Embed Size (px)
Citation preview

Gene expression in Lactococcus lactis
Maarten van de Guchte I, Jan Kok and Gerard Venerna
Department of Genetics. Centre of Biological Sciences. UniLersity of Groningen. Haren. The Netherlands
Received 24 April 1991
Revision received 28 June 1991
Accepted 2 Ju I y 1991
Key words: Lactococcus lactis; Gene expression signa!; Heterologous gene expression; Translatiol\alcoupling; Protein secretion
SUMMARY cherichia coli and Bacillus subtilis. This featureallowed the expression of a number of L. lactis-derived genes in the latter bacterial species. Sev-eral studies have indicated, however, that in spiteof the similarities, the expression signals from E.coli, B. subtilis and L. lactis are not equallyefficient in these three organisms.
Lactic acid bacteria are of major economicimportance, as they occupy a key position in themanufacture of fermented foods. A considerablebody of research is currently being devoted to thedeveIopment of lactic acid bacterial strains withimproved characteristics, that may be used tom'ke fermentations pass of more efficiently, or tomuke new applications possible. Therefore, andbecause the lactococci are designated 'GRAS'organisms ('generally recognized as safe') whichmay be used for safe production of foreign pro-teins, detailed knowledge of homologous and het-erologous gene expression in these organisms isdesired. An overview is given of our currentkIJwledge concerning gene expression in Lacto-coccus lactis. A general picture of gene expres-sion signals in L. lactis emerges that shows con-siderable similarity to those observed in Es-
2. INTRODUcrION
~dence lo~ G. Venema, Department of Genetics,Centre of Biological Sciences, University of Groningen, Kerk-C,J '0, 9751 NN Haren, The Netherlands.
p; 'ent address. National Food Biotechnology Centre, Uni-
versity ColleRe. Cork. lreland.
Lactic acid bacteria, including members of the
genera Lactobacillus, Lactococcus, Leuconostoc,Pediococcus, and Streptococcus, have long beenknown for their use in the manufacture of fer-mented foods. These include a broad range ofproducts derived from a variety of raw materialssuch as vegetables, cereals, me at and milk [1].The fermentations not only serve the preserva-tion of the food [2], they also add to the develop-ment of flavour and texture of the products [3].Moreover, some cultured milk products are dis-cussed to have certain health and additional nu-tritional benefits [4].
These features explain the major economicimoortance of the lactic acid bacteria for the food

industry, and the dairy industry in particular.Therefore, considerable efforts are made to un-derstand and improve the important characteris-tics of lactococcal starter strains. Increasing ourknowledge of genetics and physiology of theseorganisms will almost certainly lead to the devel-opment of starter strains with improved proper-ties, whereas the acquisition of more specificknowledge of the rules governing (heterologous)gene expression may be used to add desirablenew traits. In addition, since the lactococci aredesignated 'GRAS' organisms ('generally recog-nized as safe') [5], they may be used in newapplications for the production of commerciallyimportant heterologous proteins in a safe way.Depending on the nature and application of newlysynthesized proteins, several expression strategiesare conceivable. One option is to (over)producethe new protein intracellularly, af ter which thecells can be easily pelleted in order to harvest theprotein. The second option is to make the lacticacid bacteria secrete the new protein into theculture medium. For special applications, e.g. incheese making, a combined strategy may be pre-ferred, in which the protein is produced intracel-Iularly, and subsequently released by a deliverysystem based on autolysis or induced lysis of thehost [I].
In prokaryotes, transcription is immediatelyfolIowed by the translation of the nascent RNAmessenger [8,9]. Translation is initiated at theribosome binding site, which comprises the fol-lowing conserved elements: the Shine Dalgamo(SD) sequence that is complementary to the 3'-end of the 16S-rRNA (E. coli 16S-rRNA has the3'-end sequence: 3' AUUCCUCCA-5' [10]), andthe translation al start codon. These two elementsare usually separated by a window of approxi-mately 10 bases (from the central AGGA stringof the SD sequence to the start codon). In thewindow, and also immediately upstream of theSD sequence, A and U are preferred nucleotides[11]. In general, Gram-positive translation initia-tion sites have SD sequences that show a highercomplementarity to the 16S-rRNA sequence thanGrani-negative SD sequences [12].
3. GENE EXPRESSION IN PROKARYOTES
As in eukaryotes, gene expression in prokary-otes is governed by the processes of transcriptionand translation. In the cascade of events thatresults in the production of a protein, a promotersequence in the DNA mediates the interactionbetween the RNA-polymerase and the DNA toinitiate transcription. In Escherichia coli twohexameric sequences in the promoter area arewell-conserved: the so-calIed -35 region(TTGACA), and the -10 region (TATAAT) [6].Usually, these hexamers are separated by a se-quence of 17 :tl nucleotides, while the startingpoint of transcription is usually situated 7 :tlnucleotides downstream of the -10 hexamer.The same features are conserved in promoters
.active in vegetative cells of the Gram-positivebacterium Bacil/us subtilis m.
4. GENE EXPRESSION IN LACTOCOCCUSLACT1S
The first successful transformation of Lacto-coccus lactis ssp. lactis protoplasts with plasmidDNA was reported in 1982 [13]. Since that time,several modified transformation protocols havebeen described [14-18], including the now gener-ally preferred method of electrotransformation[19-22]. Plasmid-free host strains became avail-able such as the widely used L. lactis ssp. lactisstrains MGI363 [23] and ILI403 [24], allowing therapid analysis of transformation and gene cloning
experiments.General purpose gene cloning vectors were
developed, either based on cryptic L. lactis ssp.cremoris plasmids [25,26], or on non-lactococcalplasmids of Gram-positive origin [14,27]. The for-mer class of vectors offered the advantage ofbeing able to replicate in several hosts, includingE. coli and B. subtilis, thereby facilitating DNAmanipulation and selection procedures. Fromthese vectors, special purpose vectors were de-rived to studyspecific problems. Among these arepromoter-probe vectors [28-32], terminator-probevectors [32], vectors for the isolation of secretionsignals [33,34], expression vectors [35], secretionvectors [36], and vectors for integration into the
chromosome [37].

Owing to these developments, the species L./,cris became accessible to sophisticated geneticresearch techniques, and the cloning and analysisof a gradually increasing number of homologousgenes and expression signals became possible.The accumulating information on lactococcalgenes and their expression signals is currentlybeing used to establish the expression of heterol-()gous genes of interest in L. /acris. In this paper,l lta available on homologous and heterologousgene expression in L. /acris wil! be reviewed.
4.1. HomologolIS gene expression
4.1.1. Cloning and characterization of lactococcale.tpression signals using promoter and terminatorr ?be vectors
A straightforward approach to obtain moreinsight in the organization of gene expressionsignals emp!oys promoter probe vectors. The firstvector of this kind for use in L. lactis was de-scribed by Van der Vossen et al. [32,38]. Thisvector, pGKV210, contained the promoterlessch!oramphenico! acetyltransferase (cat-86) genef lm B. pumilus, preceded by a multiple cloningsite. In this multiple cloning site, chromosoma!DNA fragments obtained from L. lactis ssp. cre-moris Wg2 were inserted. The ligation mixtureswere used to transform L. lactis ssp. lactisMG1363, either direct I y or via precloning in B.subtilis. Subsequently, the chloramphenicolccetyltransferase activity in the transformants wasd.;termined. Some of the chromosomal DNA in-serts that acted as a promotel' for the expressionof the cat-86 gene were further characterized bynieans of DNA sequencing, and determination ofthe starting points of transcription. The mainconclusion that could be drawn from this workwas that the lactococcal promoter sequencescosely Iesembled the consensus E. coli and B.subtilis U43 promoter sequences. Because of thebroad host range nature of the plasmids used inthis study, it was possible to make a comparisonof chloramphenicol acetyltransferase activity lev-els in L. lactis and in B. subtilis. The activitylevels observed in B. subtilis were, on the aver-a~e, about 10 to 20 times higher than those in L.( 1is. However, since the overall expression level
was measured in this system, in addition to differ-ences in promoter activity also differences inplasmid copy number, mRNA stability, transla-tion (initiation) efficiency, or product stability maybe involved. Some of these considerations alsoapply to the comparison of different promoterswithin one strain of L. /actis using this system, asis evident from the results obtained by Koivula etai. [30], who isolated DNA fragments with pro-moter activity from the chromosome of L. /actisssp. /actis, in an approach similar to that used byVan der Vossen et al. Several promoters werecharacterized by DNA sequencing, mapping ofthe transcriptional starting points, measurementof CA T activities, and measurement of the rela'tive amounts of cat gene-specific mRNAs. Theresults showed a discrepancy between the relativepromoter strenghts in L. /actis as determined bythe measurement of CAT activities, and thosedetermined by means of mRNA measurements,indicating that CAT activity can only be used toobtain a rough initial estimate of promoterstrength. Furthermore, CA T activities obtained inL. /actis were substantially (about SOO-fold on theaverage) lower than in B. subti/is, while themRNA levels in both species were roughly thesame, indicating host-specific differences in trans-lation (initiation) efficiency, or product stability.
Several other promoter probe vectors compa-rable to pGKV210 have been described [28,29],that either employed the promoterless cat-86gene of B. pumi/is, or the cat-/94 gene of Staphy-/ococcus aureus. Recently, Simons et al. [31] de-scribed the vector pNZ336, that utilizes the ho-mologous phospho-/3-galactosidase (la cG) geneof L. /actis ssp. /actis.
Once a suitable promoter has been cloned invectors of the type described above, they can beconveniently used as terminator probe vectors, byinserting DNA fragments in between the pro-moter and the reporter gene. Van der Vossen[39] used this approach to show that the pre-sumed transcription terminator downstream ofthe L. /actis ssp. cremoris Wg2 protease gene [40]was functional in L. /actis ssp. /actis. Again, dataobtained from this kind of experiments should beinterpreted with care, because of the indirectmode of rneasurement of transcript formation:

Table 1
Lactococcus /actis-derived transcription initiation sites
TTTTTT~GAAGAAGGCGAAAAATGG~ATTTAGGT~CTGTT
CCTAAGACTG~AAAAGAGCAAATTTTGA~AGTATT~AATT
GTGAGCT~AGAAAAAAACTTCACAAAATGC~AGGT~GTAA
AGCTAAACTC~TACTTGATTTTATGT~AATTAATG~TGTA
ACATTAAATTC~GGGAGAGATAGGTTTGA~ATAAT~TTGT
AAAGAAAGAC~TTGTTGTTGAAAAATGC~ACATAA~TCCGA
TGACTACGAATCAGGCGGTTGTAGGTTCGAATCCTACCGCTTGCAIAAATA
AGTTGACC~AAAAACTGAAAATCTGT~AAATAATG~TT
TACTA~CAAAATTCTGTCAAATTTGA~AAGGTACG~GTAAG
TGTCATCAAGCGACCT~GGGCATTATGT~TAAGCT~TGAAG
ATTTTCG~TTGTTCTTCAATAGTATATAA~AGTAT~TAATA
ACCCTACGC~TAGTTAAGATTATATTA~ATTATAT~TATT
TAAACGGCT~AAATTCTGAAGTTTGTTAGA~GATT~C
TAACATTTG~CGAGTTTTATTTTTATATAATC~AGATITATAA
ACATTTG~AGTTTTATTTTTATATAATC~AGATTT~TAAAA
TCATA~TA~TGGGGTAGGAATAG~ATGTTT~TTCAA
TAATTTT~TTTTTTATTTGTTTTTTTA~AGATAAC~CCGTT
TCGAATTTTT~TATTTTCAAAGAATCCGT~CTAAC~ATC
TATGAAAAA~GATGTAACTCCGTTGAC~GGATGCA~AAGAT
TTCATTATT~ATCCTCACTAGTTATACA~ATTTGG~TTTTT
TATGGAAAAATA~AGCAAACTAAGGAGGG~GAATG~CCGAC
AAAATGTAAGA~TTACTAAAACAGTAACT~AACTG~AGGTA
1323222527134472445365553142451443001123426355
778598857573955X6646476779887796869FYY966X773Z
Y4689868X6996656997Y7647877898974X9657888866X2
1644024212351532114032512312001341203113201420
T TG T A T T AAA A T~
~ 17 TATAAT---7---fu
4
815
5
17
20
II
211
17
20
231
231
221
19
18
17
18
16
17
Putative -35 and -10 hexanucleotides are doubly underlined. Starting points of transcri;;ti;;n are singly underlined. (a), spacingbetween putative -35 and -10 hexanucleotides; (b). spacing between putative -10 hexanucleotide and the starting point oftranscription; (1-5), P21, P23, P32, P44. P59 [38]; (6-10). PI. P1O, P21. ss45, ss80 [30]; (11, 12), pWV05 prlP, prlM [39.40.97]; (13).nisin precursor [98]; (14. 14a). p9B4-6 ORFA1-lcnMa, ORFB1-lcnA ([42]. M. van Belkum personal communication); (15). usp45[43]; (16). lacR [99]; (17-21), Pal, Pf2. Pa3. Pg2, Pfl [41]. Promoters (17-21) are lactococcal phage promoters. The proposed -35and -10 hexanucleotides in entry (7) are situated upstream of the sequence shown [30]. GA TC, number of G. A, T and C residuesat each position totalled over entries (1-7). (9-12) and (14-21). Entries (8) and (13) were left out because of the inaccuracy in thedetermination of the transcription starting point. X- 10, y- II. Z = 13. F -14. Figures printed in bold face indicate that theresidue indicated occurred at least 10 times. or at least 5 times more than the corresponding complementary residue (i.e. G versusC. and A versus T). ., seemingly preferred nucleotides in L. laclis (as derived from this table); .., consensus E. colj and B.
sublilis 1743 promoter [6,7]; Pu, Purine residue.
insertion of possible terminator sequences, orother sequences, in front of the coding sequencemay interfere with translation initiation, ratherthan terminate transcription.
4.1.2. Transcription initiation signals of Lactococ-cus lactis-derived genes
Now that several lactococcal genes have beencloned and sequenced, their expression signals
15
20
20
14

can be compared with those obtained using thepromoter-probe strategy described in the preced-i:; paragraph. In Table 1 a compilation of lacto-coccal promoter seQuences is shown. Entries 1 to10 were isolated using promoter probe vectors[30,38], whereas entries 11 to 16 were isolatedtogether with specific L. lactis genes. Entries 17to 21 are lactococcal bacteriophage promotersthat were isolated using a promoter probe vectorr41]. Only promoters of which the functionality;as confirmed by mapping of the starting points
of transcription are included in the Table. Be-cause all known lactococcal promoters also func-tion in B. subtilis and E. coli, an obvious way toanalyse these seQuences is to look for the pres-ence of the weIl conserved seQuences constitutinga consensus promoter in these organisms [6,7].SeQuences resembling these so-called -35\TTGACA) and -10 (TATAAT) boxes can in-deed be recognized in the L. lactis promotersand are doubly underlined in Table 1. It is oftennot possible, however, to unambiguously identifyone particular hexanucleotide as the only possible-35 or -10 analog in the lactococcal seQuences.Therefore, the seQuences in Table 1 were alignedwith respect to the starting points of transcrip-tion, and the numbers of G, A, T, and C residuesat each position were totalled (Table I, bottorn).This procedure indicated a strong prevalence ofA residues at the starting point of transcription(underlined in Table I), and of T residues at the.-1 position. while A residues are presentthroughout the transcription initiation sites ~tud-ied, very few A residues were observed at the -Iposition and at the position immediately down-stream of the starting point of transcription. A-35 TTG seQuence can be recognized, as weIl asa region which is rich in A residues, and there-fore resembles a consensus -10 seQuence. Thedistance separating these -35 and -10 se-Quences seems to comprise a few nucleotidesmore than the 17 to 18 usua:lly found in B. subtilisand E. coli. It is important, however, to realizethat only a small set of promoters has been anal-ysed. The whole promoter area contains very fewC or G residues. Inspection ofTable 1 reveals thepresence of a TGN seQuence directly precedingthe -10 hexanucleotide in seQuences 1 to 6 and
8 to 10, but not in sequences II to 21. S.:v~ral ofthe former promoters had been selected f"r theiractivity in B. subtilis, however, in which this TGsequence is strongly conserved [7,38]. Th..,r.:lore,an inadvertent selection of promoters conlainingthis sequence may have occurred.
Promoters 14 and 14a contain exact I y th.., samesequence. Also the sequences upstream of thepromoters, which contain an open readin~ framethe function of which is yet unknown, ar.: iu.:nti-cal for at least 1 kbp (M. van Belkum, P...rsonalcommunication). The sequences downstr..'am ofthe promoters are identical up to the 7th ...odonof the coding regions [42]. In spite of this highdegree of similarity, a 2-bp differenc.: in thestarting points of transcription was obser\.:u [42].
It is generally believed that high levels of geneexpression result from promoters that clos.:ly re-sembIe the consensus sequence. This uo.:s notseem to be an absolute requirement, how..'v.:r, asis apparent from sequence nr. 15 in Tahl..' I. Noclear -35 sequence is present in this pr, Imoterbelonging to the gene usp45, which was s..'I.:.:tedbecause of the high level production (s.:..'r.:tion)of its gene product [43].
4.1.3. Translation initiation signals of La(.t()(.()CCI/Slactis-deriued genes
Table 2 gives a compilation of L. lactix-ll~rivedtranslation initiation sequences. In all th~1i~ se-quences, the translational start codon i1i I\UG,although it is known ~ha~ UUG and GU(i .ICt asstart codons in L. lactis in the expreSSi()11 ()f theB. pumilis cat-86 gene [38], and of the 11. xllhtilisnprE gene [44], respectively. As far as av.lilable,the 30 nucleotides preceding the tran1ilationalstart codon, and the 27 nucleotides followillg thiscodon are shown in Table 2. In the regillll pre-ceding the start codon, the putative SD se-quences are underlined. These sequel1~~1i arecomplementary to the 3'-end of L. lactix 16S-rRNA (3'-tJCUUUCCUCCA-5' [45]). l'Ilc fre~el1ergies of comp'ementa(ity of the putalivc SDsequences and the 16S-rRNA (~G), and Ihc 1iizesof the windows are also given in Tablc 2. Thelatter range from 4 to 15 bases, the most l.lIl11l11onvalue being 9. These sizes correspond III thoseusuaIly observed in E. coli and B. subtilix [ 12,46].

TabJe 2
Laclococcus laclis.derived Iranslation initiation sites
UAUGUUAAAAUAGCUUCU~UAUAGCCAUGAGUUUAAAUUUAAGAGAUCUUGAAUAU
8--"etSe'.LeuAsnLeuA'.gA.pLeuGluTy'.
AAUUAAAAAAGAU~AAAUUUCAUGACUUACGCAGAUCAAGUUUUUAAACAA
7--"etTh,.Ty'.AlaAspGlnValPheLysGln
UAUUACGGAGGAUUUAAAAUGCGCUUUAACCAUUUUUCAAUUGUUGAC
9--"etA'.gPheAsnH;sPheSe'.lleValAsp
CAAAUGCUUUUUUUGAAAGGACUUACACUUAUGACUAAAACACUUCCUAAAGAUUUUAUU
9--"etTh'.LysTh'.LeuP'.oLysAspPhelle
AAAUAGAUUUUUAGAACAG~UAGGUAAAUGAUAACUUUGCAACACCAAGAUUGGGAA
8--"etlleTh,.LeuGlnH;.GlnAspT,.pGlu
GCUACUUGUUUUUGAUAAGGUAAUUAUAUCAUGGCUAUUAAAAAUACUAAAGCUAGAAAU
9--"etAlalleLysAsnTh'.LysAlaA'.gAsn
UUGAUUGGAGUUUUUUAAAUGGUGAUUUCAGAAUCGAAAAAAAGAGUU
9--"etVallleSe'.GluSe'.LysLysA'.gVal
UUAUAAAAAU~AUUAUUAUGAAAAAUCAAUUAAAUUUCGAAAUCCUA
~---7--"etLysAsnGlnLeuASnPheGlulleLeu
UAUAAAAAUUGAAAGGAUUCAGGUACUAAAAUGAAAAAAGAUGAAGCAAAUACAUUUAAA----
---3--===6=="etLysLysAspGluAlaAsnTh'.PheLys
CCAGUACACUAA~CUUACAAAUUAAUGAAAAAUGAUAAUUUUUUAAUAAAUAGA
10--"etLysAsnAspAsnPheLeulleAsnA,.g
CGGGUUGCACCAUU6AGGAUUAGUUAAGAUAUGAAAAAAAAACAAAUAGAAUUUGAAAAC
-14=======11=="etLysLyslysGlnlleGluPheGluAsn
CUAAUAAAAAAGAACUGAGGUUUAGAGUUAAUGAAAAAAAAAGUUGAUACAGAAAAACAA
12--"etLysLysLysValAspTh'.GluLysGln
ACCGUGGGAACAAAUGUACACUAUCGGUUGGGGUCAUUAUGGA
-4--"etTy'.Th,.lleGlyT,.pGlyH;sTy'.Gly
AUGAAUAAAAAUGACAGCGAGGAUAUAUCAAUGAACUAUUUUAAAGGUAAACAAUUUCAA
-10====7=="etAsnTy'.PheLysGlyLysGlnPheGln
AAGGUUUCUCGCUCAGGUUUCUAUGAAUACAUGCAUCGUCGUCCUUCAAAACAACAAGUG
15--"etH;sA'.gA'.gP'.oSe'.LysGlnGlnVal
AUGAAACUUUUGGAAAGUGGAGGAUAUUGGAUGCAAAGGAAAAAGAAAGGGCUAUCGAUC
9--"etGlnA'.gLysLysLysGlyLeuSe'.lle
ACUGUAAGCAUUUCAGAGGAGACCGAAUCGAUGAAGAAAAAAAUGCGCCUUAAAGUAUUA
10--"etLysLysLys"etA,.gLeuLysValLeu
AAUGGGAGGAAAAAUUAAAAAAGAACAGUUAUGAAAAAAAAGAUUAUCUCAGCUAUUUUA
--5--"etLysLysLysllelleSe'.AlalleLeu
UACAAAAUAAAUUAUAAGGAGGCACUCAAAAUGAGUACAAAAGAUUUUAACUUGGAUUUG
10--"etSe'.Th'.LysAspPheAsnLeuAspLeu
GGUAAAAAAAUAUUCGGAGGAAUUUUGAAAUGGCAAUCGUUUCAGCAGAAAAAUUCGUA
11--"etAlalleValSe'.AlaGluLysPheVal
CGUGAUGUGUGAGGGAAAGGAGUCGCUUUUAUGGCCAAAAGUGGACUUUAUACAGGCGUA
9--"etAlaLysSe'.GlyLeuTy'.Th'.GlyVal
UAUAAGACAGAUAUAAAUGGAGAUAGAAUUAUGAUGAAUCACCCGCAUUCAAGUCAUAGG
9--"et"etAsnH;sP'.oH;sSe'.Se'.H;slle
-9.4(2)
-14.4(3)
-14.0(4)
-9.4(5)
-8.4(6)
-9.4(7)
-12.8(8)
-14.0
/-1.4-14.0
(9)
(10)
(II) -9.4
/-9.4-9.4(12)
(13)
-9.4
1-9.4-7.5
(14)
(15)
-14.(16)
-11.6(17)
(18) -4.6
(19) -17.8
(20) -14.4
(21) -16.2
(22)

13O
35O
41
456
789
10II12
131415
36
45
10
347
36
10
66
10
646
37
10
789
4II4
O
2
O
4
1
O
7
2
1
3
2
3
3
3
6
3
O
3
15
4
4
14
8
3
15
4
2
2
7
2
3
4
4
2
6
2
1
6
1
1
6
4
2
7
2
2
4O
O6O
46O
24O
27O
68O
63
619
4
1
6
2
O
6
O
18
19
20
21
22
23
4
2O
34O
2OO
3OO
515
4
O15
6
5
12
1
2
15
25
26
27
28
29
30
6
4
1
7
16
O3
O
17
IIO0
Nucleotide frequencies in Ihe 5'-parts of the 22 coding sequences presenled in Table 2. The AUG Iranslational start codon covers
positions + 1- + 3. Pos, position nr .; Obs, observed frequency; Min and Max, minimal and maximal frequencies possible without
changing Ihe amino acid sequence, respeclively.
Notes to Table 2.The functionality of the translation initiation sites has been inferred from expression studies (entries (1-13,16-19,22», lacZtranslational fusions (entries (I, 20, 21)), or homology to known DNA or protein sequences (entries (14, 15». L1G, free energy(kcal/mol) of complementarity between the SD sequence (doubly underlined) and the 3'-end of L. lactir 16S-rRNA (3'UCUUUCCUCCA-5' [45]) [100]. Figures preceding the deduced amino acid sequences indicate the size of the window between theSD sequence and the translational start codon. (I), mieR [lol]; (2), thyA [102]; (3), X-PDAP [103,104]; (4), pbg (lacG) [29,105,106];(5), pTR2030 hsp [107]; (6), pWV01 repA [108], nearly identical to pSH71 repA [29]; (7), pWV01 ORFC [108], and pSH71 ORFC[29]; (8-12), p9B4-6 ORFAI = IcnMa, ORFA2 = IcnMb, ORFA3 = lciM, ORFB2 = IciA, ORFC2 -lcjB ([42]; M. van Belkum
personal communication); (13), ~vML3 lysin [109]; (14), ISSI putative transposase [110] and IS946 putative transposase [III],nearly identical to: ISSIN and ISSIW putative transposases [112]; (15), IS904 putative transposase [113,114]; (16,17), pWV05 prtP,prtM [40,97]; (18), usp45 [43]; (19), nisin precursor [98,113,115]; (20,21), ORF32, ORF44 [38]; (22), citP [116]. Sequencesdownstream of the (putative) starting points of transcription are given for entries (2, 3, 7, 8, 13) and (20). Entries (9, 10, II) and (12)show sequences that are part of an operon. The translational stop codons of the upstream open reading frames are printed in boldface.

~].j.=".."
.,. ~" "
-O.0 "0" o
1- u
-0
.c
~
~
~ I~
8
.,.
-0,",0000000 r-N NO00 0...0...' N...N- -N 0...'0
00 0-0-00 ~N N- 00000..'. -..,...,.- -0 :!:0 '
0 00-0000..0 N- -0 0.. 0"'
-N...N O...ONOO ",.. 00 ON N... 0-
0"""00-0
OM-o 00000- oM '.'0 ."000000-'.'0 0'.'0 MO--0°
1-o."r 0-00'.'0 r-- -,., -0 -0- M~ -M...'.' M- MM~ 0...00...MM'.' ---,., :
r~""0-0~ ."".".0--0- 00 ~N ".-0 ".-00-- --0""- -0- ~~"" ".0~-0~-N".- N -"" ""-0,,,,...NN ~-N".N...N-."NN"" -N,iv
-"""N 0-0000 ~o NO00 000"' Nr--",- 0- -",N --0-00
°",-oN 0-0000 ",0 -000 ON N- -N=~ 00 N-"' 0"'0-00
--coo- 0...0
..:;
00
..,.
"~
0..t 8
c15
~- N...0...ON ~... :e-C -0 ...0- N.'..-N ~;:- -C0-00.'..0
ON~N NNO...No r-N No N~ r -0.'..- 0 ., C or-N NNN -N --
NN-C... ooON0 .,..r- 0-- 00 ",N -"'~... -~ ...r-~ ~-CON-----N- N ---
r-- O...N-C"'... ~o -0 -CN 0-", ...0-r-r- -C... ~ r C",...N-N- N N N-
N-C O-ONr-o ~... ~N -0 ...~ "'~ :?;- -~r- ~-C---C-
0 '.. NNOO ON N... -0 -~ N... O-N .,..O-C ~~O-C~...N -N --
"5
>-5 :i ~
=.:J
-'.'00-0
-=0'.'00
-00- -No --00-0
Mo -0 N-Oo --oNMM-
r--",~~ 000 r--r-- ~-o -0 ~r-- -~ !'I "' 0 or--", "'a-"'""" I
..""Nr- N-"'..'r-- -COO -C0- 00 r-N N" "0--Co ..", N..'O O-N"'-..'°1,,"'-C"' -c" 0-"' ..,-c -" N-","' -".., ..'NN-N-,
,
-NOON ..",,--r-- ON "'" No ..'-C",N 0-- NOC..' N"..'--C- j;~---"-
..'N"- °,,-00-CN -c"' ",N -0 ..,r- -"'-c- N~" N"'0"'-r-1
-oa-r--o ===N"".' ~~ ~,., =- =~ N~ ~r-'.'", Nr- NX'.' r-'.',.""r ---N ,.,
-~a-~ 0,.,=,.,"'= ~~ ~"' =- N-O N::) -~r-N ,.,- "'::,., ~~--O~...
~~
0< 0 ~u o<~u ~uo<~u o<~u 0 ~u o<~u <0<« ~ ~~ uuuu oouuuu uuuu 0 « ~~~~ 0«« < ~~ uuuu «~~~~ «« ~ ~~ 0000 ~~~
.--~ ~ .--0 o-~o -o"'OonNO NonNO "' N- -~ -00--
0..,. N 0--0000""""' --NO N No OMN- 00-
-"' --0-0000---000-0 -0 0-"'0 00-
0,., "' ,.,- 000000-"'000'.""- """ 00'.'0 -00
-= ...~,., 0-",0 "'00'.""- 0",-0 "',., 0"",,00-0
"' .,.00.,.0.,. "'.'.NN.'.O 0"'.'.0000 ON.,.O CC.,.
",-0 -0 N"' .'."""0 r-",oN O 0~00 ~ N~-oO 0-0N -.,. "'.,. .,.N .,.
.-N N N0- -~..."' O-N",-oNN ..'.0-.-N ..,. ..'.0- N-ON... N..'.r-0 -0- ...0--0- Nr ' Or--oO r- ...0- -o~N.- -
-..,. -N N N
0- 0 OOM ",...0... 'C 'C O-MO00 000 "0-0... 0 ",1" ~ 'C... M- MM ~ ...M --"'-
s ..,
.., "'- 0-00"'-00-00-0"'0 "' ..00-..'000-
"' -"' 0"'00..'0--"'- 0"'-- "'0000000-
~ 00-00000
." "~ f
00000 oo-",No oNoO 0 -0
e~
~"' ~
Co ~~ >-
1- 1-
"0;>
0...000
-0c
~
""~~~~ -'"Q " ~ ..
~"~]",MCo
~'"-
U~~~~
~~ 0N ~ ..~° O .~ .----'"0
~..'."5f.."' ~ c ..',,--0
-" -.-"'-
.0 II ~ -
~N .~
O<C.'a= ~ .
~o:g~" 0- ..
E .-;~" .--
..~:f.~
.." "
g-,,::J~-g II ~ >
u- > ..
.<~;;:;"0... -o~ .-
U~O .O ..M
..~ -
"0 C-
.U "i' .g OÓ
'"..-0'-'"-
O-U
cO'-5.-c. c ~
E " ~
'" .E "c u
.I E ..<.CO~< " c.
.;~"" .=
~~~~*
f..U~;;"'~...c.~~
-O:E';'~-O ,,-.-
O ~"'~
c. c
o~Q~,"-"' :sf....-c.c " '" .~
--> 7". :s
~~~~E" ~ ..ó e-c.N..-'" ..-u "0
~ < ""0 , --
c-~..'"
"--" "
u
~bIlO:,,"
"-Nt:"..-U"--" ~ C
.cOt..."'..
-..,o:oE.= ° O ~ 8
]8~c.~~N" .U
=0:-,,'?+
~!c;.I1.::'.O.--~
u -.,
c .U- .."" c= ..0: -::' f..0-- -.
~~0~~--.~ o CC "' ." c. .-OO",~-"O--C-8 i'f II ~ .~,,-N 0.c~Q~C-" "",0:,,"E
""~0"5'"u ~ ~
.-~ --"
"0~c.. ..c.
, 0 "
~"--"~..~ .II -~
~"0-0"'t;: ..~ ..c
---0
"M~°"O.cO~-
f..=:0=.8
0..'0 -.., , '0 "' ..'",-oN "'~ NO..' 00-,- , --
NN ~ MN MO...O N-O~...O M...o-- 0 M... M 00--N N- -
"'~ "' --'.'0-0 -N '.'N 0 0~ -0 -0- -0-00-- 00-.,. -,.,- ---N --N --
"1!:: 0- ~ 0 0 --0..' 0 N'C 'C ~.., -..'..."100-
..'N , N..'ON ~N"1"1"'- --N ° O-N 0.."C- 00-N 'C -
~- .-"'... ONON N- ",.0-"' .-.0 ..Na-N 00--N ---
-0"' --N ",f---M o-Mf---of '. Mf--f--- -0..'."' "'-"'0- 0-0-0-0 N MN -M- NN-NN- -0"",,-0 ..'.N N--oN
~~ ,0.., 0"'0- ..,-"' ..'..."OM ~.-- "'.--"'..'.00-
"'00 ,0 """ -"""0 """0.0-0 .000.-- -..,.~- 00-
""" 0"'-0 "' 0"'.0- 00-
,."" "' '.'0 "'~"'o .0"'--.0- ~0--- ,..,.. "'..."',.. 00--"' ---"'
""" .0"' 0-",- , ,"' 00-'.."'0 ,."., "'..."'- 0-0

.
codons covering these positions, j.e. the observed ';
nucleotide frequencies more or less equal the !mjnimal frequencies. Of the positions mentioned, ,
only at positjons + 21 and + 30 there is no strongneed for an A jn the coding sequence (the minj-mal frequency is 0), and yet this residue prevajls.Concerning the positions +4, +5, +7, +8, + 10,+ 11 and + 20, the questjon arises as to whethera certain nucleotide is preferred in the mRNA, orwhether the preference applies to a certain (setof) amino acid(s) in the gene product. Table 2shows that there is no clear preference of oneparticular amino acid in the deduced protejnsequence at either of the positions mentioned.Analysis of an extended number of translationjnitiation sequences is requjred to reveal whetherthe seemjng nucleotide preferences are real pref-erences. Although at some positions G, A, U andC residues are present in more or less equalnumbers (e.g. positions + 13, + 16, + 17), at manyother positions there seems to be a bias towardsA and U residues. This may ret1ect the generallyhigh (A + T) content of L. lactis (see also Section4.1.4 on codon usage).
Most 4G values range from -8 to -14 kcal/mol,with a mean value of -11 kcal/mol. In thisrespect the SD sequences of L. lactis resembiethose of E. coli, where 4G values ranging from-9 to -12 kcal/mol are common, with a meanof -11 to -12 kcal/mol [12]. In B. subtilis, onthe other hand, 4G values ranging from -14 to-19 kcal/mol are common, with a mean value of-16 to -17 kcal/mol [12]. In the area precedingthe SD sequence often long stretches of A or Uresidues are present.
The SD sequence, the start codon, and thewindow between the two, are generally consid-ered to be the main determinants of translationinitiation efficiency. Additional sequences, 10-cated outside of this region, may have an effecton the efficiency of translation initiation, how-ever. The nucleotide sequence of the mRNA perse may influence the interaction between themRNA and the ribosome [8,12,47,48]. AIterna-tively, secondary structures may be formed in themRNA that influence the efficiency of translationinitiation [47,49].
In order to examine whether certain nu-cleotides are preferred at certain positions withinthe coding sequence, the frequencies of occur-rence of the nucleotides present in the codingsequences from Table 2 were listed in Table 3.Together with the observed frequency of a givennucleotide at a given position, the minimal andmaximal frequencies of that nucleotide at thatposition are given. The maximal frequency wasobtained by introducing that nucleotide at thatposition in all coding sequences where it waspossible to do this without affecting the aminoacid sequence of the gene product, i.e. by usingsynonymous codons. Similarly, the minimal fre-quency was obtained by avoiding the use of thenucleotide of interest at a given position as muchas possible. When the AUG start codon is set tocover positions + 1 to + 3, a strong prevalence ofA residues can be seen at positions +4, +5, +7,+8, +10, +11, +20, +21 and +30. A similarhigh preference of A residues in this region wasobserved in B. subtilis [12]. From Table 3 it isclear, however, that the high incidence of Aresidues at the majority of these positions is di-:ectly linked to the amino acids encoded by the
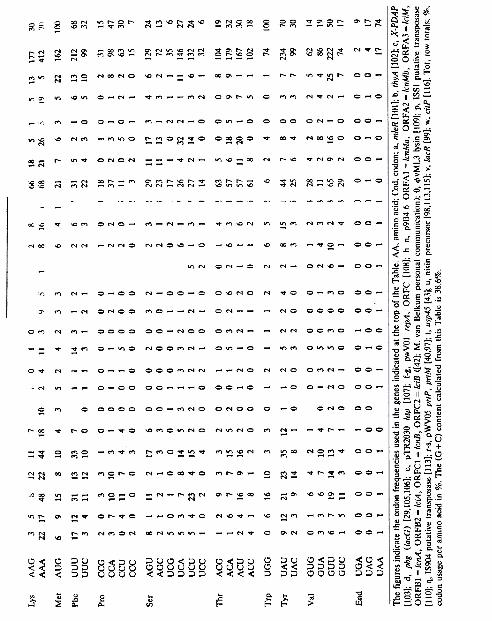
4.1.4. Codon usage in Lactococcus lactisLittle is known about codon usage in L. lactis.
Kok et al. [40] made an analysis of the codonusage in the L. lactis ssp. cremoris protease gene,and Porter et al. [50] analysed the L. lactis ssp.lactis pbg gene. Table 4 includes these data andthose of a number of other lactococcal genes. TheTable shows that the codon usage can varymarkedly between genes. Although in many casesthis variation may be a consequence of chancebecause of the small numbers of the variouscodons involved per gene, some differences arestriking, as, for example, the noticeable underre-presentation of the CAG codon for glutamine inusp45. Another example concerns isoleucinewhich is encoded predominantly by the AUUcodon in most genes. In addition to this codon, alarge number of AUA codons is present in thehsp gene, whereas in the prtP gene a large num-ber of AUC codons is found. Furthermore, in theprtP gene the AAG and AAA codons for lysineare equally represented, whereas the overall fig-ures (column: %) show a marked preference for

Table 5
G Jon usage in L. lactis, B. subtils and E. coli
LI Bs Ec AA cod LI 85 EcAA cod LI Bs Ec AA cod LI Bs Ec AA cod
Ala GCG 13 22 31 Gin CAG 24 48 76 Li.;- UUG 24 14 9 Ser AGU 24 11 6GCA 29 29 22 CAA 76 52 24 UUA 30 24 7 AGC 13 26 26GCU 40 27 26 Glu GAG 19 30 27 CUG 8 21 68 UCG 6 11 11GCC 18 22 21 GAA 81 70 73 CUA 1I 6 2 UCA 27 20 7
Arg AGG 5 III Gly GGG 1I 13 8 CUU 20 25 7 UCU 24 23 23AGA 24 22 I GGA 26 32 5 CUC 8 10 7 UCC 6 9 27CGG 4 14 3 GGU 43 24 48 Lys AAG 30 27 24 Thr ACG 19 24 18CGA 19 12 3 GGC 21 31 39 AAA 70 73 76 ACA 32 45 7CGU 36 22 56 His CAU 71 67 54 Phe UUU 68 57 37 ACU 30 15 25CGC 1I 19 36 CAC 29 33 46 UUC 32 43 63 ACC 18 16 50
Asn AAU 70 56 26 lIe AUA 18 13 3 Pro CCG 15 40 65 Tyr UAU 70 65 40AAC 30 44 74 AUU 58 50 36 CCA 47 21 16 UAC 30 35 60
Asp GAU 68 61 46 AUC 23 37 61 CCU 30 30 12 Val GUG 14 24 27GAC 32 39 54 End UGA 9 21 17 CCC 7 9 7 GUA 19 21 22
Cys UGU 74 68 43 UAG 17 8 8 MeI AUG GUU 50 31 36UGC 26 32 57 UAA 74 71 75 Trp UGG GUC 17 24 15
,he figures represenl Ih~ codon usage in L. lactis (LI) (Table 4). B. subtilis (Bs) [117] and E. coli (Ec) [117J. expressed inpercenlage per amino acid. AA. amino acid; cod. codon; ...100%.
For some amino acids L. lactis and E. coli showopposite codon preferences, while B. subtilis takesan intermediate position, as for instance withrespect to the asparagine and glutamine codons.For some amino acids, the codon choice of L.lactis resembles that of B. subtilis, whereas forothers it resembles that of E. coli. However,neither L. lactis nor B. subtilis show the clearpreference of E. coli for one or two codons inthose cases where 6 codons are available, i.e. forarginine and leucine.
In the preceding paragraphs an overview hasbeen given of some features that are, or are likelyto be, important in gene expression. It was shownthat also in L. lactis considerable variation existsin promoter sequences, translation initiation se-quences, and codon usage. Unfortunately, little isknown about the optimal characteristics of eachof these features, and their possible interactions,necessary to achieve high level gene expression.Whereas it is common belief that high level ex-pression is obtained using promoters and ribo-some binding sites with high similarity to theconsensus sequences, it is intriguing that theusp45 gene [43], selected for its high level expres-sion, lacks both.
!he AAA codon. In !he same gene, all four codonsfor threonine are equally represented, whereashe overall figures seem to favour !he ACA and
ACU codons. In usp45 these two codons are !heonly ones present for threonine. At this time wecan only guess about !he meaning of these differ-ences. A\so in other bacterial species, and higherorganisms, a considerable within-species diversityin codon usage has been observcd [51]. Especiallyin E. coli this diversity has been associated withgene expression, with a strong bias towards aparticular subset of codons in highly expressedgenes, and a more even codon usage in moder-ately expressed genes [51]. In contrast, a markedlyunbiased codon usage has been reported for B.subtilis [52].
In Table 5, !he overall codon usage in L. lactisas given in Table 4 is compared with !ha! of E.coli and B. subtilis. In general, L. lactis shows apreference for codons with an A or a U at !hewobble position, which is reflected in the high(A + T) content of !he organism (approx. 62%[53], Table 4). L. /actis, E. coli and B. subtilisresembie each other in !he preference for fiomecodons, e.g. GAA for glutamic acid, AM forlysine, and UAA as !he translational stop codon.

4.2. Heterologous gene expression
Relatively few studies have dealt with the ex-pression of heterologous genes in L. lactis. In thefollowing paragraphs an overview of heterologousgene expression in L. lactis is presented.
4.2.1. Antibiotic resistance markersThe first class of heterologous genes expressed
in L. lactis is composed of genes specifying an-tibiotic resistances. Plasmids from non-Iactococ-cal Gram-positive bacteria, or derivatives thereof,carrying these antibiotic resistance markers wereused in conjugation experiments, or as genecloning vectors [14,27,54]. A1ternatively, the genesspecifying antibiotic resistance were used to labelcryptic lactococcal plasmids in order to use theseas selectable gene cloning vectors [25,26]. Thefact that lactococcal expression signals closelyresembIe those generally present in Gram-posi-tives, al1owed the expression of these heterolo-gous genes to sufficient levels without the needfor further adjustments. This noticeable featuresubsequently permitted the rapid development ofthe genetic analysis of L. lactis. Arnong the an-tibiotic resistance-conferring genes most com-monly used are the chloramphenicol transferase(cat) gene of the S. aureus plasmid pC194 [55],and the MLS resistance genes of the S. aureusplasmid pE194 [56] and the Enterococcus faecalisplasmid pAMfJ1 [57]. In addition, the B. pumiliscat-86 gene [30,38], and the E. coli TEM-fJ-lactamase gene [33,34] have been used to isolateand study lactococcal expression and secretion
signals, respectively.
to be a useful tool in the study of translation[59,60]. Recently, Haandrikrnan et al. [61] usedthe lacZ gene in combination with the Cyamopsistetragonoloba (Guar) cDNA gene encoding a-galactosidase, to study divergent expression sig-nals. The a-gal gene was fused to the signalsequence of the B. subtilis a-amylase gene [62],and both galactosidases were provided with L.lactis-derived transcription and translation initia-tion signals. The B. stearothermophilus and B.licheniformis a-amylase genes, deprived of theirsignal sequences, have been used to isolate andstudy lactococcal secretion signals [33,36].
Other heterologous gene products producedby L. lactis have potential applications in dairyindustries, such as the bovine prochymosin (Sec-tion 6; [63,64]), the B. subtilis neutral protease(Section 6; [44]), and severallysozymes ([35]; Vande Guchte et al., unpublished results).
Lysozymes (EC 3.2.1.17), defined as 1,4-{3-N-acetylmuramidases cleaving the glycosidic bondbetween the Cl of N-acetylmuramic acid and theC4 of N-acetylglucosamine in the bacterial pepti-doglycan [67,68], may find an application becauseof their antimicrobial activity against several bac-teria involved in food spoilage and food-bornedisease [69- 71]. L. lactis was transformed to pro-duce these enzymes using a lactococcal expres-sion vector that contained gene expression signalsand a short 3'-truncated open reading frame oflactococcal origin [35]. Copy DNA encoding themature hen egg white Iysozyme was fused in-frameto the lactococcal open reading frame, whichresulted in the production of a hen egg whiteIysozyme fusion protein in L. lactis [35]. Also, theunfused mature hen egg white Iysozyme codingcDNA was expressed in L. lactis, albeit withreduced efficiency [65]. The differences in theexpression levels of the fused and unfused geneare probably (partly) due to differences in theefficiency of translation initiation [59,60].
An enzyme closely resembling the hen eggwhite Iysozyme, is encoded by the E. coli bacte-riophage T4 e gene. In order to express this genein L. lactis, it was provided with L. lactis-specificexpression signals that were positioned immedi-ately upstream of the coding sequence [65]. Alter-natively, the gene was expressed by means of
4.2.2. Other heterologous genesHeterologous genes other than antibiotic resis-
tance markers have been expressed in L. lactis bytranscriptional or translational fusion to L.lactis-derived expression signals. Among these isthe E. coli lacZ gene, encoding {3-galactosidase.De Vos and Simons [58] fused this gene to theninth codon of the L. lactis ssp. cremoris SKllprtP gene, and showed that the hybrid gene couldprovide the lactose-deficient L. lactis ssp. lactisstrain MG1363 with the ability to grow on lac-tose. The lacZ gene has, also in L. lactis, proven

trmslationaJ coupIing to a short 3'-truncated L.{,c/is derived open reading frame [65].
The E. co/i bacteriophage À Iysozyme, the ÀRgene product, was produced in L. /ac/is af ter thelysozyme coding sequence and translation initia-tion signals were transcriptionally fused to a 3'-truncated lactococcal open reading frame [66]. Inthis configuration, efficient expression of the ÀR,'ene was shown to be dependent on the transla-;on of the preceding lactococcal open reading
frame.
5. IMPROVEMENT OF GENE EXPRESSIONIN LACTOCOCCUS LACTIS
gene dosage. One reason for this is, that in food-grade applications it may be preferabie to insert aheterologous gene at a specific site in the chro-mosome of the host. In a fixed copy numbersituation, gene expression may be enhanced byimprovement of the efficiency of transcriptioninitiation. Since, as described in paragraph 4.1.2.,it is as yet not clear what exact I y constitutes anoptimal transcription initiation region, the mostobvious strategy to enhance transcription is toreplace the promoter present upstream of a geneof interest with another, more efficient, pro-moter. This strategy was applied by Van derVossen ([39]; unpublished results), who isolated anumber of promoter sequences from the chromo-some of L. lactis ssp. cremoris Wg2, and usedthese to improve the transcription of the L. lactisssp. cremoris prtP and prtM genes in L. lactisssp. lactis, resulting in elevated mRNA levels. Aconcomitant increase in protease activity was ob-served.
A second important determinant of gene ex-pression is the efficiency of translation initiation.The characteristics of a translation initiation re-gion that are considered to be of prime impor-tance in determining translation initiation effi-ciency are: the extent of complementarity be-tween the SD sequence and the 3'-end of 16S-rRNA, the start codon, and the window betweenthese two sequences [12,46]. Furthermore, natu-ral ribosome binding sites tend to have a lowpotential for secondary structure formation[49,74], which is in part a consequence of thegenerally low (G + C) content of these sites. Thenucleotide sequence in the region surroundingthe start codon might also be important in estab-lishing additional contacts between the ribosomeand the mRNA [8,12,47,75]. Changes introducedin a ribosome binding site to increase the comple-mentarity to the 16S-rRNA, and to convert thewindow to a size that is frequent I y observed, canin some cases increase gene expression [59]. How-ever, this method, or that of precisely fusing acoding sequence to an existing ribosome bindingsite, may not be generally preferabie, because theexchange of coding sequences and the introduc-tion of changes in ribosome binding sites caneasily affect mRNA secondary structure forma-
In the preceding paragraphs, it was shown that"everal heterologous genes could be expressed inL. lactis employing L. lactis-specific expressionsignals. Depending on the nature of the heterolo-gous gene product and its application, however,improved gene expression may be desired. Simi-larly, enhancement of the expression of homolo-gous genes may be advantageous for certain ap-11ications. Several strategies can be envisaged toaccomplish enhanced gene expression. One wayis to increase the gene dosage, for example byincorporating the gene in a high copy numberplasmid, or by multiple insertions of the gene intothe chromosome. Several high copy number plas-mids for use in L. lactis have been described, for
example pCK2I [72], pIL253 [27], pNZ12 [25],and derivatives thereof. De Vos et al. [63] re-ported an increased production of the L. lactisssp. cremoris SKll protease af ter cloning of thegenes involved in its production, prtP and prtM ,on a plasmid with a copy number higher than thatof the plasmid that originally carried these genes.Recently, Leenhouts et al. [73] realized the inte-gration of the L. lactis ssp. cremoris Wg2 prtPand prtM genes into the chromosome of L. lactisin a Campbell-like way. The protease activitydisplayed by the transformants was shown to bedependent on the number of copies of the genespresent, although no linear relationship was ob-
served. IIn the following we will focus on methods of
increasing gene expression without affecting the

tion in the translation initiation region, and thusaffect gene expression [60]. In fact, results ofmany changes introduced in ribosome bindingsites with the aim to improve the interactionbetween the ribosome and the mRNA, may to aconsiderable extent reflect changes in mRNAsecondary structure [46]. Therefore, a configura-tion may be preferred in which translation of agene of interest is coupled to that of a precedingefficiently expressed open reading frame [59].
The principle of translational coupling was firstrecognized by Oppenheim and Yanofsky [76], andhas since been described for many naturally oc-curring and artificial systems [77-83]. It ensuresthe efficient translation of a gene by making itdependent on the translation of a gene immedi-ately preceding it. Translation of the upstreamgene may serve to resolve secondary structuresthat would otherwise occlude the ribosome bind-ing site of the downstream gene, thereby render-ing this ribosome binding site accessible to freeribosomes [76]. Alternatively, ribosomes that havetranslated the first gene may proceed to translatethe second gene af ter a translational restart [76].Especially in the latter situation, translation ofthe downstream gene would profit from an effi-cient initiation of translation of the upstreamgene. Therefore, if an efficient translation al cou-pling could be realized, it would be possible tomake use of efficient expression signals of anupstream gene, without disturbing the delicateinterplay that may exist between transcriptionand translation signals and the (5'-end of the)coding sequence. In reference [59] a model sys-tem is described in which the juxtaposition of thestop and start codons of two tandemly arrangedopen reading frames, the lactococcal ORF32 andthe E. coli lacZ, was varied, in order to optimizethe translational coupling betWeen the twoopenreading frames in L. lactis. This study showed theresult of translational coupling, which in this casemost likely occurred via a translational restartmechanism, to be critically dependent on theconfiguration in which the two open readingframes were juxtaposed. Only configurations inwhich the stop codon of the upstream ORF32and the start codon of the downstream lacZ wereI;ontiguous or partially overlapping, resulted in
translational coupling. The best result was ob-tained with an AUGA configuration, in whichUGA represents t:i1eStop codon of the upstreamORF32, and AUG the start codon of lacZ. It wasshown that tï=ansrational coupling could be usedto enhance the expression of lacZ as comparedto the expression of this gene in the absence ofORF32.
Other processes that may affect gene expres-sion, but are difficult to optimize, if at all, are therates of transcription and translation. The lattervariabie may be correlated to codon usage, al-though opinions are divided on this issue[8,51,84-86]. In E. coli, a strong correlation isobserved between high levels of gene expression,the use of a particular subset of codons, and thehigh abundance of specific tRNAs [51,85], sug-gesting a causal connection between these fac-tors. Bonekamp et al. [84], however, have pre-sented evidence suggesting that translation ratesof individual codons do not reflect the concentra-tions of the corresponding tRNAs or the frequen-cies with which the codons are used in E. coli.Furthermore, several authors state that initiation,and not elongation, is usually the rate-controllingstep in translation [8,87]. For L. lactis, there isinsufficient inforrnation to answer the questionwhether a particular codon usage might raisetranslation efficiency (paragraph 4.1.4.).
No inforrnation is available on the possibilityto increase the abundance of specific mRNAs byimproving the rate of transcription. As in transla-tion, elongation in transcription usually seems notto be rate-Iimiting [8]. mRNA stability, however,may be improved by the addition of a 3'-exten-sion that is capable of secondary structure forrna-tion, such as a rho-independent transcription ter-minator, thus protecting the transcript fromdegradation by exonuclease activity [88,89].
In conclusion, strategies to improve gene ex-pression heavily depend on improvement of initi-ation of transcription and translation. Althoughtranscription initiation signals, translation initia-tion signals and coding sequences are oftenthought of as distinct box es that can easily beexchanged to improve gene expression, there arestrong indications that this is an oversimplifica-tion of reality. Therefore, it may be preferabIe to

use an undivided set of expression signals, i.e. ther omoter, the ribosome binding site, and (the5'-part of) the coding sequence of a highly ex-pressed gene, to express a heterologous gene,provided that translation of the latter can beefficiently coupled to translation of the former.The feasibility of this approach has been demon-strated in references [59] and [65], describing theexpression of the E. coli lacZ, and the E. coli
Jcteriophage T4 e gene in L. lactis, respectively.
6. SECRETION OF HETEROLOGOUS PRO-TEINS
Various approaches have been used in order) make L. lactis secrete hetero!ogous gene prod-
Jcts. Van Asseldonk et al. [43] screened severalL. lactis strains for secreted proteins by SDS-polyacrylamide gel electrophoresis of culture su-pernatants, and showed !ha! alf strains producedextracelfular proteins, one of which, banding at50-60 kDa, was particularly abundant. The chro-mosomalfy located gene usp45, encoding this pro-cin in L. lactis ssp. lactis MG1363, was iso!ated.
DNA and protein sequence analyses revealed !ha!thc gene most probably contained a signa! se-quence of 27 codons. This signal sequence wasused to direct !he secretion of !he B. stearother-mophilus a-amylase [36]. Similarly, !he signa! se-quence of !he L. lactis ssp. cremoris SKll celfcnvelope-associated protease gene, which mostlikely has a length of 33 codons, could be used toeffect secretion of !he bovine prochymosin [63], aprotein of major importance in !he dairy industry.Bath signal peptides conform to !he structura!characteristics of sigrial peptides sensitive to sig-nal peptidase I cleavage, as described by VonHeijne and Abrahmsen [90].
In !he strategy described above, secreted ho-mologous gene products were identified, afterwhich !he corresponding genes were cloned andana!ysed. The putative signa! sequences weresubsequently used to direct !he secretion of het- .
erologous gene products. Another approach wasused by Pérez-Martinez et al. [33] and Sibakov etal. [34]. In both studies !he 5'-truncated E. coliTEM-{3-!actamase gene was used as a reporter
gene in a signa[ sequence probe vector. In addi-tion, Pérez-Martinez et al. [33] used the 5'-trun-cated B. licheniformis a-amylase gene as the re-porter gene. Fragments of the L. lactis ssp. lactischromosome were cloned upstream of thesegenes, after which gene expression and secretionof the gene products was studied. In both studiessecretion of the heterologous gene products wasobserved with signal sequences resembling thosedescribed by Von Heijne and Abrahmsen [90].Furthermore, Pérez-Martinez reported species-specific differences in the amounts of proteinsecreted. The chromosomal DNA inserts had notonly been selected for the presence of a signalsequence, however, but also for the presence ofgene expression signals. Therefore, it is not clearwhether the differences observed in protein pro-duction in E. coli, B. subtilis, and L. lactis weredue to species-specific requirements with respectto the signal sequences, or with respect to thegene expression signals. An effect of species-specific signal sequence requirements on secre-tion efficiency is quite conceivable, as pointed outby Von Heijne and Abrahmsen [90], who showedthat signal sequences from Gram-positive bacte-ria tend to be longer than those used in E. coli.
The functionality of a heterologous signal se-quence in L. lactis has been demonstrated by thesecretion of the B. subtilis neutral protease usingthe nprE gene specific signal sequence [44]. Thesecretion of this enzyme by L. lactis may providea solution to the problems involved in the appli-cation of the purified enzyme to accelerate theprocess of cheese ripening, which are the oftenuneven distribution and low levelof retention inthe cheese curd [91-95]. The B. subtilus nprEgene, encoding the neutral protease, was pro-vided with L. lactis derived transcription signals,and introduced into L. lactis [44]. The originalNpr signal peptide, which most probably contains27 amino acids [96], served to direct the secretionof the enzyme into the culture medium. Like theabove-mentioned signal sequences derived fromL. lactis, the nprE signal sequence resembles thecanonical signal sequences described by Von Hei-jne and Abrahmsen [90].
The signal sequence of the B. subtilis a-amylasegene has been used successfully to secrete the

.[ZI[-611[ .S-S- "l°!qoJ"!W .UOJ!AU3 "lddV ."!P"W
P"Z!I!q"Js ,("""!10WSO U! "U!",(18 ,!I!A\ UA\OJ8 SiJOUl"J"
.dss S!1"vl sn",,°:10I"v7 JO .uO!J"JodoJI""I" ,(q .UO!I"W
.JOJSU"JI ,(;)u"nb"JJ.'!8!H (6861) .d"\ .s"N pu" .H .010H [OZ)
"J.a .U018U!,!s"t\\ .A8010!qOJ"!W JOJ f.I"!,,oS U",,!J"WV
.[[Z-6ZZ .dd .(.sp3 .\II .}\ .ss!JJn:J pU1! .f"f .!II"JJ"d)
S"!J"u"D 1"":>0:>0Jd"JIS .ui .uO!J"JodoJJ""I" ,(q S!1"ol
sn":10:10ld.JIS JO UO!I"WJOJSU"J.L (L861) ")J.S .J"PU"IJ"H [61)
.L8S-Z-[8SZ .[S- "l°!q
-OJ"!W .UOJ!AU3 "lddV .SJS"ldoJOJd S!1"vl sn":10:10ldaJIS
jO UO!J"WJOJSU"JI pu" UO!I""JSU"JI Uo S"W,(zU" "!JAI
.0"10Jd JO 1""JJ3 (L861) )\.f .0pUO)\ pu" .V.S .A\°'1S°t\\ [81)
.ZOII-OOII.OS
"l°!qoJ"!W .UOJ!AU3 "lddV .Sls"ld010Jd S!1"vl sn":>0:>01
.d.JIS JO UO!J"WJOJSU"JI P!WS"ld Uo SUo! +,":J JO 1""JJ3
(S-861) .S ."I"A!S pu" .W..V .OJS!W!".L ..V .1'!8!Jt\\ uoA [LI]
.6Z[-[Z[ .6t .11"'l "l°!qoJ"!W SW3d .S!lilqns snl/i"v8
puc sil"vl sn"jo"old.JIS JO UO!SnJ IS"ld010Jd "!J"U"8J"lu!
f.q J"jSU"JI P!WSCld pu" SiljVl snj:10:10ld"JIS Jo UO!ICW
-lOJSU"JI p""u",!u"-"Wosod!l (8861) .D ."w"u"A pu"
.a ."!1"1 J"p U"/\ ."f .'10)\ ..W.!1.W"f .UOSSO/\ J"p U"/\ [91)
.S6[-t6[
,S- "l°!qoJ"!W .UOJ!AU3 "lddV .VNa P!WS"ld ,(q slS"ld
.010Jd SiljVl snj:>0:10ld.JIS jO UO!I"WJOjSU"JI ,(;)U,,!j!JJ"
.,!8!H (9861) .:J.W .U!do,!:J pu" .V .Jln"no}\ ..a .UOW!S [SI)
.6SZ-ZSZ .8t
"l°!qoJ"!W .UOJ!AU3 "lddV .8U!UOI" J"ln""IOW U! "sn pu"
UO!I"Z!W!ldo .SIS"ldoJOJd SiljVl sn,,:>0:10ld.JIS JO UO!J"W
.JOJSU"JI P!WS"ld (t861) 11 .,(")\"W pu" .)\"f .0pUO)\ [tl]
SIZI-[lli .[t "l°!qoJ"!W .UOJ!AU3
"lddV .VNa P!WSCld ,(q SIS"ldoJOJd S!1"vl snj:>0jold"JIS
jO UO!I"WJOJSU"J.L (ZS61) 11 .,(")\"W pu" .)\"f .0pUO)\ [[I)
1d .OPU"IJO .sS"Jd "!W"P""V
.1[-1 .dd ."\1 10/\ ï.P3 ..va .n"uqna) !"!""!I "'!1 jO
A8010!!I Jcln",,10W "'!.L uI .S!lilq"S sn//!jvf1 U! ,(J!"!J!""ds
l"UO!1"ISU"J.L (S-S6[) .:J"f .ZI!A\OU!q"}\ pu" t\\.d .J"8"H [Zl)
.St-1 .Lt .A"}\ "l°!qoJ"
.!W .S"""UC8JO pu" .S"JO,(j""n" .S"JO,(j,,:>0Jd U! S!S"'!IU,(S
U!"10Jd JO UO!I"!I!U! JO UOS!J"dw°:J ([861) .W .'1"ZO)\ [11)
.9t[I-Zt[1 ïL VSf1 .!"S .p""V "l1"N .:>0Jd .s"J!S
8u!pu!q "WOSoq!J pu" sl"ld!JI "SUOSUOU 01 ,(J!J"IU"W
."Idwo" .VN}\ I"WOSoq!J S91 ilOj V!'1"!J.'ljS3 JO ""u"nb
."s l"U!WJ"I.,[ "'!.L (tL61) 1 .OUJ"81"a pu" "f ."U!lfS [01)
.S-6[-Z6[
.691 ""U"!"S .UO!I"" U! s"u"8 1"!J"J""q JO UO!J"Z!I"nS!A
(OL61) .V.:J .S"WO'!.L pu" .V.!I .ol"'lw"H .1.0 .J""!W [6)
.L901-[901 .t "l°!qoJ"!W
[ "l°W .SUO!IS"nb A\"U pu" PIO "!IOj vi'lj.u.'ljS3 U! UO!I
I ."!I!U! UO!I"ISU"J.L (0661) .W .snj,("Ja pUT! .N .s"nb""f [8)i
r .9t[-6[[ .9S1 "I"u"D .u"D
I"lOW .S!lilqns snl/!jof1 U! UO!I"ISU"JI pu" UO!Jd!J"SU"JI JO UO!I"!I!U! ",!II"u8!s 1"'!1 s""u"nb"s "P!IO"ljnN (Z861)
.}\ .'1"!SOl pu" "f .0J"d .1.V .U!",!SU"UOS "W .su",!d"JS
..D .""'l "f"d.S .""!JD"l "N .8u"1 "d.:J .U"JOW [L]
ï9[Z
-[t[Z .SI .s,,}\ Sp!"V "!"I"nN .s""u"nb"s J"IOWOJd ilOj
.a JO S!s,(l"uV (L861) .d.}\ .SPIOU,("}\ pu" .!1.:J .,("IJ"H [9]
.PLZ-~9Z '6E .qs8unlj"sJod .SIJ!Mlj"I!W J'I'!)!.u'ZU'J!) pun U'I!'"lj"!18QW .ui'll!wsu,q" U! U,w
-s!u.8JOOJ"!W 'IJ'pU~J'A lj"S!I'U'!) (L86!) .W 'J'qn,.L [~].88!-~L!
'L8 .A'~ 'l°!qOJ,,!W SW3d ..!J,I".q P!". "!1".1 WOJjSl!j'u,q I.UO!I!JlnU pu. ljll.'H (066!) .3.S 'pu.I!I1!!) [p]
.8P!-IE! 'L8 .A'~ 'l°!qOJ,,!W SW3d .JOA.IJ 'S"lj"UO .!J'I".q P!". ,,!p.I jO p.dw! 'lj.L (066!) .d.N 'uosiO [EJ
.P9!-6PI 'L8 .A'~ 'l°!qOJ,,!W SW3d .SUO!I.1-U'WJ'j P"j pu. pOOj U! .!J,p.q P!". ,,!p.I jO S'!I!A!p."!IS!uo8.IUV (066!) .f'i\\ 'zso8oJqoa pu. .3.S 'u'J8pu!, [Z]
.PI-E 'L8 .A'~ 'l°!qOJ,,!W SW3d ..!J,I".q P!". "!1-".1 U! SIU'W'AOJdW! 'Jnlnj pu. lu,s,Jd :f.8°loulj"'10!qJOj SUO!I."!lddV (066!) .V.)! 'U!MPI.8 pu. .,., 'A.)!"W [I]
S3JN3~3d3~
.:Ju:J8 sn08
-010J:J1:Jlj :Jlj1 1° Uo!ss:JJdx:J 1U:J!:)!n:J ljS!ldwo:):)\J01 pOlj1:JW :Jlq\J:)!ldd\J AII\JsJ:J/I!un 1S0W :Jlj1 :Jq A\Jw
:Ju:J8 sn08010WOlj (p:J1\J:)UnJ1-,() p:Jss:JJdx:J Al1u:J!:)
-!11:J u\J 1° 1\Jlj1 01 :Ju:J8 sn08010J:J1:Jlj \J 10 UO!1
-\JISU\JJ1 :Jlj1 8u!ldno:) 1U\J1JOdW! :Jq OSI\J A\Jw :JS
J;)d Uo!8:JJ 8U!po:) (:Jlj110 1J\Jd-,~) :Jlj11° :J:)u:Jnb:Js
:Jp!10:JI:)nu :Jlj1 pU\J :JJm:)nJ1S f.j\JpUO:):JS VN~w
'(:J1!S 8u!pu!q :JWOSOq!J pU\J J:J10WOJd) sl\Ju8!sUo!ss:JJdx:J p:J/IJ:JSUO:) AII\JJ:Ju:J8 :Jlj1 01 UO!1!PP\JU! ':J:)U!S .sl\Ju8!s Uo!ss:JJdx:J :Ju:J8 :)!1!:):Jds-s!I;JUl 7
10 :Jsn S:JJj\JW Wn!J:J1:)\Jq S!lj1 U! Uo!ss:JJdx:J :Ju:J8
sn08010J:J1:Jlj 01 lj:)\JOJdd\J PJ\J/l\JO11lj8!\JJ1S 1S0W
:Jlj1 'S!I;JUl .7 U! A:lu:J!:)!11:J J!:Jlj1 :JU!WJ:J1:JP SI\Ju8!sUo!ss:JJdx:J :Ju:J8 :Jlj1 10 S:JJm\J:J1 lj:)!ljN\ :JJn:)sqo
SU!\JW:Jl 1! SV .SWS!U\J8JO :J:JJlj1 :JS:Jlj1 U! 1u:J!:)!11:JAII\Jnb:J 10U :JJ\J S!1;JUl .7 pU\J s!I!lqns .H '!I°;J
.:I WOJ1 SI\Ju8!s Uo!ss:JJdx:J :Jlj1 'S:J!1!J\JI!W!S :Jlj11°
:J1!ds U! 1\Jlj1 'J:J/I:JN\Olj 'p:J1\J:)!PU! :J/I\Jlj S:J!pmS I\JJ:J-/I:JS .s:J!:):Jds 1\J!J:J1:)\Jq J:J11\JI :Jlj1 U! s:Ju:J8 p:J/I!J:Jp
-S!I;JUl .7 1° J:Jqwnu \J 1° Uo!ss:JJdx:J :Jlj1 P:JN\OII\J
:JJm\J:J1 S!lj.L .S!I!lqns snll!;JuH pu\J !1°;J U!l{;J!.1;Jl{;J-s:I U! p:J/IJ:JSqo :JSOlj1 S:JlqW:JS;)J lj:)nw f.j:J/I
:Jlq\JI!\J/I\J N\OU \J1\Jp :Jlj1 WOJ1 8u!8J:Jw:J S!I;JUl .7 U!
SI\Ju8!s Uo!ss:JJdx:J :Ju:J8 10 :JJm:)!d I\JJ:Ju:J8 :Jlj.L
S)!~VW3~ DNIqn'JNOJ .L
.S!1:J°1.7 U! s;J:Ju;Jnb;Js Il!u8!s sno8o(oJ;)};Jq Jo ;Jsn ;)q}
Jo ;Jldwl!x;J J;Jq}OUI! 8U!P!AOJd '([19] :Z"rv UO!}:J;JS>
;)SI!P!SO}JI!I1!8-v ( Jl!n[) > oqo1ouO30.11;J1 S!sdoU.Io,{.J

[37] Leenhouls, K.J., Kok, J. and Venema. G. (1989) Camp-
bell-Iike inlegralion of helerologous plasmid DNA into
the chromosome of Lactococcus /actis ssp. /actis. Appl.
Environ. Microbiol. 55, 394-400.
[38] Van det Vossen, J.M.B.M.. Van der Lelie. D. and
Venema. G. (1987) Isolation and characterization of
Streptococcus cremoris Wg2-specific promoters. Appl.
Environ. Microbiol. 53, 2452-2457.
[39] Van der Vossen, J.M.B.M. (I988) Development and use
of hosl-veclor systems for the characterization of lacto-
coccal expression signals. PhD thesis. University of
Groningen. Groningen.[40J Kok, J., Leenhouts, K.J., Haandrikman A.J., Ledeboer
A.M. and Venema, G. (I988) Nucleo)ide sequence of
the cell wall proteinase gene of Streptococcus cremoris
Wg2. Appl. Environ. Microbiol. 54, 231-238.
[41J Lakshmidevi, G., Davidson. B.E. and Hillier, A.J. (1990)
Molecular characterizalion of Ihe Lactococcus /actis ssp.
cremoris temperate bacleriophage BK5- T and identifi-
cation of a phage gene implicated in the regulation of
promoter activity. Appl. Environ. Microbiol. 56. 934-
942.
[42] Van Belkum. MJ.. Hayema, BJ., Jeeninga. R.E.. Kok,
J. and Venema, G. (1991) Organization and nucleolide
sequence of two laclococcal bacleriocin operons. Appl.
En\liron Microbiol. 57, 492-498.
[43] Van Asseldonk. M., Rutten, G., Oleman. M., Siezen.
RJ., De Vos. W.M. and Simons, G. (1990) Cloning of
usp45, a gene encoding a secreted protein from Lacto-
coccus /actis ssp. /octis MG1363. Gene 95.155-160.
[44J Van de Guchte, M.. Kodde. J., Van der Vossen,
J.M.B.M., Kok, J. and Venema, G. (1990) Heterologous
gene expression in Loctococcus /actis ssp. /actis. synthe-
sis. secrelion. and processing of the Baci/1us sub(i/is
neutral protease. Appl. Environ. Microbiol. 56, 2606-
2611
[45] Ludwig, W., Seewaldl, E., Klipper-Balz. R., Schleifer.K.H., Magrum, L., Woese, C.R., Fox. G.E. and Stacke.
brandl, E. (1985) The phylogenetic position of Strepto-
coccus and Enterococcus. J. Gen. Microbiol. 131. 543-
551.
[46J Gold. L. (I988) Pos\lranscriplional regulatory mecha.
nisms in Escherichio co/i. Ann. Rev. Biochem. 57, 199-
233.
[47J Dreyfus, M. (I988) Whal constitules the signal for the
iniliation of protein synthesis on Escherichia coli
mRNAs? J. Mol. Biol. 204, 79-94.
[48] Tomich, C.S.C., Olson. E.R., Olsen, M.K., Kayles, P.S.,Rockenbach, S.K. and Hatzenbuhler, M.T. (I989) Effecl
of nucleotide sequences directly downstream from Ihe
AUG on Ihe expression of bovine somalolropin in E.
co/i. Nucleic Acids Res. 17, 3179-3197.
[49] De Smit, M. and Van Duin, I. (I990) Control of
prokaryolic Iranslational initiation by mRNA secondary
slructure. 10" Progress in Nucleic Acid Research and
Molecular Biology (Cohn, W.E. and Moldave, K., Eds.),
Vol. 38, pp. 1-35. Academic Press, San Diego, CA.
[21J PoweJ, I.B.. Achen, M.G.. Hil1ier, A.J. and Davidson,
BE. (1988) A simple and rapid method for genetjc
transformatjon of lactjc streptococci by electroporation.
Appl. Environ. Mjcrobiol. 54, 655-660.
[22J Van der Lelie, D.. Van der Vossen, J.M.B.M. and
Venerna, G. (1988) Effect of plasmid incompatibiljty on
DNA transfer to Slreplococcus cremoris. Appl. Environ.
Microbjol. 54, 865-87J.
[23J Gasson, M.J. (1983) Plasmid complemenls of Slreplo-
coccus laclis NCDO 712 and other lactic Slreplococcj
after protoplast-induced curing. J. Bacteriol. 154, 1-9.
4J Chopin, A.. Chopin, M.C.. Moil1o-Batt, A. and Lan-
gel1a, P. (1984) Two plasmid-determined restrictjon SYS-
tems jn Slreplococcus laclis. Plasmid 11,260-263.
[25J De Vos, W.M. (1986) Gene cloning jn lactic strepto-
coccj. Neth. Mjlk Dairy J. 40, 141-154.
[26] Kok, J.. Van der Vossen, J.M.B.M. and Venerna, G.
(1984) Construction of plasmjd cloning vectors for lactic
streptococci whjch also repljcate in Bacil/us sublilus and
Escherichia coli. Appl. Environ. Microbiol. 48,726-731.
27] Sirnon, D and Chopin, A. (1988) Conslruction of a
vector plasmjd family and jts use for molecular cloning
jn Slreplococcus laclis. Bjochimie 70, 559-566.
[28] Achen, M.G.. Davidson, B.E. and Hil1ier, A.J. (1986)
Conslruction of plasmid vectors for the detection of
streptococcal promoIers. Gene 45, 45-49.
[29J De Vos, W.M. (1987) Gene cloning and expression in
lactic slreptococci. FEMS Microbjol. Rev. 46, 281-295.
[30J Koivula, T.. Sibakov, M. and Palva, I. (1991) Isolation
and characlerjzaljon of LaclococcuS laclis ssp. laclis
promoters. Appl. Envjron. Microbjol. 57, 333-340.
[31] Sjmons, G.. Buys, H.. Hogers, R.. Koenhen, E. and De
Vos, W.M. (1990) Construction of a promoter-probe
vector for lactic acjd bacterja using the lacG gene of
LaclococcUS lactis. Dev. Ind. Microbiol. 31,31-40.
[32] Van der Vossen, J.M.B.M.. Kok, J. and Venerna, G
(1985) Constructjon of cloning-, promoter screening-
and terminator screenjng shutlle vectors for Bacil/us
subtilis and Slreptococcus laclis. Appl. Environ. Micro-
bjol. 50, 540-542.
[33J Pérez Martinez, G., Smilh, H.. Bron, S.. Kok, J. and
Venerna, G. (1990) Protein export functions isolated
frorn Lactococcus lactis DNA. FEMS Microbiol. Rev.
87, P23.
[34J Sibakov, M.. Koivula, T.. Von Wright, A. and Palva, I.
(1991) Secretjon of TEM-13-laclamase wjth sjgnal se-
quences isolated frorn the chromosome of Laclococcus
laclis ssp. laclis. Appl. Environ. Microbjol. 57,341-348.
[35J Van de Guchle, M.. Van der Vossen, J.M.B.M.. Kok, J.
and Venerna, G. (1989) Construction of a lactococcal
expression vector: expression of hen egg while Iysozyme
jn Laclococcus laclis ssp. laclis. Appl. Envjron. Micro-
biol. 55, 224-228.
[36J Simons, G.. Van Asseldonk, M.. Rutten, G., Braks, A..
Nijhujs, M.. Hornes, M. and De Vos, W.M. (1990)
Analysjs of secretion signals of laclococci. FEMS Micro-
biol. Rev. 87, P24.

[50] Porter, E.V, and Chassy, B.M. (1988) Nucleotide se-
quence of the Jj-o-phosphogalactoside galactohydrolasegene of Laclobacillus casei: comparison to analogous
pbg genes of other Gram-positive organisms. Gene 62,
263-276.
[51] Sharp, P.M., Cowe, E., Higgins, D.G., Shields, D.C.,
Wolfe, K.H. and Wrighl, F. (1988) Codon usage pat-
terns in Escherichia co/i, Bacillus sublilis Saccharomyces
cerevisiae, Schizosaccharomyces pombe, Drosophilamelanogasler and Homo sapiens; a review of the consid-
erabie wilhin-species diversity. Nucleic Acids Res. 16,
8207-8211.[52J Ogasawa, N. (1985) Markedly unbiased codon usage in
Bacillus sublilis. Gene 40, 145-150.
[53J Holt, J.G. (1986) Bergey's Manual of Systematic Bacte-
riology, Vol. 2. Williams & Wilkins, Ballimore, MD.
[54] Gasson, M.J. and Davies, F.L. (1980) Conjugal transfer
of the drug resistance plasmid pAMJj in the lactic
streptococci. FEMS Microbiol. Letl. 7,51-53.
[55] Horinouchi, S. and Weisblum, B. (1982) Nucleolide
sequence and functional map of pC194, a plasmid that
specifies inducible chloramphenicol resistance. J. Bacte-
riol 150, 815-825.[56] Horinouchi, S. and Weisblum, B. (1982) Nucleotide
sequence and functional map of pE194, a plasmid that
specifies inducible resistance to macrolide, lincosamide,
and streptogramin type B anlibiolics. J. Bacteriol 150,
804-814
[57] Brehm, J., Salmond, G. and Minton, N. (1987) Sequence
of the adenine methylase gene of the SlreplococcUS
faecalis plasmid pAMJjI. Nucleic Acids Res. 15,3177.
[58J De Vos, W.M. and Simons, G. (1988) Molecular cloning
of lactose genes in dairy lactic streptococci: Ihe phos-
pho-Jj-galactosidase genes and their expression prod-
ucts. Biochimie 70,461-473.
[59] Van de Guchte, M., Kok, J. and Venerna, G. (1991).
Distance-dependent translational coupling and interfer.
ence in Laclococcus laclis. Mol Gen. Genet. 227,65-71.
[60] Van de Guchle, M., Van der Lende, T., Kok, J. and
Venerna, G. (1991) A possible contribulion of mRNA
secondary structure to translation initiation efficiency in
Laclococcus laclis. FEMS Microbiol Lett. 81, 201-208.
[61] Haandrikman, AJ. (1990) Development and use of a
broad-host-range vector for the simultaneous analysis of
divergent promoters. PhD Thesis, University of Gronin-
gen, Groningen.
[62] Overbeeke, N., Termorshuizen, G.H.M., Giuseppin,L.F., Underwood, D.R. and Verrips, C.T. (1990) Secre-
tion of the a.galactosidase from Cyamopsis le-
Iragonoloba (Guar) by Bacillus sublilis. Appl Environ.
Microbiol 56, 1429-1434.
[63] De Vos, W.M., Vos, P., Simons, G. and David, S. (1989)
Gene organization and expression in mesophilic lactic
acid bacteria. J. Dairy Sci. 72, 3398-3405.
[64] KIessen, C., Schmidt, K.H., Ferretti, JJ. and Malke, H.
(1988) Tripartite streplokinase gene fusion vectors for
gram-posilive and gram-negative procaryotes. Mol. Gen. 1
Genet. 212, 295-300. i
[65] Reference omitted. i
[66] Referenc" omitted. i
[67] Jolles, P. and Jolles, J. (1984) What's new in Iysozyme t
research? Mol. Cell. Biochem. 63, 165-189. {,
[68] Schleifer, K.H. and Kandier, 0. (1972) Peptidoglycan i
types of bacterial cell walls and their taxonomic implica-
tions. Bacteriol. Rev. 36, 407-477.
[69] Crawford, R.J.M. (1987) The use of Iysozyme in the
prevention of late blowing in cheese, Bulletin of Ihe
International Dairy Federation no.216. International
Dairy Federation, Brussels.
[70J Hughey, V.L. and Johnson, E.A. (1987) Antimicrobial
activity of Iysozyme against bacteria involved in food
spoilage and food-borne disease. Appl. Environ. Micro-
biol. 53,2165-2170.[71J Hughey, V.L., Wilger, P.A. and Johnson, E.A. (1989)
Antibacterial aclivity of hen egg white Iysozyme against
Lysteria monocytogenes Scott A in foods. Appl. Environ
Microbiol. 55, 631-638.
[72] Gasson, M.J. and Anderson, P.H. (1985) High copy
number plasmid veclors for use in laclic streptococci.
FEMS Microbiol. Lett. 30, 193-196.
[73J Leenhouts, K.J., Gieterna, J., Kok, J. and Venema- G
(1991) Chromosomal stabilization of the proteinase
genes in Lactococcus laclis. Appl. Environ Microbiol.
57, 2568-2575.
[74J Ganoza, M.C., Kofoid, E.C., Marlire, P. and Loui" BG
(1987) Potential secondary struclure at translation-ini.
tiation sites. Nucleic Acids Res. 15, 345-360.
[75J Sprengart, M.L., Fatscher, H.P. and Fuchs, E. (1990)
The initiation of translation in E. coli. apparent base
pairing between the 16SrRNA and downstream se-
quences of the mRNA. Nucleic Acids Res. 18, 1719-
1723.[76J Oppenheim, D.S. and Yanofski, C. (1980) Translational
coupling during expression of the tryptophan operon of
Escherichia coli. Genetics 95, 785-795.
[77J Das, A. and Yanofsky, C. (1984) A ribosome binding
site sequence is necessary for efficient expression of the
distal gene of a translationally-coupled gene pair. Nu-
cleic Acids Res. 12, 4757-4768.
[78J Gatenby, A.A., Rothstein, S.J. and Nomura, M. (1989)
Translational coupling of the maize chloroplasl alpB
and alpE genes. Proc. Natl. Acad. Sci. USA 86, 4066-
4070.
[79] Harms, E., Higgins, E., Chen. J.W. and Umbarger. H.E.
(1988) Translational coupling between the ilL"D and ilL"A
genes of Escherichia coli. J- Bacleriol. 170.4798-4807.
[80] Lindahl. L., Archer, R.H., McCormick, J.R., Freedman,L.P. and Zengel. J.M. (1989) Translational coupling of
the two proximal genes in the SIO ribosomal protein
operon of Escherichia coli. J. Bacleriol. 171, 2639-2645.
[81] Little, S., Hyde, S., Campbell, C.J., Lilley, RJ. and
Robinson. M.K. (1989) Translational coupling in the

threonine operon of Escherichia co/i K-12 J" Bacleriol"
171, 3518-3522
11!2] Schoner, B"E", Hsiung, H"M", Belagaje, R"M", Mayne,N"G and Schoner, R"G" (1984) Role of mRNA transla-
tional efficiency in bovine growth hormone expression
in Escherichia co/i" Proc" Natl" Acad" Sci" USA 81,
5403-5407"[83] Schiimperli, D", McKenney, K", Sobieski, D"A and
Rosenberg, M" (1982) Translational coupling at an in-
tercistronic boundary of the Escherichia coli galactose
operon" Cell 30, 865-871"
[1!4] Bonekamp, F", Dalbge, H", Christensen, T and Jensen,
K"F" (1989) Translation rates of individual codons are
nat correlated with tRNA abundances ar with frequen-
cies of utilization in Escherichia co/i" J" Bacteriol" 171,
5812-5816"[85J Grosjean, H" and Fiers, W" (1982) Preferential codon
usage in prokaryotic genes" the optimal codon-anti-
codon interaction energy and the selective codon usage
in efficiently expressed genes" Gene 18, 199-209"
[86] Ohno, S (1988) Codon preference is but an illusion
created by the construction principle of coding se-
quences" Proc" Natl" Acad" Sci" USA 85, 4378-4382
[87] Gualerzi, C"O" and Pon, C"L" ( 1990) Initiation of mRNA
translation in prokaryotes Biochemistry 29, 5881-5889"
[88] Brawerman, G" (1987) Determinants of messenger RNA
stability Cell 48, 5-6"
[89] Kubo, M", Higo, Y" and Imanaka, T" (1990) Biological
threshold values of procaryotic gene expression which is
conlrolled by the DNA inverted repeat sequence and
the mRNA secondary structure" J" Ferment" Bioeng" 69,
305-307"
[90] Von Heijne, G" and Abrahmsen, L" (1989) Species-
specific variation in signal peptide design; implications
for protein secretion in foreign hosts" FEBS Lett" 244,
439-446"[91J Ardo, Y" and Pettersson, H"E" (1988) Accelerated cheese
ripening with heat treated cells of Lactobaci/1us he/L'eti-
cus and a commercial proteolytic enzyme" J" Dairy Res"
55, 239-245"[92] Law, B"A" (1984) The accelerated ripening of cheese"
In" Advances in the Microbiology and Biochemistry of
Cheese and Ferme~ted Milk (Davies, F"L" and Law,
B"A.. Eds"), pp" 209-228" Elsevier, London"
[93] Law, B"A" and King, J"S" (1985) Use of liposomes for
proteinase addition to Cheddar cheese" J" Dairy Res" 52,
183-188"[94] Law, B"A" and Wigmore, A" (1982) Accelerated cheese
ripening with food grade proteinases" J" Dairy Res" 49,
137-146"
[95J Law, B"A" and Wigmore, A"S" (1983) Accelerated ripen-
ing of Cheddar cheese with a commercial proteinase
and intracellular enzymes from starter streptococci" J"
Dairy Res" 50, 519-525"
[96] Yang, M"Y.. Ferrari, E" and Henner, D"J" (1984) Cloning
of the neutral protease gene of Baci/1us subti/is and the
use of the cloned gene to create an in vitro-derived
deletion mutation. J. Bacteriol. 160, 15-21.
[97] Haandrikman, A.J.. Kok, J.. Laan, H.. Soemitro, S..
Ledeboer, A.M.. Konings, W.N. and Venerna, G. (1989)
Identification of a gene required for maturation of an
extrace1lular lactococcal serine proteinase. J. Bacteriol.
171, 2789-2794.
[98] Buchman, G.W.. Banerjee, S. and Hansen, J.N (1988)
Structure, expression, and evolution of a gene encoding
the precursor of nisin, a small protein antibiotic J Biol.
Chem. 263, 16260-16266.
[99] van Rooijen, R.J. and De Vos, WM. (1990) Molecular
cioning, transcriptional analysis, and nucleotide se-
quence of lacR, a gene encoding the repressor of the
lactose phosphotransferasesystem of Lactococcus lactis.
J. Biol. Chem. 265, 18499-18503.
[100] Tinoco, 1.. Borer, P.N.. Dengier, B.. Levine, M.D.. Uh-
lenbeck, O.C.. Crothers, D.M. and Gra1la, J. (1973)
Improved estimation of secondary structure in ribonu.
cieic acids. Nature New Biol 246, 40-41
[lol] Renault, p.. Gai1lardin, C. and Heslot, H (1989) Prod-
uct of the Laclococcus laclis gene required for malolac-
tic fermentation is homologous to a family of positive
regulators. J. Bacteriol 171,3108-3114
[102] Ross, P.. O'Gara, F. and Condon, S. (1990) Cloning and
characterization of the thymidylate synthase gene from
Lactococcus lactis ssp lactis Appl Environ. Microbiol
56, 2156-2163.[103] Mayo, B.. Kok, J.. Venerna, K.. Bockelm"nn, W.. Teu-
ber, M.. Reinke, H. and Venerna, G (1991) Molecul"r
cioning and sequence analysis of the x-prolyl dipeptidyl
aminopeptidase gene from Lactacoccus lactis ssp. cre-
maris. Appl Environ. Microbiol 57,38-44.
[104] Nardi, M.. Chopin, M.. Chopin, A.. Cals, M. and Gripon,
J. (1991) Cloning and DNA sequence analysis of an
x-prolyl dipeptidyl aminopeptidase gene from Lactocac-
cus laclis ssp. laclis NCDO763. Appl Environ. Micro-
biol 57,45-50.
[105] Boizet, B.. Vi1leval, D.. Slos, p.. Novel M.. Novel G.
and Mercenier, A. (1988) Isolation and structural analy-
sis of the phospho.p-galactosidase gene from Strepta.
coccus laclis Z268. Gene 62, 249-261.
[106] De Vos, W.M. and Gasson, M.J. (1989) Structure and
expression of the Laclacoccus laclis gene for phospho-
p.galactosidase (lacG) in Eschericha cali and L. lactis.
J. Gen. Microbioll35, 1833-1846.
[107] Hill C., Mi1ler, L.A. and KJaenhammer, T.R. (1990)
Nucleotide sequence and distribution of the pTR2030
resistance determinant (hsp) which aborts bacterio-
phage infection in lactococci. Appl Environ. Microbiol
56, 22S5-2258.
[108] Leenhouts, K.J.. Tolner, B.. Bron, S.. Kok, J.. Venerna,G. and Seegers, J.F.M.L. (1991) Nucleotide sequence
and characterization of the broad.host-range lactococcal
plasmid pWVOI. Plasmid 26, 55~66.
[109] Shearman, C., Underwood, H.. Jury, K. and Gasson, M.

[114] Rauch, P.J.G., Beerthuyzen, M.M. and De Vos, W.M,
(1990) Nucleotide sequence of 15904 from LaC/ococcus
/ac/is ssp. /ac/is strain NIZO R5. Nucleic Acids Res 18.
4253-4254.[[15] Kaletta, C. and Entian, K (!989) Nisin, a peptide an-
tibiotic: cloning and sequencing of the nisA gene and
posttranslational processing of its peptide product. J,
Bacteriol. 171, 1597-1601.
[[16] David, S" Van der Rest, M.E" Driessen, A.J.M" Si.
mons, G. and De Vos, W.M. (1990) Nucleotide se.
quence and expression in Esch.richia ca/i of th. Lac/a-
coccus /aC/is citrate permease gene. J Bacterioc 172,
5789-5794.
[117] Piggot, P.J. and Hoch, J.A. (!985) Revised genetic link-
age map of Bacillus sub/i/is. Microbioc Rev. 49, [58-179,
(1989) CJoning and DNA sequence analysis of a Lacto-
coccus bacteriophage Iysin gene. MoJ. Gen. Genet. 218,
214-221.
[110] PoJzin, K.M. and Shimizu-Kadota, M. (1987) Identifica-
tion of a new insertion eJement, simiJar to gram-nega-
tive IS26, on the lactose plasmid of Streptococcus lactis
ML3. J. BacterioJ. 169,5481-5488.[111] Romero, D.A. and KJaenhamrrier, T.R. (1990) Charac-
terization of insertion sequence IS946, an iso-ISS1 eJe-
ment, isolated from the conjugative lactococcaJ pJasmid
pTR2030. J. BacterioJ. 172,4151-4160.
[112] Haandrikman, A.J., Van Leeuwen, C., Kok, J., Vos, P.,
De Vos, W. and Venema, G. (1990) Insertion eJements
on JactococcaJ proteinase plasmids. AppJ. Environ. Mi-
crobiol.56, 1890-1896.
[113] Dodd, H.M" Horn, N. and Gasson, M.J. (1990) AnaJysis
of the genetic determinant for production of the peptide
antibiotic nisin. J. Gen. Microbiol. 136,555-566.





























