Embed Size (px)
Citation preview

Eur J Biochem 116, 109-116 (1981) (; FEBS 1981
General Shape and Hapten-Induced Conformational Changes of Pig Antibody against Dinitrophenyl A Small-Angle Scattering Study
Labzlo CSER. Frant ikk FRANgK, lrina A. GLADKIKH, Alexandr B. KUNCHENKO, and Yuriy M. OSTANEVICH
Laboratory of Neutron Physics, Joint Institute for Nuclear Research, Dubna; Central Research Institute for Physics, Hungarian Academy of Sciences, Budapest; and Institute of Molecular Genetics, Czechoslovak Academy of Sciences, Prague
(Received June 2, 1980/January 19, 1981)
Pig antibodies against dinitrophenyl were studied by neutron small-angle scattering and X-ray small-angle scattering with particular attention to the analysis of cross-section plots and determination of the radii of gyration of cross-section. The experimentally determined molecular parameters R, (radius of gyration), R, and R,z (two different radii of gyration of cross-section characterizing every antibody sample) show that the shapes of the two antibody types, precipitating and non-precipitating, are similar. The non-precipitating antibody is slightly more compact.
The parameters R,, R,1 and Rq2 of complexes of antibodies with the hapten, 8-dinitrophenyI-5,8-aza-4-0~0- octanoic acid, are smaller than those of the free antibody. This indicates that a conformational change is induced by the binding of the hapten. The character of the change of parameters is consistent with a view that the observed contraction of the molecule proceeds via similarity transformation.
In order to design a model of a pig antibody molecule, isolated building blocks of the molecule, the Fab and Fc fragments, were first studied. A comparison of the scattering curves with various models of fragments showed, however, that the isolated fragments acquire in solution elongated rod-like shapes.
Over 300 tentative models of the intact antibody molecule, built of small identical spheres, were constructed before a good fit with the experimental data was achieved. The most probable models have a cavity in the Fc part and the Fab parts are either fully extended or slightly bent downwards to the Fc part.
Although detailed information on the three-dimensional structure of immunoglobulin domains is available from X-ray analysis of crystalline immunoglobulin fragments [I], much uncertainty still exists concerning immunoglobulin size, shape and dynamic behaviour in solution. In particular, the charac- ter of conformational changes of antibody molecules following the binding of specific ligands, antigens or haptens, is far from being perfectly understood [2,3].
A vast body of experimental data leads to seemingly con- troversial conclusions. Some methodical approaches failed to detect any changes that could be correlated with antigen or hapten binding. These were for example, the circular dichro- ism study [4], molecular motion studies of an antibody- Gd(lI1) complex [ 5 ] , or examination of accessibility of the antibody hinge region to reduction or to proteolysis [6]. In other systems investigated, however, small but significant changes of molecular parameters were observed, indicating that the liganded antibody acquired an altered conformation. This conclusion was drawn using following methodical ap- proaches: the hydrogen-deuterium exchange [7], circular polarization of fluorescence [S], temperature-jump relaxation [9], fluorescence depolarization [lo], and electron spin reso- nance of spin-labelled probes [I 1,121.
X-ray small-angle scattering employed by Pilz et al. [I 3,141 for investigation of free and liganded rabbit antibody to the poly(u-alanyl) group and to p-azophenyl-/?-lactoside group
showed, that binding of hapten was accompanied by a de- crease of the radius of gyration. In our previous neutron small- angle scattering investigations [I51 of two types of pig anti- bodies to dinitrophenyl a similar effect was observed : the radii of gyration of antibody-hapten complexes were signifi- cantly smaller than those of free antibodies. In another study Pilz et al. [I61 demonstrated that radii of gyration of antibody Fab’ and F(ab’)Z fragments were not influenced by hapten binding. The authors conclude that the contraction effect must be due to the presence of the Fc part in the intact molecule. The inter-heavy chain disulfide bridge seems to be essential for the transition occurring upon reaction with hapten. Antibodies with an interrupted disulfide bridge did not display any significant change of molecular parameters related to hapten binding [I 71.
At high values of the transferred momentum, commonly designated as it’, both X-ray [13,14,18,19] and neutron [20] small-angle scattering experiments yield similar typical pat- terns of the so-called cross-section plot [19], i.e. In ( I x x) versus xz. The cross-section plot regularly consists of two straight-line parts with different slopes. Pilz et al. [I 3,141 concluded that this type of scattering curve can be explained by assuming that immunoglobulin G molecules are T-shaped.
0 ; H is the scattering angle, i is the wave-
__ __ 4 n 2 z ’ x = . ~- sin 012 =
length. n 1.

In the present paper we describe the results of neutron and X-ray small-angle scattering studies in pig antibodies to dinitrophenyl. The range of experimental conditions was extended to higher values of x. Neutron small-angle scattering curves obtained in this range displayed new features pertinent to hapten-induced antibody conformational changes. By fitting the experimental curves with computed scattering curves of a number of model bodies, a probable model of pig antibody molecule was constructed.
MATERIALS AND METHODS
Ant ihodies
Antibodies to dinitrophenyl were isolated from sera of pigs immunized by dinitrophenylated bovine immunoglobulin [21,22]. Precipitating antibody, also called early antibody [15], was isolated from sera collected at an early phase of the immune response [23,24]. The non-precipitating, or late [I 51 antibody was isolated from sera collected at a late phase of the immune response [23,24]. Both antibody types were of the immunoglobulin G class ( M , = 150000). The preparations used in the present work were the same as those used earlier [ 151. The association constant of E-dinitrophenyl-L-lysine binding was 6.7 x los M- l for the precipitating and 1.8 x l o7 M-' for the non-precipitating antibody.
An tibody Frugmen ts
Fab and Fc fragments of non-specific pig immunoglo- bulin G were prepared by cleavage with trypsin and separated by gel chromatography and ion-exchange chromatography [25]. Their purity was checked by immunoelectrophoresis using properly absorbed rabbit antisera.
An iihody- Hap ten Complexes
The hapten used in all experiments was 8-dinitrophenyl- 5,8-aza-4-oxooctanoic acid. It was synthesized as described before [15]. Antibody- hap ten)^ complexes were prepared by dissolving weighed amounts of hapten in antibody solution. The hapten was added in a slight excess over the stoichiometric ratio, i.e. 2.2 mol hapten/mol antibody. According to the values of the association constant, more than 90% of the binding sites in both antibody types are expected to be occupied by the hapten under the conditions used.
Preparation of Sumples .for Small-Angle Scattering
Neutron scattering was performed with 2 % (w/v) anti- body dissolved in 100 mM sodium phosphate buffer. In order to achieve maximum contrast with minimum incoherent back- ground, the buffer was prepared in deuterium oxide and adjusted to apparent pH 6.36. For X-ray scattering 2 (w/v) antibody solutions were prepared in 100 mM sodium phos- phate buffer pH 6.4 using ordinary water as the solvent.
The antibody solutions were equilibrated by dialysis against the relevant buffer. Possible aggregates were removed from the solutions by centrifugation at 6000 x g for 20 min. The absence of aggregates and dimers was checked by sedi- mentation velocity analysis in an ultracentrifuge Spinco model E.
Small-Angle Scattering Technique The neutron scattering experiments were performed at the
time-of-flight small-angle scattering facility with axial-sym- metric geometry of the Neutron Physics Laboratory of the Joint Institute of Nuclear Research (Dubna, USSR) [26]. X-ray scattering experiments were carried out using the Kratky small-angle scattering camera at the protein Research Institute, Academy of Sciences of the USSR (Poustchino, USSR). X-ray intensity distributions were in all cases cor- rected for collimation distortion using described methods [27].
Radii of gyration of cross-section and their errors were evaluated from neutron scattering curves by the method of least-squares fitting. The points of a straight-line part of the scattering curve were approximated by a first-order linear function of xz. It would have been meaningless to perform a similar evaluation for the X-ray data to estimate the error of the radii of gyration of cross-section because of the existence of strong correlation between the points arising upon correc- tion for collimation distortion effect.
Model Computation and Design
Because of random orientation of the dissolved molecules it is impossible to obtain the three-dimensional shape of the molecule from the small-angle scattering intensity curve alone. However, from the known shape one can always evaluate the scattering intensity distribution and compare it with the observed one. Thus, when some independent knowledge on the shape of the molecule is available (e.g. from stereo- chemical data or X-ray crystallography) the trial and error approach makes it possible to construct meaningful molec- ular models. The procedure consists of generation of a set of possible models and, upon calculation of corresponding intensity curves, selection of those models that give the best fit with experimental data.
Earlier i t was shown by Pilz et al. [I81 that the scattered intensity distribution, including the Guinier range and the two straight-line parts of the cross-section curve, contains enough information for comparison with models of anti- bodies and their fragments. In the present work we used the method of hard spheres [28] to evaluate the scattered intensity curves of pig antibodies and to construct molecular models. This method utilized a generalization of the Debye formula
where , f i and ,h are the scaltering amplitudes of atoms or nuclei situated at points i and k , respectively, rik is the distance between the above points.
According to the above-mentioned method [28] the body of the molecule can be constructed, in a good approximation, from a number of homogeneous identical spheres. Then formula (1) may be transformed into
here g (x) = (9 n/2) [J3/2(x R)I2. ( x R ) - ~ is the scattering inten- sity curve of a sphere having a radius R, n is the number of spheres filling the body, Ai represents the number of distances ~ i k = rj, m is the number of different distances rj and J is the Bessel function.
For simplicity, and also to cut down the computer time, the spheres should be arranged in a simple way, e.g. their
I l l
centres may form a primitive cubic lattice. The diameter of the spheres must have a value less than the resolution of the small-angle experiment, i.e. 1 - 2 nm. This computer model can be visualized by real spheres stuck together. Models illustrated in this paper were prepared from steel balls with a diameter of 5 min and were stuck with picein.
RESULTS Antibodies to Dinitrophenyl and Their Complexes with Hapten
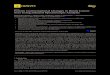
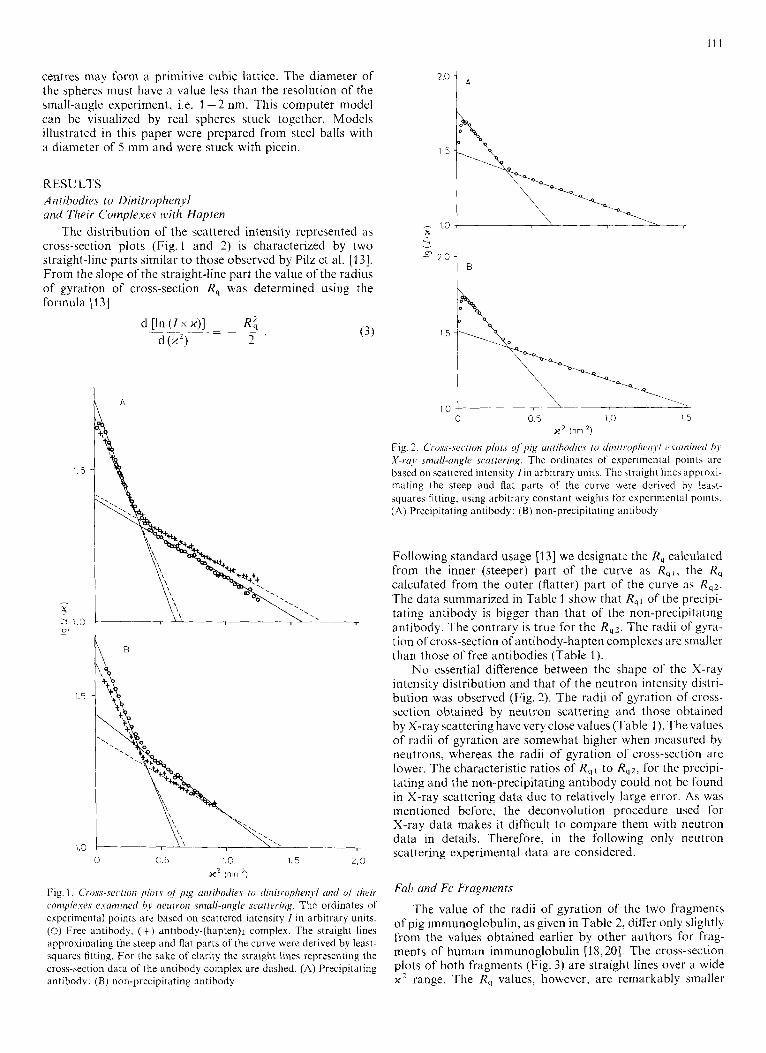
The distribution of the scattered intensity represented as cross-section plots (Fig.1 and 2) is characterized by two straight-line parts similar to those observed by Pilz et al. [13]. From the slope of the straight-line part the value of the radius of gyration of cross-section R, was determined using the formula [I 31
- - d [In ( I x x)] d ( x ' ) ~~- -
1.5
0 0.5 1.0 I. 5 2.0 x2 (nrn-')
Fig. I . Cross-seciion / J / ( J / , S of pig unrihodies lo clirzirrophrnj~l and of rlzrir. conzplcst~s e Tarnineif hy neutron snrall-cmgle scattering. The ordinates of experimental points are based on scattered intensity I in arbitrary units. (0) Free antibody. (+) antibody-(hapten)z complex. The straight lines approximating the sleep and flat parts of the curve were derived by least- squares fitting. For Llie sake of clarity the straight lines representing the cross-section data of the antibody complex are dashed. (A) Precipitating antibody; (B) non-precipitating antibody
2.0 i .
4
2.0 1
Fig. 2. C'i.os,s-.sc.c,rion p1ot.c of pig nnrihodic.c. ro dinilro/~heii>,i vcinzintd hy X-r.o,r smull-angle scarteriirg. The ordinates of exper~mental points are based on scattered intensity f in arbitrary units. The straight lines approxi- mating the steep and flat parts of the curve were derived by least- squares fitting, using arbitrary constant weights for experimental points. (A) Precipitating antibody; ( B ) non-precipitating antibody
Following standard usage [13] we designate the R, calculated from the inner (steeper) part of the curve as R q I , the R, calculated from the outer (flatter) part of the curve as R,'. The data summarized in Table 1 show that Rql of the precipi- tating antibody is bigger than that of the non-precipitating antibody. The contrary is true for the Rq2. The radii of gyra- tion of cross-section of antibody-hapten complexes are smaller than those of free antibodies (Table 1).
No essential difference between the shape of the X-ray intensity distribution and that of the neutron intensity distri- bution was observed (Fig. 2). The radii of gyration of cross- section obtained by neutron scattering and those obtained by X-ray scattering have very close values (Table 1 ). The values of radii of gyration are somewhat higher when measured by neutrons, whereas the radii of gyration of cross-scction are lower. The characteristic ratios of R q l to Rq2, for the precipi- tating and the lion-precipitating antibody could not be found in X-ray scattering data due to re!atively large error. As wa5 mentioned before, the deconvolution procedure used for X-ray data makes it difficult to compare them with neutron data in details. Therefore, in the following only neutron scattering experimental data arc considered.
Fa17 ond Fc Fragments
The value of the radii of gyration of the two fragments of pig immunoglobulin, as given in Table 2, differ only slightly from the values obtained earlier by other authors for frag- ments of human immunoglobulin [18,20]. The cross-section plots of both fragments (Fig. 3) are straight lines over a wide x2 range. The R, values, however, are remarkably smaller
112
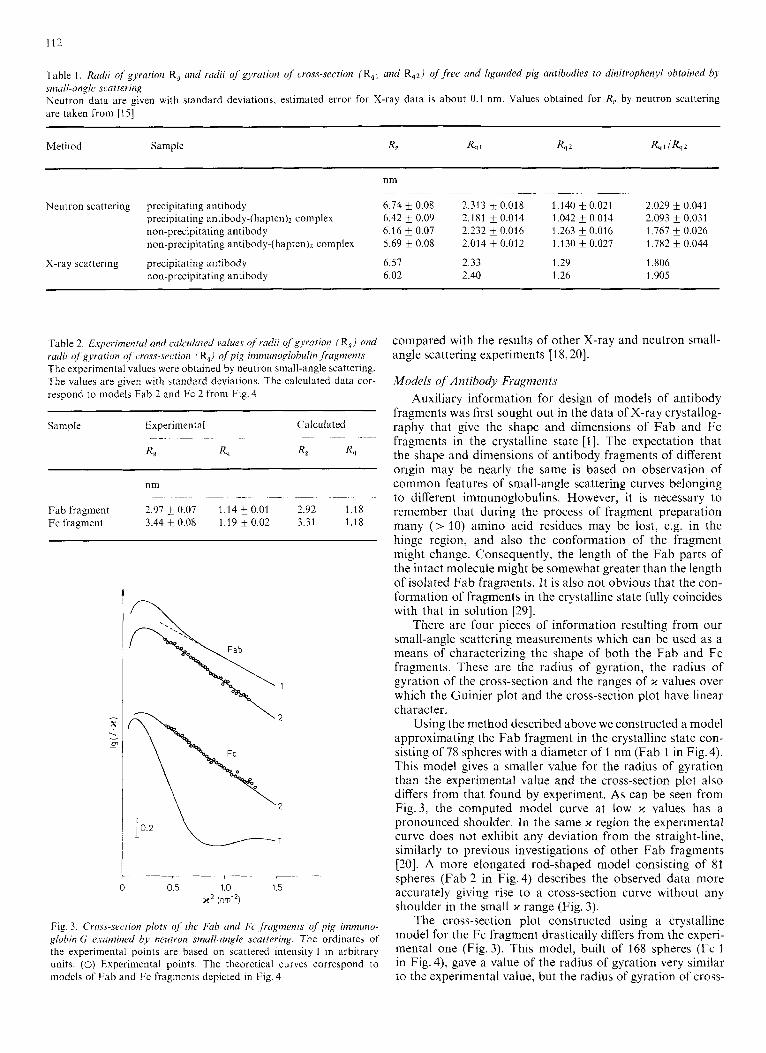
Table I , Rudii of gyration R, and radii of gyration of cross-section (R,1 and Rq2/ of f ree and liganded pig antibodies to dinitrophenyl obtained by small-angle Jcattering Neutron data are given with standard deviations, estimated error for X-ray data is about 0.1 nm. Values obtained for R, by neutron scattering are taken from [I51
Method Sample R, R, 1 R, 2 R,llR,2
nm ~~~ ~~ ~ ~~~~ ~~~~~
Neutron scattering precipitating antibody 6.74 0.08 2.313 f 0.018 1.140 f 0.021 2.029 i 0.041 precipitating antibody-(hapten)z complex 6.42 f 0.09 2.181 k 0.014 1.042 f 0.014 2.093 k 0.031 non-precipitating antibody 6.16 & 0.07 2.232 i 0.016 1.263 f 0.016 1.767 & 0.026 non-precipitating antibody-(hapten)z complex 5.69 ?r 0.08 2.014 0.012 1.130 f 0.027 1.782 f 0.044
X-ray scattering precipitating antibody 6.57 2.33 1.29 I .806 non-precipitating antibody 6.02 2.40 1.26 1.905
Table 2. E.xperimmtul und c.uk.uluied values of rudiii of gyvurion (R,) and radii of gyration of cross-seclion (%) o/pig immunoglohuli?~ fragments The experimental values were obtained by neutron small-angle scattering. The values are given with standard deviations. The calculated data cor- respond to models Fab 2 and Fc 2 from Fig. 4
Sample Lxperimental CdiCUldted ~ ~ ~ -~ ~
R , R, R, R,
nm - ~ ~ __ -. ~
Fab fragment 2 9 7 + 0 0 7 1 1 4 + 0 0 1 2 9 2 1 1 8 Fc fragment 3 4 4 k O O X 1 1 9 k O O 2 731 118
~~~ ~
modcls of Fab and Fc fragments depicted i n Fig 4 to the experimental value, but the radius of gyration of cross-
t 0 0.5 1.0 1.5
x 2 (nm-2)
Fig. 3. Cross-section plots of the Fuh and Fc f?ugments (If pig immuno- globin G e..wmined bv neutruiz smalf-angle scattering. The ordinates of the experimental points are based on scattered intensity 1 in arbitrary units. (0) Experimental points. The theoretical curves correspond to
compared with the results of other X-ray and neutron small- angle scattering experiments [I 8,201.
Models qf Antibody Frugmmts Auxiliary information for design of models of antibody
fragments was first sought out in the data of X-ray crystallog- raphy that give the shape and dimensions of Fab and Fc fragments in the crystalline state [I]. The expectation that the shape and dimensions of antibody fragments of different origin may be nearly the same is based on observation of common features of small-angle scattering curves belonging to different immunoglobulins. However, it is necessary to remember that during the process of fragment preparation many (> 10) amino acid residues may be lost, e.g. in the hinge region, and also the conformation of the fragment might change. Consequently, the length of the Fab parts of the intact molecule might be somewhat greater than the length of isolated Fab fragments. It is also not obvious that the con- formation of fragments in the crystalline state fully coincides with that in solution [29].
There are four pieces of information resulting from our small-angle scattering measurements which can be used as a means of characterizing the shape of both the Fab and Fc fragments. These are the radius of gyration, the radius of gyration of the cross-section and the ranges of 2c values over which the Guinier plot and the cross-section plot have linear character.
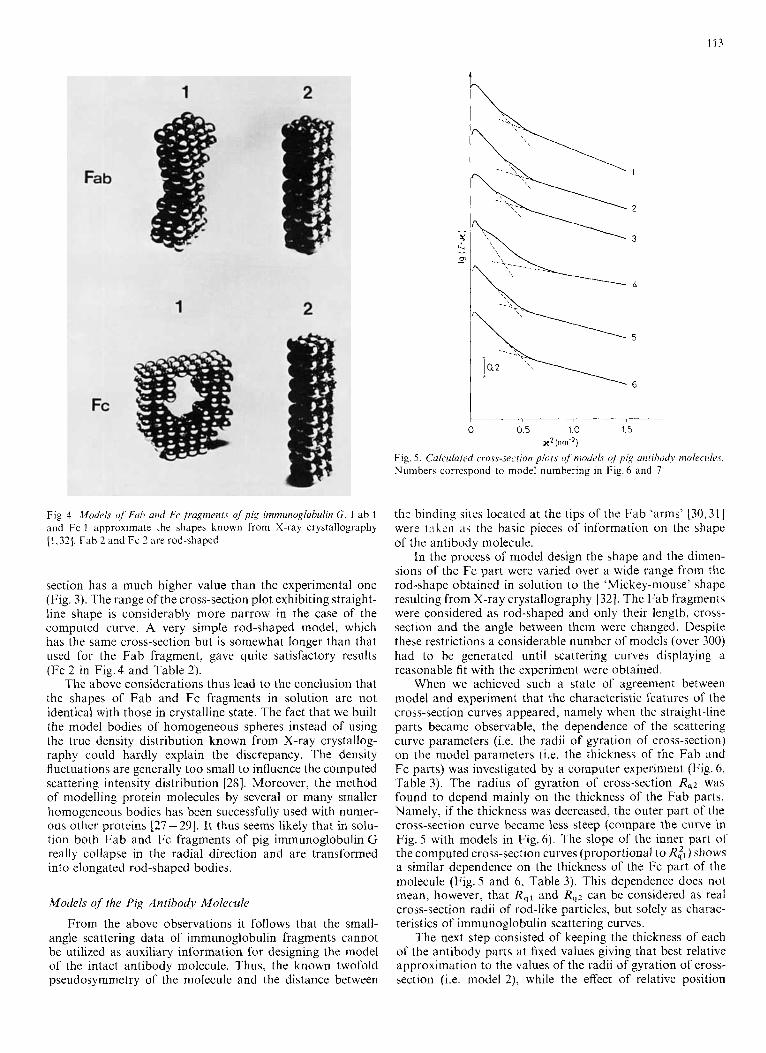
Using the method described above we constructed a model approximating the Fab fragment in the crystalline state con- sisting of 78 spheres with a diameter of 1 nm (Fab 1 in Fig. 4). This model gives a smaller value for the radius of gyration than the experimental value and the cross-section plot also differs from that found by experiment. As can be seen from Fig.3, the computed model curve at low 1c values has a pronounced shoulder. In the same 2c region the experimental curve does not exhibit any deviation from the straight-line, similarly to previous investigations of other Fab fragments [20]. A more elongated rod-shaped model consisting of 81 spheres (Fab 2 in Fig. 4) describes the observed data more accurately giving rise to a cross-section curve without any shoulder in the small i( range (Fig. 3).
The cross-section plot constructed using a crystalline model for the Fc fragment drastically differs from the experi- mental one (Fig. 3). This model, built of 168 spheres (Fc 1 in Fig. 4), gave a value of the radius of gyration very similar
113
0 L 0.5 1.0 1.5
x2 ( n d )
Fig. 5. Culculuted c.ro.vs-section plors of’niode1.s ( I / pig trrirrhoc!,~ molec.ul~~.s. Numbers correspond to model numbering in Fig. h and 7
Fig. 4. Mode/,s of’ Fuirh and Fe fragments of pig imniunoglohulin G. Fab 1 and Fc 1 approximate the shapes known from X-ray crystallography [l , 321. Fah 2 and Fc 2 are rod-shaped
the binding sites located at the tips of the Fab ‘arms’ [30,31] were ta!,c.n ;is the basic pieces of information on the shape of the antibody molecule.
In the process of model design the shape and the dimen-
section has a much higher value than the experimental one (Fig. 3). The range of the cross-section plot exhibiting straight- line shape is considerably more narrow in the case of the computed curve. A very simple rod-shaped model, which has the same cross-section but is somewhat longer than that used for the Fab fragment, gave quite satisfactory results (Fc 2 in Fig.4 and Table 2).
The above considerations thus lead to the conclusion that the shapes of Fab and Fc fragments in solution are not identical with those in crystalline state. The fact that we built the model bodies of homogeneous spheres instead of using the true density distribution known from X-ray crystallog- raphy could hardly explain the discrepancy. The density fluctuations are generally too small to influence the computed scattering intensity distribution [28]. Moreover, the method of modelling protein molecules by several or many smaller homogeneous bodies has been successfully used with numer- ous other proteins [27 - 291. It thus seems likely that in solu- tion both Fab and Fc fragments of pig immunoglobulin G really collapse in the radial direction and are transformed into elongated rod-shaped bodies.
Models of the Pig Antibody Molecule
sions of the Fc part were varied over a wide range from the rod-shape obtained in solution to the ‘Mickey-mouse’ shape resulting from X-ray crystallography [32]. The Fab fragments were considered as rod-shaped and only their length, cross- section and the angle between thein were changed. Despite these restrictions a considerable number of models (over 300) had to be generated until scattering curves displaying a reasonable fit with the experiment were obtained.
When we achieved such a state of agreement between model and experiment that the characteristic features of the cross-section curves appeared, namely when the straight-line parts became observable, the dependence of the scattering curve parameters (i.e. the radii of gyration of cross-section) on the model parameters (i.e. the thickness of the Fab and Fc parts) was investigated by a computer experiment (Fig. 6, Table 3). The radius of gyration of cross-section R,z was found to depend mainly on the thickness of the Fab parts. Namely, if the thickness was decreased, the outer part of the cross-section curve became less steep (compare the curve in Fig. 5 with models in Fig. 6). The slope of the inner part of the computed cross-section curves (proportional to R:,) shows a similar dependence on the thickness of the Fc part of the molecule (Fig. 5 and 6, Table 3). This dependence does not mean, however, that R,I and Rq2 can be considered as real cross-section radii of rod-like particles, but solely as charac-
From the above observations it follows that the small- angle scattering data of immunoglobulin fragments cannot be utilized as auxiliary information for designing the model of the intact antibody molecule. Thus, the known twofold pseudosymmetry of the molecule and the distance between
teristics of immunoglobulin scattering curves. The next step consisted of keeping the thickness of each
of the antibody parts at fixed values giving that best relative approximation to the values of the radii of gyration of cross- section (i.e. model 2), while the effect of relative position
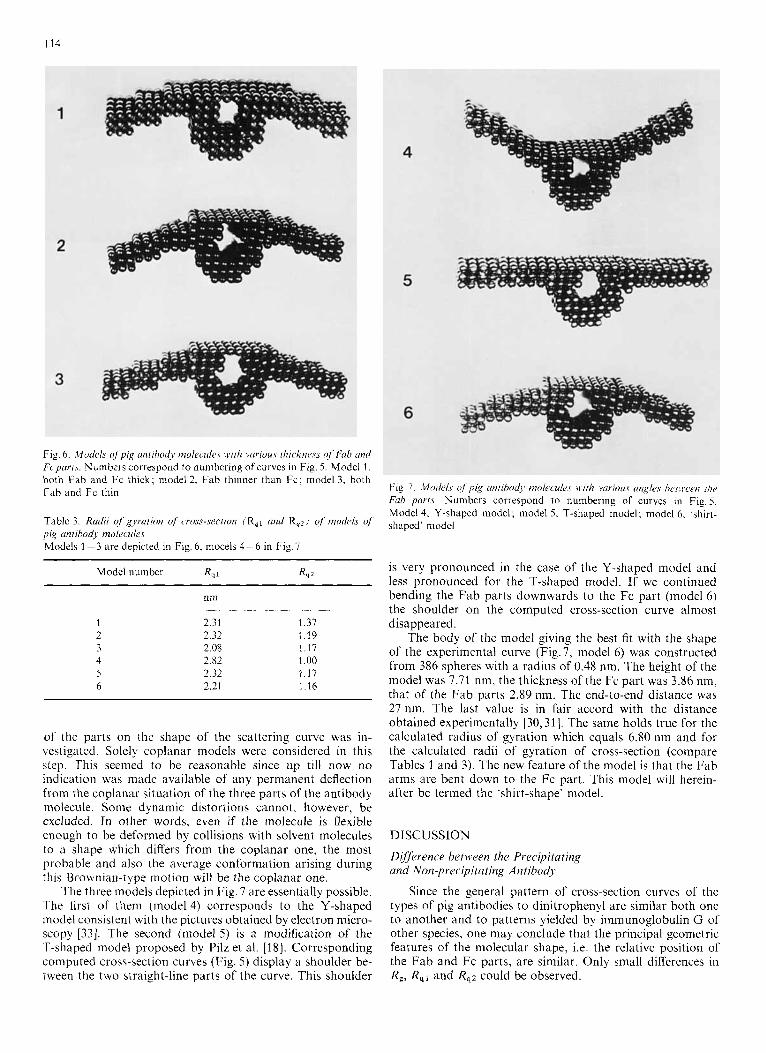
1 I4
Fig. 6. Models ofpig utifihodj mo1ecule.c. with vciriuus i1iicknr.r.s of Fab und Fcpcrrts. Numbers correspond to numbering of curves in Fig. 5. Model 1 . both Fab and Fc thick; model 2, Fab thinner than Fc; model 3, both Fab and Fc thin
Table 3. Radii of’ gyration of cross-section (R,, und R,z) of models of pig antibody molecules Models 1 - 3 are depicted in Fig. 6, mocels 4 - 6 in Fig. 7
Model number R , 1 R , 2
nm
2 31 1 3 1 2 12 119 2 08 1 1 1 2 82 1 0 0 2 32 1 1 7 2 21 1 1 6
~ ~
of the parts on the shape of the scattering curve was in- vestigated. Solely coplanar models were considered in this step. This seemed to be reasonable since up till now no indication was made available of any permanent deflection from the coplanar situation of the three parts of the antibody molecule. Some dynamic distortions cannot, however, be excluded. In other words, even if the molecule is flexible enough to be deformed by collisions with solvent molecules to a shape which differs from the coplanar one, the most probable and also the average conformation arising during this Brownian-type motion will be the coplanar one.
The three models depicted in Fig. 7 are essentially possible. The first of them (model4) corresponds to the Y-shaped model consistent with the pictures obtained by electron micro- scopy [33]. The second (model 5) is a modification of the T-shaped model proposed by Pilz et al. [18]. Corresponding computed cross-section curves (Fig. 5 ) display a shoulder be- tween the two straight-line parts of the curve. This shoulder
t i g . 7. .Motlc.ls o/ j 7 i , ~ uiiiihixlj moleculc~s will? vurious citzglcs h c t w c w / / I ( ,
Fub p a r i s . Numbers correspond to numbering of curves in Fig.5. Model 4. Y-shaped model; model 5 , T-shaped model; model 6, ‘shirt- shaped’ model
is very pronounced in the case of the Y-shaped model and less pronounced for the T-shaped model. If we continued bending the Fab parts downwards to the Fc part (model 6) the shoulder on the computed cross-section curve almost disappeared.
The body of the model giving the best fit with the shape of the experimental curve (Fig.7, model 6) was constructed from 386 spheres with a radius of 0.48 nm. The height of the model was 7.71 nm, the thickness of the Fc part was 3.86 nm, that of the Fab parts 2.89 nm. The end-to-end distance was 27 nm. The last value is in fair accord with the distance obtained experimentally [30,31]. The same holds true for the calculated radius of gyration which equals 6.80 nm and for the calculated radii of gyration of cross-section (compare Tables 1 and 3). The new feature of the model is that the Fab arms are bent down to the Fc part. This model will herein- after be termed the ‘shirt-shape’ model.
DISCUSSION
Difference between the Precipitating and Non-precipitating Antibody
Since the general pattern of cross-section curves of the types of pig antibodies to dinitrophenyl are similar both one to another and to patterns yielded by immunoglobulin G of other species, one may conclude that the principal geometric features of the molecular shape, i.e. the relative position of the Fab and Fc parts, are similar. Only small differences in R,, R,1 and Rq2 could be observed.

Table 4. Contraction ratio.r o / the radii of gyration arid of the radii of x j w t ion of cross-sw i ion 6 is the inean contraction ratio of i o K g , a R q l and ? R q Z . 6 is the mean 0 of precipitating and non-precipitating antibody. Both p and 5 are weighted mean values calculated from the Q R ; with weights = u;’, where oi ctands for the errors of Q~~ ( i = g, q l or q2). The wci@ilcd errors of 0 and
Contraction ratio Value for
are calculated using the usual formula u,” = C 7 l-’?:
precipitating non-precipitating antibody antibody
P K g 1.05 + 0 0 2 1.08 * 0.02 Q K q 1 1.06 0.01 1.11 _+ 0.01 Q R q Z 1.10 s o . 0 2 1.12 k 0 . 0 3 Q 1.065 f 0.008 1.105 f 0.009 Q 1.083 & 0.006
Because the molecular weight of both antibody types is the same, i.e. 150000, the smaller radius of gyration of the non-precipitating antibody indicates that the mass of the non-precipitating antibody is distributed more compactly compared with the precipitating antibody. This view is in agreement with the recent observation that the distance be- tween the two binding sites of the same antibody molecule is smaller in the case of non-precipitating antibody than with the precipitating antibody [30,31].
The radii of gyration ofcross-section R,z which correspond to the cross-section of the Fab part of the molecule [IS] suggest that the Fab parts of the non-precipitating antibody are thicker by about 10% than the Fab parts of the precipi- tating antibody. As far as the difference between the R,, values is concerned, it is not possible to make a straight- forward conclusion from it, since the main contribution of the Fc part (Fig. 5 ) is influenced by smaller contributions of the Fab parts. We may thus conclude that the shapes of the precipitating and non-precipitating antibodies are similar, but the Fab ‘arms’ of the latter are likely to be shorter and thicker. The picture based on the neutron small-angle scat- tering data is not unlikely considering previous knowledge. The overall length of the Fab fragment depends necessarily on the angle between its constituent parts, i.e. the variable and the constant domains. This angle is known to vary in different species of Fab fragments studied by X-ray crystallog- raphy [I] .
Effkcr of Hupten Binding
Our recent finding that the radii of gyration of liganded pig antibodies are smaller than those of free antibodies [I51 was in agreement with earlier reports ofPilz et al. [13,14,17,18] on rabbit antibodies. These authors, however, observed only a change of the radii of gyration; no significant change of the radii of gyration of cross-section was found. Our present data obtained with pig antibodies show clearly that both R,1 and R,z values change as a result of complex formation with the hapten.
Looking for some rule of the change of different parameters of the scattering curves, we produced contraction ratios of all mutually corresponding radii, i.e.
R, (free antibody) R, (liganded antibody) R, I (free antibody) R,, (liganded antibody)
QRg -= - - - -- - -~
(4b) - ( I = 1,2). _ _ Q R q , = -
The values summarized in Table 4 show that all values are close to their common average 5. This means that all three radii belonging to the free antibody may be transformed into the radii of corresponding antibody-(hapten)* complexes when multiplied by i. Moreover, if one multiplies the ab- scissa of the scattering curve of free antibody by 5, the trans- formed scattering curve coincides with that of the antibody- (hapten)2 complex within the range of experimental un- certainties.
The similarity of the shape of the scattering curves of free and liganded antibody leads to a suggestion that thc whole antibody molecule contracts via similarity transformation when binding the hapten and that the relative positions of the Fab and Fc parts do not change significantly during the contraction. This view must be taken with some reservation, however, because there is no unique relation between the shape and the observed small-angle scattering intensity distri- bution, i.e. more than one shape can yield the same scattering pat tern.
A contraction of the antibody molecule such that the shape does not change significantly would only be possible if, first, all parts of the molecule are involved, and second, the free antibody molecule is not packed with maximum density. The latter requirement is obviously fulfilled, because the existence of interdomain channels and clefts in immuno- globulin fragments has been shown even in the crystalline state.
It is necessary to bear in mind that the antibody mole- cules display significant segmental flexibility. Also the inter- domain clefts obviously fluctuate between closed and open states. Consequently, the distances between the centres of gravity of individual domains are subject to fluctuations. Small-angle scattering methods present smoothed average values of the molecular parameters. The concentration of the antibody molecule upon hapten binding may also be inter- preted as a shift of the equilibria between open and closed states of the clefts towards the closed states in the liganded form. We suppose that the contraction of the molecules con- cerns mostly or solely changes of interdomain clefts and that the chain folding in individual domains does not undergo any rearrangement resulting in higher packing density of the domains.
Since the contraction of the radii of gyration of cross- section was not observed in the case of rabbit antibodies [13, 141, it remains an open question whether the nature of the molecular contraction discussed above is specific for pig antibodies or whether it has a more general validity. Con- formational changes following hapten binding might be analo- gous in other species but not exactly the same.
Shape of Antibody Mo1cJcmle.r and qf Their Fsugmcnts
When determining the shape of large multi-domain mole- cules such as the molecules of antibodies, it is tempting to approach the problem by studying first the shape of indivi- dual constituent parts of the molecule, i.e. the shape of enzymatic fragments Fab and Fc. This approach, however, failed, because the fragments were found to acquire elongated rod-like shapes in solution. This observation is in agreement with earlier results of Pilz el al. [34] obtained with fragments of a human immunoglobulin. Thus, a model built of three rod-like particles would give a scattering curve very different from the experimental one. The relative positions of the domains in the antibody molecule obviously depend on the integrity of the molecule

116
The auxiliary information for designing iiiodels of the whole molecule was thus obtained from other sources. The shape of the Fc fragment in its crystalline form could be included almost without change in the model giving a good approximation of the experimental scattering curve. The loose character of the CH2 domain, as revealed by a crystallo- graphic study [32], was thus found to be consistent with the character of the cross-section plots yielded by antibodies in solution.
The models proposed in the present paper visualize the gross conformational features of pig antibodies but do not demonstrate the fine differences between the precipitating and the non-precipitating antibody. The large value of the radius of gyration (6.80 nm) and the distance between the tips of the Fab parts (27 nm) of model 6 are close to the experimental values of the precipitating antibody which is assumed to exist in an extended form [30]. A more accurate fit of modelled and experimental values could be achieved by refinement of the choise of the thickness of different parts of the molecule. However, such a refinement would require elementary spheres for model construction which were no more than half the size of these used above. Since the com- puter time increases proportionally with the 6th power of the reciprocal diameter of the spheres, the diameter of the spheres has to be chosen realisticly.
Both the T-shape model of Pilz et al. [18] and our ‘shirt- shape’ model describe rigid molecules. This is, of course, not a perfect description, since immunoglobulin molecules are endowed with segmental flexibility. The experimentally deter- mined scattered intensity is in fact a superposition of scattered intensity of molecules at different momentary conformations. The ‘shirt-shape’ model display a better fit with the shape of the scattering curve (cf. Fig. 1 and 9, whereas the T-shape model shows a closer agreement with the experimental Rql value (cf. Tables 1 and 3). The fact that two different rigid models may be in reasonable agreement with the experimental intensity distribution produced by flexible molecules allows us to suppose that in solution the antibody molecules may exist both in the T-shaped and in the ‘shirt-shaped’ form.
A detailed quantitative analysis of the molecular flexibility will be performed after further experimental data are ob- tained on the dynamic character of the molecule.
The authors are indebted to Dr 1. N. Serdyouk (Institute of Protein Research. Poustchino) for valuable consultations and for allowing us carry out the X-ray measurements. We thank Prof. I. M. Frank for his continuous interest in this work. We thank also Dr G. A. Medgyesi (Institute of Haematology, Budapest) for valuable discussion.
REFERENCES
1. Davies, D. R., Padlan, E. A & Segal, D. M. (1978) Annu. Rev. Biochem. 44, 639 - 667.
2. 3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14. 15.
16
17.
18.
19 20.
21 22.
23 24
28
26
27.
28
29 30.
31.
32.
33 34
Metzger, H . (1974) Adv. Immunol. 18, 169-207. Huber, R., Deisenhofer, J., Colman, P. M., Matsushinia, M. & Palm,
Cathou, R. E., Kulczycki, A, , Jr & Haber, E. (1968) Biochemistry,
Burton, D. R., Forstn, S., Frantk, F. & Novotny, J . (1979) FEBS
Wright, J . K., Engel, J. & Jaton, J.-C. (1978) Eur. J . Immunol. 8,
Liberti, P. A,, Stylos, W. A,, Maurer, P. H. & Callahan, H. J. (1972)
Schlessinger, J., Steinberg, I . Z., Givol, D., Hochman, R. & Pecht,
Lancet, D. & Pecht. I. (1976) Proc. Nut1 Acad. Sci. USA, 73, 3849-
Tumerman, L. A, , Nezlin, R. S. & Zagyansky, Y. A. (1972) FEBS
KBivarainen, A. 1. & Nezlin, R . S. (1976) Immunochemistry, I S ,
Timofeev, V. P., Dudich, 1. V., Sykulev, Yu. K., Nezlin, R. S. &
Pilz, I., Kratky, O., Licht, A. & Sela, M. (1973) Biochemistry, 12.
Pilz, I . , Kratky, 0. & Karush, F. (1974) Eur. J . Biochem. 41, 91 -96. Cser, L., Frantk, F., Gladkikh, 1. A,, Nezlin, R. S., Novotny, J. &
Pilz, I., Kratky, O., Licht, A. & Sela, M. (1978) Biochemistry, 14,
Pilz, I . , Schwarz, E., Durchschein, W., Licht, A. & Sela, M. (1980)
Pilz, I . , Puchwein, G., Kratky, O., Herbst, M., Hadger, O., Gall,
Pilz, I . (1970) Allg. Prakt. Chem. 21, 21 -29. Cser, L., Gladkikh, I. A., Kozlov, Zh. A,, Nezlin. R . S., Ogieve-
tskaya, M. M. & Ostanevich, Yu. M. (1976) FEBS Lef t . 68, 283- 287.
W. (1976) Nature (Lond.) 264, 418-420.
7, 3958 - 3964.
Lett. 102,249-252.
309 - 314.
Biochemistry, 11, 3321 - 3326.
I. (1978) Proc. Natl Acad. Sci. USA, 72, 2778-2779.
3883.
Lett. 19, 290-292.
1001 - 1010.
Frantk, F. (1979) FEBS Lett. 102, 103-106.
4998 - 8008.
Ostanevich, Yu. M. (1977) FEBS Lett. 80, 329-331.
1326 - 1333.
P ~ o c . Nail A c ~ d . Sci. USA, 77, 117-121.
W. E. & Edelman, G. M. (1970) Biochemistry, 9, 211 -219.
Frangk, F. & Simek, L. (1971) Eur. J. Immunol. I , 300-302. Frantk, F., Doskotil, J. & Simek, L. (1974) Immunochemistry, 11.
Frantk, F. (1978) FEBS Symp. 36, 63-78. Frantk, F., OISovska, Z. & Simek, L. (1979) Eur. J , Immunol. Y,
Novotny, J. & Frantk, F. (1978) Ahstr. 4th Eur. Immunol. Meet., p. 96, Budapest.
Gladkikh, I. A., Kunchenko, A. B., Ostanevich, Yu. M. & Cser, L. (1 978) Joint Institutefor Nuclear Research Reports (Soobshchenya OIYAI) P3-11487, Dubna.
Schedrin, B. M. & Feigin, L. A. (1966) KristallogrGfiya, 11. 183- 163.
Rolbin, Yu. A., Kayoushina, R . L., Feigin, L. A. & Schedrin, B. M. (1973) Kristallogrufya, 18, 701 -705.
Kratky, 0. & Pilz, I. (1978) Q. Rev. Eiophys. I f , 39-70. Cser, L., Frantk, F., Cladkikh, 1. A., Nezlin, R. S., Novotny, J. &
Ostanevich. Yu. M. (1978) FEBS Lett. 93, 312-316. Cser, L., Frantk, F., Gladkikh, I. A., Novotny, J. & Ostanevich,
Yu. M. (1980) Immunol. Letr. I , 188-189. Deisenhofer, J., Colman, P. M., Huber, R., Haupt, H. & Schwick,
G. (1976) Hoppe-Seyler’s Z. Physiol. Chem. 357, 438-448. Valentine, R. C. &Green, N. M. (1967) J . Mol. Biol. 27, 618-617. Pik, I., Schwarz, E. & Palm, W. (1976) Eur. J . Biochem. 71,239 - 247.
803 - 809.
696 - 701.
L. Cser and I. A. Gladkikh, Kozponti Fizikai Kutato Intezet, Magyar Tudominyos Akadtmia, H-1528, Budapest, pf. 49, Hungary
A. B. Kunchenko and Yu. M. Ostanevich, Ob’edinenny institut yadernych idedovaniy, Dubna, 101 000 Moskva, Glavpochtamt p.ya. 79, USSR
F. Frantk, 6stav molekularni genetiky, Ceskoslovenski akddemie vgd, Videfiski ulice 1083, CS-14200 Praha 4, Czechoslovakia