Embed Size (px)
Citation preview

Current Eye Research
Received on May 22, 1997; revised on July 24 and accepted on October 7, 1997
© 1998 Oxford University Press
Gelatinase concentration in tears of corneal-grafted patients
Claire D. Barro
1
, Jean-Paul Romanet
2
, Alaoui Fdili
2
, Michelle Guillot
1
and Françoise Morel
1
1
Laboratoire d’Enzymologie, Grepi Upres EA 2019 and
2
Service d’Ophtalmologie, Centre Hospitalier Universitaire de Grenoble,
B.P. 217-38043 Grenoble cedex 9, France
Abstract
Purpose.
Gelatinolytic enzymes, which degrade type IV base-ment membrane collagen, have been shown to be expressed bycorneal cells, either constitutively (gelatinase A or MMP-2) orafter induction (gelatinase B or MMP-9). Our aim was to deter-mine whether an enhanced MMP-9 and eventually MMP-2concentration in tears could be evidenced in the case of corneal-graft failure.
Methods.
The amount of MMP-2 and MMP-9 gelatinolyticenzymes was measured by quantitative zymography in tears oftwenty-one controls (84 samplings) and in tears of twenty-three corneal-grafted patients in a one-year post-graft follow-up study.
Results.
The mean MMP-2 values in controls were of 8.4(
±
7.3) pg/10
µ
g protein and the mean MMP-9 values in con-trols were of 73 (
±
76) pg/10
µ
g protein. No active gelatinaseform was detected in any of controls, but in all cases of cornealgraft failure, the active forms of both enzymes were present,and enzyme concentrations were higher than control values.All patients had significantly higher MMP-9 values than con-trols at each sampling time (p < 0.0001). The “corneal-graftfailure” patient group had statistically significant higher MMP-9 concentrations in tears than the “successful-graft” patientgroup at one month (p = 0.0312), four months (p = 0.0158) andone year (p < 0.01) after the graft. The presence of activeMMP-9 was highly significant of graft failure four months andone year after the graft (p < 0.0001). In contrast, MMP-2increase was delayed, with significantly higher MMP-2 valuesthan controls in all patients at four months (p = 0.0231) andone year (p = 0.0001) after the graft, but MMP-2 values couldnot discriminate between patient groups.
Conclusions.
In our study, all cases of graft failure showedabnormally high levels of the active forms of metalloproteinaseenzymes, and these values far exceeded the maximum controlconcentration. MMP-9 measurements in tears made between
one and four months after corneal transplantation, and while localcorticotherapy is steadily established, should help in predictingcorneal graft rejection. Curr. Eye Res. 17: 174–182, 1998.
Key words:
gelatinases; corneal transplantation; tears; human
Introduction
Gelatinases belong to the matrix metalloproteinase family ofenzymes (MMPs or matrixins). Matrixins are known to play anessential role in connective tissue remodelling, which occurs ina number of physiological processes such as trophoblastimplantation, embryogenesis, development and wound healing(1). The expression of these enzymes is tightly regulated, andtheir activity is specifically inhibited by tissular inhibitors(TIMPs) (2); abnormally high enzyme levels as a result ofenzyme/inhibitor imbalance leads to diverse pathological statessuch as rheumatoid arthritis, metastatic tumor progression orfibrotic diseases.
There are two members of the gelatinase subclass: the 72kDa gelatinase (gelatinase A or MMP-2) (3) and the 92 kDagelatinase (gelatinase B or MMP-9) (4). Gelatinolytic enzymesdisplay substrate specificity for type IV basement membranecollagen, and like all MMPs, are secreted as inactive zymo-gens. The loss of the 10 kDa propeptide after a conformationalchange or after cleavage by other proteases results in the activeform of the enzymes. The proenzyme forms of MMP-9 andMMP-2 bind to TIMP-1 and TIMP-2, respectively, while theactive enzymes are equally inhibited by both TIMP-1 andTIMP-2.
Matrix metalloproteinases and their inhibitors have beenshown by several authors to play a dramatic role in eye physi-ology and pathology (5, 6). They are present in the aqueoushumor (7), and they may play a role in controlling the aque-ous humor outflow (8). The increase in trabecular stromelysinand gelatinase B in response to laser trabeculoplasty accountsfor the efficiency of this treatment in open-angle glaucoma (8).TIMP-3 is expressed in the human retinal pigment epitheliumwhere it may be crucial for the maintenance of a healthy retina
Correspondence: Dr. Claire D. Barro, Laboratoire d’Enzymologie, CHU Greno-ble, B.P. 217-38043 Grenoble cedex 9, France; Email: francoise. [email protected]

Gelatinase concentration in tears of corneal-grafted patients
175
(9, 10). Both gelatinases have been shown to be secreted byuveal melanoma cell lines, providing clues for the mechanismsof tumor metastasis in uveal melanoma (11).
Gelatinolytic metalloproteinases were shown to be expressedby corneal cells (12, 13). Corneal stromal cells constitutivelyexpress the latent form of MMP-2 while MMP-9 expressionwas shown to be induced by corneal damage such as thermalburns. MMP-9 is synthesized primarily by the epithelial cellsof the upper corneal layer. Recently, failure of rat cornealcells to re-epithelialize after injury was found to be correlatedwith an over-expression of MMP-9, while interstitial collage-nase and stromelysin synthesis in injured rabbit corneas corre-lated more closely with stromal healing (14). Increased levels ofinterstitial collagenase, stromelysin 1, MMP-9 and TIMP-1 weredescribed in the aqueous humor of patients with uveitis (15).
Gelatinases may well be involved in corneal graft rejectionas well as in corneal ulceration or defective corneal trophicity,which represent some of the clinical features of corneal graftfailure. The aim of our study was to determine whether anenhanced MMP-9 and eventually MMP-2 secretion could befound in tears of corneal-grafted patients presenting the clini-cal signs of graft failure: corneal ulceration, deleterious trophi-cal defects, and corneal graft rejection.
Materials and methods
Tear sampling
Lachrymal fluid was collected, using the Schirmer assay proce-dure: the end of a sterile paper strip carefully placed in theinferior eyelid conjunctiva, so that the paper soaked up tears bycapillary action for five min. Sampling was effected at thesame time. Results in patients were compared to those in con-trol subjects.
Control subjects
The control group was made up of twenty-one healthy volun-teers, aged 23 to 45 (mean age: 32), of both sexes. Control sub-jects underwent a full ophthalmologic examination to ascertainthe presence of any conjunctival and corneal pathology (in par-ticular, infectious); they were devoid of any therapeutics. Tearsampling was effected twice in both eyes of each control sub-ject: the second sampling occured 48 to 72 h after the first one.Two samples from each eye constituted 84 samplings, whichwere considered control values.
Corneal-grafted patients
We investigated twenty-three patients who underwent a cornealgraft as treatment of corneal diseases of diverse etiologies,namely bullous keratopathy (8), keratoconus (4), previous cor-neal graft failure (5), keratitis sequellae (6) such as: post-zosterian keratitis (1), abcess sequellae (2), perforation (1) andalso constitutive dystrophies (2). The patient group ranged inage from 23 to 91 with a mean of 60 years old, and consisted often women and thirteen men: they were referred to as patient 1to patient 23, according to chronology of corneal trans-plantation. Sampling was done as previously described, at the
following rates: the day before the corneal graft (day 1), oneweek after the corneal graft (day 8), one month later (month1), four months later (month 4) and one year after the graft(month 12), representing five distinct samples from eachpatient. Patients underwent a complete clinical and ophthal-mologic examination at each sampling time, as well as acheck of significant biological parameters, to detect graftrejection, viral infection or intolerance to the immunosup-pressive therapy. The overall compliance was good but fourof our patients did not attend the year 1 follow-up: one ofthem (patient 11) because of a severe eye perforation whichled to an evisceration and the other three (patients 7, 15 and16) simply did not come.
Gelatin zymography
To evaluate the gelatinolytic activity, tear samples wereelectrophoresed under non-reducing conditions on an SDS-polyacrylamide gel copolymerized with gelatin as alreadydescribed (16). Gelatin was added to a final concentrationof 0.5 mg/ml to a 10% standard Laemmli acrylamide mixtureof the running gel. Lachrymal liquid was eluted from eachpaper strip by 500
µ
l of 150 mM Tris-HCl buffer (pH 7.6) con-taining 50 mM NaCl and 0.05% (w/v) polyoxyethylene 23 lau-ryl ether (Brij 35, Sigma). Protein content was determined bythe “enhanced-BCA” assay protocol (Pierce). For each patientsample, 1, 5 and 10
µ
g of protein were loaded into wells of a5% stacking gel, while for control samples 2.5, 5 and 10
µ
g ofprotein were loaded in the same conditions. More diluted sam-ples were used when enzyme activities were too high and didnot fit with the gelatinase standard curve. After electrophoresiswas completed, the gel was washed twice in 2.5% (w/v) TritonX-100 for 15 min at room temperature and incubated overnightat 37
8
C in 50 mM Tris-HCl buffer (pH 7.6) containing 5 mMCaCl
2
, 2 mM ZnCl
2
, 2% (w/v) Triton X-100 and 0.04% (w/v)NaN
3
. After incubation, the gel was stained for 20 min with0.25% (w/v) Coomassie blue R-250 in acetic acid-methanol-water (1:5:4), destained in acetic acid-ethanol-water (7:26:67).Gelatinolytic activity produces a clear band against thecoloured background of stained gelatin. When the gels wereincubated in the presence of a metal chelator (10 mM EDTA),the gelatinolytic bands disappeared, proving they were indeedmetalloproteinases. Molecular weight markers allowed us todetermine the relative molecular weight of each gelatinolyticband observed (Fig. 1A): two main bands were observed: at 92kDa, latent MMP-9 and at 68 kDa, latent MMP-2. Activatedforms were present as lower molecular weight bands: 82 kDafor MMP-9 and 62 kDa for MMP-2. High molecular weightforms (120–130 and 220 kDa) were complexed forms ofMMP-9, as indicated below.
Gelatinolytic bands were quantified by densitometry scan-ning (17) (Desaga CD 600 densitometer), referring to the linearpart of a purified gelatinase standard curve. Briefly, proteinsfrom human neutrophils extracellular medium after exocytosisfrom gelatinase granules were fractionated by DEAE-Sephacelchromatography and gelatinase was subsequently purified bygelatin-agarose chromatography (18). The protein amounts

176
C. D. Barro
et al.
used for the gelatinase standard curve were: 30, 50, 80, 100,150, 200, 500, 750 and 1000 pg. A curve was established foreach sample correlating protein
m
g deposited to a gelatinasequantity in pg: the final result was read in the linear part ofthe curve.
APMA activation of purified MMP-2 and MMP-9
Purified human MMP-2 (0.2 mg/ml, Valbiotech, France) andhuman neutrophil MMP-9 (0.15 mg/ml) (18), were activatedby p-aminophenyl-mercuric acetate (APMA, Sigma): briefly,10
µ
l of purified enzyme (corresponding to 800 pg MMP-2 and600 pg protein for MMP-9) were mixed with an equal volumeof 150 mM Tris-HCl (pH 7.6), 50 mM NaCl, 22.5 mM CaCl
2
containing 0.05% (w/v) Brij 35 and with 10
µ
l of 9 mMAPMA and subsequently incubated for 1 h at 37
8
C. The reac-tion was stopped by dilution in gel loading buffer, under nonreducing conditions. Purified enzymes, activated or not byAPMA (MMP-2 was already partially activated because offreeze-thaw cycles), were run on gelatin zymography in paral-lel with tear samples.
Western blotting
Proteins in tear samples were separated by SDS-PAGE (10%acrylamide) electrophoresis under reducing conditions as wellas coloured protein molecular weight standards (Amersham)and transferred to a 0.45
µ
m nitrocellulose membrane by elec-troblotting. Non-specific binding sites were blocked byimmersing the membrane in 0.5% (w/v) casein in 0.05 M Tris-HCl, 0.2 M NaCl, 0.05% (w/v) Tween-20 (TBS-Tween). Thenitrocellulose membrane was incubated with rabbit anti-humanMMP-9 polyclonal antiserum generated against purifiedMMP-9 (18) and diluted 1/200 in TBS-Tween for 2 h at roomtemperature. After washing in TBS-Tween, the membrane wassubsequently incubated with a 1/2000 dilution of horseradishperoxidase-labelled second antibody, for 1 h at room tempera-ture. Detection was performed with the Enhanced ChemiLumi-nescence (ECL) detection system (Amersham) as indicated bythe manufacturer, and the nitrocellulose membrane was exposedto Hyperfilm ECL (Amersham) for two s. Non-immune rabbitanti-serum (primary omission western) was used in the sameconditions (1/200 in TBS-Tween) and no band was evidenced(not shown). The same procedure was applied to detect a 25kDa protein of the lipocalin family (NGAL), associated withlatent MMP-9 (19, 20): we used a rabbit anti-recombinantNGAL antibody (1/1000 in TBS-Tween), which was kindlyprovided by Dr. Borregaard (19).
Statistical analysis
Control values were expressed as means
±
standard deviations(SD). Graphs were plotted using means
±
standard errors (SE).Comparisons were made using the non-parametric Mann-Whitney U test with p values corrected for
ex-aequo.
Results
MMP-2 and MMP-9 identification in tears
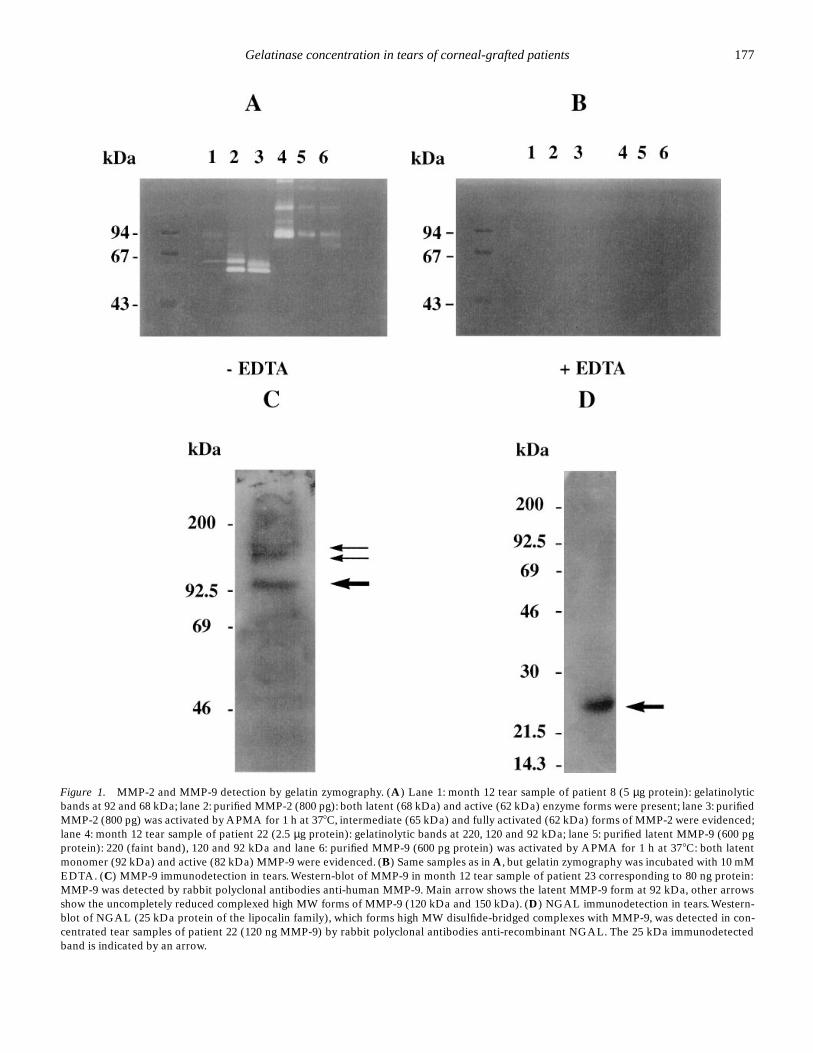
Figure 1A shows a gelatin zymography of purified enzymesrun in parallel with patients’ samples. Latent purified MMP-2had an apparent molecular weight (MW) of 68 kDa (lane 2)and the active form was already present at 62 kDa, as thisenzyme was partly activated by freeze-thaw cycles. AfterAPMA activation (lane 3), the gelatinolytic bands were at 65kDa (intermediate form) and at 62 kDa (active enzyme). Thepatient sample (lane 1) contained a major gelatinolytic band at68 kDa as latent MMP-2 and a faint one at 92 kDa correspond-ing to latent MMP-9: both gelatinolytic bands were regularlypresent in tears. Latent purified MMP-9 (lane 5) had a MW of92 kDa and a gelatinolytic band at 82 kDa appeared afterAPMA activation of the purified enzyme (lane 6) correspond-ing to activated MMP-9. High MW bands (120–135 kDa and220 kDa) corresponded to complexed forms of latent MMP-9:the 220 kDa was shown to be a disulfide-bridged homodimerof pro-MMP-9 while the 130 kDa form was identified as adisulfide-bridged complex of monomer and a 25 kDa protein(NGAL) which belongs to the lipocalin family (19, 20). Thepatient sample (lane 4) contained a strong gelatinolytic activityat 92 kDa and at 120 kDa and 220 kDa (regularly present intear samples too), which corresponded to the size of latentpurified MMP-9. All these gelatinolytic bands disappeared inthe control zymography incubated with 10 mM EDTA, provingthese activities were metalloproteinases (Fig. 1B).
MMP-9 in patient samples could be identified by Western-blotting with rabbit polyclonal antibodies directed againsthuman MMP-9 (Fig. 1C). MMP-2 in tears could not be identi-fied by immunoblotting because of the tiny amount of MMP-2in tears and of the limited available sample volume. The 25kDa NGAL protein was identified in tear samples (Fig. 1D) asa unique 25 kDa band under reducing conditions, with a rabbitanti-recombinant NGAL antibody.
Gelatinase concentration in tears of control subjects
The mean MMP-2 concentrations were of 8.4 (
±
SD 7.3) pg/10
µ
g protein while the mean MMP-9 concentrations were of 73(
±
SD 76) pg/10
µ
g protein. The distribution of MMP-2 andMMP-9 concentrations among controls are shown on Figure2A and B, respectively. More than 80% of the controls hadMMP-2 values lower than 20 pg/10
µ
g protein (Fig. 2A).Eighty-eight percent of the controls had MMP-9 values lowerthan 200 pg/10
µ
g protein (Fig. 2B). Variability of the sampleconcentration was overcome by the fact that each measurementwas compared to the same total protein quantity, as describedunder Materials and methods.
The most relevant feature is that none of the controls had anyactive form of the enzymes: only the latent, inactive form waspresent in control samples.
Gelatinase concentration in tears of corneal-grafted patients
177
Figure 1. MMP-2 and MMP-9 detection by gelatin zymography. (A) Lane 1: month 12 tear sample of patient 8 (5 µg protein): gelatinolyticbands at 92 and 68 kDa; lane 2: purified MMP-2 (800 pg): both latent (68 kDa) and active (62 kDa) enzyme forms were present; lane 3: purifiedMMP-2 (800 pg) was activated by APMA for 1 h at 378C, intermediate (65 kDa) and fully activated (62 kDa) forms of MMP-2 were evidenced;lane 4: month 12 tear sample of patient 22 (2.5 µg protein): gelatinolytic bands at 220, 120 and 92 kDa; lane 5: purified latent MMP-9 (600 pgprotein): 220 (faint band), 120 and 92 kDa and lane 6: purified MMP-9 (600 pg protein) was activated by APMA for 1 h at 378C: both latentmonomer (92 kDa) and active (82 kDa) MMP-9 were evidenced. (B) Same samples as in A, but gelatin zymography was incubated with 10 mMEDTA. (C) MMP-9 immunodetection in tears. Western-blot of MMP-9 in month 12 tear sample of patient 23 corresponding to 80 ng protein:MMP-9 was detected by rabbit polyclonal antibodies anti-human MMP-9. Main arrow shows the latent MMP-9 form at 92 kDa, other arrowsshow the uncompletely reduced complexed high MW forms of MMP-9 (120 kDa and 150 kDa). (D) NGAL immunodetection in tears. Western-blot of NGAL (25 kDa protein of the lipocalin family), which forms high MW disulfide-bridged complexes with MMP-9, was detected in con-centrated tear samples of patient 22 (120 ng MMP-9) by rabbit polyclonal antibodies anti-recombinant NGAL. The 25 kDa immunodetectedband is indicated by an arrow.
178
C. D. Barro
et al.
Gelatinase concentration in tears of corneal-grafted patients
MMP-2 values
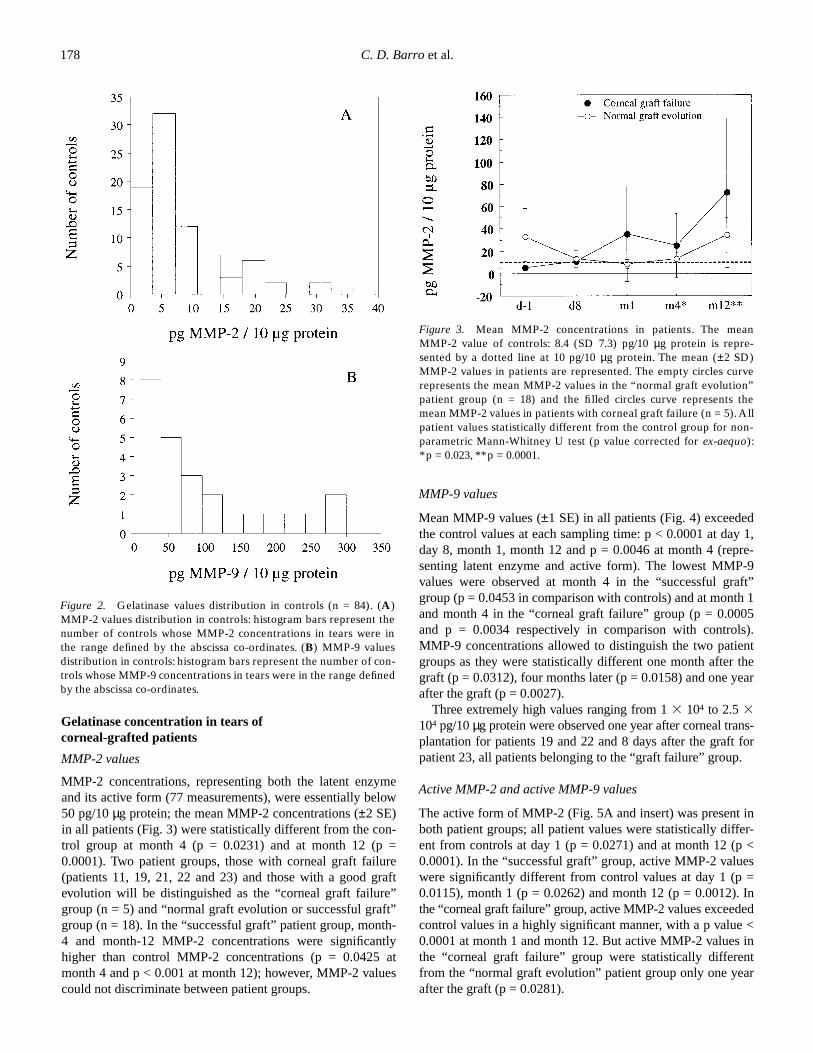
MMP-2 concentrations, representing both the latent enzymeand its active form (77 measurements), were essentially below50 pg/10
µ
g protein; the mean MMP-2 concentrations (
±
2 SE)in all patients (Fig. 3) were statistically different from the con-trol group at month 4 (p = 0.0231) and at month 12 (p =0.0001). Two patient groups, those with corneal graft failure(patients 11, 19, 21, 22 and 23) and those with a good graftevolution will be distinguished as the “corneal graft failure”group (n = 5) and “normal graft evolution or successful graft”group (n = 18). In the “successful graft” patient group, month-4 and month-12 MMP-2 concentrations were significantlyhigher than control MMP-2 concentrations (p = 0.0425 atmonth 4 and p < 0.001 at month 12); however, MMP-2 valuescould not discriminate between patient groups.
MMP-9 values
Mean MMP-9 values (
±
1 SE) in all patients (Fig. 4) exceededthe control values at each sampling time: p < 0.0001 at day 1,day 8, month 1, month 12 and p = 0.0046 at month 4 (repre-senting latent enzyme and active form). The lowest MMP-9values were observed at month 4 in the “successful graft”group (p = 0.0453 in comparison with controls) and at month 1and month 4 in the “corneal graft failure” group (p = 0.0005and p = 0.0034 respectively in comparison with controls).MMP-9 concentrations allowed to distinguish the two patientgroups as they were statistically different one month after thegraft (p = 0.0312), four months later (p = 0.0158) and one yearafter the graft (p = 0.0027).
Three extremely high values ranging from 1
3
10
4
to 2.5
3
10
4
pg/10
µ
g protein were observed one year after corneal trans-plantation for patients 19 and 22 and 8 days after the graft forpatient 23, all patients belonging to the “graft failure” group.
Active MMP-2 and active MMP-9 values
The active form of MMP-2 (Fig. 5A and insert) was present inboth patient groups; all patient values were statistically differ-ent from controls at day 1 (p = 0.0271) and at month 12 (p <0.0001). In the “successful graft” group, active MMP-2 valueswere significantly different from control values at day 1 (p =0.0115), month 1 (p = 0.0262) and month 12 (p = 0.0012). Inthe “corneal graft failure” group, active MMP-2 values exceededcontrol values in a highly significant manner, with a p value <0.0001 at month 1 and month 12. But active MMP-2 values inthe “corneal graft failure” group were statistically differentfrom the “normal graft evolution” patient group only one yearafter the graft (p = 0.0281).
Figure 2. Gelatinase values distribution in controls (n = 84). (A)MMP-2 values distribution in controls: histogram bars represent thenumber of controls whose MMP-2 concentrations in tears were inthe range defined by the abscissa co-ordinates. (B) MMP-9 valuesdistribution in controls: histogram bars represent the number of con-trols whose MMP-9 concentrations in tears were in the range definedby the abscissa co-ordinates.
Figure 3. Mean MMP-2 concentrations in patients. The meanMMP-2 value of controls: 8.4 (SD 7.3) pg/10 µg protein is repre-sented by a dotted line at 10 pg/10 µg protein. The mean (±2 SD)MMP-2 values in patients are represented. The empty circles curverepresents the mean MMP-2 values in the “normal graft evolution”patient group (n = 18) and the filled circles curve represents themean MMP-2 values in patients with corneal graft failure (n = 5). Allpatient values statistically different from the control group for non-parametric Mann-Whitney U test (p value corrected for ex-aequo):*p = 0.023, **p = 0.0001.

Gelatinase concentration in tears of corneal-grafted patients
179
Active MMP-9 was present (Fig. 5B and insert) in bothpatient groups (patients 11, 19, 22, 23 and with lower valuesfor patients 8, 14, 15) and was totally absent from the controlgroup. Active MMP-9 in all patients was significantly differentfrom the control group: the lowest values were observed at day1 (p = 0.0362) and month 4 (p = 0.0058) while active MMP-9significantly increased in all patients one week and one monthafter the corneal graft (p < 0.0001) and also one year later (p =0.0004). The two patient groups could be distinguished by theiractive MMP-9 concentrations: day 1 and month 4 values in the“normal graft evolution” group were not statistically differentfrom controls, while active MMP-9 concentrations in the “cornealgraft failure” group were significantly different from controls fromday 1 to month 12 (p < 0.0001). The most interesting result is thatmonth-4 and month-12 active MMP-9 values in the “graft failure”group were statistically higher from those of the “successful graft”group (p = 0.0004 and p = 0.0275, respectively).
Discussion
Mean MMP-9 values in patients’ tears were significantlyhigher than controls at each sampling time. Before the graft(day 1), this result was to be expected as all transplantationcandidates had corneal abnormalities such as keratitis, eye dry-ness, corneal dystrophy or ulceration. Day 1 MMP-9 valueswere not predictive of the graft issue as high MMP-9 values at
Figure 4. Mean MMP-9 concentrations in patients. The meanMMP-9 value of controls: 73 (SD 76) pg/10 µg protein is representedby a dotted line at 73 pg/10 µg protein. Mean (±1 SE) MMP-9patients values are represented with a logarithmic Y axis scale. Theempty circles curve represents the mean MMP-9 values in the “nor-mal graft evolution” patient group (n = 18) and the filled circlescurve represents the mean MMP-9 values in patients with cornealgraft failure (n = 5). *All patient values statistically different fromthe control group for non-parametric Mann-Whitney U test (p valuecorrected for ex-aequo): p < 0.0001. **Corneal graft failure groupsignificantly higher than normal graft evolution at m1: p = 0.0312,m4: p = 0.0158 and m12: p = 0.0027.
Figure 5. Active MMP-2 and MMP-9 in tears of corneal grafted patients. (A) Active MMP-2 in tears of corneal grafted patients. Mean (±2SE) active MMP-2 concentrations are represented by the empty circles curve in patients with a normal graft evolution and by the filled circlescurve in patients with corneal graft failure. *Corneal graft failure statistically different from normal graft (p = 0.0281). Insert: Gelatin zymogra-phy of tears (patient 11, day 30) showing the active band of MMP-2 at 62 kDa and the upper band at 68 kDa of latent MMP-2 (arrows); molec-ular weight markers are on the left side. (B) Active MMP-9 in tears of corneal grafted patients. Mean (±1 SE) active MMP-9 concentrations inpatients with a normal graft evolution (empty circles curve) and in patients with corneal graft failure (filled circles curve) are represented witha logarithmic Y axis scale. *Corneal graft failure statistically different from normal graft evolution at m4 (p = 0.0004) and m12 (p = 0.0275). Insert:Gelatin zymography of tears (patient 19, month 1): three main gelatinolytic bands (arrows) correspond to the high MW complexed form ofMMP-9 (120 kDa), latent MMP-9 (92 kDa) and activated MMP-9 (82 kDa), the upper band at 220 kDa is not pointed with an arrow; molecularweight markers are on the left side.

180
C. D. Barro
et al.
day 1 in patients 10 and 18 were not linked with an abnormalgraft evolution, although patients 19 and 23 had high MMP-9values at day 1 with an active form for patient 23 who sufferedfrom a post-zosterian keratitis with a bad graft evolution. Incontrast, day 1 MMP-2 values in patients were not statisticallydifferent from those in controls.
One week after the graft (day 8), MMP-2 values in patientswere not significantly different from controls, in contrast withMMP-9 values. Active MMP-9 was present for patients 22 and23. Patient 22 was grafted for a post-surgical bullous keratopa-thy: at the time of sampling, an eye dryness and a superficialpunctuated keratitis were observed. This early MMP-9increase was to be expected as the sampling was done close tothe surgical act, meaning local changes of the extra-cellularmatrix; the induction of MMP-9 secretion in the upper corneallayer accounts for its early detection in tears. The immunosup-pressive treatment was represented by a local corticotherapyand was identical for all patients except patient 21 who alsoreceived cyclosporin. Corticoids strongly inhibit matrix met-alloproteinases: the enzyme induction had to be very signifi-cant close to surgery in order to raise MMP-9 values in tears.This induced secretion of MMP-9 after surgery explains highconcentrations at day 8 in patients whose corneal graft wassuccessful.
One month after the corneal graft, MMP-2 concentrationswere still not significantly different from control MMP-2 val-ues, in contrast with MMP-9 concentrations in patients’ tears.Interestingly, active MMP-2 was significantly higher than con-trols in the “graft failure” group at month 1 sampling time andthis was illustrated in our study by patient 11 who had thehighest MMP-2 value at month 1 with an active form: the evo-lution was towards a severe eye perforation, which led to anevisceration. Patient 9 had high month 1 MMP-9 values: shewas 91 years old and had a reduced lachrymal secretion.
Four months after the graft, at distance from surgery andwhile the immunosuppressive treatment was maintained,MMP-2 values in patients’ tears raised and became signifi-cantly higher than controls while MMP-9 concentrations weredecreasing: MMP-2 levels evolution in patients’ tears did notparallel that of MMP-9 levels.
Active MMP-9 values were significantly higher in the “cor-neal graft failure” group compared to the other patient groupfour months and one year after the graft and, together withmonth-1, month-4 and month-12 global MMP-9 values,allowed us to discriminate between the two patient groups. Theone month time point seemed the most suitable in our study fordetection and/or prediction of graft failure. It was interesting toknow whether global MMP-9 values at one month could bepredictive of graft evolution. Table 1 shows a contingency tablewhere a threshold of 500 pg MMP-9/10
µ
g protein discrimi-nates between patient groups: the positive predictive value forgraft failure is 80% while the negative predictive value is 94%:this observation needs to be confirmed on a larger number ofpatients.
MMP-2 and active MMP-2 values at month 12 exceededcontrol values one year after the graft, when the immunosup-pressive treatment was stopped and subsequently, metallopro-
teinase inhibition. An additional sampling time after month 12would have answered whether MMP-2 and MMP-9 values inpatients’ tears decreased to control values or not.
The highest MMP-9 values linked to high MMP-2 valuesand active MMP-2 one year after transplantation were found inpatients 21, 22 and 23. Patient 21 had to undergo a new cornealgraft (and exhibited high MMP-9 values in spite of cyclosporintreatment) as the initial graft was ulcerated. Patient 23 whounderwent a corneal graft for a post-zosterian keratitis did notheal. Patient 22 had an endothelial graft rejection in the previ-ous months.
Although higher MMP-9 and MMP-2 concentrations werepresent in patients’ tears compared to controls, and the maxi-mal values correlated with healing defects or graft failure, fewof them were active forms of the enzymes: this could be partlyexplained by the fact that sampling was unique at one time andthe secreted enzyme may not be fully active at that precisemoment; moreover, tears reflect rather indirectly what is hap-pening in corneal cells.
It has been shown (12) that MMP-2 was the only gelatinasefound in normal, undamaged human cornea. The origin of theMMP-9 we observed in tears of healthy controls was probablypartly conjunctival, as the sampling method may pick up con-junctival cells as well as tears. In patients, MMP-9 of cornealorigin, when no conjunctival pathology was involved but the cor-nea damaged, would be present in tears in higher quantities thanthose found among controls. The link between high MMP-9 andMMP-2 values and eye dryness raises the question of a lympho-cytic infiltration of the conjunctiva with a likely slight involve-ment of the cornea. MMP-9 and MMP-2 probably become acti-vated by other proteases such as serine proteases or granzymessecreted by infiltrating T lymphocytes and cytotoxic T cells.
There are numerous relevant factors which, in conjunctionwith our indirect clinical observations, may help understandingof why enhanced gelatinase values in tears correlate with cor-neal damage and graft failure. The first one is the fact that theeye is an immune-privileged site due to the constitutive expres-sion of the Fas ligand (FasL) mRNA and protein in the cornealepithelium, endothelium, iris, ciliary body and retina (21). Pro-tection of vision from dangerous immune reactions is main-tained by apoptosis of undesired Fas positive cells, mediatedby Fas-FasL interactions (22). A functional soluble form ofFasL, which is a member of the TNF
a
family, has been
Table 1.
Contingency table for MMP-9 values at one month:if MMP-9 > 500 pg/10
µ
g protein, then “1,” else (<500 pg/10
m
g protein): 0. This table shows the frequencies observed forgraft failure when a threshold of 500 pg/10
µ
g protein is givenfor MMP-9 values at one month.
0 1 Totals
Graft failure 1 4 5Normal graft 16 1 17
Totals
17
5
22

Gelatinase concentration in tears of corneal-grafted patients
181
described in activated T lymphocytes. As membrane-boundTNF
a
was reported to be processed by metalloproteases, it hasbeen suggested that a similar protease may be involved in theprocessing of membrane-type FasL (23). Regulation of cellsurface FasL expression by metalloproteases has been con-firmed by others (24, 25). The authors stressed that activehuman soluble FasL may function as a pathological agent withsuch deleterious effects as systemic injury.
Secondly, it has been shown that human T lymphocytes,which are the main actors of the immune response in graftrejection, synthesize MMP-9 (26, 27). We already mentionedthe recent report (14) showing the significant role of MMP-9 inre-epithelialization defect after corneal injury. It would beinteresting to know whether the enhanced MMP-9 activitymeasured in tears could be responsible for an excessive solubleFasL secretion in the cornea, leading to tissue damage.
Thirdly, it appears clinically that problematic corneas aftergraft are essentially those which presented a neovasculariza-tion. A recent report (28) has brought evidence that theangiotensin-converting enzyme inhibitor, captopril, directlyinhibited neovascularization
in vivo
: this effect was shown tobe mediated by the ability of captopril to directly inhibit theendothelial-cell-derived MMP-2 and MMP-9. Both gelatinasesare required by endothelial cells to respond to an angiogenicstimulus and contribute to neoangiogenesis.
Taken together, all these processes suggest the dramaticinvolvement of MMP-2 and MMP-9 in corneal defects observedin several cases of graft failure, although many surroundingevents are required to result in corneal damage. The enhancedexpression and secretion of MMP-9 and MMP-2 by cornealdamaged cells may represent both an alert signal which mustbe checked and quite an easy target for local therapeutics (29–31).
Our study evidenced the predominant correlation betweenMMP-9 increase in tears and graft failure clinics: global MMP-9 values were significantly different in the “graft failure” groupcompared to the “normal graft” group from one month (p =0.0312) to one year (p = 0.0027) after the graft. The presenceof active MMP-9 was highly predictive of graft failure fourmonths (p = 0.0004) and one year (p = 0.0275) after the graft.
MMP-2 increase in tears was delayed compared to MMP-9and could not help discriminate between the “graft failure”patient group and the “normal graft evolution” patient group.In case of graft failure and in the eye-dryness syndrome, theuse of local metalloproteinase inhibitors would be helpful. Tearsampling (which has the significant advantage of non-invasivenessfor patients and ease of sampling), made between one monthand four months after corneal graft to determine MMP-9 andMMP-2 values, would be a simple test to help in predictinggraft evolution.
Acknowledgments
We thank Dr. Jean-Luc Bosson for his expert advice in statisti-cal analysis. We thank Pr. M. Mouillon and Pr. A. Hadjian forwelcoming us in the Ophthalmology Department and Enzy-mology Laboratory, respectively.
References
1. Matrisian, L. M. (1992) The matrix-degrading metallopro-teinases.
BioEssays
,
14,
455–463.2. Matrisian, L. M. (1994) Matrix metalloproteinase gene
expression.
Ann. N.Y. Acad. Sci.
732,
42–50.3. Collier, I. E., Wilhelm, S. M., Eisen, A. Z., Marmer, B. L.,
Grant, G. A., Seltzer, J. L., Kronberger, A., He, C., Bauer, E. A.and Goldberg, G. I. (1988) H-ras oncogene-transformedhuman bronchial epithelial cells (TBE-1) secrete a singlemetalloprotease capable of degrading basement membranecollagen.
J. Biol. Chem.
14,
6579–6587.4. Wilhelm, S. M., Collier, I. E., Marmer, B. L., Eisen,
A. Z., Grant, G. A. and Goldberg, G. I. (1989) SV-40-transformed human lung fibroblasts secrete a 92-kDatype IV collagenase which is identical to that secretedby normal human macrophages.
J. Biol. Chem.
264,
17213–17221.5. Girard, M. T., Matsubara, M., Kublin, C., Tessier, M. J.,
Cintron, C. and Fini, M. E. (1993) Stromal fibroblasts syn-thesize collagenase and stromelysin during long-term tis-sue remodeling.
J. Cell. Sci.
104,
1001–1011.6. Matsubara, M., Zieske, J. D. and Fini, M. E. (1991) Mech-
anism of membrane dissolution preceding corneal ulcer-ation.
Invest. Ophthalmol. Vis. Sci.
32,
3221–3237.7. Ando, H., Twinning, S. S., Yue, B. Y. J. T., Zhou, X., Fini,
M. E., Kaiya, T., Higginbotham, E. J. and Sugar, J. (1993)MMPs and proteinase inhibitors in the human aqueoushumor. Invest. Ophthalmol. Vis. Sci. 34, 3541–3548.
8. Parshley, D. E., Bradley, J. M. B., Samples, J. R., Vanbus-kirk, E. M. and Acott, T. S. (1995) Early changes in matrixmetalloproteinases and inhibitors after in vitro laser treat-ment to the trabecular meshwork. Curr. Eye Res. 14, 537–544.
9. Jones, S. E., Jomary, C. and Neal, M. J. (1994) Expressionof TIMP-3 mRNA is elevated in retinas affected by sim-plex retinitis pigmentosa. FEBS Lett. 352, 171–174.
10. Ruiz, A., Brett, P. and Bok, D. (1996) TIMP-3 is expressedin the human retinal pigment epithelium. Biochem. Bio-phys. Res. Commun. 226, 467–474.
11. Cottam, D. W., Rennie, I. G., Woods, K., Parsons, M. A.,Bunning, R. A. D. and Rees, R. C. (1992) Gelatinolyticmetalloproteinase expression patterns in ocular melanoma.Invest. Ophthalmol. Vis. Sci. 33, 1923–1927.
12. Fini, M. E. and Girard, M. T. (1990) Expression of col-lagenolytic/gelatinolytic metalloproteinases by normalcornea. Invest. Ophthalmol. Vis. Sci. 31, 1779–1788.
13. Fini, M. E. and Girard, M. T. (1990) The pattern of metal-loproteinase expression by corneal fibroblasts is altered bypassage in cell culture. J. Cell. Sci. 97, 373–383.
14. Fini, M. E., Parks, W. C., Rinehart, W. B., Girard, M. T.,Matsubara, M., Cook, J. R., West-Mays, J. A., Sadow, P. M.,Burgeson, R. E., Jeffrey, J. J., Raizman, M. B., Krueger,R. R. and Zieske, J. D. (1996) Role of matrix metallopro-teinases in failure to re-epithelialize after corneal injury.Am. J. Pathol. 149, 1287–1302.
15. Digirolamo, N., Verma, M. J., McLuskey, P. J., Lloyd, A.

182 C. D. Barro et al.
and Wakefield, D. (1996) Increased matrix metalloprotein-ases in the aqueous humor of patients and experimentalanimals with uveitis. Curr. Eye Res. 15, 1060–1068.
16. Zaoui, P., Barro, C. and Morel, F. (1996) Differentialexpression and secretion of gelatinases and tissue inhibitorof metalloproteinase-1 during neutrophil adhesion. Bio-chim. Biophys. Acta, 1290, 101–112.
17. Girard, M. E., Matsubara, M. and Fini, M. (1991) Trans-forming growth factor-b and IL-1 modulate expression ofmetalloproteinases by corneal stromal cells. Invest. Oph-thalmol. Vis. Sci. 32, 2441–2454.
18. Morel, F., Berthier, S., Guillot, M., Zaoui, P., Massoubre,C., Didier, F. and Vignais, P. V. (1993) Human neutrophilgelatinase is a collagenase type IV. Biochem. Biophys. Res.Commun. 191, 269–274.
19. Kjeldsen, L., Johnsen, A. H., Sengelov, H. and Borregaard,N. (1993) Isolation and primary structure of NGAL, anovel protein associated with human neutrophil gelatinase.J. Biol. Chem. 268, 10425–10432.
20. Kolkenbrock, H., Hecker-Kia, A., Orgel, D., Kinawi, A.and Ulbrich, N. (1996) Progelatinase B forms from humanneutrophils. Complex formation of monomer/lipocalinwith TIMP-1. Biol. Chem. 377, 529–533.
21. Griffith, T. S., Brunner, T., Fletcher, S. M., Green, D. R.and Ferguson, T. A. (1995) Fas ligand-induced apoptosisas a mechanism of immune privilege. Science, 270, 1189–1192.
22. Wilson, S. E., Li, Q., Weng, J., Barry-Lane, P. A., Jester,J. V., Liang, Q. and Wordinger, R. J. (1996) The Fas-Fasligand system and other modulators of apoptosis in thecornea. Invest. Ophthalmol. Vis. Sci. 37, 1582–1592.
23. Tanaka, M., Suda, T., Takahashi, T. and Nagata, S. (1995)Expression of the functional soluble form of human Fasligand in activated lymphocytes. EMBO J. 14, 1129–1135.
24. Mariani, S. M., Matiba, B., Bäumler, C. and Krammer,P. H. (1995) Regulation of cell surface APO-1/Fas (CD95)ligand expression by metalloproteases. Eur. J. Immunol.25, 2303–2307.
25. Kayagaki, N., Kawasaki, A., Ebata, T., Ohmoto, H., Ikeda,S., Inoue, S., Yoshino, K., Okumura, K. and Yagita, H.(1995) Metalloproteinase-mediated release of human Fasligand. J. Exp. Med. 182, 1777–1783.
26. Weeks, B. S., Schnaper, H. W., Handy, M., Holloway, E.and Kleinman, E. (1993) Human T lymphocytes synthe-size the 92 kDa type IV collagenase (gelatinase B). J. Cell.Physiol. 157, 644–649.
27. Montgomery, A. M. P., Sabzevari, H. and Reisfeld, R. A.(1993) Production and regulation of gelatinase B byhuman T cells. Biochim. Biophys. Acta, 1176, 265–268.
28. Volpert, O. V., Ward, W. F., Lingen, M. W., Chesler, L.,Solt, D. B., Johnson, M. D., Molteni, A., Polverini, P. J.and Bouck, N. P. (1996) Captopril inhibits angiogenesisand slows the growth of experimental tumors in rats. J.Clin. Invest. 98, 671–679.
29. Kenagy, R. D., Nikkari, S. T., Welgus, H. G. and Clowes,A. W. (1994) Heparin inhibits the induction of three matrixmetalloproteinases (stromelysin, 92-kD gelatinase, andcollagenase) in primate arterial smooth muscle cells. J.Clin. Invest. 93, 1987–1993.
30. Paemen, L., Martens, E., Norga, K., Masure, S., Roets, E.,Hoogmartens, J. and Opdenakker, G. (1996) The gelati-nase inhibitory activity of tetracyclines and chemicallymodified tetracycline analogues as measured by a novelmicrotiter assay for inhibitors. Biochem. Pharmacol. 52,105–111.
31. Amin, A. R., Attur, M. G., Thakker, G. D., Patel, P. D.,Vyas, P. R., Patel, R. N., Patel, I. R. and Abramson, S. B.(1996) A novel mechanism of action of tetracyclines:effects on nitric oxide synthases. Proc. Natl. Acad. Sci.USA, 93, 14014–14019.










![GRAFTED TOMATO - Iserv1].pdf · GRAFTED TOMATO Grafted onto ... Grafting joins the top part of one plant (the scion) to the root ... (TPIE) - January 18-20, 2012 Spring Trials in](https://img.pdfslide.net/doc/110x75/5aa1ea047f8b9a436d8c452d/grafted-tomato-1pdfgrafted-tomato-grafted-onto-grafting-joins-the-top-part.jpg)


















