing proline 1183
Figure 5. CID mass spectra of deprotonated VGVAPG at two collision energies.
the CID mass spectra of protonated H-VGVAPG-OH at twocollision energies. The dominant fragment ions are the y00
2 andb4 ions arising by cleavage of the amide bond N-terminalto proline. (The b3 ion arises largely, if not completely,by further fragmentation of the b4 ion.) In the same vein,fragmentation of the MHC ions of tripeptides of sequenceH-Xxx-Yyy-Pro-OH shows44 dominant primary cleavage toform the y00
1 ion.However, it does appear to be necessary to have at least
two amino acid residues N-terminal to proline to observe theproline effect in the fragmentation of protonated peptides.
Thus, the MHC ions of the tripeptides H-Xxx-Pro-Yyy-OHshow44 greater cleavage of the amide bond C-terminal toproline than the amide bond N-terminal to proline. Asa further example, Fig. 7 shows the breakdown graph,obtained by cone-voltage CID, for protonated H-Gly-Pro-Gly-Gly-OH where it is observed that the y00
3 ion is a relativelyminor product, the major primary fragmentation productinvolving cleavage of the amide bond C-terminal to prolineas part of the reaction sequence MHC ! b2 ! a2 ! P, withthe last step involving formation of the proline iminium ionby fragmentation of the a2 ion.48 Vaisar and Urban43 also
Copyright 2005 John Wiley & Sons, Ltd. J. Mass Spectrom. 2005; 40: 1173–1186

1184 A. G. Harrison and A. B. Young
Figure 6. CID mass spectra of protonated VGVAPG at two collision energies.
Figure 7. Breakdown graph for protonated GPGG.
have reported that protonated H-Ala-Pro-Leu-Gly-OH givesthe b2 ion as the major primary fragment ion.
The ‘proline effect’ in positive ion mass spectrometry ofpeptides has been attributed41 to the high proton affinityof proline or, alternatively,8,42,47 to the strain involved in
the formation of a protonated bicyclic oxazolone49,50 uponfragmentation of the amide bond C-terminal to proline.Recent computations44 have shown that both factors playa role in favoring N-terminal cleavage over C-terminalcleavage in proline-containing protonated peptides, but thereclearly also is an effect of the neutral(s) lost in cleaving theN-terminal amide bond. When there are two amino acidresidues N-terminal to proline, a stable diketopiperazine51 ora stable oxazolone9,52,53 neutral is produced when the y00
n ion isformed. On the other hand, when there is only one amino acidresidue N-terminal to proline, the neutrals accompanying y00
n
formation are an imine plus CO9,54 making the fragmentationreaction more energy-demanding. However, in the negativeion mode, when there is only one amino acid residueN-terminal to proline, the neutral accompanying y ionformation is most likely (Scheme 7) a ketene that is morestable than an imine plus CO. Thus, the proline effect isstill observed in the negative ion mode even when there isonly one amino acid residue N-terminal to proline as for thetripeptides H-Xxx-Pro-Yyy-OH discussed above.
The origin of the proline effect in the negative ion modeis not entirely clear. As initially formed, the y ions arepostulated to be N-deprotonated species. It is likely thatthe secondary amine of proline is more acidic than theprimary amine of other amino acids. For example, we notethat dimethyl amine is 7 kcal mol�1 more acidic than methylamine (N deprotonation in both cases).55 Thus, formationof the N-deprotonated species is likely to be more facile forthe proline-containing peptides. One also should note thatthe CID mass spectra of the y2 ions derived from H-Xxx-Pro-Ala-OH and H-Xxx-Xxx-Pro-Ala-OH are very similar to
Copyright 2005 John Wiley & Sons, Ltd. J. Mass Spectrom. 2005; 40: 1173–1186

Fragmentation of deprotonated peptides containing proline 1185
the CID mass spectrum of deprotonated H-Pro-Ala-OH. Inthe latter case, one would not expect the initial ionizationprocess to remove the proline amino hydrogen; it is likelythat rearrangement of the initially formed N-deprotonatedspecies to the more stable O-deprotonated species does occurfor y ions. The gas phase acidities (for both N- and O-deprotonation) and the barriers for proton shifts are understudy computationally.
CONCLUSIONS
The fragmentation reactions of deprotonated peptides con-taining proline have been elucidated through a combinationof MS2 and MS3 experiments, deuterium labelling and accu-rate mass measurements. The results show that there is asubstantial proline effect in that cleavage of the amide bondN-terminal to proline is prevalent and leads to the appro-priate y ion. However, it is clear that this proline effect isnot as pronounced as that observed in the fragmentation ofprotonated peptides containing proline. Further results arepresented indicating that the 00b2 ions formed from deproto-nated tripeptides have a deprotonated oxazolone structure.
AcknowledgementThis work was supported by the Natural Sciences and EngineeringResearch Council (Canada) through a research grant to AGH andan equipment grant to the Department of Chemistry, which madepossible the purchase of the QStar.
REFERENCES1. Hunt DF, Yates JR III, Shabanowitz J, Winston S, Hauer CR.
Protein sequencing by tandem mass spectrometry. Proc. Natl.Acad. Sci. U.S.A. 1986; 83: 6233.
2. Biemann K. Sequencing of peptides by tandem massspectrometry and high-energy collision-induced dissociation.Methods Enzymol. 1990; 193: 455.
3. Biemann K. Primary studies of peptides and proteins. InBiological Mass Spectrometry: Present and Future. Matsuo T,Caprioli RM, Gross ML, Seyama T (eds). John Wiley & Sons:New York, 1993; 275.
4. Papayannopoulos IA. The interpretation of collision-induceddissociation tandem mass spectra of peptides. Mass Spectrom.Rev. 1995; 14: 49.
5. O’Hair RAJ. The role of nucleophile-electrophile interactions inthe unimolecular and bimolecular gas-phase ion chemistry ofpeptides and related systems. J. Mass Spectrom. 2000; 35: 1377.
6. Schlosser A, Lehmann WD. Five-membered ring formationin unimolecular reactions of peptides: a key structuralelement controlling low-energy collision-induced dissociationof peptides. J. Mass Spectrom. 2000; 35: 1382.
7. Polce MJ, Ren D, Wesdemiotis C. Dissociation of the peptidebond in protonated peptides. J. Mass Spectrom. 2000; 35: 1391.
8. Wysocki VH, Tsaprailis G, Smith LL, Breci LA. Mobile andlocalized protons: a framework for understanding peptidedissociation. J. Mass Spectrom. 2000; 35: 1399.
9. Paizs B, Suhai S. Fragmentation pathways of protonatedpeptides. Mass Spectrom. Rev. 2005; 24: 508.
10. Waugh RJ, Bowie JH. A review of the collision-induceddissociation of deprotonated dipeptides and tripeptides. Anaid to structural identification. Rapid Commun. Mass Spectrom.1994; 8: 169.
11. Bradford AM, Waugh RJ, Bowie JH. Characterization ofunderivatized tetrapeptides by negative ion fast atombombardment. Rapid Commun. Mass Spectrom. 1995; 9: 677.
12. Bowie JH, Brinkworth CS, Dua S. Collision induced fragmenta-tion of the [M–H]� parent anions of underivatized peptides. An
aid to structure identification and some unusual negative ioncleavages. Mass Spectrom. Rev. 2002; 21: 87.
13. Jai-nkuknan J, Cassady CJ. Anion and cation post-source decaytime-of-flight mass spectrometry of small peptides: Substance P,angiotensin II and renin substrate. Rapid Commun. Mass Spectrom.1996; 10: 1678.
14. Jai-nkuknan J, Cassady CJ. Negative ion post-source decay time-of-flight mass spectrometry of peptides containing acidic aminoacid residues. Anal. Chem. 1998; 70: 5122.
15. Ewing NP, Cassady CJ. Dissociation of multiply chargednegative ions from hirudin (54–65), fibrinopeptide B and insulinA (oxidized). J. Am. Soc. Mass Spectrom. 2001; 12: 105.
16. Clipston NL, Jai-nkuknan J, Cassady CJ. A comparison ofnegative and positive ion time-of-flight post-source decay massspectrometry for peptides containing basic residues. Int. J. MassSpectrom. 2003; 222: 363.
17. Marzluff EM, Campbell S, Rodgers MT, Beauchamp JL. Low-energy dissociation pathways of small deprotonated peptides inthe gas-phase. J. Am. Chem. Soc. 1994; 116: 7787.
18. Harrison AG. Sequence-specific fragmentation of deprotonatedpeptides containing H or alkyl side chains. J. Am. Soc. MassSpectrom. 2001; 12: 1.
19. Roepstorff P, Fohlman J. Proposal for a common nomenclaturefor sequence ions in mass spectra of peptides. Biomed. MassSpectrom. 1984; 11: 601.
20. Biemann K. Contribution of mass spectrometry to peptide andprotein structure. Biomed. Environ. Mass Spectrom. 1988; 16: 99.
21. Eckersley M, Bowie JH, Hayes RN. Collision-induced dissocia-tion of deprotonated peptides: dipeptides and tripeptides withhydrogen and alkyl ˛-groups. An aid to structural determina-tion. Org. Mass Spectrom. 1989; 24: 597.
22. Harrison AG. Effect of phenylalanine on the fragmentation ofdeprotonated peptides. J. Am. Soc. Mass Spectrom. 2002; 13: 1242.
23. Harrison AG. Characterization of ˛- and �-glutamyl dipeptidesby negative ion collision-induced fragmentation. J. MassSpectrom. 2004; 39: 136.
24. Waugh RJ, Bowie JH, Gross ML, Vollmer D. Collision induceddissociation of deprotonated peptides. Dipeptides andtripeptides containing proline. Int. J. Mass Spectrom. Ion Processes1994; 133: 165.
25. Harrison AG. Energy-resolved mass spectrometry. Acomparison of quadrupole cell and cone voltage collision-induced dissociation. Rapid Commun. Mass Spectrom. 1999; 13:1663.
26. van Dongen WD, van Wijk JIT, Green BM, Heerma W,Haverkamp J. Comparison between collision induceddissociation of electrosprayed protonated peptides in the up-front region and in a low-energy collision cell. Rapid Commun.Mass Spectrom. 1999; 13: 1712.
27. Harrison AG. Fragmentation reactions of alkylphenylammonium ions. J. Mass Spectrom. 1999; 34: 1153.
28. Makowiecki J, Tolonen A, Uusitalo J, Jalonen J. Cone voltage andcollision cell collision-induced dissociation of triphenylethylenesof pharmaceutical interest. Rapid Commun. Mass Spectrom. 2001;15: 1506.
29. Bure C, Lange C. Comparison of dissociation of ions inan electrospray source or a collision cell in tandem massspectrometry. Curr. Org. Chem. 2003; 7: 1613.
30. Smith RD, Loo JA, Barinaga CJ, Edmonds CG, Udseth HR.Collisional activation and collision-activated dissociation oflarge multiply-charged polypeptides and proteins produced byelectrospray ionization. J. Am. Soc. Mass Spectrom. 1990; 1: 53.
31. Chen H, Tabei K, Siegel MM. Biopolymer sequencing usinga triple quadrupole mass spectrometer in the ESI nozzle-skimmer/precursor ion MS/MS mode. J. Am. Soc. Mass Spectrom.2001; 12: 846.
32. Chass GA, Marai CNJ, Harrison AG, Csizmadia IG. Fragmen-tation reactions of a2 ions derived from deprotonated dipep-tides – A synergy between experiment and theory. J. Phys. Chem.A 2002; 106: 9095.
Copyright 2005 John Wiley & Sons, Ltd. J. Mass Spectrom. 2005; 40: 1173–1186

1186 A. G. Harrison and A. B. Young
33. Styles ML, O’Hair RAJ. The [M–H–CO2]� anion of glycylglycine undergoes rearrangement in the gas phase. RapidCommun. Mass Spectrom. 1998; 12: 809.
34. Harrison AG, Siu KWM, El Aribi H. Amide bond cleavage indeprotonated tripeptides: a newly discovered pathway to 00b2ions. Rapid Commun. Mass Spectrom. 2003; 17: 869.
35. Bradford AM, Waugh RJ, Bowie JH, Vollmer DL, Gross ML.Collision-induced dissociation of deprotonated dipeptidemethyl esters containing H, alkyl or benzyl ˛ side chains. Int. J.Mass Spectrom. Ion Processes 1994; 136: 143.
36. Busch KL, Glish GL, McLuckey SA. Mass Spectrometry/MassSpectrometry. Techniques and Applications of Tandem MassSpectrometry. VCH Publishers Inc: New York, 1988.
37. Chass GA, Marai CNJ, Setiadi DH, Csizmadia IG, Harrison AG.A Hartree-Fock, MP2 and DFT computational study of thestructures and energies of 00b2 ions derived from deprotonatedpeptides. A comparison of method and basis set used on relativeproduct stabilities. J. Mol. Struct. (Theochem) 2004; 675: 149.
38. Harrison AG, Young AB. Fragmentation of deprotonated N-benzoylpeptides. Formation of deprotonated oxazolones. J. Am.Soc. Mass Spectrom. 2004; 15: 446.
39. Gaskell SJ, Reilly MH. Hybrid tandem mass spectrometry ofpeptides above mass 1000. Rapid Commun. Mass Spectrom. 1988;2: 188.
40. Bean MF, Carr SA, Thorne GC, Reilly MH, Gaskell SJ. Tandemmass spectrometry of peptides using hybrid and 4-sectorinstruments. A comparative study. Anal. Chem. 1991; 63: 1473.
41. Schwartz BL, Bursey MM. Some proline substituent effects onthe tandem mass spectrum of protonated pentaalanine. Biol.Mass Spectrom. 1992; 21: 92.
42. Vaisar T, Urban J. Probing the proline effect in CID of protonatedpeptides. J. Mass Spectrom. 1996; 31: 1185.
43. Vaisar T, Urban J. Gas-phase fragmentation of mono-N-methyl peptides. Analogy with solution-phase acid-catalyzedhydrolysis. J. Mass Spectrom. 1998; 33: 305.
44. Grewal RN, El Aribi H, Harrison AG, Siu KWM, Hopkinson AC.Fragmentation of protonated tripeptides: the proline effectrevisited. J. Phys. Chem. B 2004; 108: 4899.
45. Loo JA, Edmonds CG, Udseth HR, Smith RD. Primary sequenceinformation from intact proteins by electrospray ionizationtandem mass spectrometry. Science 1990; 248: 201.
46. Loo JA, Edmonds CG, Smith RD. Tandem mass spectrometryof very large molecules. 2. Dissociation of multiply chargedproline-containing proteins from electrospray ionization. Anal.Chem. 1993; 65: 425.
47. Tabb DL, Smith LL, Breci LA, Wysocki VH, Lin D, Yates JR. III.Statistical characterization of ion trap tandem mass spectra fromdoubly charged tryptic peptides. Anal. Chem. 2003; 75: 1155.
48. Harrison AG, Young AB, Schnoelzer M, Paizs B. Formation ofiminium ions by fragmentation of a2 ions. Rapid Commun. MassSpectrom. 2004; 18: 1635.
49. Yalcin T, Khouw C, Csizmadia IG, Peterson MR, Harrison AG.Why are B ions stable species in peptide mass spectra? J. Am.Soc. Mass Spectrom. 1995; 6: 1165.
50. Yalcin T, Csizmadia IG, Peterson MR, Harrison AG. Thestructure and fragmentation of Bn (n ½ 3) ions in peptide massspectra. J. Am. Soc. Mass Spectrom. 1996; 7: 233.
51. Nold MJ, Wesdemiotis C, Yalcin T, Harrison AG. Amide bonddissociation in protonated peptides. Structures of the N-terminalionic and neutral fragments. Int. J. Mass Spectrom. Ion Processes1997; 164: 137.
52. Paizs B, Suhai S, Harrison AG. Experimental and theoreticalinvestigation of the main fragmentation pathways of protonatedH-Gly-Gly-Sar-OH and H-Gly-Sar-Sar-OH. J. Am. Soc. MassSpectrom. 2003; 14: 1454.
53. Paizs B, Suhai S. Towards understanding the tandem massspectra of protonated oligopeptides 1: Mechanism of amidebond cleavage. J. Am. Soc. Mass Spectrom. 2004; 15: 103.
54. Paizs B, Schnoelzer M, Warnken U, Suhai S, Harrison AG.Cleavage of the amide bond of protonated dipeptides. Phys.Chem. Chem. Phys. 2004; 6: 2691.
55. Lias SG, Bartmess JE, Liebman JL, Holmes JL, Levin RD,Mallard WG. Gas-phase ion and neutral thermochemistry. J.Phys. Chem. Ref. Data 1988; 17(Suppl. 1): 1.
Copyright 2005 John Wiley & Sons, Ltd. J. Mass Spectrom. 2005; 40: 1173–1186





























