Embed Size (px)
Citation preview

Vox Sang. 16: 361-372 (1969)
Functional Properties of Stored Red Cells
C..H. DE VERDIER Department of Clinical Chemistry, University Hospital, Uppsala
During the last two decades many efforts have been made to improve storage conditions for erythrocytes. This has resulted in prolonged storage time and higher erythrocyte viability after transfusion. A great number of biochemical parameters has been studied but, except for viability, the functional adequacy of the product has not until recently been investigated. If the relations between function and biochemical parameters were well known, i t would be sufficient and in many cases simpler to make the biochemical studies only. Little is known, however, and these questions will be dealt with after a brief presentation of the biochemical changes taking place during storage.
I will restrict this introduction and only discuss two procedures for preservation, e.g. ACD- or supplemented ACD-solutions for storage at +4"C and low glycerol - liquid nitrogen systems for storage at
When an erythrocyte is stored in ACD-solution, the conditions for the erythrocyte is changed as indicated in figure 1,e.g. with respect to temperature and the concentration of glucose, hydrogen ion, lactate and adenine. These conditional changes will impose biochemical changes, some of which will be irreversible in the sense that the cells will have less chance to survive in the circulation; other changes will be reversible in the sense that they will be normalized sooner or later after reentry into the circulation.
Decreases in enzymic activities during storage must always be irreversible as the mature erythrocyte lacks the ability to synthesize proteins. Marked decrease in hexokinase and phosphofructokinase has been described by BLUM and HEIM [I] and by JACOBASCH and RADERECHT [2]. AKERBLOM et al. have carefully studied the treatment
-196°C.

362 Optimal Use of Human Blood
ACD- solulion
Temperature
Decrease in enzyme activities Effect on individual enzymee?
Effect on ATP consumption? (Na+-K+ pump?) Glucose concentration
Continuous decrease Not critical
Lactate concentration Continuous decrease Effect through increase in hydrogen ion concentration
Hydrogen ion concentration Irutam and continuous increase Inhibition of glycolysis Decrease in DPG concentration
Continuous decrease Decrease in ATP, ADP and AMP concentration
Instant decrease
Adenine concentration
Fig. 1. Transfer of erythrocytes from the circulation to ACD-solution a t $ 4 T means changes in conditions that will impose biochemical changes.
of the hemolysate before the analyses and have not been able to confirm any drastic decrease in the enzymic activities. The lowest measured value was 75% of the initial phosphofructokinase activity after 45 day6 in ACD solution [3]. The two well-known and most important biochemical changes are : 1. Rapid decrease in 2,3-diphos- phoglycerate (DPG). 2. Decrease in ATP. According to recent in- vestigations by ASKARI and RAO [4] and by BANASCHAK [S], these
,i' ADP
ATP 2.3-DPG
IMP
inosine bp hypoxanlhine Ji ribose 1-P
Fig. 2. Effect of ATP and 2,3-diphosphoglycerate on AMP deaminase.
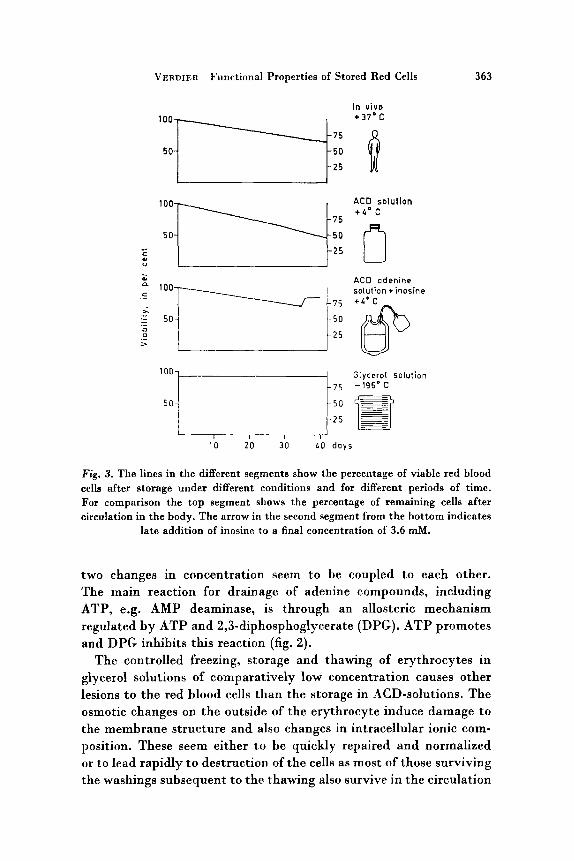
VERDIER Functional Properties of Stored Red Cells 363
In v i v o t 37' c
ACD solution t 4 O C
A C O adenine solution t inos ine
G l y c e r o l solu!ion - 196" C
10 20 30 60 d a y s
Fig. 3. The lines in the different segments show the percentage of viable red blood cells after storage under different conditions and for different periods of time. For comparison the top segment shows the percentage of remaining cells after circulation in the body. The arrow in the second segment from the bottom indicates
late addition of inosine to a final concentration of 3.6 rnM.
two changes in concentration seem to be coupled to each other. The main reaction for drainage of adenine compounds, including ATP, e.g. AMP deaminase, is through an allosteric mechanism regulated by ATP and 2,3-diphosphoglycerate (DPG). ATP promotes and DPG inhibits this reaction (fig. 2).
The controlled freezing, storage and thawing of erythrocytes in glycerol solutions of comparatively low concentration causes other lesions to the red blood cells than the storage in ACD-solutions. The osmotic changes on the outside of the erythrocyte induce damage to the membrane structure and also changes in intracellular ionic com- position. These seem either t o be quickly repaired and normalized or t o lead rapidly to destruction of the cells as most of those surviving the washings subsequent to the thawing also survive in the circulation
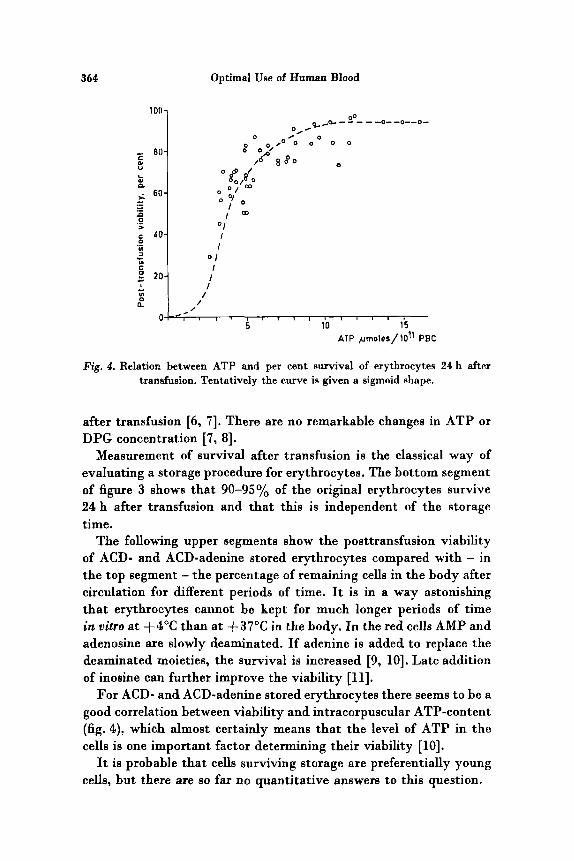
364 Optimal Use of Human Blood
00 - - 0
-0-
ATP ~rnoles/lO" PBC
Fig. 4. Relation between ATP and per cent survival of erythrocytes 24 h after transfusion. Tentatively the curve is given a sigmoid shape.
after transfusion [6, 71. There are no remarkable changes in ATP or DPG concentration [7, 81.
Measurement of survival after transfusion is the classical way of evaluating a storage procedure for erythrocytes. The bottom segment of figure 3 shows that 90-95y0 of the original erythrocytes survive 24 h after transfusion and that this is independent of the storage time.
The following upper segments show the posttransfusion viability of ACD- and ACD-adenine stored erythrocytes compared with - in the top segment - the percentage of remaining cells in the body after circulation for different periods of time. It is in a way astonishing that erythrocytes cannot be kept for much longer periods of time in vitro a t +4"C than a t +37"C in the body. In the red cells AMP and adenosine are slowly deaminated. If adenine is added to replace the deaminated moieties, the survival is increased [9, lo]. Late addition of inosine can further improve the viability [ll].
For ACD- and ACD-adenine stored erythrocytes there seems to be a good correlation between viability and intracorpuscular ATP-content (fig. 4). which almost certainly means that the level of ATP in the cells is one important factor determining their viability [lo].
It is probable that cells surviving storage are preferentially young cells, but there are so far no quantitative answers to this question.

VERDIER Functional Properties of Stored Red Cells 365
The main functions of the erythrocyte are listed in table I. I will soon return to the important question of gas transport but will first only mention that the erythrocytes complete an important mission as iron store and pH buffer. The first function is fairly independent of their viability, the second one is prevented by acid preservation solutions or accumulation of lactic acid.
Table I . Functions of the human erythrocytes
1. Oxygen transport (lung-tissue) 2. Carbon dioxide transport (tissue-lung) 3. Iron store 4. Buffer capacity
The first important discovery concerning the oxygen transport of stored erythrocytes was made by VALTIS and KENNEDY [12] who showed that erythrocytes stored for a week or a longer time in ACD- solution had an oxygen dissociation curve that was moved to the left, i.e. the hemoglobin had a higher affinity for oxygen. I n 1967, BENESCA and BENESCH [13] and CHANUTIN and CURNISH [l4] independently showed that phosphocompounds like ATP and DPG could move the S-shaped oxygen dissociation curve of hemoglobin solutions to the right. If one assumes that the periferal tissue wants t o keep a constant oxygen pressure, i t is evident that hemoglobin without DPG can deliver much less oxygen than hemoglobin with DPG can do.
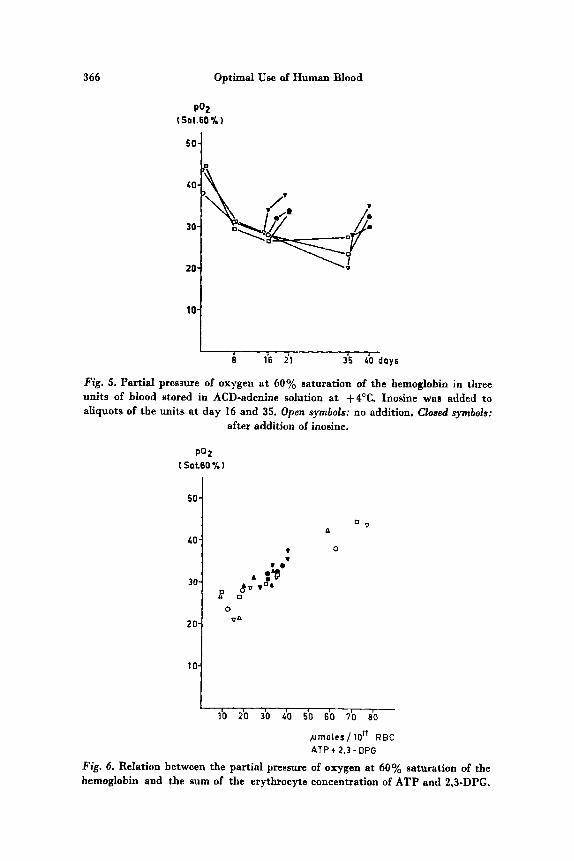
AKERBLOM et at. [15] have simultaneously investigated the con- centration of DPG and ATP and the position of the oxygen dissociation curve and have found the same shift as described by VALTIS and KENNEDY. In fig. 5 the position of the curve is expressed as PO, a t oxygen saturation 60%. We were able to reverse this shift by the addition of a DPG-generating substance as inosine. In fig. 6 is presented the relation between the position of the oxygen dissocia- tion curve (expressed as PO, at saturation 60%) and the sum of the molar concentrations of ATP and DPG. It is evident that there is a relationship between the content of the two phosphocompounds and the position of the oxygen dissociation curve but the clinical import- ance of this shift of the curve for stored erythrocytes is still an unanswered question. Investigations must follow several lines : The rate of resynthesis of DPG in erythrocytes reentering the circulation must be determined. The oxygen tension must be directly or in-
366
50-
40-
30-
20-
10-
Optimal Use of Human Blood
D V a
* 0 * *. & :'8
8 v v a h B n 0
v 4
PO2 1 Sat.6OX 1
I G
8 16 $1 35 40 days
Fig. 5. Partial pressure of oxygen a t 60% saturation of the hemoglobin in three units of blood stored in ACD-adenine solution a t +4"C. Inosine was added t o aliquots of the units a t day 16 and 35. Open symbols: no addition. Closed symbols:
after addition of inosine.
PO2 [ Saf.60 % I
flumoles/ 10" R B C ATP+ 2.3-OPG
Fig. 6. Relation between the partial pressure of oxygen at 60% saturation of the hemoglobin and the sum of the erythrocyte concentration of ATP and 2,3-DPG.
VERDIER Functional Properties of Stored Red Cells 361
c bottom
CtOP g-
8-
7-
6-
5-
4-
3-
A T P
5 6 7 8 PH
\
\ \
5 6 7 8 PH
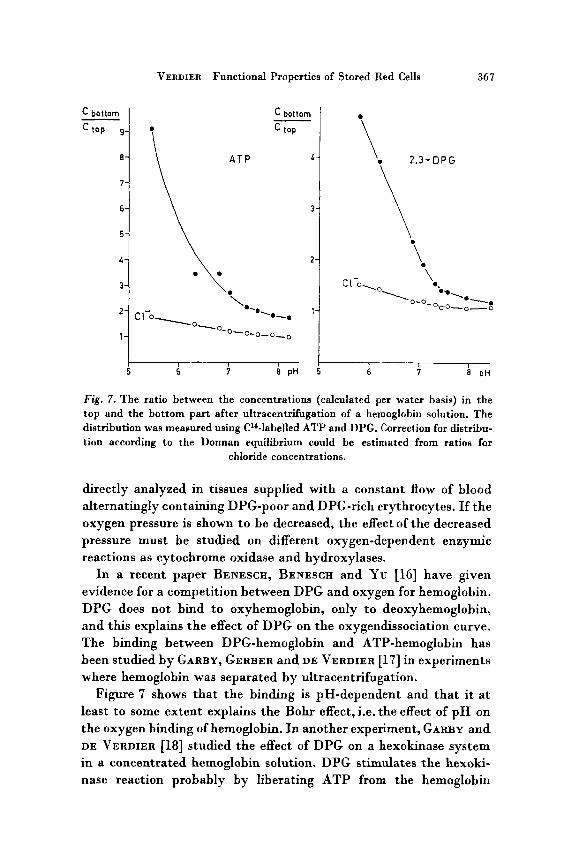
Fig. 7. The ratio between the concentrations (calculated per water basis) in the top and the bottom part after ultracentrifugation of a hemoglobin solution. The distribution was measured using C'*-labelled ATP and DPG. Correction for distribu- tion according to the Donnan equilibrium could be estimated from ratios for
chloride concentrations.
directly analyzed in tissues supplied with a constant flow of blood alternatingly containing DPG-poor and DPG-rich erythrocytes. If the oxygen pressure is shown to be decreased, the effect of the decreased pressure must be studied on different oxygen-dependent enzymic reactions as cytochrome oxidase and hydroxylases.
In a recent paper BENESCH, BENESCH and YU [16] have given evidence for a competition between DPG and oxygen for hemoglobin. DPG does not bind to oxyhemoglobin, only to deoxyhemoglobin, and this explains the effect of DPG on the oxygendissociation curve. The binding between DPG-hemoglobin and ATP-hemoglobin has been studied by GARBY, GERBER and DE VERDIER [17] in experiments where hemoglobin was separated by ultracentrifugation.
Figure 7 shows that the binding is pH-dependent and that it a t least to some extent explains the Bohr effect, i.e. the effect of p H on the oxygen binding of hemoglobin. In another experiment, GARBY and DE VERDIER [18] studied the effect of DPG on a hexokinase system in a concentrated hemoglobin solution. DPG stimulates the hexoki- nase reaction probably by liberating ATP from the hemoglobin

368 Optimal Use of Human Blood
molecules. We have here a fascinating system with a t least three components, oxygen, ATP and DPG competitively binding to the hemoglobin molecule and thus creating several intricate regulation mechanisms. One example of such a system may be the increased ion transport through the membranes of deoxygenated erythrocytes described by e.g. MCMANUS [19].
This new concept of interaction between enzymic cofactors, oxygen and hemoglobin will probably have an impact on the organization of erythrocyte preservation and distribution of blood products.
References
1. BLUY, K.-U. und HEIM, W.: Untersuchungen an liinger gelagerten Blut- konserven: Nucleosideffekt und Hexokinaseaktivitat. Blut 14: 76-82 (1966).
2. JACOBASCEI, G. und RADERECHT, H.-J. : Die Veranderung der Enzymkapazitaten von Hexokinase, Phosphofruktokinase und Puruvatkinase roter Blutzellen bei Konserviening in ACD-zitratfreien Stabilisatoren. Acta biol. med. german. 17:
3. AKERBLOM, 0.; ERIKSSON, A.; GARBY, L. and DE VERDIER, C.-H.: (unpublished results).
4. ASKARI, A. and RAO, S. N.: Regulation of AMP deaminase by 2.3-diphospho- glyceric acid: A possible mechanism for the control of adenine nucleotide metabolism in human erythrocytes. Biochim. biophys. Acta 151: 198-203 (1968).
5. BANASCHAK, H. : Die Aktivierung der Adenylsauredesaminase irn Erythrozyten durch Nukleotide. Folia haemat., Lpz. (in press).
6. KRIJNEN, If. W.; DE WIT, J. J. FR. M.; KUIVENEIOVEN, A. C. J.; Loos, J. A. and PRINS, H. K.: Glycerol treated human red cells frozen with liquid nitrogen. Vox Sang. 9: 559-572 (1964).
7. AKERBLOM. 0.; DE VERDIER, C.-H.; ERIKSSON, A.; GARBY, L. and HOGMAN, C. F. : Rapid freezing of erythrocytes using glycerol in low concentration. Bibl. Haemat. 29: 814 (1968).
8. AKERBLOM, 0.: (Unpublished results). 9. SUGITA, Y. and SIMON, E. R.: The mechanism of action of adenine in red cell
preservation. J. clin. Invest. 44: 629-642 (1965). 10. AKERBLOM,, 0.; DE VERDIER, C.-H.; FINNSON, M.; GARBY, L.: HOGMAN, C. F.
and JOEIANSSON, S. G. 0.: Further studies on the effect of adenine in blood preservation. Transfusion 7: 1-9 (1967).
11. AKERBLOM,, 0.; DE VERDIER, C.-H.; ERIBSSON, A.; GARBY, L. and HOGMAN, C. F.: Supplementation with adenine and inosine to stored blood. Effects on erythrocyte viability and metabolism. Folia haemat., N.F. (in press).
12. VALTIS, D. J. and KENNEDY, A. C.: Defective gas-transport function of stored red blood-cells. Lancet 1954 i : 119-124.
13. BENESCH, R. and BENESCH, E.: The effect of organic phosphates from the human erythrocyte on the allosteric propertiee of hemoglobin. Biochem. bio- phys. Res. Comm. 26: 162-167 (1967).
275-284 (1966).

VERDIER Functional Properties of Stored Red Cells 369
14. CMNUTIN, A. and CURNISK, R. R. : Effect of organic and inorganic phosphates on the oxygen equilibrium of human erythrocytes. Arch. Biochem. Biophys.
15. ~ C E R B L O M , 0.; DE VERDIER, C.-H.; GARBY, L. and HOGMAN, C. F.: Restoration of defective oxygen-transport function of stored rcd blood cells by addition of inosine. Scand. J. din. Lab. Invest. (in Press).
16. BENESCH, R.; BENESCH, R. E. and Yu, C. I.: Reciprocal binding of oxygen and diphosphoglycerate by human hemoglobin. Proc. nat. Acad. Sci., Wash. 59:
121: 96-102 (1967).
526-532 (1968). 17. GARBY, L.; GERBER, G. and DE VERDIER, C.-H.: (unpublished work). 18. GARBY, L. and DE VERDIER, C.-H.: Binding of ATP and 2,3-DPG to herno-
globin: Effect on erythrocyte glycolysis. Folia haemat., N.F. (in prcss). 19. McMANus,T.: Comparative biology of red cells. Fed. Proc. 26: 1821-1826
(1967). 20. BENESCK, R. and BENESCH, R. E.: Oxygenation and ion transport in red cells.
Science 160: 83 (1968). 21. ZURCHER, C.; LOOS, J. A. and PRINS, H. K.: Hereditary high ATP content of
human erythrocytes, Proc. 10th Congr. int. SOC. Blood Transf., Stockholm 1964;
22. BEUTLER, E. and DURON, 0.: Effect of pH on preservation of red cell ATP. Transfusion 5: 17-24 (1965).
23. BEUTLER, E. and DURON, 0.: The preservation of red cell ATP: The effect of phosphate. Transfusion 6: 124-129 (1966).
24. WOOD, L. and BEUTLER, E.: The viability of human blood stored in phosphate adenine media. Transfusion 7: 401-408 (1967).
25. G~NTEERBERC, €I. and ROST, J.: The true oxidized glutathione content of red blood cells, obtained by new enzymic and paperchromatographic methods. Analyt. Biochem. 15: 205-210 (1966).
26. ELLMAN, G. L. and LYSXO, H.: Disulfide and sulfhydryl compounds in TCA extract of human blood and plasma. J. Lab. clin. Med. 70: 518-527 (1967).
27. SRIVASTAVA, S. K. and BEUTLER, E. : Analyt. Biochem. (in press, 1968).
pp. 549-556 (1965).
Author’s address: Dr. C. H. DE VERDIER, Department of Clinical Chemistry, Uni- versity Hospital, Uppsala (Sweden).
Discussion
E. BEUTLER. DE VERDIER has raised the very important question of whether transfused cells, which may be viable but have a very low 2, 3 DPG level, actually can carry oxygen t o the tissues. As a matter of fact, we can be quite sure that they do not. The question then comes up, as t o how soon they will he able to carry oxygen to the tissues: will they do it in 10 min, or after 1 h, or does i t take several hours before they regenerate enough 2, 3 DYG so they can release oxygen when they get to the tissues.

370 Optimal Use of Human Blood
We have carried out very preliminary studies in an in uitro system, which we hope to extend to an in uiuo system shortly. These studies suggest that the regenera- tion is fairly rapid; that within an hour or so, stored red cells which have been partially depleted of DPG, regenerate substantially their 2, 3 DPG levels. But these results are very preliminary and need to be carried out under more physiologic conditions and particularly they need to be carried out in uiuo.
C. H. DE VEIIDIER. There must be a minimum value for the time needed for resynthesis of DPG. In fresh red cells the DPG concentration is about 4 mM, and the rate of glucose consumption is about 2 mmoles per liter blood cells per hour. That means that maximally 4 mmoles of DPG can be synthetised per liter blood cells per hour. But I do not think that all of the glucose could go to DPG, because then there would be only consumption and no production of ATP in the red cell which would thus very rapidly be depleted of ATP. I would guess that maximally 25% of the glucose can be converted to DPG. Therefore resynthesis of DPG must be a question of hours, a t least more than an hour.
T. J. GREENWALT. VALTIS and KENNEDY [12] were the first ones to study how quickly the oxygen dissociation power of hemoglobin recovered and they showed it was a matter of hours. I think that the clinical experience in the past indicates, that i t must be fairly rapid, because huge amounts of preserved blood have been given and have functioned in patients. We know this from clinical experience. And as BENESCH and BENESCH [20, 161 have pointed out, there is no effect until the value drops below about two thirds of the normal level.
E. BEUTLER. I think that the reason for the good clinical results may be that the depletion of 2, 3 DPG gets to the level that Dr. GREENWALT mentioned only after ten days or two weeks of storage. Maybe the surgeons who keep insisting on having fresh blood knew something, because until now we have been focusing all our attention on the viability of infused cells. That is what an 21 days of ACD blood dating is based on. But probably only a small proportion of the blood given is really 21 days old, so that when an individual gets massive infusion of blood, some of it is quite fresh. For this reason one does not observe clinically the effect that one would predict if 2, 3 DPG-depleted blood were being given. C. H. DE VERDIER. Of course I do not mean that stored red cells with low DPG
are useless as oxygen carriers, but i t may be that in a tissue with a low circulation, e.g. in a patient in shock, the oxygen tension of the cells in the tissue may fall under a critical level.
H.K.PRINs. In Holland we know a family with elevated pyruvate kinase activity as well as elevated ATP levels of the red cells, and this goes together with a very low content of 2, 3 DPG [Zl]. The only thing we can see in those patients is a somewhat increased hemoglobin level, therefore from this experience I would say that either the DPG content of the red cells is not so important, or that perhaps the increased level of ATP has a benificial effect on the oxygendissociation. As you mentioned already, ATP is a competitive binder to hemoglobin. Would you think that this double level of ATP has enough binding capacity for hemoglobin, so that it can take the place of DPG?
c. H. D E VERDIER. This family is an interesting experiment of nature, but I do not think we can draw a conclusion from it, for I suppose that your patients could easily increase their minute volume or the flow of blood to the tissues and I suppose you did not measure that. It is very difficult to design experiments in

VERD~ER Functional Properties of Stored Red Cells 371
this direction and I suppose that is the reason why nobody so far has done such experiments.
E. BEUTLER. With regard to the first part of Dr. DE VERDIER’S introduction I might say that we, too, looked at some of the enzymes (hexokinase, phospho- fructokinase) in ACD-adenine stored red cells. We quite agree with Dr. DE VERDIER on the basis of preliminary observations that we do not find any great changes in enzyme levels. I should like to comment also on the commonly made statement that ACD-blood is acidified blood.
According to our studies [22], the pH of blood collected in ACD formula B is approximately 7.4 or 7.5 a t 4°C. Many of the studies which have suggested that ACD-blood has a pH 6.8 or 7.0 are based on pH measurements a t 25°C; temperature greatly affects the pH of blood, causing a shift of about 0.5 in the pH. In our view, therefore, the effect of ACD is not to acidify the blood, but to prevent i t from becoming so alkaline that the hexokinase step outruns the rest of glycolysis and actually results in depletion of ATP.
With regard to the correlation of ATP levels and viability, we worked for a number of years, trying t o develop preservatives which optimise red cell ATP levels for a long period of time, so that we might be able to develop a preservative which would preserve cells for 60 or 70 days; and we can now present a preservative which maintains ATP levels for 50-70 days [23], but unfortunately in our hands [24] the correlation between viability and ATP levels breaks down after 45 or 50 days. I would like to ask Dr. DE VERVIER whether the very nice correlation which he found between ATP levels and viability was based on studies carried out after only 35 days of storage or whether it was a longer period.
C. H. DE VERDIER. Well, as you mentioned, the ATP level is one factor that is in correlation with viability, and I quite agree that there are certainly a lot of other factors and that they may be more significant after longer periods of storage. The points that we have in our correlation curve were all collected during a storage period of about 42 days; which was the maximum storage time.
H. W. KRIJNEN. Now I would like to invite Dr. BEUTLER, as the chairman of the Glutathione-workshop, to present a summarizing report of the activities of the workshop.
E. BEUTLER, I must say that, when Dr. PRINS first extended to me an invitation to participate in this workshop, I had some misgivings whether it would be possible to go into a strange laboratory with strange equipment and t o try to reproduce, within one or two days, rather complex techniques having to do with the estimation of reduced and oxydized glutathione. Still i t seemed that until we actually tried it we would not know whether such a thing could be done.
The problem which we approached was a very straightforward one: two groups had reported rather widely divergent results for levels of oxydized glutathione in red blood cells; one group had reported 40 pM 1251, another group about 140 p M [26] and after the workshop was organized a third group, attempting to compromise the matter, reported 5 pM as a correct level [27]. Thus the problem posed in the workshop was a clear-cut one.
One of the most interesting things that emerged from the workshop, that some- what surprized several of us, was that people who had to a considerable extent given the technical performance of laboratory tests over to technicians and assistants in

372 Optimal Use of Human Blood
the laboratory, were actually still able to work in the laboratory and in a large measure reproduce the results which their technicians obtained.
It was also possible in this type of workahop for the investigators who had re- ported different values, to work from aliquota of the very same blood sample prepared in the same way, and therefore to eliminate one of the possible sources of disagreement. It was aIso possibIe for us to quickly agree that the methods of estimation at the end of the preparation of the protein-free filtrate were the same. Thus the participants in the workshop were able to focus attention on the critical step which had to do with the alkylntion of reduced glutathione by N-ethylmaleimide and the method of preparation of the protein-free filtrate.
It is too early for me to present to you the final conclusion of the workshop, because although I think tremendous progeas has been made in these three or four days that we worked in the laboratory, there are still a number of experiments which remain to be done.
I think that one of the most important conclusions, which we can draw already, is that, after careful preparation, it is possible to carry out such a workshop. The staff of the Institute must be congratulated for making this type of a workshop possible. I believe that for carefully chosen biochemical topics i t has become obvious to me that such workshops can be very useful.





























